
Abstract
Spinal cord injury (SCI) leads to extensive bone loss and high incidence of low-energy fractures. Pulsed electromagnetic fields (PEMF) treatment, as a non-invasive biophysical technique, has proven to be efficient in promoting osteogenesis. The potential osteoprotective effect and mechanism of PEMF on SCI-related bone deterioration, however, remain unknown. The spinal cord of rats was transected at vertebral level T12 to induce SCI. Thirty rats were assigned to the control, SCI, and SCI+PEMF groups (n = 10). One week after surgery, the SCI+PEMF rats were subjected to PEMF (2.0 mT, 15 Hz, 2 h/day) for eight weeks. Micro-computed tomography results showed that PEMF significantly ameliorated trabecular and cortical bone microarchitecture deterioration induced by SCI. Three-point bending and nanoindentation assays revealed that PEMF significantly improved bone mechanical properties in SCI rats. Serum biomarker and bone histomorphometric analyses demonstrated that PEMF enhanced bone formation, as evidenced by significant increase in serum osteocalcin and P1NP, mineral apposition rate, and osteoblast number on bone surface. The PEMF had no impact, however, on serum bone-resorbing cytokines (TRACP 5b and CTX-1) or osteoclast number on bone surface. The PEMF also attenuated SCI-induced negative changes in osteocyte morphology and osteocyte survival. Moreover, PEMF significantly increased skeletal expression of canonical Wnt ligands (Wnt1 and Wnt10b) and stimulated their downstream p-GSK3β and β-catenin expression in SCI rats. This study demonstrates that PEMF can mitigate the detrimental consequence of SCI on bone quantity/quality, which might be associated with canonical Wnt signaling-mediated bone formation, and reveals that PEMF may be a promising biophysical approach for resisting osteopenia/osteoporosis after SCI in clinics.
Introduction
Spinal cord injury (SCI) is catastrophic damage with neurologic dysfunction below the level of injury, resulting in a high risk of paraplegia and tetraplegia and subsequent increased premature death. 1 A SCI induces deleterious skeletal complication characterized by severe sublesional bone loss, particularly at the distal femur and proximal tibia. 2 More than 50% of persons with SCI have low-energy fractures after injury. 3 The SCI-induced bone loss results from suppressed bone formation and increased bone resorption. 4
Although the etiology is complicated, the lack of mechanical stimuli and insufficient nutrition are believed to contribute greatly to the occurrence of SCI-associated osteoporosis. 5,6 The current available non-pharmacological therapies are based mainly on applying mechanical forces to skeleton (e.g., standing, assisted walking, and physical exercise), whereas the efficacy remains controversial. 7 Moreover, these mechanically based approaches are inconvenient and difficult to implement for the majority of patients with SCI.
Pharmacological therapies such as bisphosphonates and calcitonin have proven to be effective to partially resist SCI-induced bone deterioration by inhibiting bone resorption. 8,9 Nonetheless, some non-negligible issues of bisphosphonates (potential side effects, such as musculoskeletal pain and gastrointestinal irritation) and calcitonin (high cost and side effects, such as nausea, vomiting, diarrhea, abdominal pain, and headache) limit their extensive clinical application in the SCI population. 10,11 Therefore, exploring more countermeasures with effective, safe, and economical nature for SCI-related osteoporosis is needed critically, especially for undeveloped regions/countries.
Pulsed electromagnetic fields (PEMF) stimulation, as a non-invasive biophysical approach, has been employed extensively in the treatment of patients with skeletal diseases since the approval by the Food and Drug Administration in 1979. 7 The satisfactory outcomes with PEMF therapy in the management of fresh fractures, delayed and ununited fractures, and osteoarthritis have been confirmed by several clinical and experimental studies. 12 –14
More recently, growing evidence suggests that PEMF treatment has positive effects on preserving bone mass and strength in rodent osteoporotic models induced by estrogen deficiency, disuse, and diabetes. 15,18 Some in vitro studies have also shown that PEMF has the capability to promote osteoblast differentiation and mineralization. 19 Several clinical trials have also demonstrated that PEMF can significantly enhance bone mass in patients with osteoporosis. 20,21 Despite the successful utilization of PEMF in many bone diseases, the potential osteoprotective effects and mechanism of PEMF on SCI-induced bone loss remain unknown.
The canonical Wnt (Wnt/β-catenin) signaling pathway is known to be crucial in regulating bone development and modulating bone formation. 22,23 Low bone mass phenotype developed in mice deficient for several critical components in the canonical Wnt signaling pathway (e.g., Lrp5/6, β-catenin). 24 –26 It has been reported that SCI-induced decrease of bone mass and bone formation is associated with the suppression of the canonical Wnt signaling pathway. 27
Several studies have also demonstrated that PEM Finduced enhancement of osteoblastogenesis in vitro is associated with the activation of Wnt/b-catenin signaling. 28,29 Previous in vivo studies have shown that the expression levels of critical genes in the Wnt/β-catenin pathway, such as Wnt1, p-GSK-3β, and β-catenin, were upregulated after PEMF stimulation in the normal or aging skeleton. 30 Therefore, it is necessary to study the role of the canonical Wnt pathway in PEMF-mediated regulation of bone metabolism in animals with SCI.
In this study, the effects of PEMF treatment on bone mass, microstructure, and mechanical properties in the SCI rat model were systematically evaluated via micro-computed tomography (CT), three-point bending, and nanoindentation assays. The functional activities of osteoblasts, osteoclasts, and osteocytes and their corresponding bone turnover were also examined by serum biochemical, bone histological, and histomorphometrical analyses. Further, the underlying molecular mechanisms of PEMF on bone metabolism in SCI rats were also investigated. The present study aims to provide proof-of-principle evidence for future scientific and reasonable clinical application of PEMF in SCI-related bone loss.
Methods
Animals and experimental design
Two-month–old male Sprague-Dawley rats were purchased from the Animal Center of the Fourth Military Medical University and acclimated to the environment until three months of age. All animals were housed under controlled temperature (23° ± 1°C) and humidity (50% ∼ 60%). All animal experiments were approved by the Institutional Animal Care and Use Committee of the Fourth Military Medical University and were in accordance with the guidelines and regulations for the care and protection of animals. To avoid the interference of estrogen, 4 only male rats were used in the present study.
Animals were randomized into three groups (10 rats in each group): the control group (Control), SCI group (SCI), and SCI with PEMF exposure group (SCI+PEMF). After anesthetization with intraperitoneal sodium pentobarbital (30 mg/kg), the spinal cord was completely transected at vertebral level T12 to induce a traumatic SCI in the SCI and SCI+PEMF groups. All rats were treated with penicillin G potassium (80,000 U) intraperitoneally to avoid wound infection for three consecutive days after surgery. Bladders were emptied manually at least three times per day until automatic voiding recovered. Rats in the control group received only a laminectomy without damage to the spinal cord.
After surgery, post-injury locomotor behavior was evaluated using the Basso-Beattie-Bresnahan (BBB) rating scale, and we found all rats with SCI had complete paralysis according to the BBB scores. One week after surgery, rats in the SCI+PEMF group were exposed to a 2 h/day PEMF exposure for eight consecutive weeks. The first week post-surgery is a rehabilitation period for rats after SCI, and the animal behavior and status were monitored and anti-infection treatment was performed in this period. Previous studies have also chosen the similar paradigm of initiating pharmacological therapy or biophysical intervention one week after SCI. 31 –33
The exposure duration (2 h/day, 8 weeks) of PEMF has been proven effective in improving bone mass and strength in normal and osteoporotic animals induced by aging or disuse. 18,30 Rats in the SCI and SCI+PEMF groups were observed to exhibit the symptoms of complete paralysis based on the BBB score testing throughout the experiment. Rats after SCI demonstrated symptoms of paraplegia, muscle spasticity and atrophy, and other complications (e.g., inappetence, cacotrophia, bladder dysfunction and infection) were not observed. The fluorochrome calcein (Sigma-Aldrich) was injected intraperitoneally at a dose of 8 mg/kg on 14 and four days before euthanasia, respectively.
Rats were sacrificed by an overdose of sodium pentobarbital at the end of the experiment. Blood samples were obtained from the abdominal aorta immediately for biochemical analysis. After euthanasia, the full transections of the spinal cord were further confirmed by autopsy. The soft tissues and muscle were removed gently, and bilateral femora and tibiae were harvested on the ice. Left femora were immersed in 85% ethanol for micro-CT, nanoindentation, and dynamic bone histomorphometrical analyses. Right femora were wrapped in saline-soaked gauze and stored at -80°C for three-point bending testing. Left proximal tibiae were immersed in 4% paraformaldehyde (PFA) for histological analyses of bone cells. The middiaphysis of left and right tibiae (1.5 cm length) was spun down in a centrifuge to separate out bone marrow and stored in liquid nitrogen for Western blotting and quantitative reverse transcription polymerase chain reaction (qRT-PCR) examinations, respectively.
PEMF stimulation
Rats in the SCI+PEMF group were exposed to whole-body PEMF stimulation generated by a custom-designed electromagnetic exposure device (GHY-III, FMMU, Xi'an, China). The device was made up of a signal generator and three parallel Helmholtz coils. The PEMF waveform used in this experiment consisted of a pulsed burst (burst width, 5 msec; pulse width, 0.2 msec; pulse wait, 0.02 msec; burst wait, 60 msec; pulse rise, 0.3 msec; pulse fall, 2.0 msec) at a frequency of 15 Hz. Our previous studies confirmed that this specific PEMF waveform can promote osteoblastic function and bone growth. 15,17,28
Animals were housed in cages that were aligned with the center of the coils to make sure that rats were in the center of the pulsed electromagnetic fields. A Gaussmeter (Model 455 DMP Gaussmeter; Lake Shore Cryotronics, Westerville, OH) was utilized to measure accurately the intensity of the magnetic fields, and the peak intensity of the magnetic fields was approximately 2.0 mT. Another circular coil (5 cm coil diameter, 1 mm coil diameter, 20 turns) was placed in the midcenter of the Helmholtz coils with the coil parallel to the Helmholtz coils, and the induced peak electrical field was measured to be ∼2 mV/cm.
Micro-CT scanning
The left femora were scanned using a high-solution micro-CT (GE Healthcare, Madison, WI) to obtain the three-dimensional (3D) bone microstructure. The scanner was set using the following parameters: 80-kV voltage, 80-μA current, 2.96-s exposure time, a 210-degree rotation angle, and a 0.4-degree rotation step. The 3D images were reconstructed based on the acquired two-dimensional images with an isotropic voxel size of 16 μm.
Then, the quantitative analysis of trabecular and cortical bone architecture was performed using the VGStudio Max 2.2 software (Volume Graphics, Heidelberg, Germany). For trabecular bone properties, a volume of interest (VOI) was selected starting at 0.4 mm proximal to the metaphyseal growth plate and extended 2.4 mm proximally. The trabecular microstructural indices were calculated, including trabecular bone mineral density (BMD, mg/cm3), bone volume per total volume (BV/TV), connectivity density (Conn.D, 1/mm3), trabecular number (Tb.N, 1/mm), trabecular thickness (Tb.Th, μm), and trabecular separation (Tb.Sp, mm). For the analysis of cortical bone, another VOI at the femoral middiaphysis with 1.6-mm height was chosen to obtain cortical thickness (Ct.Th, μm) and cortical area (Ct.Ar, mm2).
Three-point bending testing
To determine the whole-bone mechanical properties, right femora were destructively tested based on the three-point bending testing using a mechanical testing system (ElectroForce 3220, Bose Corp, Eden Prairie, MN). The samples were thawed in saline for 1 h before testing. The bone sample was placed between the two fixed loading points of the sample supporter (span length, 20 mm) with the physiological curvature facing up. The samples were subjected to a 2-N stabilized, and then a bending load was applied at the midpoint of the femoral diaphysis until reaching its breaking point at a vertical displacement rate of 2 mm/min. The force and displacement data were recorded, and biomechanical parameters were calculated subsequently, including ultimate load (N), yield load (N), stiffness (N/mm), and energy to failure (mJ).
Nanoindentation
After micro-CT scanning, left femoral metaphysis was dehydrated and embedded in methyl methacrylate (MMA). Then, the samples were sectioned longitudinally into approximately 1-mm–thick slices using a Leica 2500E diamond saw microtome (Leica SpA, Milan, Italy). The specimens were polished using progressive grades of silicon carbide abrasive paper (800, 1000, and 1200) under 2-min water irrigation at each grade. Then, samples were polished with a microcloth with 0.25-mm diamond suspension to reduce surface roughness, and subsequently immersed in saline for 24 h to rehydrate. After ultrasonically cleaning, specimens were transferred to an Agilent G200 nanoindentation testing system (Agilent Technologies Inc., Chandler, AZ).
The experiments were conducted according to the previously reported protocol. 28 In brief, first the tip was indented into the bone samples to 4000-nm depth at a strain rate of 0.05 sec−1. Second, the indenter was maintained at the peak load for 10 sec to diminish the potential influence of viscoelasticity and creep. Third, the tip was unloaded to 10% of the peak load at the maximum loading rate for 60 sec to calculate the thermal drift. Finally, the indenter was removed from the bone surface, and the intrinsic material properties of trabecular and cortical bone tissues were analyzed based on the obtained force-displacement data, including elastic modulus (GPa), hardness (GPa) and dissipated energy (mN*nm).
Bone histomorphometry
The left femoral specimens embedded in MMA were sectioned longitudinally to 50 μm using the diamond saw microtome to visualize the calcein fluorochrome labels using a fluorescence microscope (BX51, Olympus, Tokyo, Japan). The region that was 1 mm–3 mm proximal to the distal growth plate of the femoral metaphyseal region was defined as the VOI for dynamic trabecular bone histomorphometrical analysis. To calculate mineral apposition rate (MAR, μm/day), average distance between the double fluorochrome-labeled layers was obtained and divided by the labeling intervals of 10 days. Bone formation rate/bone surface (BFR/BS, μm3/μm2/day) was calculated as the product of MAR and mineralizing surface per BS (double-labeled plus half the single-labeled perimeter as a fraction of the total bone perimeter).
Serum biochemical analysis
Blood samples were centrifuged immediately at 12,000 rpm for 5 min after sample collection, and then the supernatant was stored at -80°C. Osteocalcin (OC) and N-terminal propeptide of type 1 procollagen (P1NP) were measured as bone formation markers using commercial enzyme-linked immunosorbent assay kits (CUSABIO Biotech Co., Wuhan, China). Serum levels of bone resorption markers were also quantified, including tartrate-resistant acid phosphatase 5b (TRACP 5b) and C-terminal cross-linked telopeptides of type 1 collagen (CTX-1). All serum measurements were performed according to the manufacturer's recommended procedures.
Histological analyses of osteocytes, osteoblasts, and osteoclasts
After fixed in 4% PFA for 48 h, left proximal tibiae were cut longitudinally into two parts to perform histological analyses of bone cells. The region that was 1–3 mm distal to the proximal growth plate of the tibial metaphyseal region was defined as the VOI for histological analysis. One part of the sample was embedded in MMA for osteocyte morphological analysis. After polishing with abrasive paper (4000 grits), the sample surface was acid-etched in 37% phosphoric acid for 10 sec, rinsed by dH2O five times for 2 min, immersed in 5% sodium hypochlorite for 10 min, and rinsed by dH2O five times for 2 min. After air drying overnight, the surface was coated with a layer of gold palladium and imaged under a scanning electron microscope (SEM, S-3400N, HITACHI, Japan). Another part of the tibial sample was decalcified in 10% ethylenediaminetetraacetic acid, embedded in paraffin, and then sectioned to 5-μm thickness.
TUNEL fluorescence staining was performed for osteocyte survival analysis using a TUNEL apoptosis detection kit (Roche, Mannheim, FRG, Germany). Nuclei were counterstained with DAPI (4', 6-diamidino-2-phenylindole). The percentage of apoptotic osteocytes was calculated by dividing the number of positively stained osteocytes by DAPI-labeled osteocytes. Toluidine blue staining and tartrate-resistant acid phosphatase staining were performed to visualize osteoblasts and osteoclasts, respectively. Osteoblast number per millimeter of bone surface (N.OB/BS, 1/mm) and osteoclast number per millimeter of bone surface (N.OC/BS, 1/mm) were then quantified.
Real-time PCR
The right tibiae were ground to powder in an RNase-free mortar filled with liquid nitrogen. Then, total ribonucleic acid (RNA) was extracted using RNAiso Plus (Takara, Ohtsu, Japan) following the manufacturer's guidelines and quantified using the Nanodrop 2000 spectrophotometer (Thermo Scientific, Waltham, MA). Complementary deoxyribonucleic acid (cDNA) was synthesized from 2-μg total RNA using the PrimeScriptTM RT Master Mix (Takara).
Real-time PCR was performed in a reaction volume of 20 μL using SYBR Premix Ex TaqTM II (Takara) in the CFX96 Real-Time PCR Detection System (Bio-Rad, Hercules, CA) under the following cycling conditions: an initial denaturation at 95°C for 30 sec plus 40 cycles consisting of a 15-sec denaturation interval at 95°C, a 15-sec annealing interval at 55°C, and a 15-sec extension interval at 55°C. The glyceraldehyde-3-phosphate dehydrogenase (GAPDH) messenger RNA was used to normalize the gene expression. The following primers synthesized by AuGCT DNA-SYN Biotechnology Co., Ltd. (Beijing, China) were used, including: Wnt1 (forward primer, 5'-CGAGGTGAAAGGGCAAGGAAAG-3'; reverse primer, 5'-TGATGAAGAGGGAGCAGGACAG-3'), Wnt3a (forward primer, 5'-TGAATTTGGAGGAATGGTCTCT-3'; reverse primer, 5'-TGGGCACCTTGAAGTATGTGTA-3'), Wnt10b (forward primer, 5'-CGAGAATGCGGATCCACAA-3'; reverse primer, 5'-CCGCTTCAGGTTTTCCGTTA-3') and GAPDH (forward primer, 5'-ACCACAGTCCATGCCATCAC-3'; reverse primer 5'-TCCACCACCCTGTTGCTGTA-3'). Statistical analysis was performed using the 2-ΔΔCt method.
Western blotting
Left tibial diaphyses were ground to powder in liquid nitrogen, and then homogenized in radioimmunoprecipitation assay buffer with 1 mM phenylmethylsulfonyl fluoride. Protein extracts were diluted 4:1 with 5 × loading buffer. Then, 20 μg protein per sample were electrophoresed on 10% Tris-glycine sodium dodecyl sulfate-polyacrylamide gel and transferred to the NC membrane (Pierce, Shanghai, China). The membranes were blocked in 5% milk in Tris-buffered saline with 0.5% Tween-20 for 1 h, and incubated overnight at 4°C with primary antibodies p-GSK3β (Abcam, Cambridge, MA), GSK3β (Abcam), β-catenin (Abcam), and β-actin (Bioworld technology, Louis Park, MN), respectively.
Then the membranes were incubated with secondary antibody for 2 h. The SuperSignal West Pico chemiluminescent substrate kit (Thermo Scientific, Rockford, IL) was used to visualize the blots according to the manufacturer's instructions and imaged using an ECL chemiluminescent system (GE ImageQuant 350, GE Healthcare).
Statistical analysis
All data were reported as mean ± standard deviation, and statistical differences were defined significant with a p value <0.05. Data were analyzed using SPSS 19.0 software (SPSS, Chicago, IL). Normal distribution was examined using the Kolmogorov-Smirnov test, and homoscedasticity was evaluated using the Levene test. Each parameter was found to obey normal distribution and homoscedasticity. Comparisons between the three groups were performed using one-way analysis of variance followed by Bonferroni post hoc analysis.
Results
Effects of PEMF stimulation on serum markers of bone turnover in rats with SCI
The SCI group showed significantly lower serum levels of osteocalcin and P1NP than the control group (Fig. 1A,B, p < 0.001, -46.1% and -36.9%, respectively). In addition, rats with SCI exhibited enhanced bone resorption when compared with the control rats, as evidenced by significant increase in the levels of serum TRACP 5b (Fig. 1C, p < 0.001, +38.3%) and CTX-1 (Fig. 1D, p < 0.01, +44.2%). The SCI+PEMF group showed significant increase in the levels of serum osteocalcin and P1NP compared with the SCI group (Fig. 1A,B, p < 0.001, +64.0% and +46.0%, respectively), whereas no significant difference was observed in the serum levels of TRACP 5b and CTX-1 between the SCI group and SCI+PEMF group (Fig. 1C,D, p > 0.05, -7.2% and -6.3%, respectively).
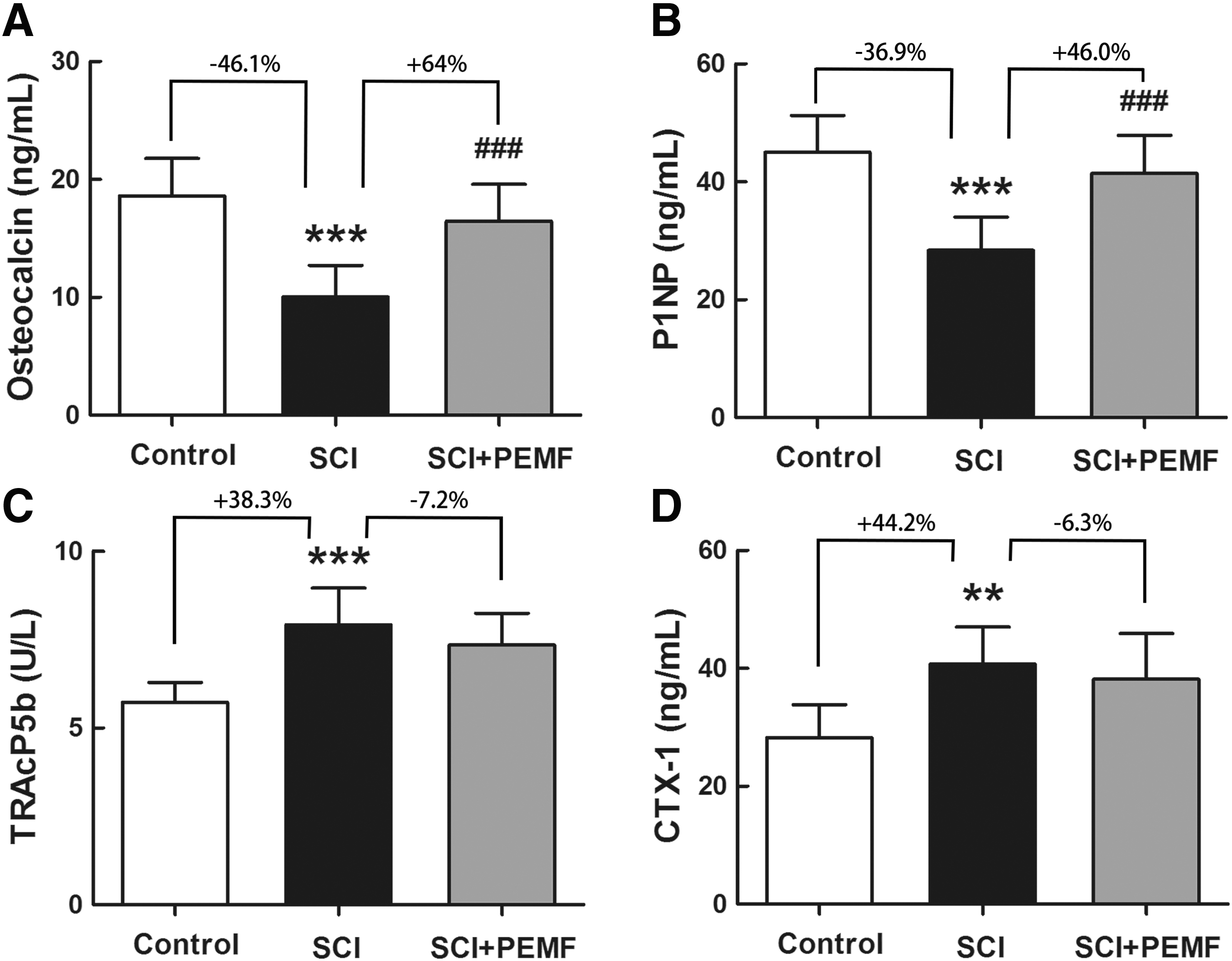
Effects of eight-week pulsed electromagnetic fields (PEMF) exposure on serum biochemical levels in rats with spinal cord injury (SCI), including bone formation indices (
Effects of PEMF on trabecular and cortical bone microstructure in rats with SCI
The SCI group exhibited reduction in trabecular bone mass and deterioration in trabecular microarchitecture compared with the control group (Fig. 2A–G), as evidenced by a significant decrease in BMD (p < 0.001, -22.9%), BV/TV (p < 0.001, -50.1%), Conn.D (p < 0.01, -32.4%), Tb.N (p < 0.001, -29.7%), and
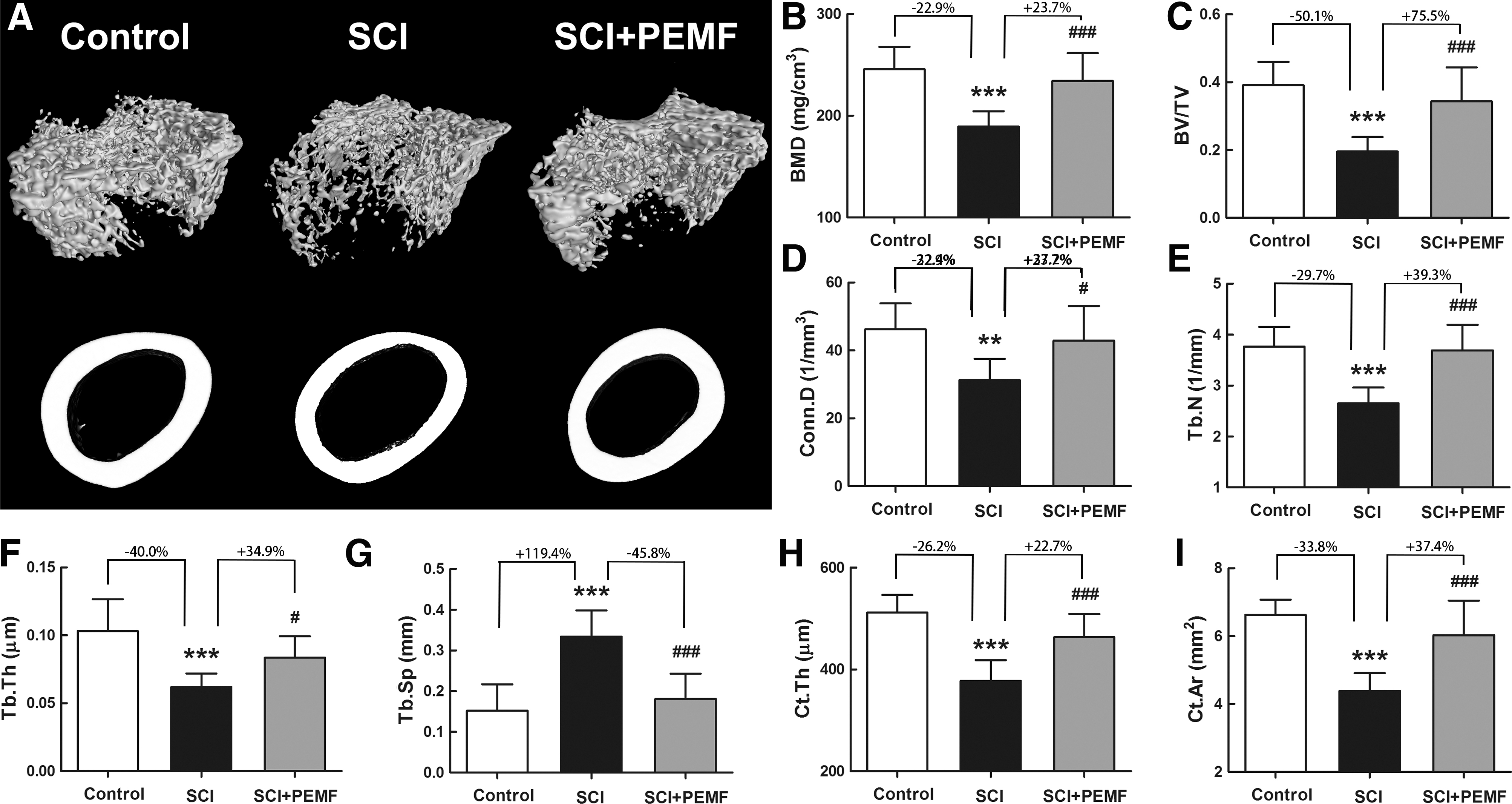
Effects of eight-week pulsed electromagnetic fields (PEMF) exposure on cancellous and cortical bone mass and microstructure in rats with spinal cord injury (SCI) as assessed by micro-computed tomography.
The PEMF exposure partially prevented trabecular bone loss in the SCI+PEMF group at the distal femora, which was characterized by increased BMD (p < 0.001, +23.7%), BV/TV (p < 0.001, +75.5%), Conn.D (p < 0.05, +37.2%), Tb.N (p < 0.001, +39.3%), and
Effects of PEMF stimulation on bone mechanical properties in rats with SCI
According to the three-point bending testing (Fig. 3), we found that the SCI group had lower ultimate load (p < 0.001, -17.1%), yield load (p < 0.01, -17.3%), stiffness (p < 0.001, -21.2%), and energy to failure (p < 0.001, -39.8%) than the control group. In comparison with the SCI group, the SCI+PEMF group showed significant increase in ultimate load (p < 0.05, +13.9%), yield load (p < 0.05, +15.0%), stiffness (p < 0.001, +22.2%), and energy to failure (p < 0.01, +35.4%), suggesting that PEMF inhibited SCI-induced deterioration of the whole-bone structural properties.
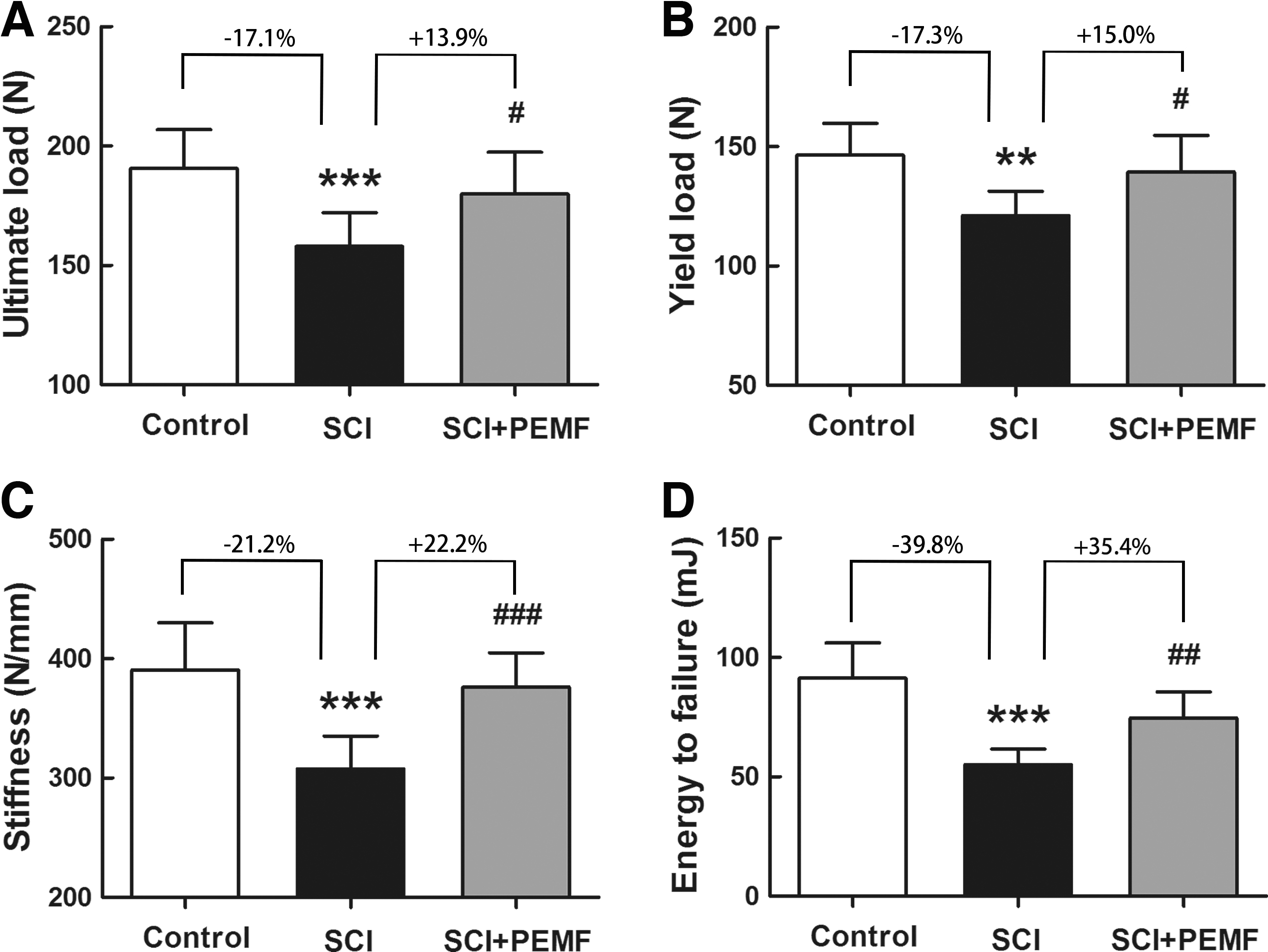
Effects of eight-week pulsed electromagnetic fields (PEMF) exposure on femoral whole-bone mechanical properties in rats with spinal cord injury (SCI) as assessed by three-point bending testing, including
The nanoindentation results demonstrated that rats with SCI experienced compromised intrinsic material properties in both trabecular bone and cortical bone (Fig. 4), as evidenced by the significant reduction in modulus (p < 0.001, -15.2% and -18.3%, respectively), hardness (p < 0.01, -23.2% and -25.0%, respectively), and dissipated energy (p < 0.001, -13.4% and -12.2%, respectively). In comparison with the SCI group, the SCI+PEMF group showed a significant increase in modulus (p < 0.01, +14.1% and +17.9%, respectively), hardness (p < 0.05, +23.6% and +25.4%, respectively), and dissipated energy (p < 0.01, +11.1% and +10.8%, respectively) in trabecular and cortical bone.

Effects of eight-week pulsed electromagnetic fields (PEMF) exposure on intrinsic material properties in both trabecular bone and cortical bone in rats with spinal cord injury (SCI) assessed by biomechanical nanoindentation testing, including
Effects of PEMF stimulation on bone histomorphometry in rats with SCI
Our dynamic histomorphometrical results (Fig. 5A,C,D) revealed that rats with SCI had greatly decreased bone formation rate when compared with the control group, as evidenced by significantly reduced MAR (p < 0.001, -31.4%) and BFR/BS (p < 0.001, -38.4%). The PEMF induced significant increase in MAR (p < 0.001, +42.0%) and BFR/BS (Pp< 0.001, +50.7%) in rats with SCI. Moreover, we also found that rats with SCI exhibited significantly lower N.OB/BS than the control rats (Fig. 5E, p < 0.001, -39.6%).
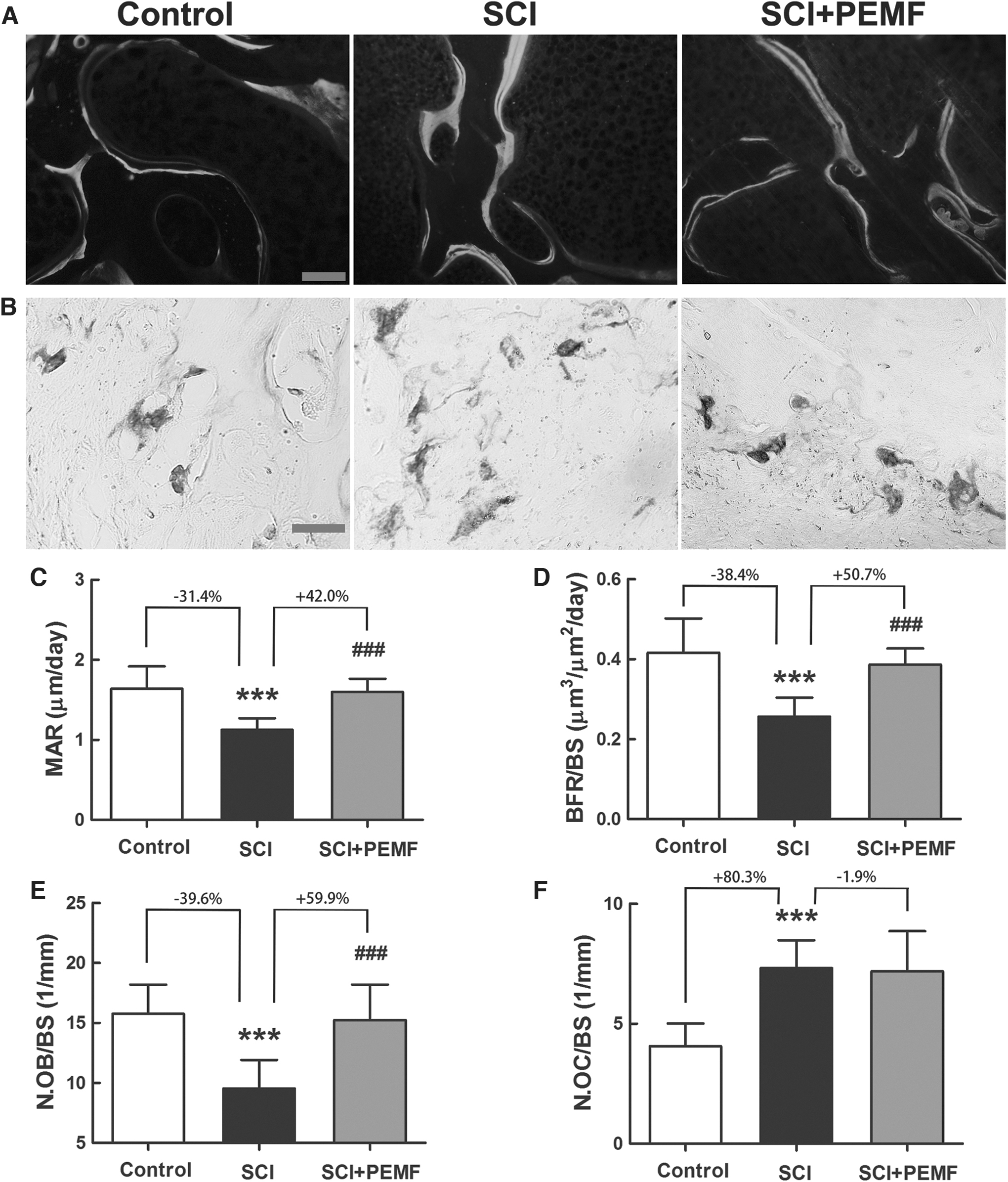
Effects of eight-week pulsed electromagnetic fields (PEMF) on bone histomorphometry in rats with spinal cord injury (SCI).
The TRAP staining results revealed that the SCI group demonstrated a significant increase in N.OC/BS when compared with the control group (Fig. 5B,F, p < 0.001, +80.3%). The PEMF partially reversed the reduction in N.OB/BS in rats with SCI (p < 0.001, +59.9%). No significant difference was detected, however, in N.OC/BS between the SCI group and the SCI+PEMF group.
Effects of PEMF on osteocyte morphology and survival in rats with SCI
The SEM results (Fig. 6A) showed that osteocytes in rats with SCI exhibited reduced dendrite number and impaired dendrite connectivity compared with those in the control rats. The TUNEL staining results also demonstrated that rats with SCI had much more TUNEL-positive osteocytes than the control rats (Fig. 6B). The PEMF exposure effectively inhibited the deterioration of osteocyte morphology and significantly increased the osteocyte dendrite number in rats with SCI (Fig. 6A,C, p < 0.01, +64.3%). Moreover, PEMF also significantly reduced osteocyte apoptosis in rats with SCI, as evidenced by decreased TUNEL+ osteocyte number in the SCI+PEMF group compared with the SCI group (Fig. 6D, p < 0.001, -55.9%).
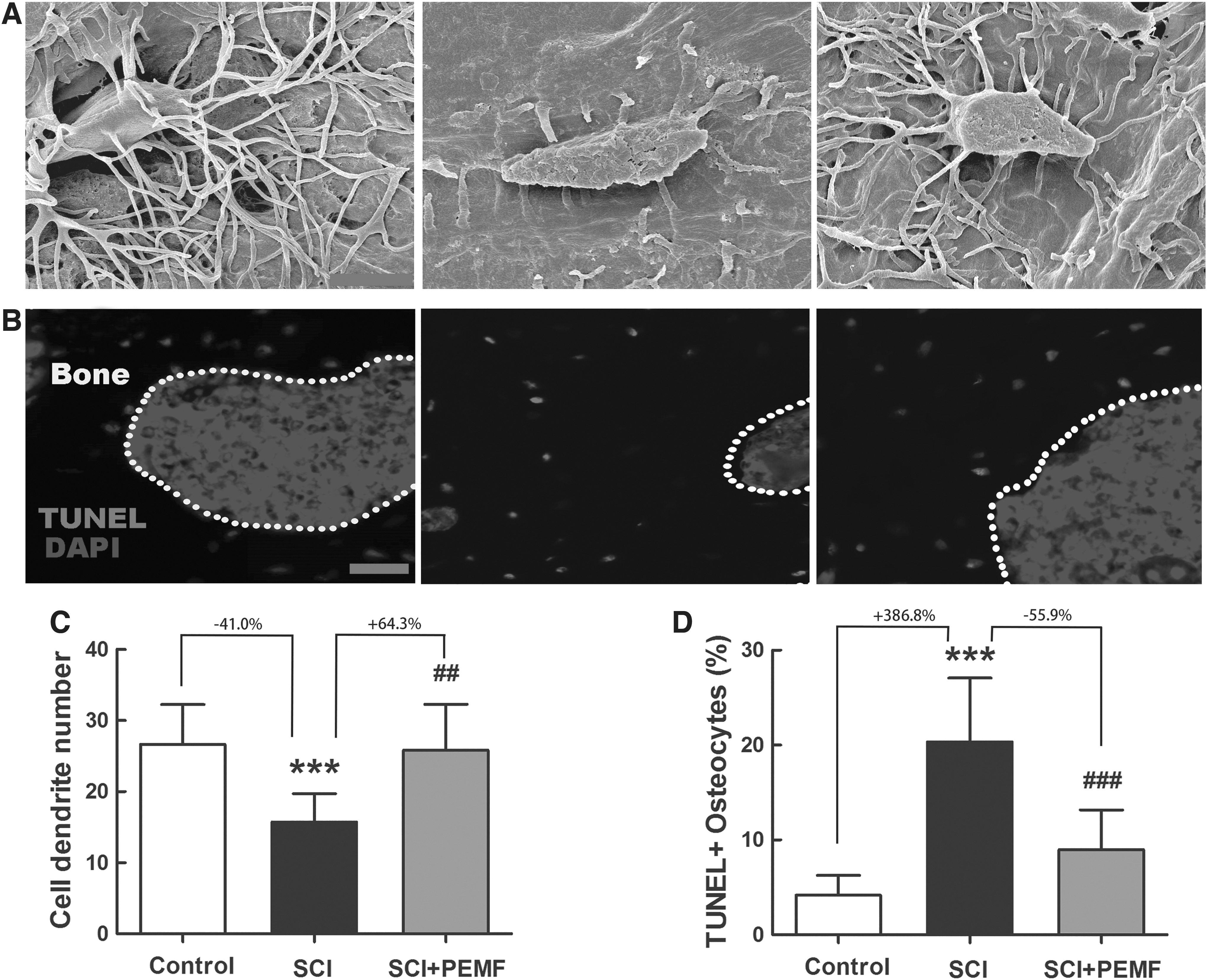
Effects of eight-week pulsed electromagnetic fields (PEMF) exposure on osteocyte morphology and survival in rats with spinal cord injury (SCI).
Effects of PEMF on skeletal Wnt/β-catenin signaling expression in rats with SCI
The real-time PCR results suggested that the gene expression levels of Wnt1 (Fig. 7A, p < 0.001, -60.2%), Wnt3a (Fig. 7B, p < 0.01, -37.7%), and Wnt10b (Fig. 7C, p < 0.001, -60.2%) in the tibiae of rats with SCI were significantly lower than those of the rats in the control group. As shown in Fig. 7D,E, SCI significantly reduced the protein level of p-GSK-3β/GSK-3β (p < 0.001, -62.4%) in the SCI group compared with the control group. Moreover, rats with SCI exhibited a significantly lower β-catenin protein level than the control rats (Fig. 7F, p < 0.001, -67.7%).
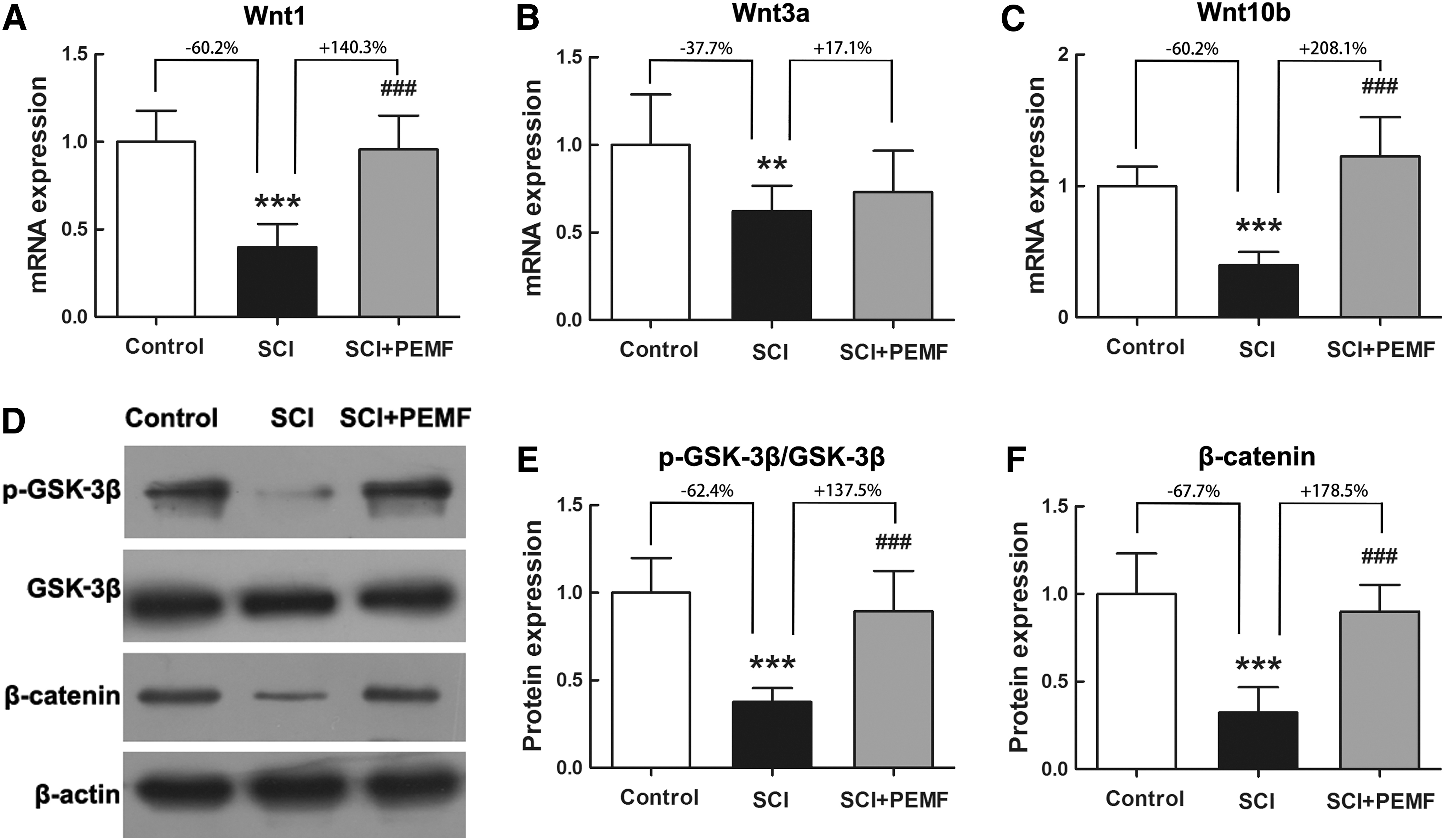
Effects of eight-week pulsed electromagnetic fields (PEMF) exposure on tibial gene and protein expression in canonical Wnt signaling in rats with spinal cord injury (SCI) via real-time polymerase chain reaction and Western blotting analyses, respectively. (
The PEMF significantly increased the mRNA levels of Wnt1 (p < 0.001, +140.3%) and Wnt10b (p < 0.001, +208.1%) in rats with SCI, whereas no significant difference was observed in the Wnt3a expression between the SCI group and the SCI+PEMF group (P > 0.05, +17.1%). In comparison with the SCI group, the protein expression levels of p-GSK-3β/GSK-3β and β-catenin were also significantly increased in the SCI+PEMF group (P < 0.001, +137.5% and +178.5%, respectively).
Discussion
Individuals with SCI experience profound changes in the skeleton, characterized by rapid bone loss and remarkably increased fracture risk. Considering the side effects, high cost, or insufficient effectiveness of most pharmacological interventions, non-pharmacological paradigms based on skeletal mechanical stimulation have received extensive research interests. 34 Patients with SCI, however, have all kinds of psychological and physiological barriers to perform exercise. 35
Substantial studies demonstrate that PEMF has the potential to be a promising therapeutic method to promote fresh/delayed fracture healing with no observable side effects. 16,36 In recent years, growing evidence has shown that PEMF has the capacity to inhibit bone loss induced by disuse or estrogen deficiency. Nevertheless, it remains unknown whether PEMF can effectively attenuate the adverse outcomes of SCI-induced bone abnormalities.
In this study, we found that PEMF preserved cancellous and cortical bone mass, microarchitecture, and mechanical strength by rescuing osteoblast- and osteocyte-mediated bone anabolism in rats with SCI, which was associated with PEMF-induced activation of the skeletal Wnt/β-catenin signaling pathway. The current study offers an exciting new avenue to resist bone deterioration in patients with SCI.
In this study, the complete transection model was used to induce SCI. The complete transection model results in complete SCI, whereas the degree of injury severity of contusion models may not be that easy to be controlled because of the individual differences of animals compared with the transection model. 37,38 Moreover, the difficulty in confidently and precisely identifying the anatomical location of the lesion and variability resulting from different types of impactors used for contusion may also potentially influence the repeatability and reproducibility of the experimental results. 39,40
Therefore, using the complete transection model may reduce the potential influence resulting from different degrees of injury severity and may be helpful for verifying the effects of PEMF on bone loss after SCI more clearly. Although contusion is more common clinically, the complete transection model can also mimic complete laceration of the spinal cord induced by stab wounds or gunshot injuries. 41,42 In addition, the complete transection animal model at the lower thoracic cord has been employed extensively to investigate the pathophysiology of SCI-induced bone loss. 32,33,43 –46
It is widely acknowledged that individuals with SCI exhibit significant reduction in trabecular bone mass, especially at the distal femur and proximal tibia. 47 Our micro-CT results showed that complete SCI for nine weeks resulted in detrimental alterations in trabecular bone mass and bone microarchitecture, including compromised trabecular number, thickness, and connectivity. Similarly, previous studies have also observed severe cancellous bone deterioration in animals after SCI. 48
Interestingly, PEMF exposure was found to almost restore the trabecular number, separation, and volume fraction in rats with SCI to the levels in the control rats. Moreover, the deterioration of trabecular bone connectivity and thickness induced by SCI were also partially inhibited by PEMF. After PEMF exposure, trabecular bone became much denser with more “plate-like” trabeculae and highly interconnected morphology, which is beneficial for bone to resist higher loads. 49 Moreover, we also found that SCI induced cortical bone thinning in rats, which was consistent with previous reports. 45 The PEMF stimulation partially mitigated the deterioration in cortical thickness in rats with SCI, revealing potential improvement of bone mechanical integrity and fracture toughness. 50
To evaluate the fracture resistance capacity of rats with SCI, three-point bending testing at whole-bone level was performed. We found that SCI led to a remarkable decrease in femoral structural properties. Several previous studies have also found similar biomechanical alterations in the skeleton of animals with SCI. 51,52 We also found that PEMF partially inhibited SCI-induced deterioration in whole-bone mechanical properties, revealing improved ability of the skeleton to resist fracture. Further, nanoindentation was performed to investigate tissue-level material properties independent of the sample size and shape. 53 We found that elastic modulus and hardness of both trabecular and cortical bone were compromised in rats with SCI.
To our knowledge, this is the first study reporting SCI-associated intrinsic mechanical properties of the skeleton. Interestingly, PEMF exposure was found to partially ameliorate SCI-induced deterioration in trabecular and cortical bone material properties. Because skeletal intrinsic elastic modulus and hardness determined by nanoindentation depend highly on the degree of mineralization, 54 this study reveals that PEMF may promote the deposition of bone-related irons (e.g., calcium, phosphate, and magnesium) into bone matrix and subsequently result in tissue-level changes in skeletal material properties.
Bone loss after SCI is known to be associated with unbalanced bone metabolism with decreased bone formation and accelerated bone resorption. 48 Serum osteocalcin and P1NP, as two sensitive and reliable biochemical markers of bone formation, were herein found to be significantly lower in rats with SCI. We also observed that serum TRACP 5b and CTX-1 (two specific bone resorption biomarkers) were increased in rats with SCI. The PEMF almost restored serum osteocalcin and P1NP levels in rats with SCI to control levels, supporting the strong anabolic action induced by PEMF. Nevertheless, PEMF exerted no observable impact on TRACP 5b or CTX-1 secretion in rats with SCI.
Consistently, the calcein double-labeling results provided more direct evidence that PEMF preserved bone formation in rats with SCI. Further, the bone histological results showed that PEMF exposure increased the osteoblast number on the bone surface in addition to enhancing bone formation rate in rats with SCI. The PEMF, however, had no inhibitory effect on osteoclast number, which was in accordance with the serum biomarker findings. Therefore, the inhibitory effect of PEMF on SCI-induced bone loss may be associated mainly with the promotion of bone formation rather than the suppression of bone resorption.
Accumulating evidence supports the critical role of osteocytes embedded in bone matrix in orchestrating bone metabolic activity by acting as direct detectors to external mechanical signals. 56 Osteocytes can sense mechanical stimulation and secrete several important cytokines to regulate the osteoblast activity via their long dendritic process. 56 Recent study has also demonstrated that osteocytes can also sense exogenous electromagnetic signals via primary cilia and transduce them into intracellular biochemical signals. 57 Specifically, several previous in vivo studies have also found that SCI is detrimental to the function and survival of osteocytes. 33,58
Our SEM and TUNEL staining findings revealed that SCI impaired the morphology and connectivity of the osteocyte network and also induced more osteocyte apoptosis. Interestingly, PEMF treatment inhibited SCI-induced morphological changes of osteocytes and also reduced the number of apoptotic osteocytes. Thus, our findings demonstrate that the osteogenic effect of PEMF in rats with SCI was also related to the improvement of osteocyte activity and survival.
Considerable evidence supports the crucial role of the canonical Wnt signaling pathway (β-catenin-dependent) in regulating bone mass and architecture and maintaining bone homeostasis. 22,23 Activation of canonical Wnt signaling enhances osteoblastogenesis and bone formation by stimulating proliferation and differentiation of osteoblasts. 59 Moreover, the canonical Wnt signaling regulates the expression of osteoblast-specific transcription factors (e.g., Runx2 and osterix). 60,61 In the canonical Wnt pathway, Wnts bind to specific frizzled receptors and LRP5/6 in a ternary complex, and thus inactivate glycogen synthase kinase 3β (GSK-3β) to induce the subsequent nuclear accumulation of β-catenin. 62
It has been reported that diminished bone mass and skeletal abnormalities after SCI are linked with the inhibition of the canonical Wnt signaling. 27 Consistent with these findings, our results also found that SCI not only downregulated the expression of canonical Wnt ligands (Wnt1, Wnt3a, and Wnt10b), but also decreased p-GSK-3β and downstream β-catenin expression. The inhibition of these important molecules of the canonical Wnt signaling induced by SCI was reversed by eight-week PEMF exposure. Our findings indicate that the anabolic response of the SCI skeleton to PEMF was at least partially associated with the activation of the canonical Wnt signaling pathway.
Conclusion
The present study clearly demonstrates that whole-body PEMF exposure significantly alleviated SCI-induced deterioration of trabecular and cortical bone mass and microarchitecture. The PEMF stimulation also had the capacity to improve bone structural and material mechanical properties in rats with SCI. The anti-osteoporotic effect of PEMF treatment resulted mainly from its strong anabolic action in rats with SCI, which was associated with the improvement of osteoblast and osteocyte activities. Further, the preservation of bone formation in SCI was associated with the activation of skeletal canonical Wnt signaling induced by PEMF. This study provides direct evidence that PEMF stimulation, as an easy and non-invasive biophysical technique, has promising therapeutic potential for SCI-induced osteoporosis in clinics.
Footnotes
Authors' Contributions
All authors contributed to the manuscript. JC, DJ, and EPL designed the research. XS, ZDY, DW, YQY, and YJD performed the experiments. XS, JC, ZDY, DW, and EPL analyzed data. XS, JC, DJ, and ZDY wrote the article. All authors have read and approved the manuscript. XS, ZDY, and DW are co-first authors.
Funding Information
The authors acknowledge the support from The National Natural Science Foundation of China (No. 51907111, 11972366, 31500760, and 51777211), Shaanxi Youth Science and technology new star project (No. 2019KJXX-087), and the Shaanxi Provincial Natural Science Foundation of China (No. 2018JQ8056 and 2020SF-084).
Author Disclosure Statement
No competing financial interests exist.