
Abstract
Apathy is a common and impairing sequela of traumatic brain injury (TBI). Yet, little is known about the neural mechanisms determining in which patients apathy does or does not develop post-TBI. We aimed to elucidate the impact of TBI on motivational neural circuits and how this shapes apathy over the course of TBI recovery. Resting-state functional magnetic resonance imaging data were collected in patients with subacute mild TBI (n = 44), chronic mild-to-moderate TBI (n = 26), and nonbrain-injured control participants (CTRL; n = 28). We measured ventromedial prefrontal cortex (vmPFC) functional connectivity (FC) as a function of apathy, using an a priori vmPFC seed adopted from a motivated decision-making study in an independent TBI study cohort. Patients reported apathy using a well-validated tool for assaying apathy in TBI. The vmPFC-to-wholebrain FC was contrasted between groups, and we fit regression models with apathy predicting vmPFC FC. Subacute and chronic TBI caused increased apathy relative to CTRL, replicating previous work suggesting that apathy has an enduring impact in TBI. The vmPFC was functionally connected to the canonical default network, and this architecture did not differ between subacute TBI, chronic TBI, and CTRL groups. Critically, in TBI, increased apathy scores predicted decreased vmPFC-dorsal anterior cingulate cortex (dACC) FC. Last, we subdivided the TBI group based on patients above versus below the threshold for “clinically significant apathy,” finding that TBI patients with clinically significant apathy demonstrated comparable vmPFC-dACC FC to CTRLs, whereas TBI patients with subthreshold apathy scores demonstrated vmPFC-dACC hyperconnectivity relative to both CTRLs and patients with clinically significant apathy. Post-TBI vmPFC-dACC hyperconnectivity may represent an adaptive compensatory response, helping to maintain motivation and enabling resilience to the development of apathy after neurotrauma. Given the role of vmPFC-dACC circuits in value-based decision making, rehabilitation strategies designed to improve this ability may help to reduce apathy and improve functional outcomes in TBI.
Introduction
The motivation to initiate goal-directed behaviors is central to adaptive functioning. Unfortunately, a marked reduction in this motivation (i.e., apathy) is a common sequela of traumatic brain injury (TBI). 1 Apathy negatively impacts quality of life in patients with TBI: it has been linked to poor treatment outcomes, caregiver distress, and loss of job opportunities. 2 Notably, apathy is highly comorbid but independent from depression, another common psychiatric symptom of TBI. 3 –5 Further, previous studies have found no relationship between time since injury and apathy, suggesting that day-to-day motivation can still be decreased in patients with chronic TBI after recovery has plateaued. 6
Despite the deleterious and lasting clinical impact of apathy in TBI, there have been relatively few attempts to elucidate its underlying mechanisms. A better understanding of the neural mechanisms shaping which patients with TBI do or do not experience apathy post-injury is essential, because such findings could inform the design of effective rehabilitation strategies for normalizing motivated behavior in the wake of a TBI event.
One potential route to apathy in TBI is a diminished capacity to determine whether a goal is valuable enough to pursue. Functional magnetic resonance imaging (fMRI) evidence in humans suggests that the reward value computations that drive goal-directed behavior are encoded in the ventromedial prefrontal cortex (vmPFC). 7 A recent voxel-based lesion-symptom mapping study provides causal evidence that this vmPFC reward value signal is linked to the motivation to perform goal-directed behaviors. Specifically, impaired performance on a value-based decision-making task mediated the association between vmPFC damage and increased apathy in patients with focal brain lesions. 8 No brain region is an island, however, and adaptive value-based decision making likely relies on the integrated functioning of distributed networks that extend beyond vmPFC.
No previous studies have examined how post-injury changes in vmPFC connectivity to the distributed networks underlying value-based decision making—in particular, dorsal anterior cingulate cortex (dACC), 9,10 lateral prefrontal cortex (LPFC), 11,12 and subcortical nuclei including amygdala and ventral striatum 13 –16 —might shape the presence or absence of clinically significant apathy in patients with TBI.
Previous studies have yielded conflicting evidence regarding whether vmPFC's functional connectivity increases or decreases after a TBI. 17 –19 This has led to differences in interpretation, with some suggesting that compromised vmPFC circuit integrity disrupts post-TBI functioning, 20 while others suggest that increased functional connectivity of vmPFC and other densely connected network hubs represents an adaptive neuroplastic response for promoting resilience during TBI recovery. 21
We used resting-state fMRI (rsfMRI) and measured vmPFC functional connectivity (FC) in a well-characterized sample of subacute TBI (≤ 2 weeks post-injury), chronic 1 TBI (≥ 1 year post-injury), and nonbrain-injured control participants (CTRLs; for another report on this sample, see 22 ). We used an a priori vmPFC seed that was causally linked to impaired reward valuation and apathy in an independent cohort of brain-injured patients 8 and computed vmPFC-wholebrain FC maps for each participant. Within the wholebrain FC map, we hypothesized that vmPFC FC to the extended value-based decision-making network (namely: dACC, LPFC, amygdala, and ventral striatum) would be associated with the degree of apathy in patients with TBI.
Given mixed findings on vmPFC FC post-TBI in previous studies 19,20 and the fact that roughly 40-50% of patients with TBI do not experience clinically significant apathy, 1,23 there were two viable outcomes we expected to observe. First, vmPFC-value network hypoconnectivity in patients with clinically significant apathy relative to CTRLs would provide evidence of a neural circuit disruption underlying apathy post-TBI.cf.,24 Alternatively, vmPFC-value network hyperconnectivity in patients with apathy scores below clinical threshold relative to CTRLs would suggest an adaptive compensatory response in the valuation network that might help to reduce apathy after neurotrauma.cf.,21
In addition, there were two unlikely possibilities: that apathy below clinical threshold would be associated with decreased vmPFC connectivity, or that clinical apathy would be associated with vmPFC hyperconnectivity. The series of analyses reported here were designed to test the validity of one or more of these vmPFC circuit hypotheses regarding apathy in TBI.
Methods
Study participants
Data were collected from 125 individuals (n = 60 patients with subacute TBI, n = 32 patients with chronic TBI, and n = 33 CTRLs). All participants were 18–55 years old and reported English as their primary language. The subacute TBI sample patients were recruited through the University of New Mexico Health Sciences Center (UNM-HSC)'s Department of Emergency Medicine after having arrived at the hospital because of a self-reported loss of consciousness (LOC) event of ≤30 min, and had a Glasgow Coma Scale score between 13 and 15. These inclusion criteria exceed the minimum requirement for diagnosing mTBI as outlined by most published recommendations. 25 Subacute patients were excluded from participating if they had a history of seizures or previous head injury.
Patients with chronic TBI were also referred by UNM-HSC's Emergency Department or from other Emergency Departments and clinics, from patient recruitment services, or through advertisements in the community. The chronic TBI sample comprised both mild and moderate injuries, and therefore eligibility allowed for LOC up to 24 h, required for head injury to have occurred 3 months to 15 years before testing, and participant must have reported at least one current cognitive symptom on the Neurobehavioral Symptom Inventory. 26 The majority of the chronic TBI group (20/26 of the final chronic TBI sample) had a single mild TBI, whereas a small subset experienced either multiple injuries or a moderate TBI. This variation in TBI history and severity was also taken into consideration (see section Control models).
A group of nonbrain-injured control participants was recruited from the community, and participants were required to self-report 0 traumatic events with LOC during a phone screen to be eligible for the study.
Participants in all three groups did not have a current or recent history of substance abuse, did not have a history of other neurological or major psychiatric disorder, and did not have any contraindications to MRI. Twenty-seven participants were excluded because of excessive head motion during MRI (n = 3 CTRL, n = 2 subacute TBI, and n = 2 chronic TBI; see section Image pre-processing), incomplete MRI or Frontal Systems Behavior Scale (FrSBe) data (n = 2 CTRL, n = 11 subacute TBI, and n = 5 chronic TBI), or not meeting LOC inclusion criteria (n = 2 subacute TBI). This left a final sample of N = 98 participants (n = 44 patients with subacute TBI, n = 26 patients with chronic TBI, and n = 28 CTRL participants; Table 1).
Participant Demographic Information
TBI, traumatic brain injury; Ed, education; Inj, injury.
In the final sample, age and sex were matched between groups (p ≥ 0.339), but there was a significant group difference in years of education (p = 0.006). Therefore, years of education was considered as a potential covariate (see section Control Models).
Neuropsychological assessment
Apathy
The FrSBe was used to obtain self-reported apathy symptoms for all participants. 27 Featured in the FrSBe are 46 total items with the apathy subscale comprising 14 items (e.g., “sit around doing nothing,” “get involved with activities spontaneously,” and “do things without being requested to do so”). Items are scored on a 5-point Likert scale (almost never, seldom, sometimes, frequently, and almost always). The raw apathy scores were the sum of 10 normally scored items and four reverse-scored items. The raw scores were converted to T-scores according to the FrSBe Professional Manual. 27 Clinically significant apathy on the FrSBe is defined as a T-score of ≥65 (grey line in Fig. 1).
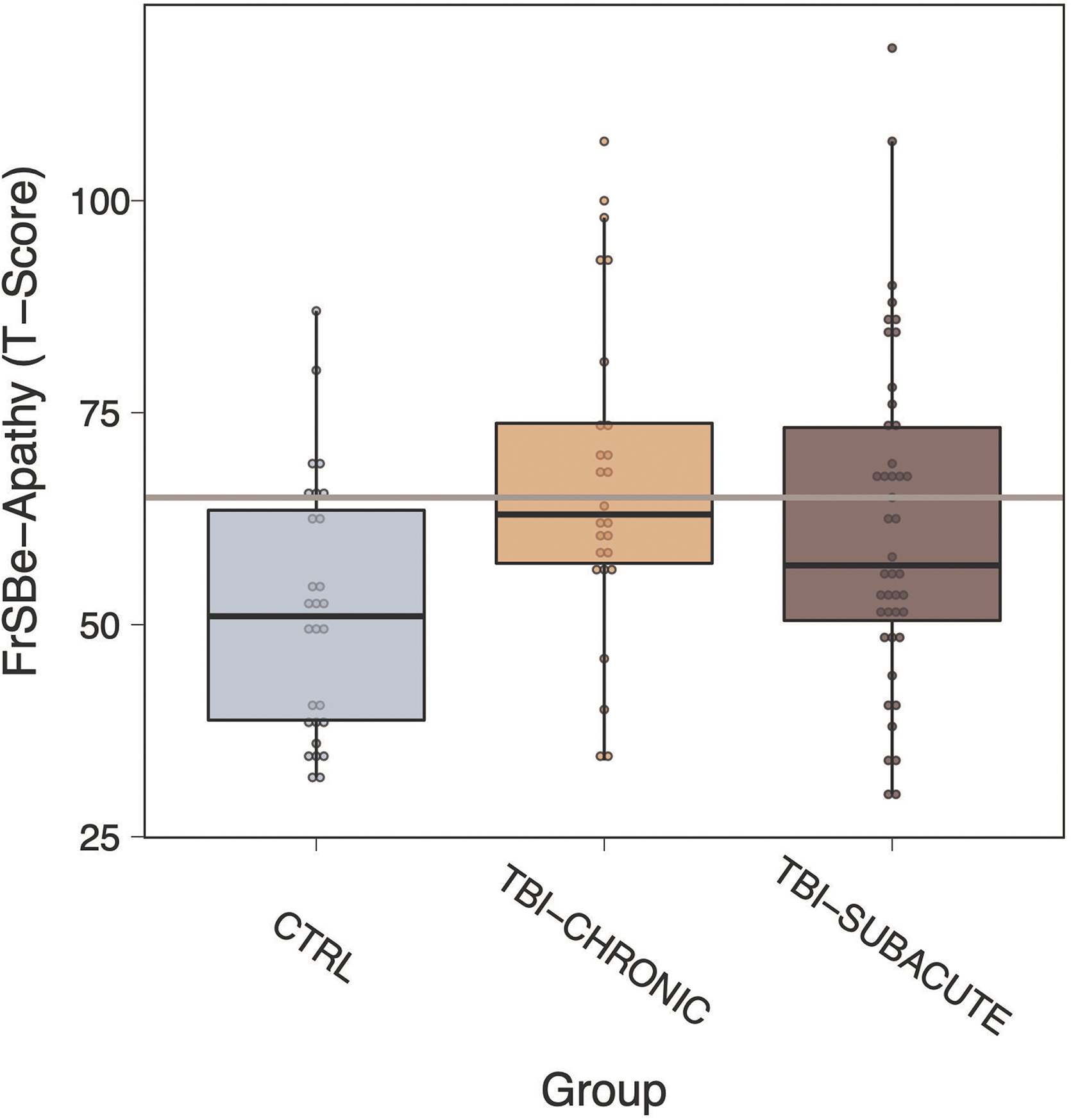
Self-reported apathy as a function of group, which was significantly higher in the subacute and chronic traumatic brain injury (TBI) groups relative to control (CTRL; all p HOLM ≤ 0.04). Gray line indicates clinically significant apathy. FrSBe, Frontal Systems Behavior Scale.
Demographics and other symptoms questionnaires
All participants completed a demographic questionnaire, and the patients provided additional information regarding the nature of their TBI (of particular importance: severity and time since injury). Participants also completed the Patient-Reported Outcomes Measurement Information System 28 battery, which provided patient-reported levels of depression, anxiety, and fatigability that were particularly important to take into account given their potential to share overlapping variance with patient apathy.
Image acquisition, processing, and analysis
Image acquisition
All MRI scans were acquired on a 3T Siemens Tim Trio system with a 32-channel phased-array head coil. The T1-weighted (T1w) structural MRI was acquired via a multi-echo MPRAGE sequence (5-echo; voxel size = 1 mm iso), and rsfMRI data were acquired with a single-shot, gradient-echo echoplanar pulse sequence using simultaneous multi-slice technology (repetition time = 460 msec; multi-band factor
Image preprocessing
Results included in this article come from pre-processing performed using fMRIprep v.1.5.0 31,32 (RRID:SCR_016216), which is based on Nipype v.1.2.2 33,34 (RRID:SCR_002502).
Anatomical data preprocessing
The T1w image was corrected for intensity nonuniformity (INU) with N4BiasFieldCorrection,
35
distributed with ANTs v.2.2.0
36
(RRID:SCR_004757), and used as T1w-reference throughout the workflow. The T1w-reference was then skull-stripped with a Nipype implementation of the
Volume-based spatial normalization to one standard space (MNI152NLin2009cAsym) was performed through nonlinear registration with antsRegistration (ANTS v.2.2.0), using brain-extracted versions of both T1w reference and the T1w template. The following template was selected for spatial normalization: ICBM 152 Nonlinear Asymmetrical template version 2009c 40 (RRID:SCR_008796; TemplateFlow ID: MNI152NLin2009cAsym).
Functional data pre-processing
For the BOLD run, the following pre-processing was performed. First, a reference volume and its skull-stripped version were generated using a custom methodology of fMRIprep. A deformation field to correct for susceptibility distortions was estimated based on fMRIPrep's fieldmap-less approach. The deformation field is that resulting from coregistering the BOLD reference to the same-subject T1w-reference with its intensity inverted. 41,42 Registration is performed with antsRegistration (ANTs v.2.2.0) and the process regularized by constraining deformation to be nonzero only along the phase-encoding direction, and modulated with an average fieldmap template. 43
Based on the estimated susceptibility distortion, an unwarped BOLD reference was calculated for a more accurate coregistration with the anatomical reference. The BOLD reference was then coregistered to the T1w reference using bbregister (FreeSurfer), which implements boundary-based registration. 44 Coregistration was configured with six degrees of freedom. Head-motion parameters with respect to the BOLD reference (transformation matrices and six corresponding rotation and translation parameters) are estimated before any spatiotemporal filtering using MCFLIRT (FSL v.5.0.9 45 ).
BOLD runs were slice-time corrected using 3dTshift from AFNI 20160207 46 (RRID:SCR_005927). The BOLD time-series were resampled to surfaces on the following spaces: fsaverage5. The BOLD time-series (including slice-timing correction when applied) were resampled onto their original, native space by applying a single, composite transform to correct for head-motion and susceptibility distortions. These resampled BOLD time-series will be referred to as pre-processed BOLD in original space, or just pre-processed BOLD.
The BOLD time-series were resampled into standard space, generating a pre-processed BOLD run in MNI152NLin2009cAsym space. First, a reference volume and its skull-stripped version were generated using a custom methodology of fMRIprep. Several confounding time-series were calculated based on the pre-processed BOLD: framewise displacement (FD), DVARS, and three region-wise global signals. The FD and DVARS are calculated for each functional run, both using their implementations in Nipype (following the definitions by Power and associates 47 ). The three global signals are extracted within the CSF, the WM, and the whole-brain masks.
In addition, a set of physiological regressors were extracted to allow for component-based noise correction (CompCor 48 ). Principal components are estimated after high-pass filtering the pre-processed BOLD time-series (using a discrete cosine filter with 128 sec cutoff) for the two CompCor variants: temporal (tCompCor) and anatomical (aCompCor). The tCompCor components are then calculated from the top 5% variable voxels within a mask covering the subcortical regions. This subcortical mask is obtained by heavily eroding the brain mask, which ensures it does not include cortical GM regions. For aCompCor, components are calculated within the intersection of the aforementioned mask and the union of CSF and WM masks calculated in T1w space, after their projection to the native space of each functional run (using the inverse BOLD-to-T1w transformation).
Components are also calculated separately within the WM and CSF masks. For each CompCor decomposition, the k components with the largest singular values are retained, such that the retained components' time-series are sufficient to explain 50% of variance across the nuisance mask (CSF, WM, combined, or temporal). The remaining components are dropped from consideration. The head-motion estimates calculated in the correction step were also placed within the corresponding confounds file. The confound time-series derived from head motion estimates and global signals were expanded with the inclusion of temporal derivatives and quadratic terms for each. 49 Frames that exceeded a threshold of 0.5 mm FD or 1.5 standardized DVARS were annotated as motion outliers.
All resamplings can be performed with a single interpolation step by composing all the pertinent transformations (i.e., head-motion transform matrices, susceptibility distortion correction when available, and coregistrations to anatomical and output spaces). Gridded (volumetric) resamplings were performed using antsApplyTransforms (ANTs), configured with Lanczos interpolation to minimize the smoothing effects of other kernels. 50 Nongridded (surface) resamplings were performed using mri_vol2surf (FreeSurfer). Many internal operations of fMRIprep use Nilearn v.0.5.2 51 (RRID:SCR_001362), mostly within the functional processing workflow. 2
FC analysis
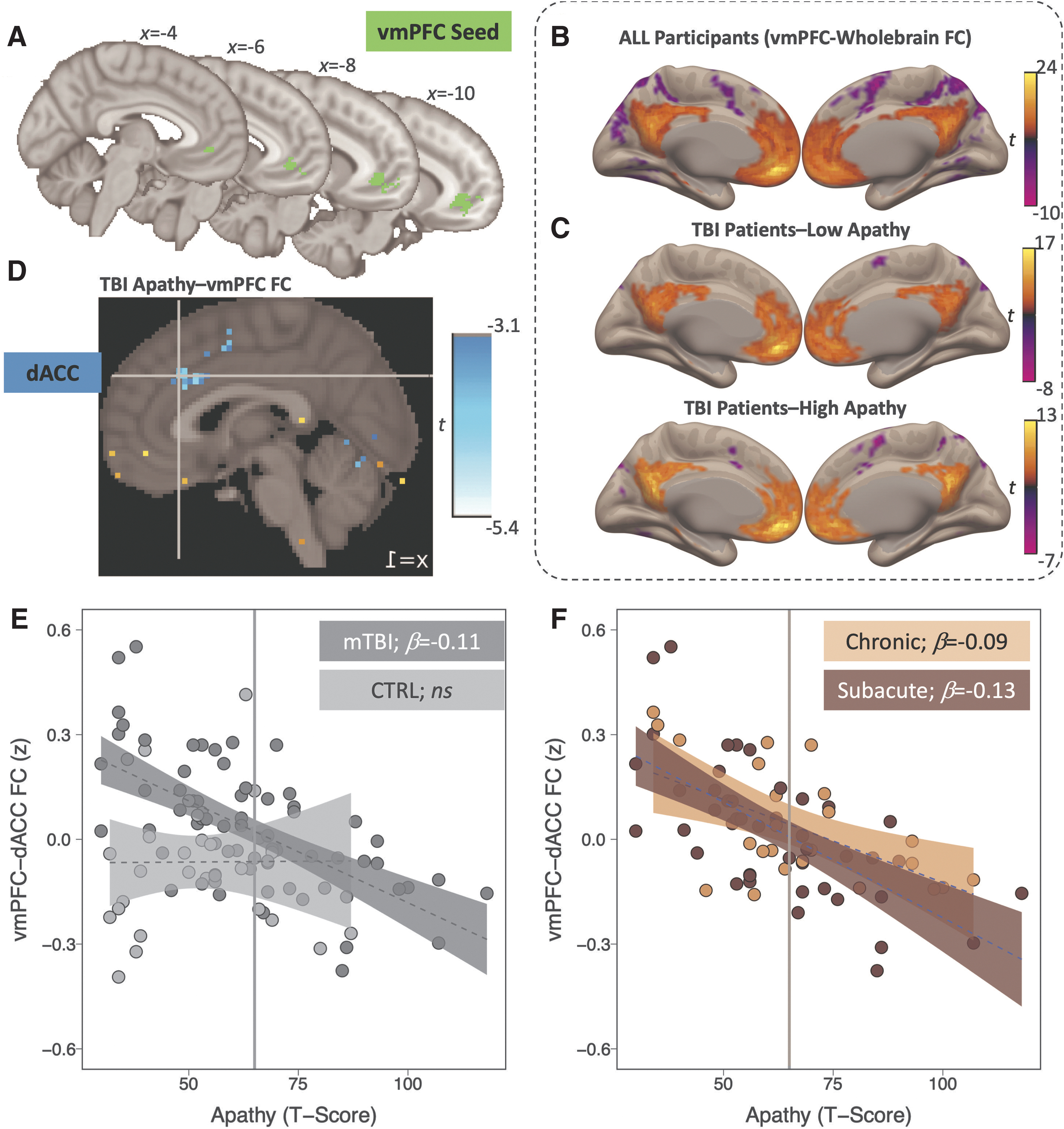
Seed-to-voxel FC maps were estimated in the CONN Toolbox v.19b 52 (RRID:SCR_009550), using a vmPFC seed from a previous publication on apathy and reward-guided decision-making in an independent chronic TBI cohort (Fig. 2A 8 ). Data input to CONN had completed pre-processing via fMRIprep, and FC was modeled using a bivariate correlation-based weighted generalized linear model. Notably, motion outliers defined via fMRIprep were excluded in first-level FC models, and we removed any participants who exceeded this scrubbing threshold on ≥50% of timepoints. 53 –55
(
In addition, in line with the standard CONN Toolbox recommendations, aCompCor component-based denoising from fMRIprep was adopted, but not tCompCor. 56,57 To briefly restate, the aCompCor procedure run within fMRIprep extracts signal components in regions of interest (ROIs) where signal fluctuations are unlikely to reflect neuronal activity (i.e., WM and CSF), and uses those components to reduce the influence of physiological artifacts in GM. 48 Second-level FC models were height thresholded at p < 0.001, and cluster corrected using a conservative whole brain familywise error correction (p FWE<0.05).
In our first step, we ran a one-sample t test to determine the wholebrain FC pattern of our vmPFC ROI, which was followed by a one-way analysis of variance (ANOVA) on vmPFC-wholebrain connectivity with group (i.e., subacute TBI, chronic TBI, and CTRL) as a factor. Next, we ran regression models examining the extent to which vmPFC-wholebrain FC was related to apathy, including both a constant and a demeaned apathy term as regressors in the model. We ran these FC regressions separately within CTRLs and TBI patients, given the significant differences in apathy across groups (see Results).
In addition, given the hypothesized association between ventral striatum-dACC connectivity and apathy in neurological disorders, 58 the TBI FC regression was also run with nucleus accumbens as the seed ROI (run separately by hemisphere; accumbens masks via the Harvard-Oxford Probabilistic Subcortical Atlas).
To provide further support that any observed effects in the TBI regression models were related to apathy per se, we ran a control model including covariates for head motion (FD), time since injury (log[days]), injury severity (given some chronic patients had a moderate TBI), the two other FrSBe subscores (disinhibition and executive dysfunctioning), as well as other affective and somatic constructs that share overlapping variance with apathy (namely: anxiety, depression, and fatigability).
Yet, statistically controlling for these variables as model covariates has limited utility, 59 and to further validate our TBI apathy regression model, we also put it through a repeated k-fold cross-validation using the caret package in R v.3.5.0 60,61 (RRID:SCR_001905). Briefly, this procedure involved dividing the dataset into k = 10 nonoverlapping folds, with each fold given the opportunity to be used as a test dataset to verify the model fit while all other folds are used as training datasets. This k-fold procedure was repeated 10 times to provide maximally reliable parameter estimates. In addition, given that the subacute group had stricter requirements for TBI verification than the chronic group, we repeated this k-fold cross-validation focusing specifically on the subacute group to confirm that any observed effects were driven by TBI-related apathy per se (see section Control models).
Last, to determine the direction of any observed apathy-related vmPFC FC effects in the TBI regression (i.e., whether they might be related to pathological changes in vmPFC FC and in patients with high apathy scores, or compensatory changes in FC that may promote resilience to apathy development after a TBI), we ran group-wise one-way ANOVAs on any significant FC effects observed in our regression contrasting CTRL, TBI patients with clinically significant apathy (TBIapathy: T ≥ 65; n = 31), and TBI patients with apathy scores below clinical threshold (TBIlow_apathy: T < 65; n = 39).
Results
Apathy
There was a significant group difference in apathy (F2,95 = 5.15, p = 0.007, h2 p = 0.10; Fig. 1). Specifically, post hoc tests revealed that apathy in the subacute TBI group (EMM = 62, 95% confidence interval [CI] = 56 to 67) was significantly higher than apathy in the CTRL group (EMM = 51, 95% CI = 45 to 58; t95 = 2.34, pHOLM = 0.043). Apathy was also significantly higher in patients with chronic TBI (EMM = 67, 95% CI = 60 to 74) relative to CTRLs (t95 = 3.12, pHOLM = 0.007). Apathy levels, however, did not appear to differ between subacute and chronic patients with TBI (pHOLM = 0.254). Further, a linear regression model did not reveal a significant effect of time since injury (in log[days]) on apathy across acute and chronic TBI groups (b = 2.47, se = 2.07, t = 1.19, p = 0.237).
vmPFC-wholebrain FC
One-sample t test and one-way ANOVA
Across the sample, vmPFC was positively correlated with the default mode network and anticorrelated to salience, dorsal attention, and sensorimotor networks (p FWE<0.05; Fig. 2B; Table 2). This canonical wholebrain FC pattern did not differ between the CTRL, subacute TBI, and/or chronic TBI groups.
Wholebrain vmPFC Functional Connectivity for All Participants
MNI, Montreal Neurological Institute; FWE, family-wise error.
Apathy regression model
Next, we examined the association between apathy and vmPFC FC separately in the CTRL and TBI groups. The CTRL group did not demonstrate any significant modulation of vmPFC FC as a function of apathy. In contrast, the TBI group demonstrated significant negative modulation of vmPFC FC to a cluster in dACC as a function of apathy (Fig. 2D; Table 3). Neither left nor right nucleus accumbens regression models evidenced any significant modulation as a function of apathy in the current dataset. Ten repeats of a 10-fold cross-validation evidenced the robustness of the vmPFC regression model across repeated resampling of training and test sets (R 2 mean = 0.443), suggesting that higher apathy scores predicted a decrease in vmPFC-dACC FC reliably across repeats within the TBI sample (b = -0.114, SE = 0.017, t = -6.59, p < 0.001).
Modulated vmPFC Functional Connectivity as a Function of Apathy in TBI Patients
MNI, Montreal Neurological Institute; ROI, region of interest; FWE, family-wise error.
This cross-validation procedure was reproduced within the subacute and chronic TBI subgroups, producing identical relationships at both injury stages (subacute: R2 mean = 0.579, b = -0.129, SE = 0.022, t = -5.87, p < 0.001; chronic: R2 mean = 0.759, b = -0.093, SE = 0.028, t = -3.27, p < 0.001). This provides further evidence that the relationship between apathy and vmPFC-dACC FC is independent of time since injury, and given the rigid TBI verification criteria within the subacute group, these results provide strong evidence that what we observed is related to TBI-related apathy symptoms per se.
Post hoc apathy group contrast
The vmPFC FC was contrasted between TBI patients with (TBIapathy) or without (TBIlow_apathy) clinically significant apathy (FrSBe clinically significant threshold ≥65). Notably, whereas vmPFC was anticorrelated to dACC in the TBIapathy subgroup, this cluster was absent in the TBIlow_apathy subgroup, although the group contrast did not reach significance after wholebrain FWE correction (Fig. 2C).
When directly comparing extracted vmPFC-dACC values between groups, FC significantly varied between CTRLs, TBIapathy, and TBIlow_apathy (F 2,95 = 13.24, p < 0.001, h 2 p = 0.22). Specifically, post hoc tests revealed that vmPFC-dACC FC did not differ between TBIapathy (EMM = -0.07, 95% CI = -0.13 to -0.01) relative to CTRLs (EMM = -0.06, 95% CI = -0.13 to -0.002; t 95 = -0.164, p HOLM = 0.870).
The vmPFC-dACC FC, however, was significantly higher in the TBIlow_apathy subgroup (EMM = 0.11, 95% CI = 0.06 to 0.16) relative to both TBIapathy (t 95 = 4.50, p HOLM <0.001), and CTRLs (t 95 = 4.19, p HOLM <0.001). Therefore, the observed negative association between apathy and vmPFC-dACC FC in the TBI group was not indicative of hypoconnectivity in patients with TBI who have clinically significant apathy, but rather was driven by vmPFC-dACC hyperconnectivity in patients with TBI who have apathy scores below clinical threshold.
Control models
We ran a series of control models to further strengthen our confidence in our results. First, given that education history differed between groups, we ran an analysis of covariance on apathy as a function of group after covarying for years of education. The effect of group on apathy remained significant after covarying for education (p = 0.011), as did the intergroup differences between CTRL and both subacute and chronic TBI patients (all p ≤ 0.03).
In addition, to confirm that vmPFC-dACC FC in the regression model was associated with apathy per se, we ran a multiple regression model of vmPFC-dACC FC as a function of apathy in TBI, including covariates for the other FrSBe dimensions (executive dysfunction and disinhibition), head motion (FD), time since injury (log[days]), injury severity (given some chronic patients had a moderate TBI), anxiety, depression, and fatigability. Compellingly, the effect of apathy on vmPFC-dACC FC remained significant after covarying for all other variables (b = -0.112, SE = 0.032, t = -3.45, p = 0.001).
Last, the vmPFC-dACC FC difference as a function of apathy group was not driven by a confounding demographic or other variable, as the TBIapathy and TBIlow_apathy subgroups were matched with respect to education history, sex, time since injury, and head motion (all ps ≥ 0.21). The TBIapathy subgroup did differ from TBIlow_apathy with respect to age (p = 0.030), but including age as a covariate did not change the effect of group on vmPFC-dACC FC (TBIlow_apathy vs. TBIapathy: p HOLM < 0.001; TBIlow_apathy vs. CTRLs: p HOLM < 0.001). Therefore, we observed a clear and lasting increase in TBI patient apathy relative to CTRLs, and a specific and robust association between apathy and vmPFC-dACC FC in TBI patients, both of which appeared to be statistically independent from time since injury or several other potential confounding variables.
Head motion control analyses
Head motion can systematically contaminate FC patterns in resting-state fMRI, and several control analyses were performed to ensure that head motion did not confound the inferences in the current study. First, we examined whether head motion (mean FD) varied systematically as a function of our key explanatory and outcome measures. Mean FD did not vary between CTRL participants and either mTBI group (subacute: p = 0.770; chronic: p = 0.304), was not correlated with either apathy scores (r 96 = -0.054, p = 0.594), and was not correlated with vmPFC-dACC FC (r 96 = -0.067, p = 0.513). Therefore, head motion was unlikely to introduce systematic variance in FC that might have confounded the current results.
In addition, we confirmed the reliability and robustness of our findings across low- and high-motion subjects. Concerning reliability, we randomly selected subsets of the fMRI time-series for each subject and computed the difference in vmPFC-dACC FC between these subsets and the same FC estimate using the full time-series.cf.62 Although vmPFC-dACC FC estimates clearly became more reliable as more data were included (b = 0.04, se = 0.008, p < 0.0000001), the estimate stabilized at ≈30% or more of the data included (Supplementary Result 1)—i.e., well below the 50% minimum subject exclusion in the current protocol.
Relatedly, given the potential for scrubbing to disrupt the temporal autocorrelation structure of resting-state fMRI time-series, we reran our analyses without scrubbing and replicated the negative association between vmPFC-dACC FC and apathy in mTBI (Supplementary Result 2).
Finally, to confirm the results were robust to the selected subject motion threshold (≥50% scrubbed timepoints), we recomputed the correlation between apathy and vmPFC-dACC FC in mTBI iteratively at thresholds ranging from 5% to 50% in steps of 5%. At each threshold, the correlation remained significant (5%: r 42 = -0.56, p < 0.0001; 10%: r 54 = -0.60, p < 0.0001; 15%: r 61 = -0.60, p < 0.0001; 20%: r 64 = -0.61, p < 0.0001; 25%: r 66 = -0.60, p < 0.0001; 30%: r 67 = -0.61, p < 0.0001; 35–50%: r 68 = -0.63, p < 0.0001). Collectively, these analyses establish that the primary results reported here were unlikely to be systematically confounded by motion, were reliable given the proportion of the data included in the analysis, and were robust regardless of scrubbing and subject-level motion exclusion criteria.
Discussion
The aim of this study was to elucidate the motivational neural circuit disruptions or adaptations that shape the presence or absence of apathy in TBI. We did not find evidence for any vmPFC or nucleus accumbens FC correlates of clinically significant apathy post-TBI. We did find novel and robust evidence, however, for a potential compensatory mechanism: vmPFC-dACC hyperconnectivity in TBI patients with apathy scores below clinical threshold. This finding replicates previous demonstrations of resting-state hyperconnectivity in TBI, 19,20 which have been argued to offset disruptions in structural connectivity 63 and help to reestablish network communication between densely connected cortical hubs. 21
Our findings suggest that vmPFC-dACC hyperconnectivity may represent one such adaptive network response to brain injury, reducing apathy in TBI patients. Given the vmPFC ROI used in the current study has been causally implicated in reward valuation, 8 this compensatory effect may be related to a preserved motivation to initiate reward-seeking behaviors in TBI patients with apathy levels below clinical threshold.
There is a growing literature implicating vmPFC and dACC aberrations in clinical amotivation across psychiatric and neurological illness. 64 A prominent functional model of vmPFC is that it encodes the anticipated value of obtainable outcomes. 7,65 Relatedly, dACC has been implicated in encoding one's willingness to exert effort to obtain rewards. 58 A deficit in reward valuation and/or willingness to exert effort may therefore represent transdiagnostic pathways to amotivation. 64 The current findings add to this work by suggesting that enhanced vmPFC-dACC FC may help to maintain motivational drive in patients who do not experience amotivation. It is difficult to gleam the specific functional role of vmPFC-dACC FC in the current resting-state study, and future task fMRI studies will be critical to determining the specific computational role of this circuit in shaping motivated behavioral outcomes in TBI.
It is unclear why our findings did not implicate any FC correlates underlying clinically significant apathy in TBI. Previous structural brain imaging studies have implicated volume loss in vmPFC in apathy symptoms in TBI, 8,66 but to our knowledge, no previous functional imaging studies have examined the neural correlates of apathy in this patient population. One possibility is that evoked activation of motivational neural circuits during value- or effort-based decision making tasks may be more sensitive to the neuropathologies underlying clinical apathy in TBI than intrinsic resting-state FC.
This approach would be in line with work on the neurobiology of apathy in Parkinson disease, where value- and effort-based decision making tasks have proven sensitive for detecting dysfunctional dopaminergic regulation of motivated behaviors, leading to clinical symptoms including apathy and impulsivity. 67 –69 A similar task-evoked fMRI approach is rarely adopted to study motivated decision-making processes and associated clinical symptoms in the TBI population, and future studies should pursue it to try to elucidate the neural correlates of clinically significant apathy in TBI.
Given the established role of vmPFC and dACC in motivated decision making, the current study suggests that rehabilitation interventions should aim to promote recruitment of these brain regions in patients with apathy. For example, vmPFC enhancement has been found to underlie reductions in attention to threat cues in anxious individuals following cognitive bias training, 70 and a modified version of this approach might be adopted to enhance reactivity to cues related to goal-directed behavior in TBI patients with clinical apathy. Further, given the utility of inhibitory vmPFC-targeted noninvasive brain stimulation to disrupt value-based decision making and decrease cue reactivity, 71,72 an excitatory version of this approach may prove fruitful for normalizing motivation and facilitating better functional outcomes for TBI patients experiencing clinically significant apathy.
Footnotes
Funding Information
This research was funded by the National Institute of General Medical Sciences (P20GM109089, PI: CWS).
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Results 1
Supplementary Results 2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.