
Abstract
Physician-prescribed rest after traumatic brain injury (TBI) is both commonplace and an increasingly scrutinized approach to TBI treatment. Although this practice remains a standard of patient care for TBI, research of patient outcomes reveals little to no benefit of prescribed rest after TBI, and in some cases prolonged rest has been shown to interfere with patient well-being. In direct contrast to the clinical advice regarding physical activity after TBI, animal models of brain injury consistently indicate that exercise is neuroprotective and promotes recovery. Here, we assessed the effect of low and moderate intensity treadmill exercise on functional outcome and hippocampal neural proliferation after brain injury. Using the controlled cortical impact (CCI) mouse model of TBI, we show that 10 days of moderate intensity treadmill exercise initiated after CCI reduces anxiety-like behavior, improves hippocampus-dependent spatial memory, and promotes hippocampal proliferation and newborn neuronal survival. Pathophysiological measures including lesion volume and axon degeneration were not altered by exercise. Taken together, these data reveal that carefully titrated physical activity may be a safe and effective approach to promoting recovery after brain injury.
Introduction
Standard clinical practice for traumatic brain injury (TBI) involves the recommendation of physical rest after injury until symptoms (e.g. headache, dizziness, nausea) resolve, followed by a gradual return to pre-injury levels of activity. 1 –3 The duration of prescribed rest is not standardized, and although the most up-to-date clinical guidelines recommend a brief period of rest (∼2 days), prolonged strict rest for ≥5 days may also be prescribed. 4 The clinical justifications given for recommending rest include preventing re-injury, maximizing recovery potential, and avoiding post-concussive syndrome. 5 This approach is potentially problematic for several reasons. First, this strategy relies on improvement of often self-reported and subjective symptoms as a guide for returning to pre-injury activity; however, resolution of TBI symptoms does not necessarily correspond to a full recovery. 6,7 Second, there is a significant cost associated with prolonged rest, including social isolation, loss of school and work time, and cardiac deconditioning. 8,9 Finally, the evidence for beneficial outcomes associated with a temporary reduction of physical activity is minimal. For example, rest does little to address the long-term consequences of brain injury, such as mood disorders and memory impairment. 10 These symptoms occur despite adherence to prescribed physical rest and are alleviated through physical activity. 11 –13 Outcomes from randomized controlled trials that assessed the effectiveness of prolonged bed rest (5–6 days) versus short-term or no rest (0–2 days) in TBI patients ranged from no difference 14 to slower symptom resolution 15 as a result of adherence to prolonged rest. On the contrary, concussion rehabilitation that includes various forms of early introduction of physical activity was found to improve mobility, 16 balance, 17 and mood, 13 as well as reducing post-concussion symptoms. 18,19
Animal models of brain injury generally report neuroprotective effects of exercise, although outcome variability caused by differences in voluntary versus forced exercise, injury severity, and exercise timing make it difficult to tease out the effectiveness of post-TBI exercise. 20 –25 Overall, physical exercise reduces neuroinflammation, improves mitochondrial function, increases the levels of neurotrophic and angiogenic factors, improves cerebral blood flow, and alleviates cognitive and stress-related dysfunction. 26 –28 Importantly, in brain injury models, these outcomes are dependent on exercise intensity; lower intensity treadmill exercise after TBI reduces neuronal cell death 29 and improves memory, 30 whereas higher intensity exercise impairs post-injury cognitive recovery, 23 highlighting the need to carefully titrate post-injury exercise intensity in order to achieve improved recovery rates. Likewise, data from stroke models indicate that synaptic plasticity, 31 neurogenesis, 32 and hippocampus-dependent memory 33 are differentially affected by physical activity based on exercise intensity. However, less is known regarding whether, and to what extent, varying intensities of exercise affect hippocampal proliferation and survival of newborn neurons following a brain injury. Therefore, using the controlled cortical impact (CCI) mouse model of TBI, the present study was designed to determine the effect of low and moderate intensity treadmill exercise on cognitive recovery and survival of newborn hippocampal neurons in brain injured mice.
Methods
Animals
All procedures were conducted on male Swiss–Webster mice (4–6 weeks of age) derived from breeders purchased from Charles River (Wilmington, MA). Mice in cohort 1 (Fig. 1A) were bred at the Ohio State University (OSU) and cohort 2 (Fig. 1B) was bred at West Virginia University (WVU). Pups were weaned at 21 days of age into a standard mouse cage with ad libitum access to food and filtered tap water. Animals at OSU were housed in a 14:10 light–dark cycle and animals at WVU were housed in a 12:12 light-dark cycle. All procedures were approved by the Institutional Animal Care and Use Committees at OSU and WVU.
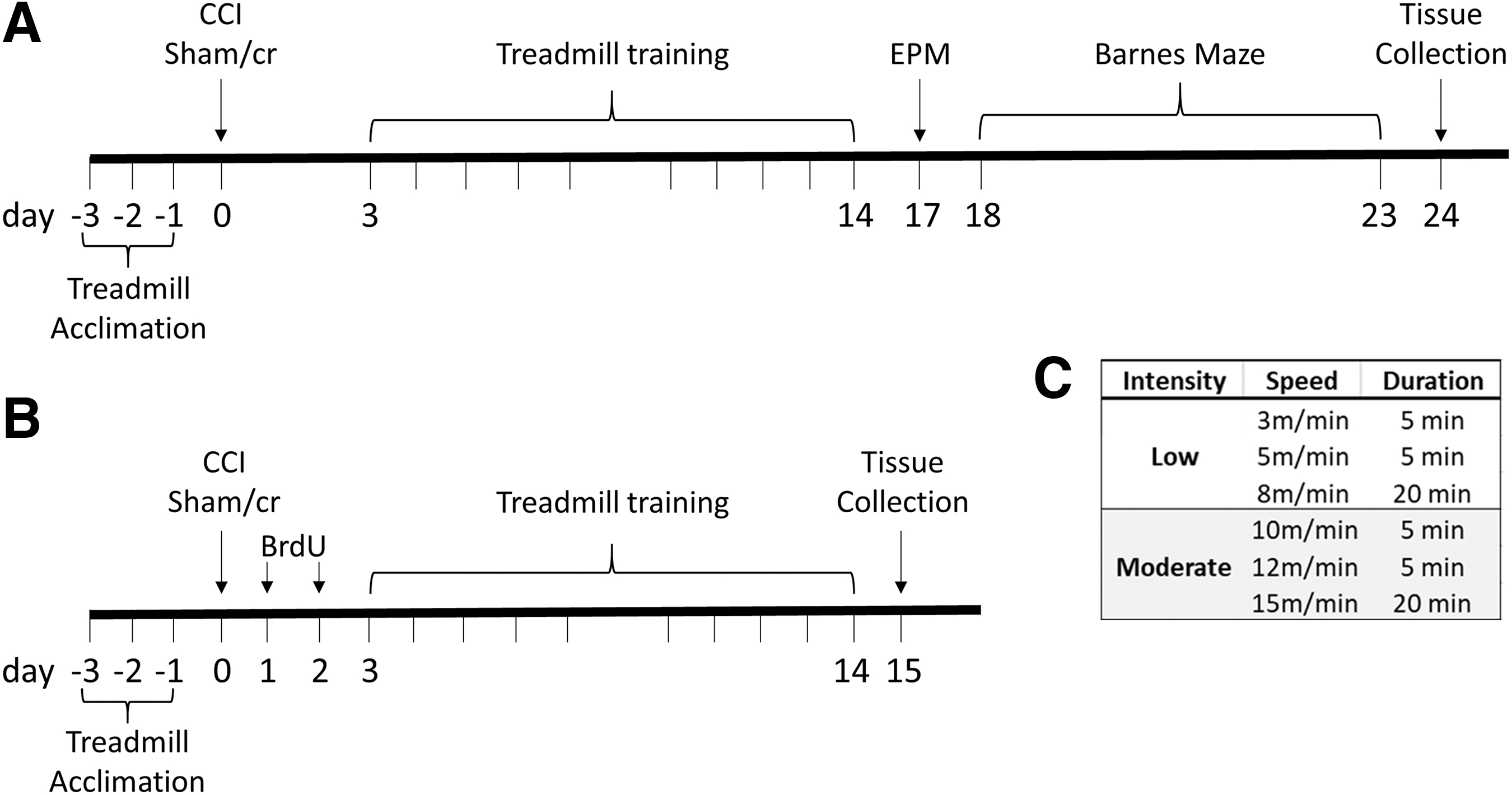
Experimental timeline. Mice underwent a controlled cortical impact (CCI) or sham/cr (craniectomy only).
TBI
TBI was induced via a CCI model using an established protocol. 34 Briefly, mice were anesthetized via inhaled isoflurane (3% in oxygen) and secured into a stereotaxic frame equipped with a heating pad (Stoelting). Their skulls were exposed and a 4 mm craniectomy was performed over the left parietal bone. A 3 mm plunger was retracted and accelerated into the brain causing a 1.2 mm deformation at a velocity of 5 m/sec with a dwell time of 200 msec. The control group (sham/cr) consisted of mice that received a craniectomy; however, the plunger was then gently lowered to the surface of the brain and immediately retracted. After CCI or sham/cr, the incision was sutured with 6/0 nylon suture and mice were replaced into their home cages. All mice were treated subcutaneously with bupivacaine (1 mg/kg) and meloxicam (5 mg/kg) for analgesia, and monitored daily post-operatively. A total of nine mice were euthanized post-operatively because of insufficient recovery (i.e., they demonstrated lethargy, dehydration, or pain not significantly improved by analgesic treatment) by day 2 after injury.
Forced exercise paradigm
All mice were acclimated to a three-lane stationary treadmill (Columbus Instruments) at a speed of 6 m/min for 3 consecutive days (10 min/day). Acclimation was performed prior to injury to avoid acute injury-induced variation in ability to perform the novel task. 23 Beginning 72 h after CCI or sham/cr, mice were randomly assigned to one of three exercise conditions (sedentary, low intensity, or moderate intensity) for 10 days (excluding the weekend) at 30 min/day. All acclimation and exercise conditions were performed at a 0 degree incline, and no negative stimuli (i.e., electric shock) were used in this exercise paradigm. Low and moderate intensity treadmill exercise conditions were adapted from previously published reports 29,35 (see Fig. 1C for specific training conditions), mice in sedentary conditions were also placed on the treadmill for 30 min daily but the treadmill was not turned on. Two mice were removed from the study because of an inability to perform the full treadmill paradigm. The following groups were formed for two cohorts (cohort 1 was used for behavioral analysis and histology, cohort 2 was used for bromodeoxyuridine / 5-bromo-2'-deoxyuridine [BrdU] labeling of newborn neurons): sham/cr sedentary (cohort 1 n = 8, cohort 2 n = 8), sham/cr low intensity (cohort 1 n = 8, cohort 2 n = 8), sham/cr moderate intensity (cohort 1 n = 7, cohort 2 n = 9), CCI sedentary (cohort 1 n = 8, cohort 2 n = 7), CCI low intensity (cohort 1 n = 7, cohort 2 n = 7), and CCI moderate intensity (cohort 1 n = 7, cohort 2 n = 8).
Elevated plus maze (EPM)
Anxiety-like behavior was measured in cohort 1 17 days post-injury (dpi) (Fig. 1A) on an EPM according to an established protocol. 36 Briefly, mice were placed in the center of a plus-shaped maze elevated 50 cm off the floor and consisting of two “open” arms and two “closed” arms. All arms were 50 cm long and 10 cm wide. The closed arms were enclosed with 40 cm high walls. Mice were recorded on the EPM for 5 min by a blinded observer, and videos were analyzed using ANY-maze (Stoelting) for number of entries into each arm and path length. The maze was cleaned with 70% ethanol after each trial. Decreased activity in the open arms is indicative of anxiety-like behavior.
Barnes maze
Spatial learning and memory performance were measured in cohort 1 18–23 dpi (Fig. 1A) by a blinded observer on the Barnes maze as previously reported. 37 Briefly, the maze (Stoelting) consists of a 91 cm diameter circular arena with 18 holes around the perimeter, with one hole leading to a dark escape box (target hole). Over a 5-day training period, mice underwent three trials/day during which they were individually placed onto the center of the maze, and video recorded for up to 120 sec/trial. Latency to find the target hole was recorded for each trial daily. On the 6th day, the escape box was removed and mice were placed on the maze for 90 sec for the probe trial. The probe trial assessed time spent in the quadrant that formerly contained the target. Behavioral assessment was performed using ANY-maze (Stoelting). The maze and escape box were cleaned with 70% ethanol after each trial.
BrdU administration
In order to label newly generated cells, mice in cohort 2 (Fig. 1B) were injected intraperiotoneally (i.p.) with BrdU (50 mg/kg in saline, Sigma Aldrich). 38 All mice were injected twice daily, 6 h apart, on 1 and 2 dpi. The timing of the injections allowed for labeling of cells that were generated after the injury, but before the start of the exercise protocol, in order to assess the effect of exercise intensity on the survival of newborn cells.
Tissue processing and immunohistochemistry
Brain tissue fixation was performed following an overdose with sodium pentobarbital (200 mg/kg i.p.) and transcardial perfusion with 4% paraformaldehyde. Mice in cohort 1 were euthanized 24 dpi and mice in cohort 2 were euthanized 15 dpi. Forty-micron sections were sliced coronally on a cryostat throughout the forebrain. Free-floating immunohistochemistry was performed for microglia (Iba1, Wako) and proliferating cells (Ki67, Abcam). Tissue was washed with 0.1M phosphate buffered saline (PBS), blocked with 10% normal goat serum (NGS), and incubated overnight with rabbit anti-Ki67 antibody (1:2000) or rabbit anti-Iba1 (1:1000) at room temperature. The following day, tissue was incubated in biotinylated goat anti-rabbit secondary antibody for 2 h, and the staining was visualized using the diaminobenzidine protocol (Vector Laboratories). Ki67-positive cells were counted in the ipsilateral and contralateral subgranular zone (SGZ). The SGZ of both upper and lower blades of the dentate gyrus were assessed in three dorsal hippocampal sections (approximate coordinates: -1.70 mm to -2.18 mm from Bregma). Ki67-positive cell counts are reported as the average of three counts per hemisphere. For microglial analysis, bilateral images were taken lateral to the cornu ammonis (CA)2/CA3 region of the hippocampus (bordering the site of direct cortical damage approximately between -1.70 and -2.18 mm from Bregma). Iba1-positive microglia were assessed in a 0.043mm2 region of interest (ROI) superimposed over a 10x image of the peri-lesional cortex, hippocampus CA1, or dentate gyrus in the ipsilateral and contralateral hemispheres using FIJI software. 39 The activation state of Iba-1 positive microglia within the ROI was assessed qualitatively based on morphological characteristics (ramified: small cell body and thin/long processes, or activated: enlarged or amoeboid cell body with shortened/thick processes). Results were averaged within each group, and data are reported as percent ramified or activated.
Double immunofluorescence staining for BrdU/doublecortin (DCX) was also performed on free-floating tissue. Tissue was washed with 0.1 M PBS, blocked with 10% normal horse serum, and incubated overnight with goat anti-DCX (Santa Cruz, 1:500) antibody at room temperature. The following day, tissue was incubated in donkey anti-goat secondary antibody (AlexaFluor 594) for 2 h, incubated with 2N HCl at 37°C for 1 h and 0.05M boric acid for 10 min, blocked with 10% NGS, and incubated overnight with rat anti-BrdU (1:200) at room temperature. On the 3rd day, tissue was incubated in goat anti-rat secondary antibody (AlexaFluor 488). BrdU- and DCX-positive cells were counted bilaterally in the SGZ and granular zone of the dentate gyrus in three dorsal hippocampal sections using the same coordinates as described. Data are reported as the average of three counts per hemisphere, as well as percent BrdU-cells that co-express DCX.
Axon degeneration
Axon degeneration was assessed using a commercial silver staining kit (FD NeuroTechnologies) according to manufacturer's instructions. Damaged axons were qualitatively assessed using an adapted version of a previously reported method (0 = few silver stained axons, 3 = dense axonal degeneration throughout white matter tracts). 37
Cresyl violet staining
Lesion volume was assessed via cresyl violet staining. Cryosections were mounted onto glass slides and hydrated in graded ethanol washes (100%, 95%, 70%). Tissue was then incubated for 3 min in 0.1% cresyl violet and destained in 95% ethanol containing glacial acetic acid. Tissue was then dehydrated, dipped in xylene, and cover-slipped with Permount. Cortical lesion volume was calculated by outlining the cortex on the ipsilateral and contralateral sides of every 10th slice in the forebrain. A total of six sections (400 μm apart) were assessed within a pre-defined region (Bregma -1.06 mm and -2.92 mm). Percent lesion volume was calculated using the following formula: [(contralateral cortex volume – ipsilateral cortex volume)/contralateral cortex volume] × 100.
Statistical analysis
Statistical analysis was performed using SPSS Version 26 (IBM Corp.) Immunohistochemistry, EPM, Barnes maze probe, and lesion volume were assessed via analysis of variance (ANOVA) (injury × activity). Barnes maze learning trials were assessed via repeated measures ANOVA (injury × activity). All significant main effects were followed up by a least significant differences post-hoc analysis. Qualitative axon damage data were analyzed via the non-parametric Mann–Whitney U test.
Results
Treadmill exercise improves neurobehavioral measures in brain-injured mice
In order to determine the effect of exercise on neurobehavioral outcomes in brain- injured mice, mice in cohort 1 underwent 10 days of sedentary, low, or moderate intensity treadmill exercise after CCI or sham/cr injury. Anxiety-like behavior was assessed via the EPM. Among sedentary mice, CCI significantly reduced open arm entries (F 1,12 = 5.728, p < 0.05) relative to sham/cr, however both low intensity (p > 0.05) and moderate intensity (p > 0.05) increased open arm entries in CCI mice to sham/cr levels. Moreover, there was a significant main effect of exercise on open arm entries among CCI-injured mice (F 2, 19 = 3.909, p < 0.05), such that moderate intensity (p < 0.05) but not low intensity exercise (p = 0.057) significantly increased open arm entry in CCI mice relative to CCI mice in the sedentary condition (p < 0.05; Fig. 2A). Similarly, CCI significantly reduced distance traveled in open arms among sedentary mice (F 1,13 = 13.942, p < 0.01). A main effect of exercise was again evident among CCI mice (F 2,19 = 4.744, p < 0.05), such that moderate intensity (p < 0.05) but not low intensity (p > 0.05) exercise significantly increased distance traveled in the open arms in CCI mice relative to CCI mice in the sedentary condition (Fig. 2B).
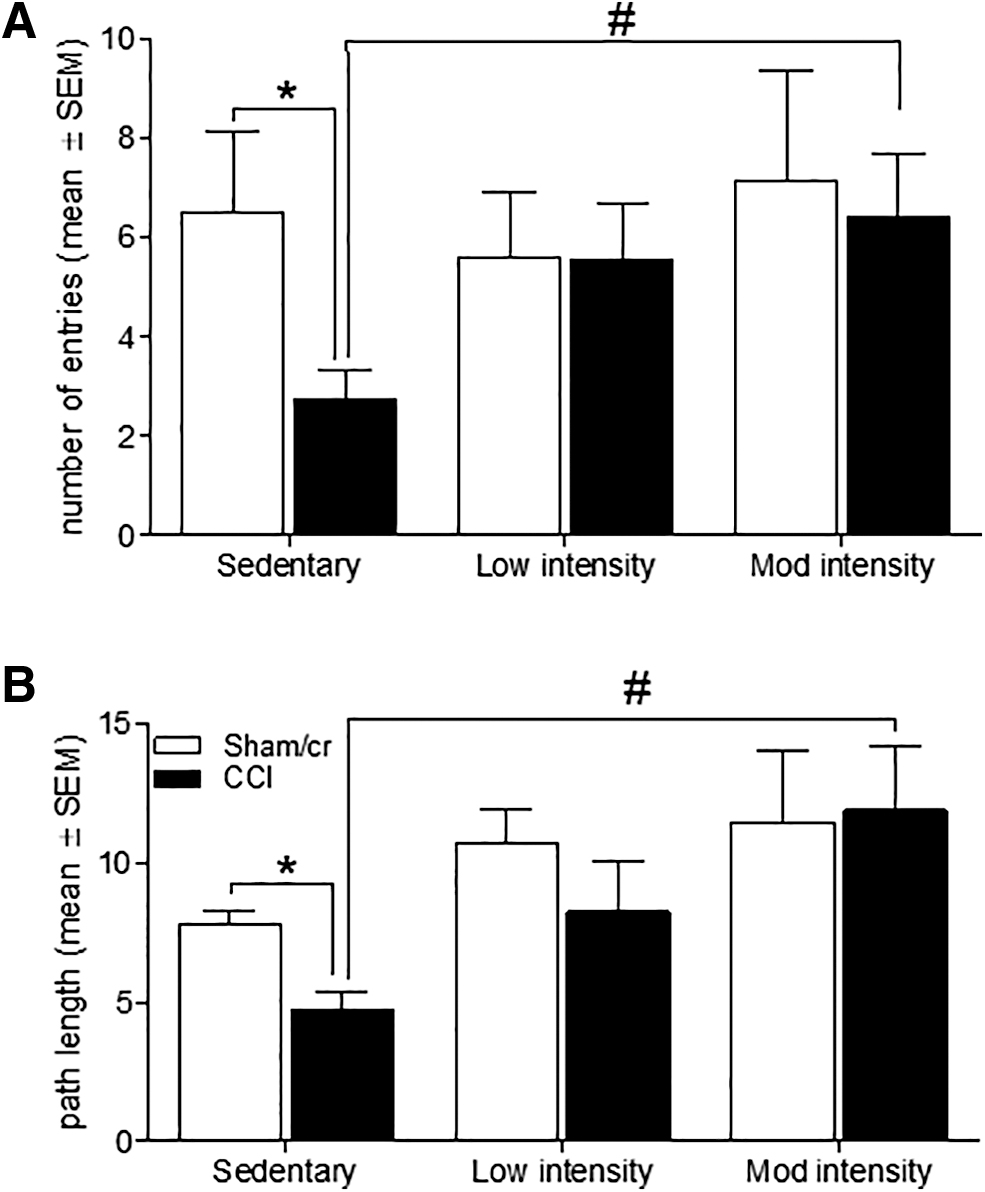
Treadmill exercise reduces anxiety-like behavior after brain injury.
Cognitive function was assessed via a hippocampus-dependent task on the Barnes maze. A repeated measures analysis revealed main effects of injury (F 1,37 = 7.506, p < 0.01) and exercise (F 2,37 = 4.088, p < 0.05) on latency to escape across all trials. Specific comparisons revealed that sedentary CCI mice had significantly longer latencies than all sham/cr mice (p < 0.05 regardless of activity level). Low-intensity exercise resulted in an intermediate effect in CCI mice such that latencies were no longer significantly different from those of sham sedentary mice (p > 0.05) but were still longer than those of sham/cr mice in either low or moderate exercise conditions (p < 0.05). Critically, moderate intensity exercise significantly reduced latency in CCI mice compared with both sedentary and low intensity CCI groups (p < 0.05; Fig. 3A). In the probe trial, an ANOVA revealed a main effect of exercise on percent time spent in the target quadrant (quadrant 3, which had contained the escape box during training sessions; F 2,36 = 3.306, p < 0.05). Low intensity exercise has an intermediate effect on the probe trial and was not significantly different from either sedentary or moderate intensity conditions (all p > 0.05). However, moderate intensity exercise significantly increased percent time in the target quadrant in CCI mice compared with the sedentary CCI group (p < 0.05; Fig. 3B).
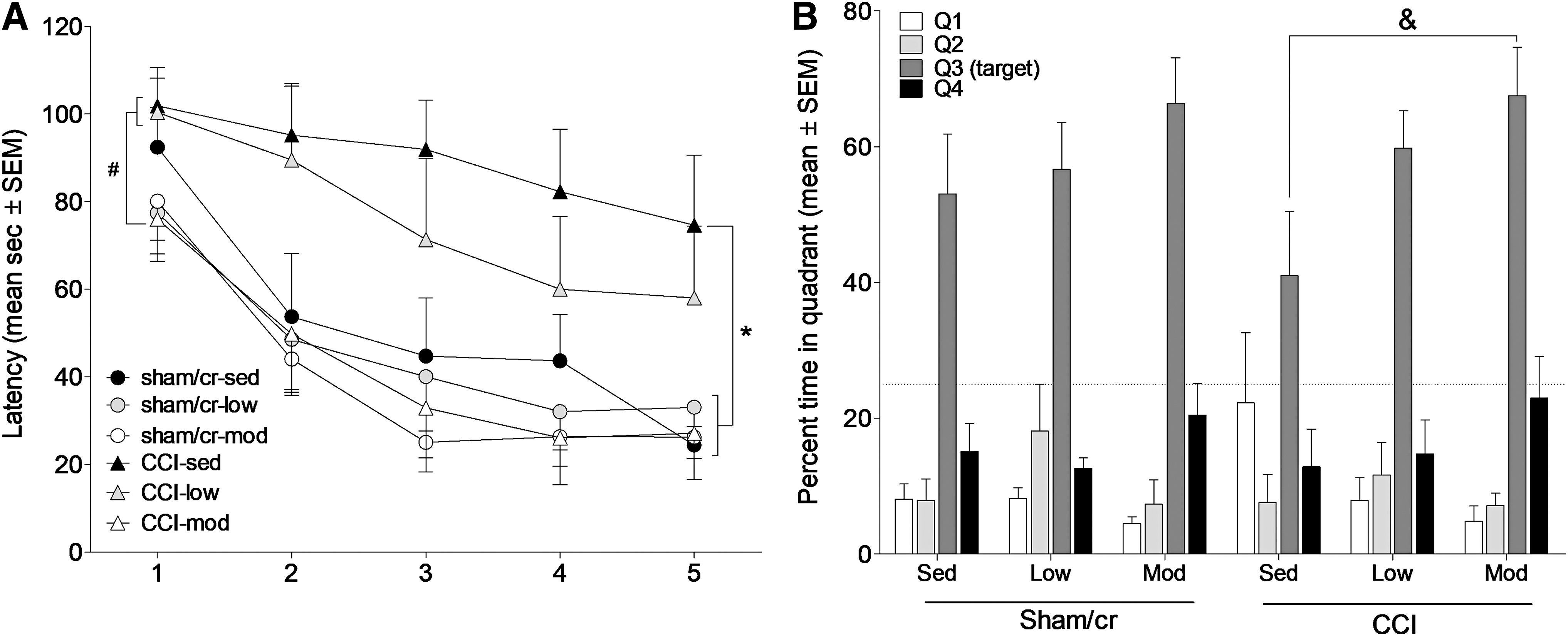
Treadmill exercise improves hippocampus-dependent learning and memory
Treadmill exercise does not exacerbate TBI-associated pathology
Mice were euthanized following behavioral analysis, and brain tissue was assessed for TBI-related pathology. Cortical lesion volume was significantly larger in CCI than in sham/cr mice (F 1,39 = 44.574, p < 0.001); however, there was no significant effect of exercise on lesion volume among CCI mice (p > 0.05; Fig. 4A and B). Similarly, axonal degeneration was significantly more severe in CCI than in sham/cr mice (U = 468.0, p < 0.001), but was not significantly affected by exercise among CCI mice (p > 0.05; Fig. 4C and D).
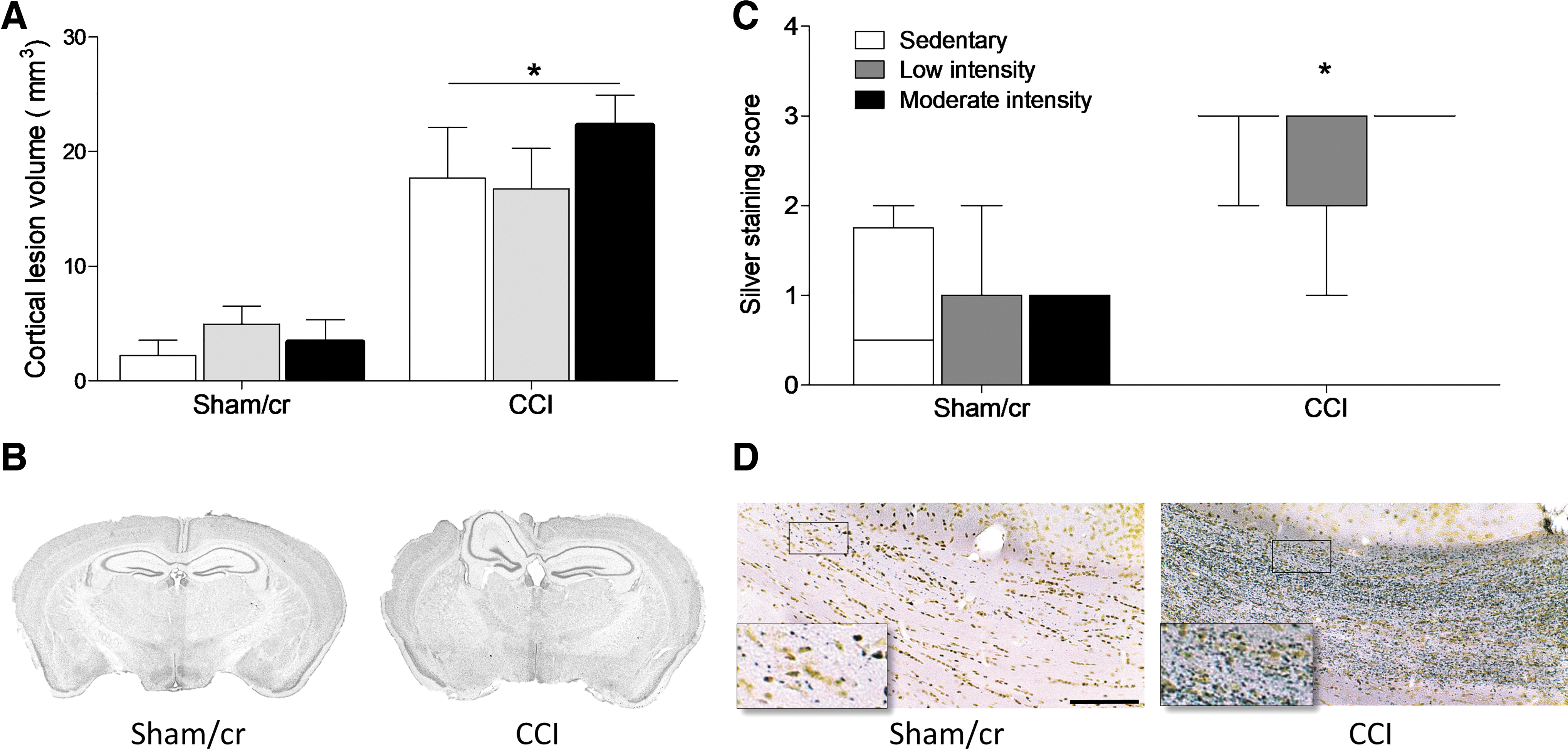
Lesion volume and axonal degeneration following controlled cortical impact (CCI).
Treadmill exercise attenuates microglial activation
Microglial activation was assessed via Iba1 immunohistochemistry in the peri-lesional cortex (and the corresponding cortical region on the contralateral hemisphere). A two-way ANOVA revealed a main effect of surgery such that CCI significantly reduced the percent of ramified microglia and correspondingly increased the percent of activated microglia in the ipsilateral (F 1,35 = 6.668, p < 0.05) but not contralateral (p > 0.05) cortex compared with sham/cr mice. A post-hoc comparison in the ipsilateral cortex revealed that whereas the CCI+sedentary group differed significantly from the sham/cr+sedentary group, neither the low nor moderate exercise CCI groups differed significantly from shams (Fig 5A and C). Similarly, in the CA1 region of the hippocampus, a two-way ANOVA revealed a main effect of surgery in the ipsilateral (F 1,35 = 8.544, p < 0.01) but not contralateral (p > 0.05) hippocampus. As in the cortex, CCI significantly decreased the percent of ramified (and correspondingly increased activated) microglial cells in the hippocampus CA1. A post-hoc comparison in the ipsilateral CA1 revealed that whereas the CCI+sedentary group differed significantly from the sham/cr+sedentary group, neither the low or moderate exercise CCI groups differed significantly from shams (Fig. 5B and C). There were no significant main effects of exercise intensity on either the cortex or hippocampus. Analysis of the hippocampus dentate gyrus revealed no significant main effects of either surgery or exercise intensity (all p < 0.05).
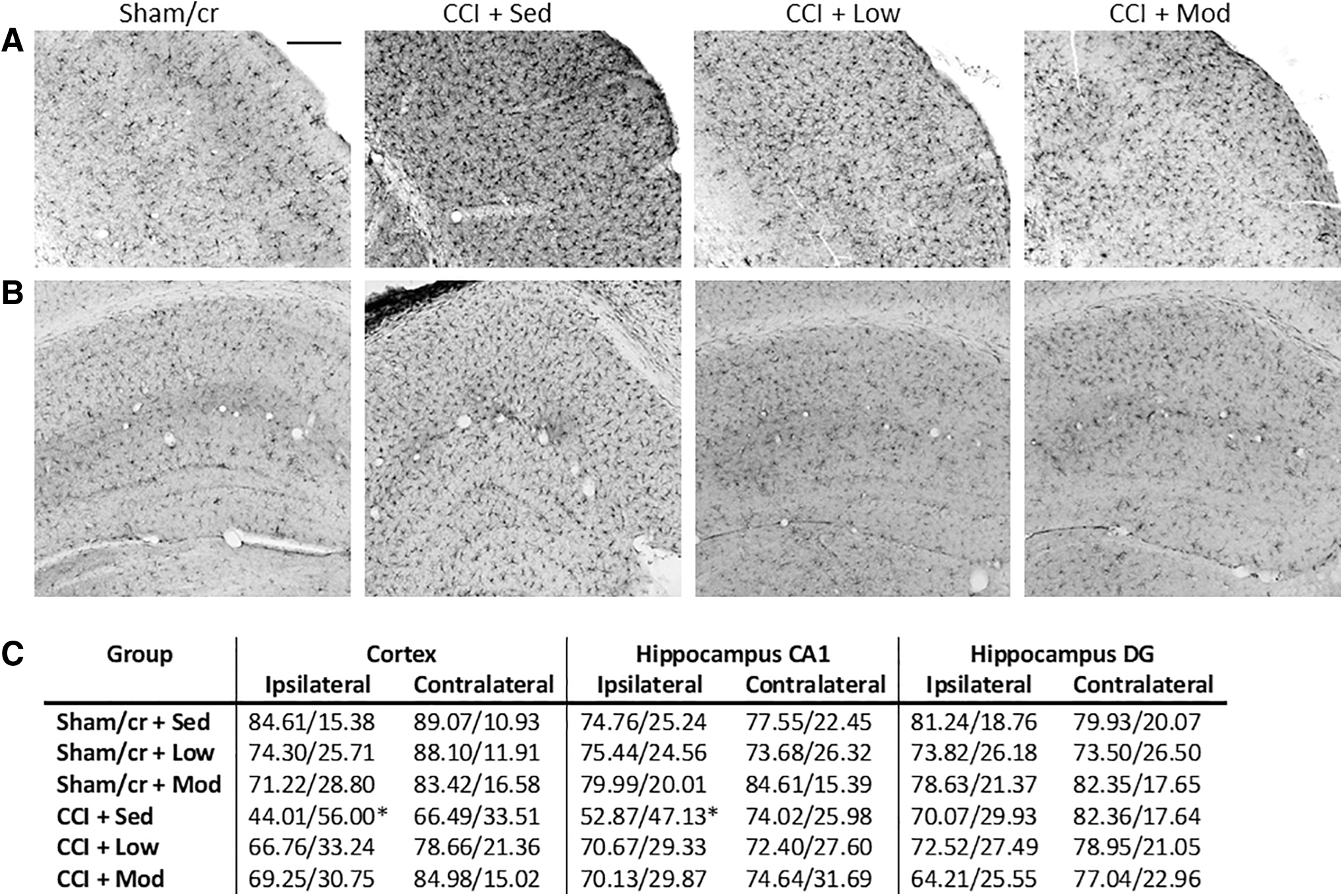
Microglial activation state following controlled cortical impact (CCI). Representative images of Iba-1 positive microglia in the
Treadmill exercise affects proliferation and survival of newborn neurons
Exercise improves cognitive function in part by increasing hippocampal neurogenesis and the survival of newborn neurons 40,41 ; however it remains unknown whether exercise intensity differentially affects these processes in the injured brain. In order to determine whether group differences in cognitive outcomes in our study were the result of greater rates of survival of newborn neurons following exercise, mice were injected with BrdU on days 1 and 2 after injury. Immediately following the completion of the exercise paradigm, mice were euthanized and tissue was collected for immunohistochemistry. Therefore, tissue collected from this cohort of mice (Fig. 1B) allowed us to examine the hippocampal profile at the same time point that the other cohort (Fig. 1A) began behavioral testing. In order to assess the effect of treadmill exercise on injury-induced hippocampal neurogenesis, mice in cohort 2 were injected with BrdU (a thymidine analog that is incorporated into the DNA of dividing cells during the S phase of the cell cycle) on days 1 and 2 following injury. Mice were euthanized after 10 days of treadmill exercise, and brain tissue was assessed for BrdU and DCX (immature neuronal marker) labeling in the hippocampus dentate gyrus. A two-way ANOVA revealed no significant effects of injury or exercise on ipsilateral dentate gyrus BrdU expression (all p > 0.05; Fig. 6A); however, among the CCI groups, there was a main effect of exercise in the contralateral hippocampus (F 2,18 = 5.091. p < 0.05).
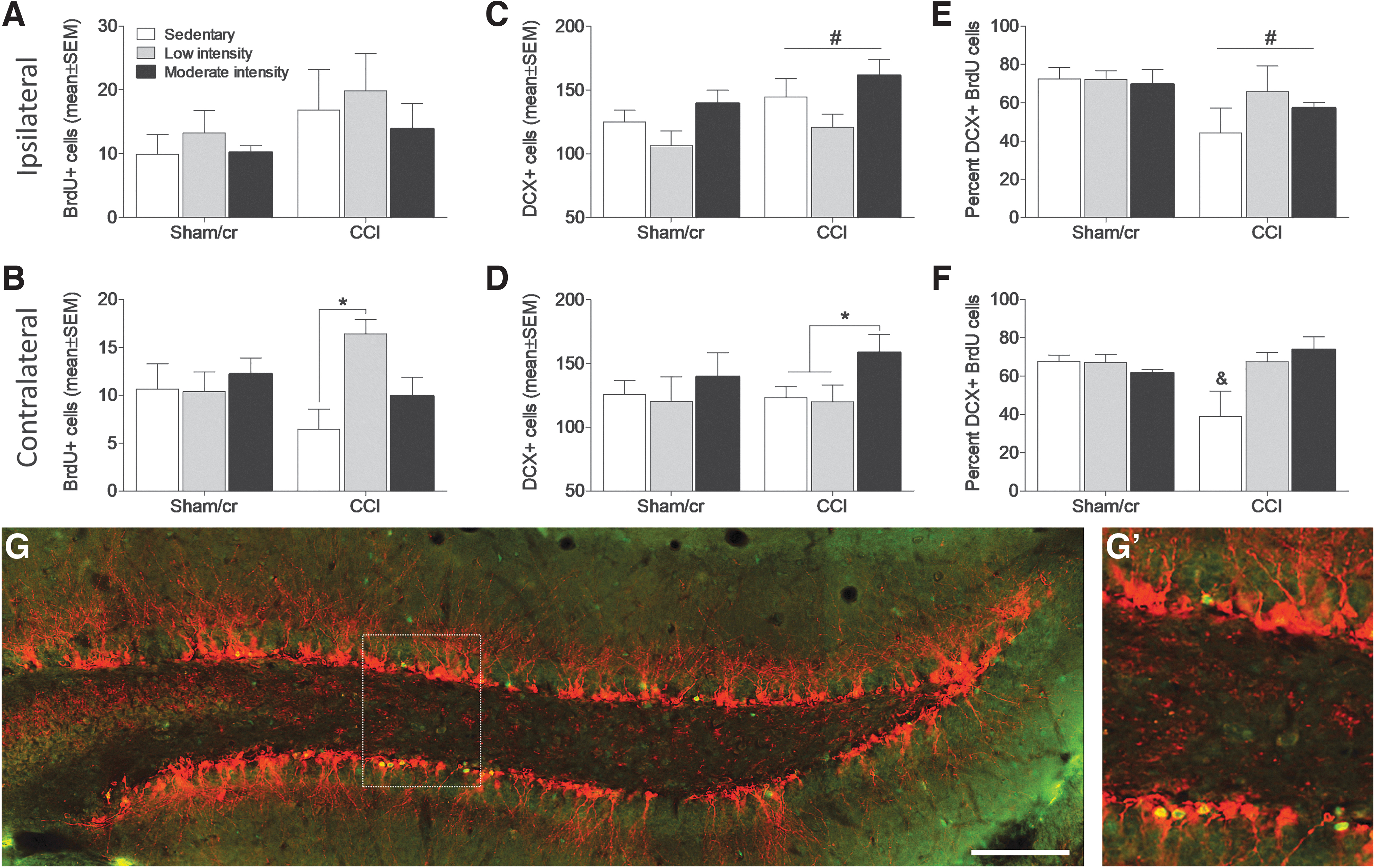
Treadmill exercise increases the number of surviving doublecortin (DCX)-positive neurons in the contralateral hippocampus after controlled cortical impact (CCI). Number (± standard error of the mean [SEM]) of bromodeoxyuridine / 5-bromo-2'-deoxyuridine (BrdU)-positive cells in the
A post-hoc analysis revealed that low intensity exercise significantly increased the number of BrdU cells in the contralateral hemisphere of CCI mice compared with sedentary conditions (p < 0.05; Fig. 6B). The total number of DCX-positive neurons was significantly increased by injury in the ipsilateral (F 1, 31 = 8.168, p < 0.01; Fig. 6C) but not the contralateral (Fig. 6D) hippocampus. However, there is a main effect of exercise among CCI mice (F 2,17 = 3.840, p < 0.05) on DCX-positive cells in the contralateral hemisphere. A post-hoc analysis revealed that moderate intensity exercise increased the number of DCX-positive cells compared with both sedentary (p < 0.05) and low intensity (p < 0.05) exercise conditions. A comparison of the percent of BrdU cells that co-label with DCX revealed that CCI significantly reduces the proportion of surviving newborn neurons (main effect of injury; F 1,31 = 5.346, p < 0.05) in the ipsilateral hippocampus (Fig. 6E). However, analysis of the contralateral hippocampus revealed both a main effect of exercise (F 2,34 = 3.356, p < 0.05) and an injury by exercise interaction (F 2,34 = 5.805, p < 0.01) such that the proportion of co-labeled BrdU and DCX cells was only reduced in the sedentary CCI group compared with all other groups (all p < 0.05, Fig. 6F).
Analysis of progenitor cell proliferation via Ki67 labeling revealed a similar pattern. Brain injury significantly reduced the number of Ki67-positive proliferating cells in the ipsilateral hippocampus (F 1,34 = 5.042, p < 0.05; Fig. 7A). However, there was a main effect of exercise in the contralateral hippocampus (F 2,34 = 3.767, p < 0.05), revealing a significant increase in progenitor cell proliferation in the moderate intensity exercise CCI group relative to sedentary CCI (post-hoc analysis, p < 0.05; Fig. 7B).
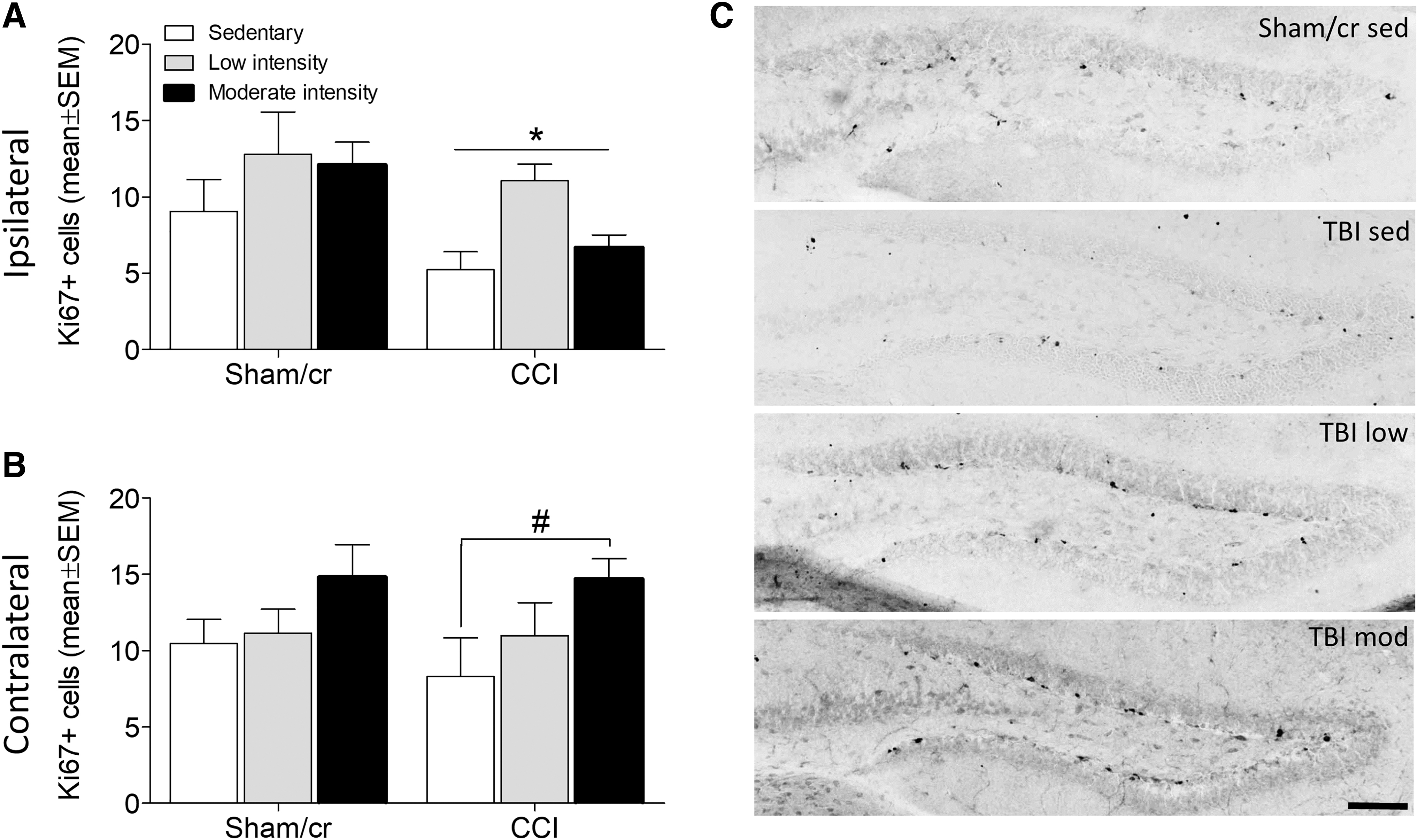
Treadmill exercise increases the number of proliferating progenitor cells in the contralateral hippocampus after controlled cortical impact (CCI). Number (± standard error of the mean [SEM]) of Ki67-positive progenitor cells in the
Discussion
Effect of treadmill exercise on functional recovery after brain injury
Standard treatment for brain injury patients generally includes some extent of physician-prescribed rest as a means to avoid repeated injury and reduce metabolic strain during a vulnerable period of post-injury recovery. 1,42 Given the highly heterogeneous nature of brain injuries (type of injury, affected brain regions, and severity), and the impact of variables such as patient age, genetics, and comorbidities, there is no standardized approach to determining the extent and duration of rest after injury. Rather, treatment guidelines rely on outward symptom resolution as an indicator that the patient can gradually return to pre-injury activities. 43,44 Unfortunately, self-reporting of symptom resolution is highly variable and may discount lasting cognitive, mood, and stress-related symptoms that may be exacerbated by restricted physical activity. 44,45 A growing body of evidence has begun to question the utility of post-injury rest, as recent randomized controlled trials and meta-analytic reviews have identified little to no benefit of rest to brain injury patients. 4,14,15,46,47 In contrast to the clinical advice regarding physical activity after TBI, animal models of brain injury indicate that under controlled conditions, exercise speeds up cognitive recovery, promotes neurogenesis, and attenuates neuroinflammation. 25,48 –51 Here, the goal was to address the effect of low and moderate intensity treadmill exercise on cognitive recovery and hippocampal plasticity after brain injury. Consistent with known neuroprotective effects of exercise, our study reveals that moderate intensity treadmill exercise reduces anxiety-like behavior and improves learning and memory to a greater extent than low intensity exercise after CCI. Moreover, we report an exercise intensity-dependent increase in hippocampal proliferation and survival of newborn neurons in the dentate gyrus.
The CCI model reproduces key clinical elements of brain injury, including pathophysiology, cognitive decline, anxiety, and depressive behaviors. 34,52 Post-injury neurobehavioral symptoms are experienced by as many as 80% of patients with mild to moderate TBI. 53 Further, symptoms such as impaired cognitive function (i.e., attention, executive function, memory), major depression, and anxiety/post-traumatic stress disorder may be exacerbated by somatic symptoms such as headache, dizziness, and sleep disturbance. 54 Research has consistently demonstrated that physical activity decreases symptoms of depression and anxiety in uninjured participants. 55 Among brain-injured patients, exercise increases vigor, reduces negative mood states, and improves cognitive recovery. 13,56 –58 Moreover, TBI patients whose rehabilitation includes physical exercise are more likely to self-report improved health and quality of life. 13,18,59 Similarly, in pre-clinical models of traumatic or ischemic brain injury, exercise improves spatial learning 23,33,48 and ameliorates depressive 60 and anxiety-like 61 behaviors.
Our data support these findings and suggest that titration of exercise intensity is key for improving functional recovery after TBI. Indeed, we report an exercise intensity-dependent dose response for both anxiety-like behavior and performance on a spatial memory task. However, there are notable discrepancies between our findings and similar work. In addition to exercise intensity, the efficacy of post-injury exercise is determined by several factors including the exercise paradigm (forced vs. voluntary), timing of onset, and injury severity. Numerous studies of post-injury exercise use voluntary wheel running rather than forced treadmill (or motorized wheel running) paradigms. Voluntary wheel running is reported to produce lower stress responses 21 and to have different effects on trophic factor upregulation and behavioral outcomes compared with forced exercise. 21,62 –64 However, neither paradigm has produced consistent results in acquired brain injury models (stroke or TBI). In the current study, forced treadmill exercise is preferred experimentally because of the greater control over both the duration and intensity of exercise. The more critical variable in determining efficacy of post-injury exercise appears to be the timing of the onset of exercise. Pre-clinical models vary significantly on the timing of exercise onset, initiating as early as the day of injury and as late as 4 months after injury. 63 Again, the outcomes are somewhat inconsistent, and whereas numerous studies suggest that early onset exercise (1–4 days after injury) promotes recovery, 20,65,66 others have shown better outcomes with delayed exercise onset. 25,48 In the current study, exercise onset occurred on day 3 after injury, which is consistent with the clinical recommendation for post-injury rest and allows for assessment of the impact of exercise within the clinical paradigm that is currently in place for brain injured patients. Finally, although similar studies have been conducted to identify optimal post-injury exercise intensity, in contrast to our findings, some studies indicate a greater benefit of low intensity exercise over moderate/high intensity exercise. This discrepancy may be the result of an earlier onset of exercise and variability in the definition of low versus moderate or high intensity exercise, as well as genetic variability in exercise tolerance (i.e., Swiss–Webster mice perform significantly better on exercise stress tests than the more commonly used C57BL/6 mice). 23,67 Overall, the findings in this study indicate that moderate exercise led to a substantial improvement on two of the most common and debilitating outcomes measures of brain injury: cognitive recovery and anxiety-like behavior.
Effect of treadmill exercise on TBI pathophysiology
The pathophysiology underlying TBI-induced cognitive deficits is multi-faceted, and includes neuroinflammation, excitotoxicity, and metabolic dysfunction, which contribute to neuronal degeneration of vulnerable hippocampal neurons. 68,69 Pathophysiological assessments in our study indicate that neither low nor moderate intensity exercise exacerbated cortical lesion volume or axon damage in brain injured mice. Similar studies using greater duration exercise paradigms have shown that exercise can reduce neuronal cell death and ultimately reduce the injury volume. 25,29,51,70 Although the exercise duration in the current study was insufficient to reduce these pathophysiological measures, it remains critically important to note that even a brief duration of exercise initiated following brain injury was sufficient to promote functional recovery without exacerbating neurodegeneration. Moreover, our data indicate that microglial activation state is attenuated by both exercise intensities resulting in proportionally greater numbers of resting (ramified) and fewer activated microglia in both the peri-lesional cortex and hippocampus in injured mice. Microglia play a critical role as a first line of defense in brain injury and are therefore rapidly upregulated in the injured brain. Although critical for the initial response to injury (metabolic product clearance, debris removal, synaptic plasticity), chronic upregulation of microglia is pro-inflammatory and neurotoxic. 71 Our data are consistent with known anti-inflammatory effects of exercise in acquired brain injury models. 25,66,72 Specifically, exercise has been shown to reduce the number of CD68-positive reactive microglia, inhibit microglia-associated pro-inflammatory cytokine production, and upregulate expression of genes that promote activation of the anti-inflammatory M2 microglial phenotype. 25,72 Importantly, the exercise paradigm used in this study significantly improved functional recovery and attenuated microglial activation in brain injured mice without exacerbating tissue damage.
Effect of post-injury treadmill exercise on proliferation and survival of newborn neurons
Cognitive impairment after TBI is believed to be driven by a combination of primary neuronal and vascular injury, as well as secondary complications including edema, neuroinflammation, and neurodegenerative mechanisms that contribute to cell death. 73 Neurogenic regions in the brain (i.e., SGZ) function as an endogenous tool of regeneration and repair, as they are highly sensitive to environmental changes caused by brain injury, and therefore rapidly respond by increasing progenitor cell proliferation. 74,75 Increasing evidence suggests that physical exercise enhances neural stem cell proliferation and neurogenesis 41,76 and promotes cognitive recovery after brain injury. 65,66,77 Given that DCX-positive newborn neurons are highly vulnerable to brain injury, 78,79 we tested whether exercise promotes newborn neuronal survival after CCI. Although our data indicate a reduction of BrdU-positive cells that express DCX in the ipsilateral hippocampus, both low and moderate intensity exercise increased the survival of newborn hippocampal neurons in the contralateral hemisphere. Brain injury typically results in an acute period of increased proliferation during the first few days of recovery, 75 and whereas proliferation typically drops off significantly after this acute period, we report that exercise dose-dependently maintains an increased proliferation rate of Ki67-positive cells in the contralateral hippocampus. Higher rates of proliferation and newborn neuron survival in the contralateral hippocampus likely reflect substantial differences in the microenvironment between the ipsilateral and contralateral hemispheres. CCI, although focused to one side of the brain in this study, can affect the contralateral side via direct damage through mechanical disruption that propagates through the tissue, death of ipsilateral cell bodies that target the contralateral side, and damage to axons that originate in the contralateral hemisphere and project to the injured zone. 80,81 Further, areas of focal injury can alter more remote cell populations via secreted factors, spreading depolarizations and a variety of other mechanisms. Newborn neurons are vulnerable to factors such as neuroinflammation, 82 oxidative stress, 83 and mitochondrial dysfunction, 84 which occur throughout the central nervous system (CNS) in this injury model, but to a much greater extent within the injured hemisphere. Exercise likely has two separate but overlapping classes of effects on neurogenesis in this paradigm. First, exercise has been shown repeatedly to drive cell proliferation and neurogenesis via the induction of growth factors (i.e., vascular endothelial growth factor and brain derived neurotrophic factor), 85,86 and activation of the cyclic adenosine monophosphate (AMP) response element-binding protein transcription factor, 87 among others. Second, exercise can likely help reduce some of the pathophysiological consequences of injury that would otherwise serve to inhibit neurogenesis. 29,51,72 As the two hemispheres both experience the exercise but are differentially affected by the CCI, it is not surprising that the two sides of the brain exhibit divergent neurogenic responses. Similar patterns of hemisphere differences in both proliferating cells and immature neuron numbers have been reported after brain injury 75 ; however, this is the first report to show that forced treadmill exercise promotes neuronal survival in the contralateral, but not ipsilateral, hemisphere. Overall, these data are consistent with known effects of exercise as a promoter of hippocampus-dependent cognitive recovery in part by increasing the survival of newborn neurons in the dentate gyrus. 40 Given that the time point at which these data were assessed corresponds to the start of behavioral testing in the first cohort of this study, these experiments suggest that exercise ameliorates cognitive deficits in brain injured mice, in part by increasing hippocampal proliferation and newborn neuronal survival.
Notable discrepancies exist between our results and previously published reports. We were unable to replicate the well-established findings of increased BrdU labeling in CCI mice, which is likely a reflection of differences in BrdU injection timing and duration. 88,89 The BrdU dosing schedule for the current study was selected in order to mark cells born after the injury, but prior to the start of the treadmill protocol. This dosing regimen allowed us to specifically assess the impact of treadmill exercise on newborn neuronal survival of the small subset of tagged cells, rather than getting an overall assessment of neurogenesis. We also note that variability in total numbers of BrdU-positive cells may also be related to age differences, as mice were injured between 4 and 6 weeks of age. This age range represents considerable variability in neurogenesis, 90 and although litters were counterbalanced in each group (therefore the age range is equally represented among all groups assessed), the difference in age likely contributed to variability in our data set.
Conclusion
Taken together, the data sets presented here indicate that moderate intensity forced exercise reduces anxiety-like behavior, improves cognitive function, and increases hippocampal proliferation and newborn neuronal survival following CCI. Critically, the benefits of exercise were achieved without exacerbating the lesion volume or axon damage, which suggests that carefully titrated physical exercise may be an effective approach for promoting recovery in TBI patients and may pose less risk to the injured brain than initially predicted. Ultimately, the outcomes of this study contribute further insight and empirical evidence of the impact of post-injury exercise that can better inform future recommendations of rest and exercise in the TBI patient population.
Footnotes
Funding Information
Research reported in this publication was supported by the National Institute of General Medical Sciences of the National Institutes of Health under award number U54GM104942, and the National Institute of Neurological Disorders and Stroke of the National Institutes of Health under award number P30NS104177.
Author Disclosure Statement
No competing financial interests exist.