
Abstract
Traumatic brain injury (TBI) alters microbial populations present in the gut, which may impact healing and tissue recovery. However, the duration and impact of these changes on outcome from TBI are unknown. Short-chain fatty acids (SCFAs), produced by bacterial fermentation of dietary fiber, are important signaling molecules in the microbiota gut–brain axis. We hypothesized that TBI would lead to a sustained reduction in SCFA producing bacteria, fecal SCFAs concentration, and administration of soluble SCFAs would improve functional outcome after TBI. Adult mice (n = 10) had the controlled cortical impact (CCI) model of TBI performed (6 m/sec, 2-mm depth, 50-msec dwell). Stool samples were collected serially until 28 days after CCI and analyzed for SCFA concentration by high-performance liquid chromatography-mass spectrometry/mass spectrometry and microbiome analyzed by 16S gene sequencing. In a separate experiment, mice (n = 10/group) were randomized 2 weeks before CCI to standard drinking water or water supplemented with the SCFAs acetate (67.5 mM), propionate (25.9 mM), and butyrate (40 mM). Morris water maze performance was assessed on post-injury Days 14–19. Alpha diversity remained stable until 72 h, at which point a decline in diversity was observed without recovery out to 28 days. The taxonomic composition of post-TBI fecal samples demonstrated depletion of bacteria from Lachnospiraceae, Ruminococcaceae, and Bacteroidaceae families, and enrichment of bacteria from the Verrucomicrobiaceae family. Analysis from paired fecal samples revealed a reduction in total SCFAs at 24 h and 28 days after TBI. Acetate, the most abundant SCFA detected in the fecal samples, was reduced at 7 days and 28 days after TBI. SCFA administration improved spatial learning after TBI versus standard drinking water. In conclusion, TBI is associated with reduced richness and diversity of commensal microbiota in the gut and a reduction in SCFAs detected in stool. Supplementation of soluble SCFAs improves spatial learning after TBI.
Introduction
The gut microbiota are severely disrupted by trauma and critical illness. 1 –4 Studies utilizing high-throughput molecular techniques to identify disturbances in the microbiota of adult and pediatric intensive care unit (ICU) patients have observed dysbiosis characterized by reduced bacterial diversity, depletion of commensal bacterial species, enrichment with nosocomial pathogens, and loss of body site specificity. This state can be characterized as “ICU dysbiosis.” 1,2 Once the microbial community has shifted, dysbiosis may persist in a stable maladaptive state with detrimental consequences for the host, 5 –7 including diarrhea, secondary infections, and chronic inflammation.
Studies of adults years after moderate–severe traumatic brain injury (TBI) 8 suggest that gut microbiome may be chronically disrupted. In vivo models of TBI in mice and rats have reported acute changes to gut microbiota occurring within the first week of injury. 3,4 The relevance of these changes to the gut microbiome on secondary brain injury or recovery from TBI has not been examined in either the pre-clinical or clinical setting.
The microbiome gut–brain axis describes bidirectional biochemical signaling between the brain and microorganisms in the gut. Microbiota-generated metabolites represent one of the primary pathways of communication between the microbiome and host and influence normal brain function and response to injury. 9 –11 Short-chain fatty acids (SCFAs) are a major class of bacterial metabolites produced by bacterial fermentation of undigested dietary fiber in the cecum and colon. SCFA signaling is relevant to blood–brain barrier (BBB) permeability, microglial polarization and function, and neurogenesis. 12 A consistent finding in critically ill humans 1,2 and corresponding murine models 3,13 is the depletion of SCFA-producing commensal bacteria. We hypothesized that following TBI, we would observe progressive depletion of SCFA-producing commensal bacteria in the gut and generation of SCFAs. Further, we hypothesized that SCFA supplementation would be neuroprotective after TBI. Our results demonstrate that SCFA-producing commensal bacteria from the Ruminococcaceae, Bacteroidaceae, and Lachnospiraceae families are progressively depleted following TBI and do not recover out to 28 days after injury. Supplementation of SCFAs in drinking water improved spatial learning in the TBI model.
Methods
Animals and surgical procedures
Studies were approved by the Institutional Animal Care and Use Committee at the University of Pittsburgh. Adult male C57BL6/J mice were purchased from Jackson Laboratories (Bar Harbor, ME), single housed with access to standard rodent chow (Prolab Isopro RMH 3000, LabDiet, St. Louis, MO) and autoclaved water ad libitum for at least 1 week prior to experimental protocol.
TBI was induced using the controlled cortical impact (CCI) model as previously described. 14,15 Briefly, mice were anesthetized with 4% isoflurane followed by continuous administration of 2% isoflurane in N2O/O2 (2:1) via nosecone and placed in a modified stereotaxic frame with ear bars to stabilize the head. A craniotomy was performed in the left parietal bone and the bone flap removed. CCI was performed with a 3-mm impactor tip fired at velocity 6 m/sec, depth 2 mm, dwell 50 msec. The skull was replaced and secured with dental cement (Koldmount, Vernon-Benshoff, Albany, NY), the scalp closed, 10% povidone iodine solution (McKesson, Irving, TX) topical applied to the scalp, and mice recovered in a temperature-controlled chamber. No systemic antibiotics were administered in the performance of the CCI model.
Experimental rigor
Mice were ordered to arrive to the housing facility at 11 weeks of age and were mixed prior to the start of the experiment. After at least 1 week, mice were moved to individual housing to prevent coprophagic effects and, in the behavioral outcome study, to prevent benefit of social enrichment. Prior to the outcomes study, mice were randomized to experimental groups and assigned a study identification number. Cages were labeled only with the mouse identification number and technicians performing the surgery, behavioral outcomes assessment, histologic analysis, SCFA analysis, and fecal collection were blinded to experimental groups. Diet and food autoclaving procedures were standardized. Addition of SCFAs to drinking water did not change the clarity of the water; however, an odor is present in SCFA supplemented water if investigators were very close to the cage.
SCFA supplementation in drinking water
Mice were randomized to receive SCFA-supplemented or standard drinking water 2 weeks prior to CCI and continued until sacrifice. For SCFA treatment, 67.5 mM sodium acetate (Sigma-Aldrich, St. Louis, MO), 25.9 mM propionate (Sigma-Aldrich), and 40 mM butyrate (Sigma-Aldrich) were added as previously described. 16,17 Water solutions were prepared and changed three times per week.
DNA extraction and analysis of 16S rRNA genes
Stool was collected from mice (n = 10) at baseline, 3 h, 24 h, 3 days, 7 days, 14 days, and 28 days after CCI. Mice were placed in a sterile chamber and fresh fecal pellets collected and stored at −80°C prior to batch analysis. The MO BIO PowerSoil DNA Isolation Kit (MO BIO Laboratories, Carlsbad, CA) was used to extract microbial DNA according to manufacturer instructions. 16S rRNA gene amplicons were produced by polymerase chain reaction amplification of V4 amplicon using 515F and 806R primers. Samples were forward and reverse sequenced on the Illumina MiSeq platform (San Diego, CA). QIIME2 was used to demultiplex raw sequence read pairs, trim reads, and the DADA2 algorithm implemented within QIIME was used to denoise reads and generate a table of operational taxonomic units (OTUs). Taxa were assigned to predicted OTUs with feature classifier plugin trained on the greengenes reference database (v13.8). Alpha and beta-diversity were calculated using Phyloseq and analysis of differentially enriched or depleted OTUs was performed with LEfSe 18 with correction for multiple comparisons.
SCFA quantification
Sample preparation
Stool samples were homogenized with 50% aqueous acetonitrile containing 10μg/mL internal standard (butyrate-d5) at a ratio of 100 mg stool to 1 mL of solution followed by vortex mixing and centrifugation at 16,000 × g for 10 min at 10°C; 60μL cleared supernatants were transferred to a clean tube and derivatized with 30 μL of 200 mM 3-nitrophenylhydrazinein 50% aqueous acetonitrile and 30 μL of 120 mM N-(3-dimethylaminopropyl)-N0-ethylcarbodiimide -6% pyridine solution in 50% aqueous acetonitrile. The mixture was reacted at 60°C for 30 min and the reaction was stopped with 0.45 mL of 50% acetonitrile.
Liquid chromatography–mass spectrometry analysis
Derivatized samples were injected (5 μL) into a Thermo Vanquish UHPLC at a flow rate of 0.25 mL/min using a binary solvent system. Solvent A consisted of water and 0.1% formic acid and solvent B consisted of acetonitrile and 0.1% formic acid. Short-chain fatty acids were separated using binary solvent elution gradient at 15% B for 2 min, 15–55% B in 9 min, and then held at 100% B for 1 min. The column was equilibrated for 3 min at 15% B between injections. SCFA were separated on a Phenomenex Kinetex C18 (2 × 150 mm, 1.7 μ) column at 40°C. SCFA were analyzed using a Thermo Q-Exactive mass spectrometer at 60,000 resolution in negative ion mode, using a data-dependent Top-5 ms2 method. The duration between collection and SCFA analysis was standardized across fecal samples.
Functional outcome
Mice (n = 10/group: naïve, naïve+SCFA, CCI, CCI+SCFA) were assessed for spatial learning and memory acquisition using the Morris water maze (MWM) as previously described. 19,20 Naïve mice were not subjected to anesthesia or sham surgery. MWM employs a 90-cm diameter pool with a 10-cm platform placed 1 cm below the water's surface to assess the mouse's ability to learn spatial relations between distal cues and escape to the platform. On Days 14–18 after CCI, the average of four trials taken from set starting positions in the pool was recorded for each mouse. On Day 19, mice were assessed for their ability to escape to a visible platform and probe trial was performed where the platform was removed and mice swimming behavior assessed for 30 sec.
Lesion volume
On Day 28 after CCI, mice (n = 10/group) were anesthetized with 4% isoflurane and sacrificed via transcardial perfusion with heparinized saline followed by 2% paraformaldehyde. The brain was dissected, cryopreserved in sucrose solution, and stored at −80°C. Coronal 10-μm frozen sections were stained with hematoxylin and eosin and lesion volume quantified using image analysis software (MCID; Imaging Research, Saint Catherine's, Ontario, Canada) by an evaluator blinded to the treatment groups (OMO). 19
Statistical analysis
Statistical analysis was performed using Prism 8 (Graphpad, La Jolla, CA). SCFA concentrations were compared by repeated measures (RM)-analysis of variance (ANOVA). When the overall ANOVA revealed a significant effect, the data were further analyzed to determine specific group differences with Dunnett's test corrected for multiple comparisons. MWM hidden platform data were analyzed by two-way RM-ANOVA. All other data were analyzed by Mann-Whitney test. All data are presented as mean ± standard error of the mean. A p value <0 .05 was considered significant.
Results
Durable alterations in the gut microbiome in mice after TBI
Microbiome analysis of fecal pellets from mice (n = 10) was performed at baseline and serially following TBI (Fig. 1). Alpha diversity, as determined by species richness (observed OTU and Shannon Index, remained stable for 72 h after TBI followed by a progressive decline in diversity without recovery at 28 days after injury. Beta diversity was analyzed by principal component analysis (PCoA) plot of abundance Jaccard distances, based on the presence or absence of bacterial families and abundance. The maximum variation was across principal component 1 and, overall, the samples obtained on Day 28 after TBI were quite distant from earlier samples. Beta diversity distances between sampling times were significant when all times were included (p < 0.001; R2 = 0.26768). We found no significant differences in group heterogeneity (PERMDISP p > 0.05), indicating that dispersions in groups did not account for this difference. Multiple pairwise ADONIS2 tests of all days revealed that differences between all days and Day 28 were significant (p < 0.05), while other comparisons did not reach significance (Supplementary Table S1).
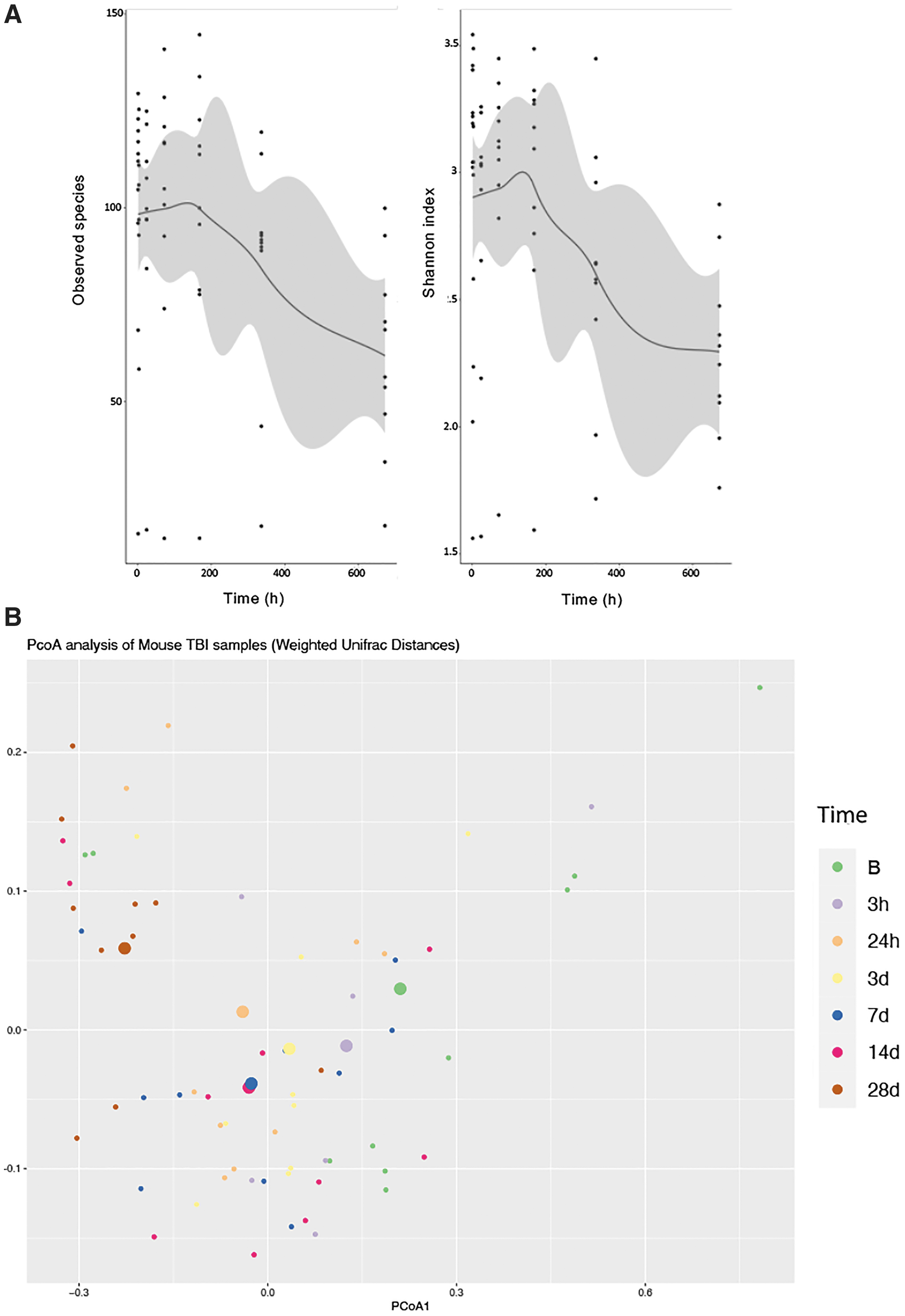
Changes in gut microbiome over time after traumatic brain injury (TBI). Adult C57BL6/J male mice (n = 10) underwent controlled cortical impact (CCI) performed and serial fecal pellets were analyzed by 16S rRNA sequencing.
Taxonomic composition of samples was performed and LefSE was used to determine enriched or depleted taxa (Fig. 2). Five taxa were observed to be significantly different (p < 0.05). Bacteria from the Verrucomicrobiaceae and Erysipelotrichaceae families were enriched over time, whereas bacteria from the Lachnospiraceae, Ruminococcaceae, and Bacteroidaceae families were depleted over time.

Taxonomic analysis of gut microbiome over time after traumatic brain injury
Total fecal SCFA concentrations are depleted after TBI
The same fecal samples used for 16S rRNA microbiome analysis also were used for SCFA quantification (Fig. 3). In Figure 3A, the relative abundance of acetate (C2), propionate (C3), and butyrate (C4) for individual mice are plotted from baseline out to 28 days after TBI. Absolute quantification of SCFAs was performed and normalized to fecal weight. Acetate concentration, the most abundant SCFA detected in the stool, was decreased from baseline at 7 days (7.1 ± 1.1 vs. 10.5 ± 1.2; p < 0.05) and 28 days (4.1 ± 0.4 vs. 10.5 ± 1.2; p < 0.0001) after TBI. Total SCFAs were decreased from baseline at 24 h (9.5 ± 0.6 vs. 13.6 ± 1.7; p < 0.05), possibly due to reduction in rodent chow consumption post-procedure, and 28 days (5.9 ± 0.6 vs. 13.6 ± 1.7; p < 0.001) after TBI. No changes were observed in fecal concentrations of butyrate and propionate.
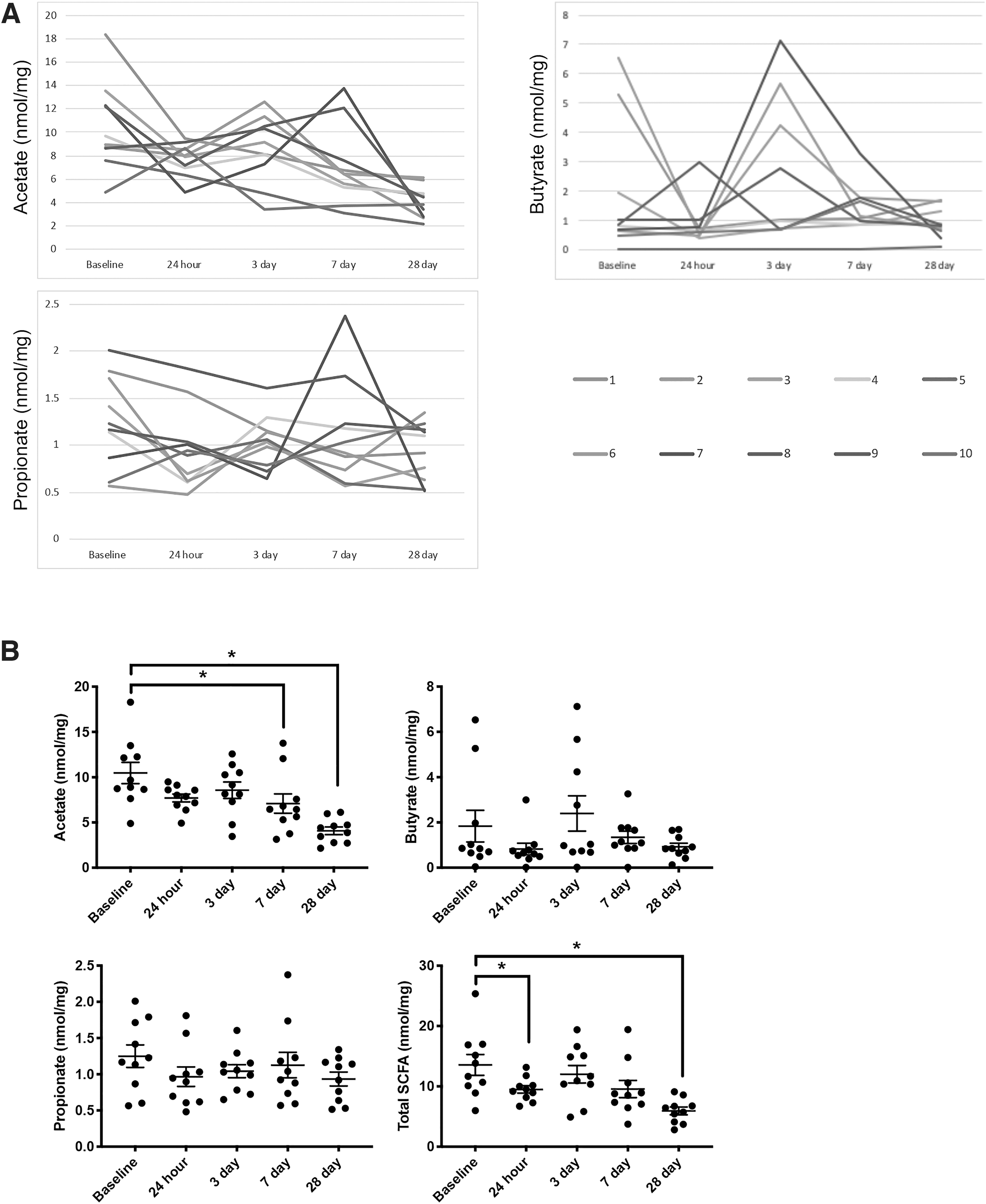
Changes in fecal short-chain fatty acid concentration over time after traumatic brain injury. Adult C57BL6/J male mice (n = 10) underwent CCI and serial fecal pellets were analyzed for short-chain fatty acid (SCFA) concentration.
In an exploratory analysis, samples that had both 16S rRNA gene sequencing and SCFA quantification performed were used to assess bacterial community profiles associated with SCFA concentration. Figure 4A shows a PCoA plot of community composition shaded for fecal acetate concentration. Axis 1 explained 74% of the variation between samples and appeared to correlate with acetate concentration. We also noticed a distinct “horseshoe” pattern in the ordination plot, another indication that most of the variation in the data was explained by the first coordinates. A linear regression analysis of SCFA concentrations versus the primary coordinates found that a correlation between Axis 1 and acetate (R 2 = 0.325, p < 0.001) and butyrate (R 2 = 0.300, p < 0.001), but not propionate. To determine differences in the gut microbiome between samples with “high” or “low” fecal acetate, we binned the samples into four groups of approximately equal size (n = 12–13 samples/group) according to acetate level (Supplementary Table S2). Group 1 was comprised of samples with acetate 2.16–4.88 nmol/mg, Group 2 comprised samples 4.88–7.31 nmol/mg, Group 3 comprised samples 7.31–9.16 nmol/mg, and Group 4 comprised samples 9.16–18.3 nmol/mg. Pairwise ADONIS comparisons of groups found significant differences between Group 1 and Groups 2, 3, and 4 (p < 0.01). Analysis of bacterial taxa enriched or depleted between these groups found a relative reduction in the Ruminococcaceae and Christensenellaceae families, which are important producers of acetate, 12,21,22 in Group 1 samples (Fig. 4B).
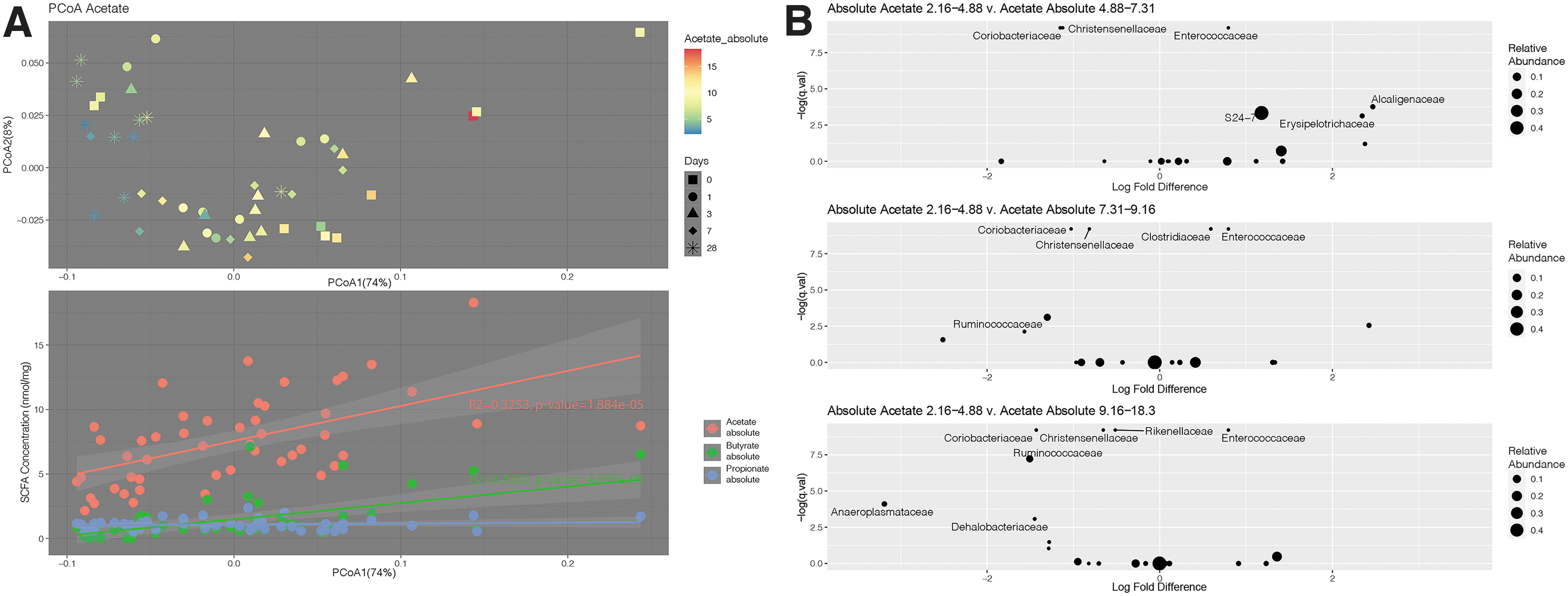
Community composition is distinct in samples with the lowest acetate concentration.
Supplementation of SCFAs via the drinking water improves spatial learning after TBI
Mice (n = 10/group) were randomized 2 weeks prior to CCI or no injury (naïve) to receive standard water or water supplemented with acetate, propionate, and butyrate. Spatial learning and memory acquisition was tested with MWM at 14–18 days post-injury (Fig. 5). No differences were observed between naïve+vehicle and naïve+SCFA mice. Compared with CCI+vehicle mice, CCI+SCFA mice had reduced latency to find the hidden platform (p < 0.05). Due to a difference in swim speed between naïve mice and mice after CCI, the mean path length over four trials was assessed. Compared with CCI+vehicle mice, CCI+SCFA mice had reduced path length to find the hidden platform (p < 0.05). On probe trial, the percentage time spent in target quadrant was assessed. As expected, CCI+vehicle mice spent less time in the target quadrant compared with Naïve+vehicle mice (17.5 ± 3.7% vs. 34.3 ± 5.6%; p < 0.05). No difference was observed on probe trial between naïve mice with or without SCFA treatment and CCI+SCFA mice.
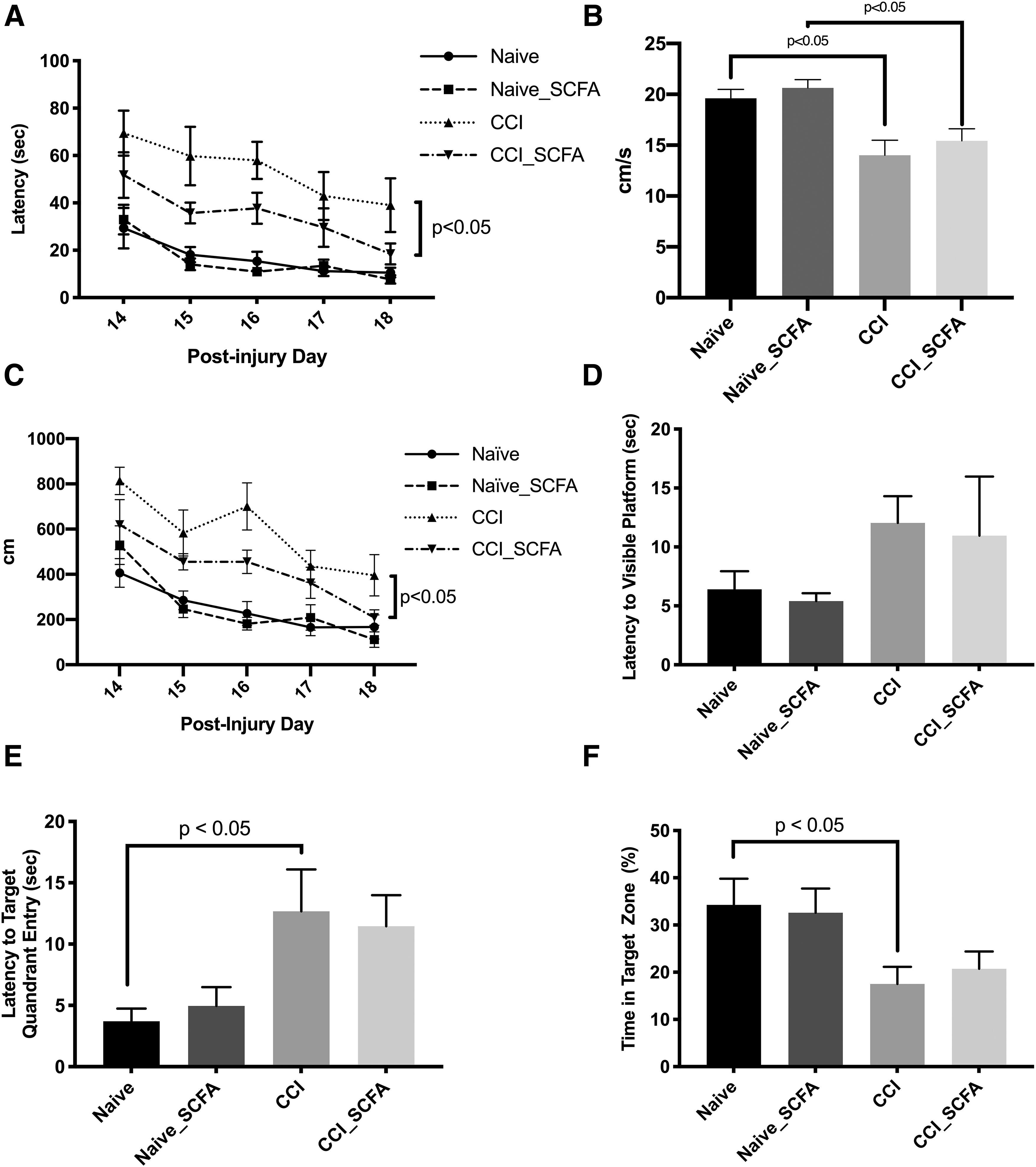
Mice receiving short-chain fatty acids (SCFAs) in drinking water after TBI demonstrate improved spatial memory acquisition. Adult male C57BL6/J mice (n = 10/group) were evaluated using Morris water maze.
SCFAs do not impact lesion volume after TBI
On post-injury Day 28, mice were sacrificed and the brain assessed for lesion volume. No significant difference was observed between CCI and CCI+SCFAs groups on lesion volume (20.4 ± 1.3 vs. 23.4 ± 1.4 mm3; p = ns) or hemispheric volume loss (34.5 ± 1.4 vs. 34.8 ± 1.9 mm3; p = ns; Fig. 6).

Histologic assessment of mice after traumatic brain injury with and without short-chain fatty acids (SCFAs) in drinking water.
Discussion
We report that microbial diversity is progressively decreased in the gut after TBI. We observed this change beginning approximately 72 h after injury with continued loss of diversity out to 28 days after injury. Bacteria from the Veruccomicrobiaceae and Erysipelotrichaceae families were enriched while bacteria from the Lachnospiraceae, Ruminococcaceae, and Bacteroidaceae families were depleted. The Lachnospiraceae, Ruminococcaceae, and Bacteroidaceae families include major SCFA producers in humans and mice. 12,23 Changes in gut microbiota and SCFA levels were analyzed in comparison to baseline samples after an acclimatization period. However, delayed effects of housing on low abundance OTUs cannot be excluded. 24 In contrast to our hypothesis of a steady and progressive decline in SCFA generation after TBI, we observed a significant decrease in total SCFAs in fecal samples at 24 h and 28 days after TBI driven largely by changes in acetate. Supplementing drinking water with the SCFAs acetate, propionate, and butyrate improved spatial learning after TBI without a difference in lesion volume.
Dysbiosis is defined as any change in the composition of commensal microbial communities relative to that found in healthy individuals. 25 The etiology of dysbiosis in critically-ill patients is likely multi-factorial and related to exposures such as antibiotics, sedation, invasive mechanical ventilation and vascular catheters. 26 Delays in initiating enteral nutrition or dietary changes from pre-morbid state could also play a role. 22 Additionally, in patients with severe TBI, the injury itself may impact the gut microbiome through vagal pathways and release of catecholamines which contribute to ileus, altered blood flow to abdominal viscera, and altered architecture of the intestinal epithelium. 13,27 In some cases, once these triggers of dysbiosis are removed, the microbiome may recover to a healthy state that closely resembles the community structure and function prior to critical illness. However, a dysbiotic microbiome may itself become stable and potentially with adverse effects on the host. 28
Urban and colleagues 8 compared the microbiome of patients with a history of moderate/severe TBI residing in permanent care facilities with age-matched controls working within the facilities and observed differences between the community structure of TBI subjects and controls. Similar to our findings, despite species differences, the authors reported a decrease in relative abundance of Bacteroidaceae spp. and an increased abundance of Verrucomicrobiaceae spp. Notably, the average time from injury was 260.0 ± 157 months. In our study, although we observed variability between individual mice, the reductions in bacterial diversity and changes in taxonomic composition did not recover 28 days after TBI despite returning the mice to the housing conditions and pre-injury diet. These findings suggest that in some patients, additional interventions may be required to rescue a stably dysbiotic microbiome. Treatments such as prescribed dietary postbiotics, functional bioactive compounds that boost the production of SCFAs by the microbiota, 29 may even be precision targeted 30 to promote the growth of potentially beneficial species. 31
Sustained impaired gut homeostasis characterized as depletion of commensal bacteria, as well as impaired gastric motility and barriers to administration of enteral nutrition in patients with critical illness and trauma, would be expected to limit the enteric and systemic availability of SCFAs. 32 The limited available evidence in patients is consistent with this hypothesis. Romick-Rosendale and colleagues 33 observed a significant reduction in fecal SCFA levels in children after hematopoietic stem cell transplant, a finding exacerbated in children receiving antibiotics with activity against anaerobic organisms. Yamashiro and colleagues 34 reported significantly lower total SCFA levels in the stool of adults with ischemic stroke.
In our study, the reduction in total SCFA concentration at 24 h after TBI, driven largely by changes in acetate, without changes detected in the gut microbiome may have been related to decreased oral intake in mice after CCI. When fermentable fibers are reduced in the diet, gut microbiota instead switch to using dietary or endogenous proteins or dietary fat as an energy source, with less SCFA production as a result. 35 In our experience, mice typically have decreased oral intake over the first several hours after CCI, but prolonged feeding intolerance or refusal is atypical. At 28 days after injury, we suspect the reduction in acetate and total SCFA concentration is due to the changes observed to gut microbiota. At this point, food consumption is indistinguishable between injured and naïve mice. Animal well-being and weight were monitored throughout the study; however, water consumption, which can be impacted by water supplements, was not directly assessed.
In an analysis of community profiles associated with high or low SCFA levels, we observed that samples with the lowest concentration of acetate were distinct from those with higher acetate concentration. In an analysis of individual taxa, we found that samples with the lowest acetate concentration were relatively depleted of bacteria from Ruminococcaceae and Christensenellaceae families. These bacterial families are important sources of acetate, as well as other SCFAs, in mice and humans. 12,21,22
Prior investigations support a neuroprotective role for SCFAs in TBI. Intravenous administration of a lipid-soluble acetate prodrug supported energy metabolism of neurons and glia. 36,37 Intraperitoneal and gastric administration of sodium butyrate reduced BBB permeability in germ-free mice. 38 –40 Additionally, the branched-chain SCFA valproic acid is anti-epileptic and neuroprotective in in vivo TBI models. 41 –45 Subclinical seizures are well described after TBI in mice 46 and though not as well established as valproic-acid, it is hypothesized that other SCFAs such as those used in our study may have anti-epileptic actions as well. 47 Acetate, which we found to be significantly reduced after TBI in fecal samples, is the primary SCFA in the systemic circulation where it can cross the BBB and act as a unique energy source for glia and possibly neurons. 36,48 –50 A potential role for SCFAs as a unique energy substrate after TBI is intriguing given previously reported findings of improved MWM performance with delayed glucose treatment when administered shortly before testing. 51
In addition, SCFAs may impact peripheral organs through their effects on systemic immunity, particularly through the regulation of lymphocyte polarization. 12,29 Recent studies have implicated activated CD8 T cells as a cause of long-term secondary injury after TBI in mice. 52 Dysbiosis and SCFA depletion weeks after TBI may contribute to a dysregulated pro-inflammatory immune phenotype and chronic secondary injury. 19,53 Our finding of improved spatial learning with supplementation of SCFAs supports further investigation of restoring SCFAs as a therapeutic target. In some settings, such as elite athletes and the military, this could represent an opportunity for a preventative approach given the relatively benign nature of dietary interventions to improve the gut microbiome and fermentation of dietary fibers, and the rapid decrease in SCFAs that we observed post-injury.
We recently published that depletion of gut microbiota with broad spectrum enteral antibiotics is associated with improved neurologic outcomes in mice after TBI. 54 In that study, although we did not detect a difference in MWM performance, mice depleted of gut microbiota prior to TBI had increased CA1 hippocampal density and reduced lesion volume. This raises the possibility of a combined therapeutic approach for patients with acute brain injury and increased gut barrier permeability 55 where the microbiome and microbial metabolites are assessed as therapeutic targets. In this approach, gut microbiota could be depleted with antibiotics to prevent harmful effects caused by translocation of bacteria or pathogen-associated molecular patterns and, simultaneously, patients are administered beneficial microbial metabolites, such as SCFAs, to prevent deficiency caused by the injury and antibiotics. This approach is highly speculative at this time. However, with the use of selective digestive decontamination already in practice in intensive care units to reduce nosocomial infection and frequent delays in initiating enteral nutrition for critically ill patients, it is relevant to consider the impact of such exposures on commensal microbiota and the loss of microbial metabolites that may be beneficial.
Conclusion
The gut microbiome is disrupted after TBI and in a sustained fashion with depletion of SCFA producing bacteria and an associated reduction of fecal SCFAs. SCFA supplementation in drinking water produced an improvement in spatial learning after TBI but did not impact lesion volume. Further study of the mechanisms of SCFA-mediated improvement in functional outcome after TBI and evaluation of interventions to raise SCFA production are warranted.
Footnotes
Funding Information
Drs. Simon, Morowitz and Clark received funding from National Institutes of Neurological Disorders and Stroke, grant number R21 NS115173. Dr. Wendell is supported by NIH grant number NIHS10OD023402. Drs. Simon and Morowitz's work was also supported by the UPMC Children's Hospital of Pittsburgh.
Author Disclosure Statement
No competing financial interests exist
Supplementary Material
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.