
Abstract
Complete spinal cord lesions interrupt the connection of all axonal projections with their neuronal targets below and above the lesion site. In particular, the interruption of connections with the neurons at lumbar segments after thoracic injuries impairs voluntary body control below the injury. The failure of spontaneous regrowth of transected axons across the lesion prevents the reconnection and reinnervation of the neuronal targets. At present, the only treatment in humans that has proven to promote some degree of locomotor recovery is physical therapy. The success of these strategies, however, depends greatly on the type of lesion and the level of preservation of neural tissue in the spinal cord after injury. That is the reason it is key to design strategies to promote axonal regrowth and neuronal reconnection. Here, we test the use of a developmental axon guidance molecule as a biological agent to promote axonal regrowth, axonal reconnection, and recovery of locomotor activity after spinal cord injury (SCI). This molecule, netrin-1, guides the growth of the
Introduction
Spinal cord injury (SCI) is a traumatic pathology that results in temporary or permanent damages in the spinal circuits and axonal tracts innervating spinal neurons. This axonal damage disrupts the information flow between brain and spinal cord, and compromises motor, sensory, and autonomic function. 1
The initial mechanical injury of ascending and descending axonal tracts and neurons localized at the epicenter of the lesion is followed by secondary damage provoked as a result of hypoxia-reperfusion, a strong inflammatory response, and the reexpression of chemorepulsive signals such as semaphorins. These factors contribute to increasing the cellular damage by triggering axonal dieback and neuronal/glial death. 2 –4 Later, the lesion transitions to a chronic stage with the formation of a glial scar that constitutes a physical and chemical barrier impeding axonal growth, tissue repair, and restoration of function.
Spinal cord injury is one of the main causes of disability among young adults worldwide 5 with the World Health Organization estimation between 250,000 and 500,000 new injuries each year. 6 Although functional impairments can be diverse, the loss of motor function particularly influences the degree of independence that affected individuals can achieve in their daily lives, directly impacting their quality of life.
To date, the only treatment that has shown any success in promoting the recovery of motor function in humans is training–based rehabilitation. 7 Therapy seems to be effective only in promoting recovery in motor incomplete lesions, however, while no significant improvement occurs in more severe injuries. 8,9 Even if the degree of preservation of local circuits is sufficient to trigger muscular activity, no voluntary movement is possible in the absence of supraspinal control. Voluntary motor control depends on descending axons originated in neurons located in the motor cortex and other supraspinal centers, jointly contacted with the spinal circuits responsible to control movement, located at and below the lesion site.
The major neuronal pathway controlling voluntary movement is the corticospinal tract (CST). It originates from pyramidal cells located in layer five of the primary motor and sensory cortices, being the only direct pathway connecting the motor cortex with the spinal cord. 10 The particular trajectory of CST allows it to transmit locomotor information to the forelimbs and hindlimbs. 11
Most SCIs in humans are anatomically incomplete, and preserved regions of white matter containing spared axons are observed even in the most severe, motor complete lesions. These axons could partially maintain synaptic interactions with the spinal circuits located below the injury site. 12 When the majority of descending pathways are interrupted by the injury, however, the source of modulation and excitation that allows the spinal circuits to be functional is suppressed. 13
Besides, both residual axons and axons with dieback as a result of the lesion show altered conduction at the margins of the lesion site. This decreases the efficiency of spontaneous reconnection between axons and target neurons and hinders spontaneous locomotor recovery. 14 This highlights the need to develop strategies to promote the regeneration of severed axons and the reconnection of these regrown axons with spared circuits below the injury site.
Axonal growth depends on both neuron-intrinsic factors and extrinsic environmental cues. The ability of central adult neurons to regenerate remains a topic of intense debate. While it is clear that regenerative capacity is greatly diminished in adulthood, 15,16 new evidence shows that regenerative programs are activated early after injury, but fail to remain active in a sustained way for mechanisms still unclear. 17 On the other side, the lesioned spinal cord is a hostile environment, and several axonal growth inhibitors are known to be upregulated or reexpressed after injury and contribute to axonal regeneration failure.
We tested the use of netrin-1 as a strategy to promote axonal regeneration across the epicenter of the lesion. Netrin-1 is a secreted protein member of the netrin family that acts as a diffusible axon guidance signal during embryonic development to establish the three-dimensional trajectory of CST. Besides its role during development, it has been shown to influence the remyelination process in adulthood. 18,19
We assessed the effect of acute administration of recombinant netrin-1 in situ in a rat model of complete spinal cord transection. 3,20,21 This precision lesion was selected because it causes minimal tissue loss and severs all axons, disconnecting spinal circuits below the injury site from all supraspinal input.
We found that a high dose of netrin-1 (2000 ng administered at 100 ng/μL) promoted significant locomotor recovery in our SCI model. This effect was because of significant axonal regeneration of descendent and ascendant axonal tracts throughout the lesion site, together with the reconnection of these regenerated axons with neuronal targets above and below the lesion site. The treatment markedly reduced lesion volume, inhibited macrophage activation and migration to the lesion, prevented axonal dieback, and preserved axonal myelination. Moreover, the treatment prevented transsynaptic degeneration. Our results highlight the therapeutic potential of netrin-1 as a biological therapy for patients with SCI.
Methods
Animals
We used male Wistar Kyoto rats from Charles River Laboratories, aged eight to 10 weeks old and weighing 200–250 g. All animal experiments were conducted in the “Laboratorio de Medicina Experimental,” Hospital Alemán, Buenos Aires, Argentina. The experimentation was approved by the Hospital Alemán Ethics Committee and was conducted according to the United States National Institutes of Health Guide for the Care and Use of Laboratory Animals. 22 The rats were housed in a temperature-controlled room at 22°C ± 2°C, with humidity of 50–60% and dark cycle of 12 h, with access to food and water ad libitum throughout the study.
Surgical procedures
Rats were anesthetized using ketamine (75 mg/kg body weight), xylazine (7 mg/kg body weight), and acepromazine (2 mg/kg body weight). Rats were placed over a heated platform to maintain the body temperature during the surgery (Kent Scientific Corporation, Torrington, CT). The blood pressure was determined, in pseudo-real time, during the spinal cord transection using a noninvasive blood pressure system (CODA® system, Kent Scientific Corporation).
Rats received complete spinal cord transection at the Th10-Th11 lamina level by a precision lesion in aseptic conditions (modification of previously described transection in 23,24 ). Briefly, after Th10-Th11 laminectomy, the dura was opened, and the spinal cord was completely transected using a needle tip (25 gauge). The severed ends of the cord after the precision lesion retracted about almost 1 mm. The lesion was inspected to check for a complete transection using a stereoscopic microscope (Stereo Zoom ST-6D). Muscles and skin were closed in layers. The netrin-1 injection is described in detail in a later section.
Before injury, the animals were randomly divided into four groups: sham, vehicle control, netrin-1 (1000 ng), and netrin-1 (2000 ng). Sham-operated rats received only Th10-Th11 laminectomy, without spinal cord transection. No post-surgical complications were observed in any of the experimental groups (sham/SCI-vehicle/SCI-netrin-1). Sham-operated rats showed no locomotor deficits after the surgical procedure.
After surgery, the rats were individually housed and received subcutaneous injection of tramadol hydrochloride (5 mg/kg body weight) as analgesia, gentamicin (5 mg/kg body weight) to prevent infections, and 6 mL lactated Ringer solution to prevent dehydration. Then, rats were kept warm (28°C) for 4 h, placed on beds of sawdust, and given manual bladder evacuation three times per day until recovery of the reflex. Moreover, hindlimb muscle stretch was performed in each rat.
Food was provided on the cage floor, and water bottles were placed to have no difficulty reaching after injury. Supplemental oral feedings, Ringer solution or/and analgesia were given as necessary.
Surgeries for retransection experiments were performed 22 days after the first spinal injury and were performed at the same thoracic level (Th10-Th11) where the first SCI took place.
Netrin-1 treatment in situ at epicenter of lesion after complete SCI
Recombinant netrin-1 (Human Netrin-1 cat No 6419-N1, R&D Systems, Minneapolis, MN) was reconstituted at 50 or 100 μg/mL in phosphate-buffered saline (PBS). At the time of injury, 20 μL of netrin-1 or vehicle (PBS) was delivered in the epicenter of the lesion, filling the entire injury area. The infusion was performed with the tip of the pipette at a right angle, and at a 45-degree angle in both cranial and caudal orientation. 3 A total of 1000 or 2000 ng of netrin-1 was administered. Delivery of netrin-1 was performed using a calibrated micropipette (Proline® Plus, Sartorius, Bohemia, NY). The whole process was conducted under a stereoscopic microscope.
Behavioral testing
To assess the effects of netrin-1 treatment in the recovery of locomotor functions after a complete SCI, we performed four behavioral tests. In all tests, the animals were acclimated before receiving the injury (starting five days before SCI) and during testing, either in the open field as well as on the ladder and in the straight field. Acclimatization was performed for two 10-min cycles before test. 3 After each trial, the platforms were cleaned using 70% ethanol.
Analysis for the four tests was performed in a sound attenuated room following a blind experimental design.
Open field locomotor test: Basso Beattie Bresnahan motor rating scale (BBB score) was evaluated in rats after SCI in an open field platform (1 m2). 25
Ladder rung coordination test: The apparatus consists of a straight platform of 1 meter length with rungs (14 cm of length and 4 mm of diameter) separated 10 mm between each other. This test is used to determine a specific walking task—the hindlimb coordination recovery after a complete SCI. Video recording was obtained in high definition. The number of steps slipped and steps missed (foot fall errors) were quantified per crossing. Three repetitions with 4 min of resting time were done for each condition in each day analyzed. The values of steps missed and steps slipped were averaged separately. 26
Hindlimb step length quantification during movement: To perform this analysis, we used a straight platform of 1 meter length with a transparent floor. Below the platform, a mirror at located at 45 degrees. The video recording was acquired focusing the mirror to obtain a ventral image of the rats. Quantification of steps length in each limb was performed importing the video images (Movie FFMPEG plug-in) to Fiji Image Software (NIH; Bethesda, MD) to trigger the analysis of length pattern with manual tracking plug-in.
Angle of displacement generated between the right and left hindlimb during the movement. Rats were placed in the same platform that was described above. Angle analysis was performed using the angle measurement from Fiji Image Software. The software determined the angle generated between the left hindlimb versus right hindlimb during movement.
Perfusion, fixation, and tissue preparation
Rats were transcardially perfused with PBS/heparin (0.05% v/v) followed by 4.2 % paraformaldehyde (PFA) in PBS. Spinal cord, brain, and sciatic nerve were removed and post-fixed for 48 h at 4°C in the same fixative solution. Then the tissues were treated with a sucrose gradient (15–30%) for 48 h to protect the tissue against ice crystal formation during freezing. 3,21 After sucrose treatment, the dura mater (meninges) was removed.
Inclusion of tissues in paraffin blocks
Axial sections of spinal cord and sciatic nerve were fixed in PFA and then in buffered formalin; then the fixed tissues were embedded in paraffin. To analyze the cellular structures, sections of 1.5 μm were cut and mounted on slides.
Immunohistochemistry
Spinal cord tissue, protected by sucrose, was frozen to obtain 30-μm cryostat longitudinal slices (cranial to caudal and dorsal to ventral) using a Leica CM 1850 cryotome. To perform the analysis, 12 serial slices (dorsal to ventral) of spinal cord were collected from each specimen (sham, vehicle control, and netrin-1). Immunohistochemical analysis was performed as described previously. 27,28
Free floating slices were first incubated with high ionic force blocking buffer (HIFBB) for 50 min and later treated for 20 min with 0.15 % Triton X-100 in PBS for permeabilization. Solutions of primary and secondary antibodies were prepared in HIFBB. Incubation time for primary antibodies was 3 h at room temperature, while secondary antibodies were incubated ON at 4°C. Nuclear labeling was performed with 4′,6-diamidino-2-phenylindole (DAPI) for 1 h at room temperature. Controls to determine nonspecific binding were performed using free floating slices treated only with secondary antibodies using different IgG isotypes. All analyses were performed by an observer blinded to experimental conditions.
Anterograde, retrograde, and transsynaptic tracing
Anterograde tracing of CST axons was performed in rats treated with vehicle control or netrin-1 25 days after complete SCI. Fluoro-Ruby (FR) dextran tracer (25% in dH2O) from Molecular Probes (D1817, Eugene, OR) was pressure injected at six different sites into the sensory-motor cortex using a microcapillary tube (Sigma-Aldrich, St. Louis, MO.), 0.5 μL per site. The microcapillary tube remained in place for 2 min after each dextran tracer delivery. The flow rate was, on average, 5 nL/sec. The process was performed under a stereoscopic microscope (Stereo Zoom ST-6D) in a apparatus Stereotaxic (Harvard Apparatus Compact Stereotaxic Instrument, Holliston, MA) following these coordinates: 1.5, 0 and −1.5 mm anterior to bregma, 2, −2 mm lateral to bregma, and 1.5 mm depth, adapted from. 29 Rats were killed 14 days after stereotaxic surgery.
Retrograde tracing of rat sensory axons at the sciatic nerve was performed as described previously. 30 In brief, 25 days after complete SCI, rats treated with vehicle control or netrin-1 were injected with 2 μL of Fluoro-Emerald (FE) dextran tracer (30% in dH2O) from Molecular Probes (D1820, Eugene, OR) into the right and left sciatic nerves to trace sensory axons in the dorsal column at lumbar segment 2 of the spinal cord. Dextran tracer was loaded into a microcapillary tube and injected using a manual syringe pump. The flow rate was, on average, 30 nL/sec. The microcapillary tube remained in the sciatic nerve for 2 min after delivery. Also, the process was performed under a stereoscopic microscope (Stereo Zoom ST-6D). Eleven days after tracer delivery, rats were killed, and the spinal cord and sciatic nerve tissues were obtained by a perfusion-fixation process.
Sensory and motor circuits were retrograde traced to explore the relationship between regenerated neuronal structures and synaptic functions across the injured spinal cord. For this purpose, we injected in the sciatic nerve of rats treated with vehicle control or netrin-1 a transsynaptic marker, 25 days after complete SCI. Two μL of wheat germ agglutinin fluorescent conjugate (WGA, 1 mg/mL, W11261, Molecular Probes) was loaded into a microcapillary tube and injected using a manual syringe pump. The flow rate was, on average, 30 nL/sec. The microcapillary tube remained in the sciatic nerve for 2 min after WGA delivery. Twelve days after injection, rats were killed, and the spinal cord and sciatic nerve tissues were obtained by a perfusion-fixation process. After that, the immunohistochemistry technique was performed as described above.
All surgical procedures were performed under aseptic conditions.
Clearing technique
Whole rat spinal cord tissues (FR CST labeled) were incubated in tetrahydrofuran (THF) (Biopack, Prod. Quim., Buenos Aires, Argentina)-distilled water [50% (vol/vol)] for 1 h, in THF-distilled water [80% (vol/vol)] for 2 h, in THF (100%) for 1.30 h, followed by fresh THF (100%) for 24 h, and dichloromethane (Biopack) (100%) for 2 to 24 h. Finally, the spinal cords were incubated in a BABB solution (mixture of benzyl alcohol [Biopack] and benzyl benzoate [Biopack] at a ratio of 1:2) until samples became transparent. 21 All incubations were developed under shaking in the dark.
Plastic embedded tissue for high-resolution fluorescence microscopy
Spinal cord and sciatic nerve from rats injected with FE were dehydrated with acetone (Biopack) for 60 min at 4°C. After that, tissues were included in historesin (Reichert-Jung, Depew, NY, catalog number: 7O-2218-500). The inclusion media (tissue plus historesin) was put in blocks. Orientation was longitudinal for the spinal cord and transversal for the sciatic nerve. The blocks were incubated at 4°C ON to reach the polymerization and hardening. Then, the blocks were cut in ultrathin sections (1 μm), 50 serial slices per condition, using a microtome (Leica RM2255) equipped with a tungsten carbide blade. The process was performed in the dark to preserve the fluorescent signal.
Image acquisition
Epifluorescence microscopy
Images were collected using a Nikon Eclipse E400 epifluorescence microscope (Nikon Instrument Group, Melville, NY) equipped with CFI Plan Apo Lambda objectives (4X-NA: 0.1; 10X-NA: 0.45; 20X-NA: 0.75, and 60X-NA: 1.4, all suitable for confocal microscopy) and Nikon camera DS-Fi1 (Nikon Instrument Group). Illumination source was through a mercury arc lamp. The beam passed through bandpass filters: Dapi cube (375/28), Alexa 488 cube (480/30), and Cy3 cube from Chroma Technology Corp., Bellows Falls, VT. Images were obtained in eight bits (TIFF format) and then were deconvolved using Huygens compute engine 3.5.2p3 64b (close platform) with Tikhonov method and Laplacian stencil. Theoretical Point Spread Function (PSF) was obtained with Diffraction PSF 3D plug-in from Fiji software.
One-photon confocal microscopy
Images were acquired using an Olympus Fluoview 1000 Confocal Microscope (Olympus Headquarters Corporate, Philadelphia, PA) equipped with 10X objective UPLSAPO, NA: 0.40; 20X objective UPLSAPO, NA: 0.75, and 60X objective PLANAPO, NA: 1.42, oil immersion). Three-dimensional images were collected taking z-series between 350–800 optical slices of Airy unit = 1 airy disk with intervals in accordance with Nyquist theory (optimum overlapping to minimize photobleaching). 21
The confocal images were collected in 12 bits per pixel, and the scan speed (time per pixel in arbitrary units Olympus) was 8.0 to 10X magnification and 4.0 to 20X and 60X magnification. The excitation wavelength laser used was 473 nm for Alexa Fluor 488 and FE; and 559 nm for Cy3 and FR. The settings of PMT voltage, gain, and offset were in average: 400, 3125 and 1000, respectively (in all cases arbitrary Olympus units).
Image processing and analysis
Assessments of scar and injury area after complete SCI
Scar area quantification was performed in whole tissue using the area measurement function from Fiji Image Software. Tissue images were previously calibrated by set-scale plug-in, and the region of interest (ROI) was delimited by freehand function. The ROI area analysis was applied five times in each image, and the values obtained were averaged.
Regarding quantification of injury area on each axial segment of spinal cord (slice thickness 30 μm), the method was the same mentioned above. Summation of the area on each axial segment was averaged in each condition.
Axonal length
The entire z-axis stacks from the spinal cord collected by confocal microscopy were imported in a calibrated format to IMARIS 3D software v6.3.1 (Bitplane Sci Software, Zurich, Switzerland). The length quantification of axonal tracts in three dimensions was performed using the measurement point function of IMARIS. Trajectory of each axonal tract was tracked manually with a measurement point from the beginning of the tract to the end of the tract, creating a filament render 3D with the shape of the axons (X/Z). Using this strategy, an accurate measure of the axonal tract length was obtained, as described previously. 31
Axonal branching
Quantification of axonal branching was performed using a projection of z-stack images from the epicenter of the lesion as well as from upstream or downstream areas. The count of sprouting was performed using filament function of IMARIS and volume viewer plug-in from Fiji software.
Axonal tracer
Images in TIFF format were imported to macro Axon Tracer for Automated Quantification of Axon Regeneration in Spinal Cord Tissue. 32 The ROI was applied to determine the lesion area. Axons were displayed with yellow mask to perform the length quantification.
CST axons and synpasin-1 cluster quantification
Quantification of CST axons located ventral to the dorsal column as well as quantification of synpasin-1 clusters was performed using the analyze particle function from Fiji Software. Circularity was defined between 0.00–1.00, showing an overlay mask in situ.
Quantification of myelinated axons (Fast blue-positive axons) was performed using the manual multi-point counting function in 60X CST images. Quantification of the CST myelinated area was performed using the freehand lines function. In all cases, analysis was performed three times for each image and then averaged.
CST area and volume quantification
Area and volume occupied by CST axons at the epicenter of the lesion were measured using confocal z-stack images imported in IMARIS 3D. To determine the area of CST, the clipping plane function was used. Longitudinal tracts were clipped in z orientation obtaining a coronal view of CST allowing the measurement of surface using the measurement point function. To determine the volume of CST, contour surface function was applied.
3D-video animation
In motion video based on each group of z-images from 1-photon confocal microscopy was performed using the animation plug-in from IMARIS 3D.
Antibodies
The primary antibodies listed below were used: mouse monoclonal IgG anti- monocyte chemokine C-C motif ligand 2 (CCL2) (1:200) for IHC from Millipore, mouse monoclonal IgG anti-neurofilament medium molecular mass (NFM) (1:500) from Invitrogen, mouse monoclonal IgG anti-βIII-Tubulin (neuronal) (1:500) for IHC from Sigma, and mouse monoclonal IgG anti-synapsin-1 (Syn-1) (1:200) for IHC from Sigma, rabbit Monoclonal IgG anti-glial fibrillary acidic protein (GFAP) (1:150) for IHC from Cell Marque.
The secondary antibodies used for IHC were DyLigthTM (488) and Cy2 TM, Cy3TM, all from Jackson ImmunoResearch. The DAPI was from Molecular Probes (Eugene, OR).
The specificity of the secondary antibodies was confirmed by omitting the primary antibody. No immunoreactivity was observed in any of these control tissue samples.
Statistical analysis
Graph-Pad Prism software Version 5.0 (Graph-Pad software, Inc., La Jolla, CA) was used for data analysis. Results are presented as mean ± standard deviation (SD) of at least six independent experiments. For each variable/parameter analyzed (i.e., BBB score, ladder rung, axonal length, axonal branching, number of CST axons, etc.), values obtained from the same animal were averaged and considered one independent observation.
Comparisons were performed using unpaired two-tailed Student t test or one-way analysis of variance (ANOVA) followed by the Tukey multiple comparison test or the Bonferroni multiple comparison test, where appropriate. Significance level was set at p = 0.05.
Results
In situ administration of high doses of netrin-1 promotes voluntary locomotor recovery after complete SCI
To test the potential of netrin-1 to promote functional recovery after SCI, we used a model of complete spinal cord transection. The complete spinal transection was performed at thoracic level 10–11, resulting in immediate paralysis of the hindlimbs; 1000 ng or 2000 ng of netrin-1 or vehicle was infused at the epicenter of the lesion immediately after the lesion was performed.
As expected, 24 h after a complete spinal cord transection, both vehicle- and netrin-treated rats showed a complete paralysis of the lower body, dragging their abdomen and tail during locomotion.
To assess the extent of recovery of hindlimb movement, we performed the BBB test (Basso-Beattie-Bresnahan locomotor rating scale). At 21 days after injury, no recovery was observed in vehicle-treated animals, presenting a BBB score of 0.78 ± 0.6. This value is consistent with slight or no movement in one of the hindlimb joints (ankle, knee, or hip), and inability of the animals to bear their own weight.
In contrast, rats treated with netrin-1 presented a marked locomotor recovery. Netrin-1–treated animals presented a straight trunk position with vertebral column parallel to the floor and lifting of the tail (Fig. S1A and Supplementary Video S1). Moreover, rats presented consistent weight-supported plantar steps in the stance during the gait (Fig. 1B). This motor recovery was reflected in a significant improvement in the BBB score values at 21 days (netrin-1 1000 ng: 11 ± 3.7, netrin-1 2000 ng: 14.9 ± 0.7; p < 0.001) (Fig. 1C).
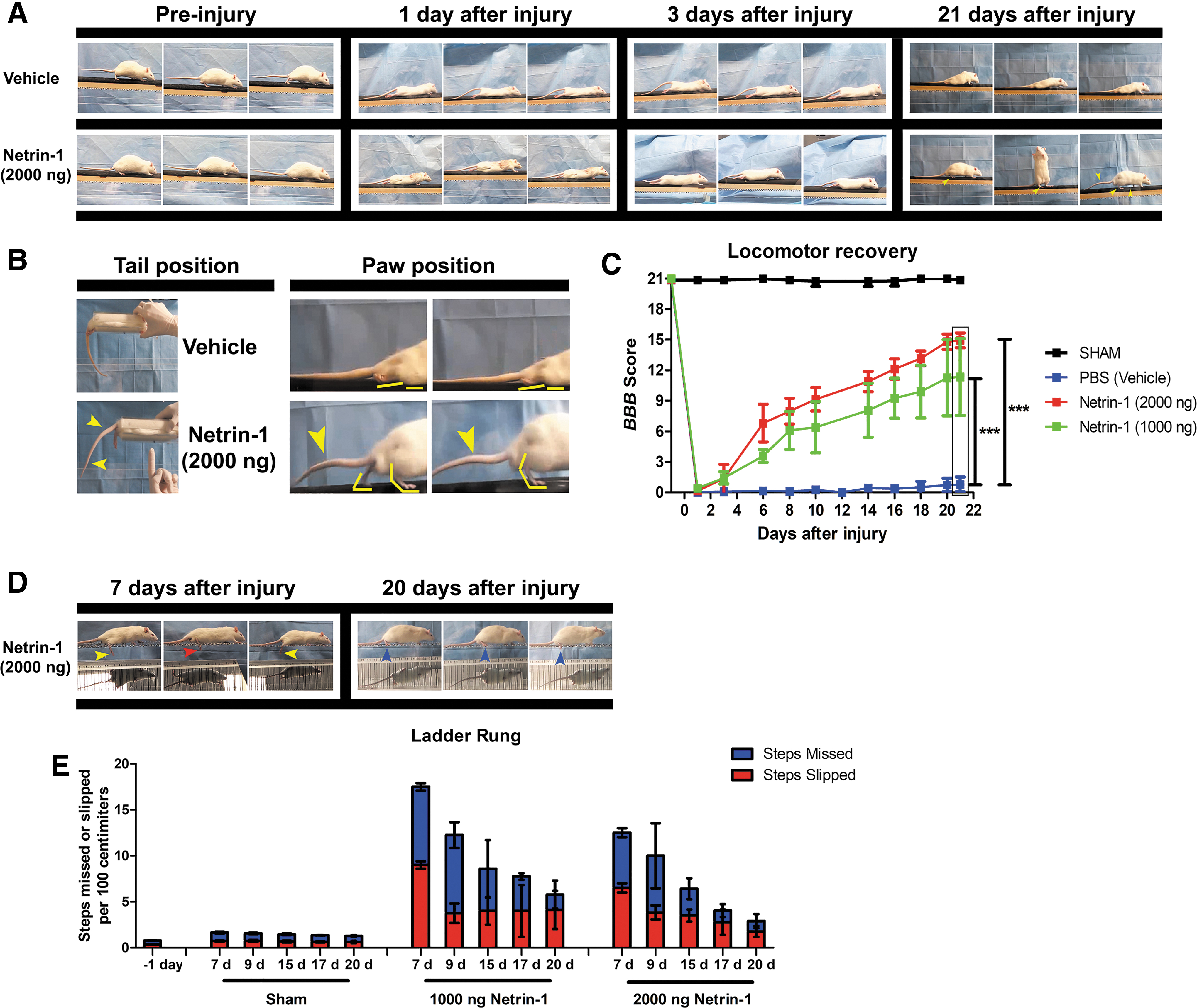
Evaluation of locomotor recovery after spinal cord injury (SCI). (
To further assess the recovery of coordinated movement we performed the ladder rung test. 33
From day 7 to day 20 after injury, we observed that rats treated with 2000 ng of Netrin-1 had a progressive decrease in the number of both slipped steps (SSday7: 6.5 ± 0.5 – SSday20: 1.76 ± 0.59) and missed steps (MSday7: 6 ± 0.5 - MSday20: 1.42 ± 0.50); as well as improved rung grasping (Fig. S1D-E, and Supplementary Video S2). Moreover, the netrin-1 effect on ladder rung performance was dose dependent, and a significant difference was observed between rats treated with 2000 ng and 1000 ng of netrin-1 on day 20 (Fig. 1E).
Although both doses of netrin-1 improved the BBB score and the performance in the ladder rung, treatment with 2000 ng of netrin-1 had a more consistent effect. For this dose, all rats in the experimental group reached the late phase of locomotor recovery (BBB score: 14.9), corresponding with recovery of locomotor coordination (BBB score ≥14). For this reason, we selected a dose of 2000 ng of netrin-1 for the following experiments.
To further characterize the extent of locomotor recovery, we measured step length and hindlimb angle. We did not find significant differences in the step length of rats treated with netrin-1 compared with sham-operated rats, for both hindlimbs. Moreover, we did not find significant differences in the length of the left step compared with the length of the right step in netrin-1 treated rats (Supplementary Fig. S1A-B and Supplementary Video S3). Also, the angle between the hindlimb and the main body axis during stepping presented no significant difference between netrin-1 treated rats and sham controls (Supplementary Fig. S1C-D).
These observations show the recovery of hindlimb function was symmetrical, and indicate that a complete bilateral transection was successfully performed in these animals. These results suggest that in situ administration of netrin-1 at the epicenter of the lesion promotes the recovery of locomotor function after a complete spinal cord transection.
In situ netrin-1 administration reduces the lesion size 24 days post-injury
As mentioned above, we performed a complete transection of the spinal cord at thoracic level 10–11, severing all axons at the lesion site without provoking significant tissue loss. Figure 2A shows the complete section of spinal cord. The systolic pressure was measured to evidence the neurogenic shock triggered by the injury (Fig. 2B). These results show that not only the lesion applied was anatomically consistent across animals, but also that the physiological effects of the lesion were reproducible.

Precision transection at thoracic 10 – 11 site. (
At 24 days after injury, the tissue discontinuity produced by the transection was bridged in all animals (Fig. 2C, top panel). Scar area was reduced in Netrin-1 treated rats compared with controls. This effect was evidenced by a digitally generated image showing the complete discontinuity of tissue at the lesion site in rats treated with vehicle (shown in white) (Fig. 2C, bottom left panel) in contrast to that observed in netrin-1 treated rats, where the preservation of tissue was evidenced by an increase in the number of pixels (ramp of color green-yellow-red) (Fig. 2C, bottom right panel).
Quantitative analysis of the scar area showed a significant decrease in lesion size in netrin-1 treated animals (p = 0.0037) (Fig. 2D). We performed a volumetric analysis of the injury by measuring the lesion area on each axial plane (dorsal to ventral) along the spinal cord, in each spinal cord. We observed a significant decrease in the injury size in netrin-1 treated rats compared with vehicle-injected controls (p = 0.0083) (Fig. 2E-F).
Netrin-1 reduces CCL2 levels
We observed an intense CCL2 fluorescent signal in the vicinity of the lesion (upstream from injury) in vehicle-treated rats (Supplementary Fig. S2A, left panel). The CCL2 signal was markedly reduced in netrin-1 treated rats (p = 0.0423) (Supplementary Fig. S2A, right panel and Supplementary Fig. S2B). Notably, in the 3D surpass image from rats treated with vehicle, we observed cells with macrophage-like phenotype displaying the CCL2 signal (left inset, white arrowheads) and CCL2 clusters inlaid into astrocyte ramifications (yellow arrowheads). In contrast, netrin-1 treated rats only presented a few clusters of CCL2 inlaid in the astrocyte processes, and no diffuse fluorescent CCL2 signal was present in the spinal cord tissue (right inset, yellow arrowheads).
Netrin-1 treatment promotes axonal regeneration across the lesion site
We analyzed a region at the epicenter of the lesion corresponding to THE gray matter-intermediate area, where the majority of regenerated axons come from propiospinal neurons. We observed no NFM-positive axons at the epicenter of the lesion site in rats treated with vehicle—only a few NFM-positive fragments. In contrast, we found a sharp NFM-positive signal indicative of axonal regrowth at the epicenter of the lesion in netrin-1 treated rats (Fig. 3A). The higher levels of NFM signal observed correspond to a significant increase in axonal length, number of regenerated axons, and number of branches (p < 0.001) (Fig. 3B).

Axonal regeneration after netrin-1 treatment. (
In addition, we analyzed the axonal structure in spinal cord white matter right upstream and downstream of the injury site. Upstream of the lesion in vehicle treated rats we found axonal tracts fragmentation (Fig. 3C, left panel and inset). This structural alteration is a common feature and reflects two different processes: axonal dieback occurring at the proximal fragment of the transected descending axons and wallerian degeneration occurring at the distal portion of transected ascending axons. Netrin-1 treated rats showed a high preservation of the structure of axonal tracts, with no signs of axonal degeneration. Moreover, shaped structures like growth-cone were present in the tips of some of these axons (Fig. 3C, right panel and inset).
When we analyzed downstream areas of white matter, a reduced number of NFM-positive axons were observed in rats treated with vehicle in comparison with netrin-1 treated rats (Fig 3D,E).
Further, in rats treated with vehicle, pre-synaptic clusters labeled with synapsin-1 were restricted only to the gray matter areas (Fig. 3D, left 3D-render inset). Conversely, netrin-1-treated rats presented a strong synapsin-1 fluorescence signal, and synapsin-1 clusters not only spread out in the gray matter but also in white matter areas. These clusters appeared localized in the vicinity of the focal plane of the axons entering/exiting white matter tracts and following the same perpendicular trajectory (Fig. 3D, right 3D-render inset).
Next, we quantified the number of axonal ramifications in white matter areas upstream of the lesion. The degree of ramification could impact the ability to form new connections and maintain neural activity after injury and could even compensate for deficits in regeneration. The tridimensional analysis showed a significant increase in the number of axonal branches in netrin-1 treated rats in comparison with vehicle (white matter volume analyzed = 4.2 e6 μm3) (p < 0.001, Supplementary Fig. S3). Moreover, the number of ramifications observed in the vicinity of the lesion site in netrin-1 treated rats corresponds to 64% of sham controls at identical metameric segments.
Regrowth of ascending and descending axonal in white matter
A representative image of a longitudinal segment of injured spinal cord shows the lesioned area, delimited by reactive astrocytes (Fig. 4A, left top panel). In netrin-1 treated rats, we found a significant number of NFM-positive axonal tracts crossing the transected white matter; however, this was not observed in vehicle-treated rats (p = 0.0011) (Fig. 4A-B). Interestingly, the 3D analysis showed that regenerated axons presented a tortuous trajectory across the lesion, outflanking areas devoid of tissue (Fig. 4A, white arrowheads point to the axonal trajectory in top row, high magnification images).

Axonal tracts regeneration in white matter. (
Surprisingly, this axonal growth follows the hystotypical pattern of the spinal cord white matter, where axons are highly aligned and oriented in the direction of the spinal cord main axis (Supplementary Video S4). Guidance comes either from other signals in the tissue or from the Netrin-1 in interaction with other components. During development, Netrin-1 is known to act locally by promoting axonal growth cone adhesion and growing by a mechanism called haptotaxis. 34,35
Corticospinal tract regeneration at the epicenter of the lesion site
In our rat model, we measured the regeneration of CST across the injury site. The FR was injected to label corticospinal axons (Fig. 5A). To perform the analysis, we used a modified clearing technique 21 (Fig. 5B,C), followed by confocal image acquisition. We observed fragmentation of CST axons immediately upstream of the lesion site in spinal cords from rats that were treated with vehicle, consistent with retrograde axonal degeneration (Fig. 5D, left panel). Netrin-1 treatment promoted the regrowth of CST across the injury site. Axons were observed in the tissue bridges connecting cranial and caudal sides of the injury, and flanking segments without tissue (Fig. 5D, red arrowheads in the right panel).
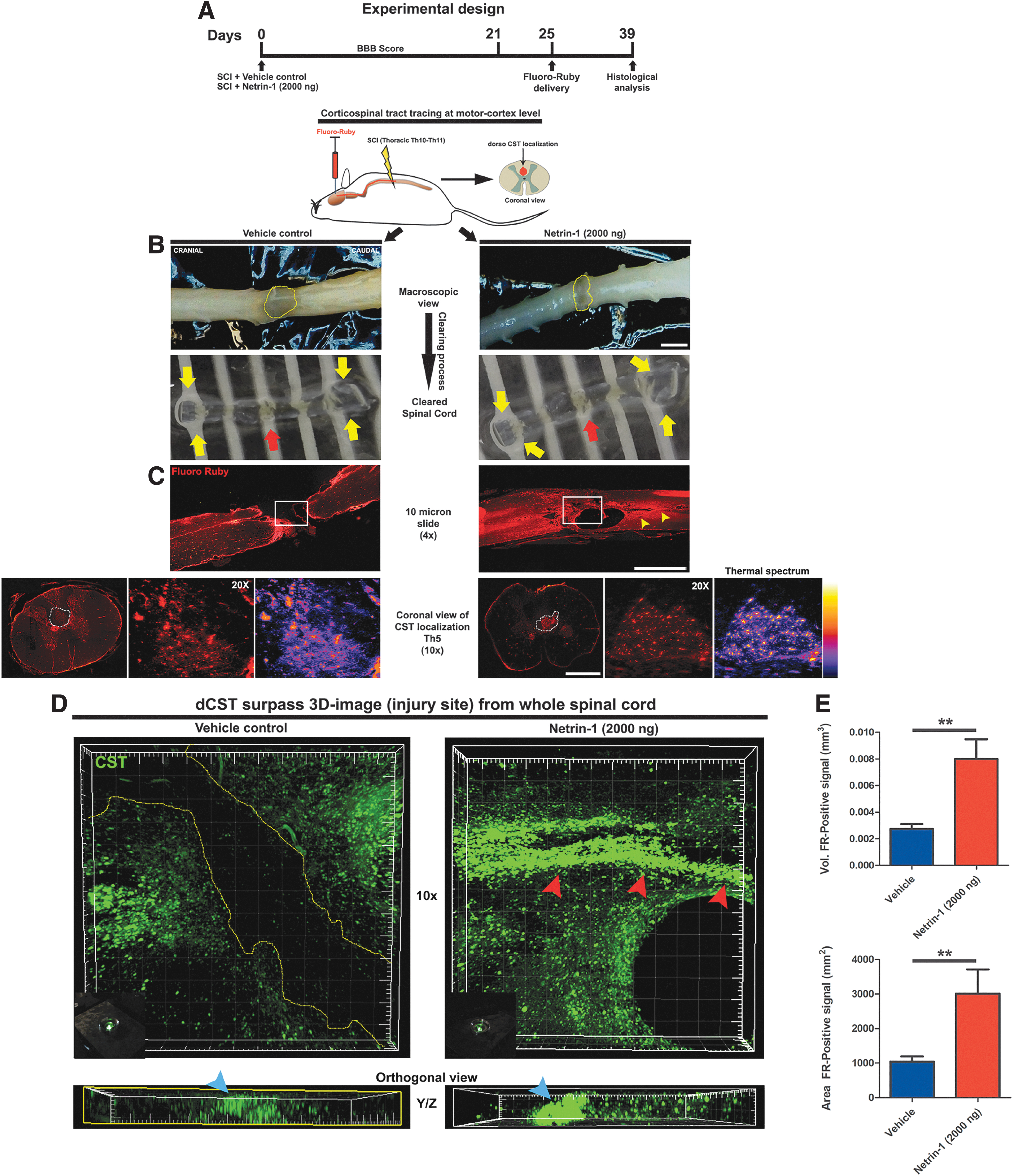
Corticospinal tract (CST) regeneration at epicenter of the lesion. (
We observed a strong CST fluorescent signal in Netrin-1 treated rats. This signal was limited to the anatomical segment of CST localization in the spinal cord (orthogonal view in Fig. 5D, blue arrowheads point to the CST localization). The quantitative analysis of the volume and area of FR-positive axonal tracts showed a significant increase in netrin-1 treated rats compared with rats treated with vehicle (p = 0.0088) (Fig. 5E).
In addition to mechanical transection of CST after initial injury, CST dieback increases the distance that the axons need to grow to reach the other side of the lesion. Moreover, axons undergoing dieback are in a nonregenerative state; as cytoskeleton collapses, growth cones retract forming retraction bulbs.
We found a significant decrease in the number of NFM-positive CST axons upstream of the lesion site in spinal cords from vehicle-treated rats, in comparison with netrin-1 treated rats (p = 0.0044) (Fig. 6A,B). Further, we observed a significant preservation of CST myelinated-axons (Fast blue labeling) of netrin-1 treated rats, whereas the number of Fast blue positive CST axons was lower in vehicle treated rats (p = 0.0009) (Fig. 6C).

Corticospinal tract (CST) integrity upstream of the lesion site. (
Axonal regrowth across the lesion prevents transsynaptic degeneration at lumbar segments
The recovery of motor function depends not only on axonal regeneration but also on proper connections being established between regrown axons and spinal circuits.
Accordingly, we determined the number of pre-synaptic interactions between descendent axonal tracts with second order motor neurons at lumbar segments after SCI.
In netrin-1 treated rats, we found a significant increase in the number of pre-synaptic clusters surrounding the motor neuron surface in the ventral horn of the lumbar spinal cord (Fig. 7A). The magnifications in Figure 7A (right panel) show representative motor neurons of lumbar spinal cord from netrin-1 treated rats. The surfaces of motor neurons are surrounded by white matter axons entering gray matter and pre-synaptic clusters of synapsin-1 localized in the vicinity of the focal plane of the axonal shaft (yellow arrowheads). In contrast, lumbar spinal cord from vehicle-treated rats showed a reduced number of pre-synaptic clusters in the surface of motor neurons (Fig. 7A magnification, left panel).
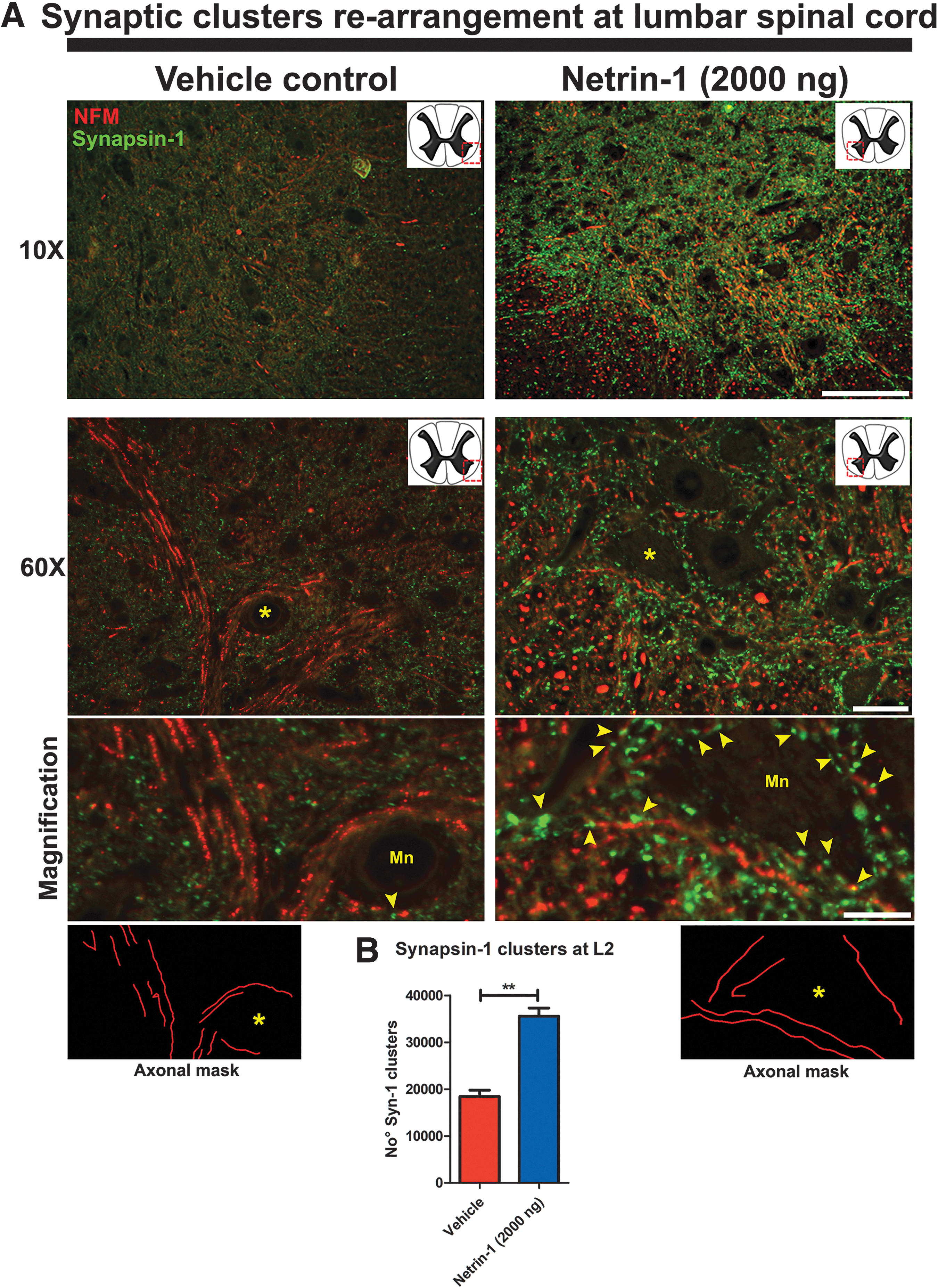
Rearrangement of synaptic clusters in gray matter at the lumbar segment. (
The axonal masks in the bottom of Figure 7A show the trajectory of the axonal tract that contacts the motor neurons. The number of synapsin-1 clusters in the ventral horn from netrin-1 treated rats was double that observed in vehicle-treated rats (p < 0.05) (Fig. 7B).
Netrin-1 treatment preserves integrity of sensory axons at the lumbar segment after SCI
Next we investigated how the thoracic SCI affected the structural integrity of ascending sensory axons at the lumbar dorsal column. For this purpose, we injected the FE axonal tracer in the sciatic nerve from rats treated with vehicle or netrin-1 after SCI (Fig. 8A,B). We performed a quantitative analysis of the sensory axonal tract localized in the spinal cord dorsal column at lumbar segment 2. We found preservation of sensory axons only in rats that were treated with netrin-1 after SCI, whereas we found a total absence of FE-positive axons in the dorsal column of rats that were treated with vehicle (Fig. 8C, red arrowheads in the magnification show the axonal shaft). The left bar graph (Fig. 8D) shows significant differences in the number of FE-positive axonal tracts in netrin-1 treated rats compared with vehicle treated rats (p = 0.0014).

Integrity of ascending sensory axons at the lumbar dorsal column after spinal cord injury (SCI). (
Moreover, we found in the dorsal column upstream of the lesion site (10 mm cranial to lesion) a significant preservation of sensory axons and axons from second order neurons in netrin-1 treated rats (p = 0.0499) (Supplementary Fig. S4).
These results support a neuroprotective/neuroregenerative role of netrin-1 distant from the lesion site, in a similar fashion to the prevention of CST dieback.
Transsynaptic reconnection after complete SCI
The next step was to investigate the transsynaptic interaction between regenerated axons and their final neural targets. For this purpose, we injected wheat germ agglutinin WGA-Alexa-488 in the sciatic nerve. The WGA is a fluorescent lectin that has the property to do transcitosis when a pre- post-synaptic contact takes place.
After injection, we analyzed the efficiency of WGA delivery into the motor neurons localized in the ventral horn of the lumbar spinal cord (Fig. 9A). Figure 9B shows the WGA fluorescent signal (white arrowheads) in the soma of motor neurons (NFM positive signal) in both vehicle- and netrin-1 treated rats. In netrin-1 treated rats, the WGA signal was also observed in axonal tracts upstream of the lesion site, colocalizing with NFM-labeling in axonal shafts (Fig. 9C, right panel). This indicates that these axons regrew across the lesion and contacted lumbar motor neurons below the lesion site. The WGA labeling was not observed in axons upstream of the lesion site in vehicle treated rats (Fig. 9C, left panel).
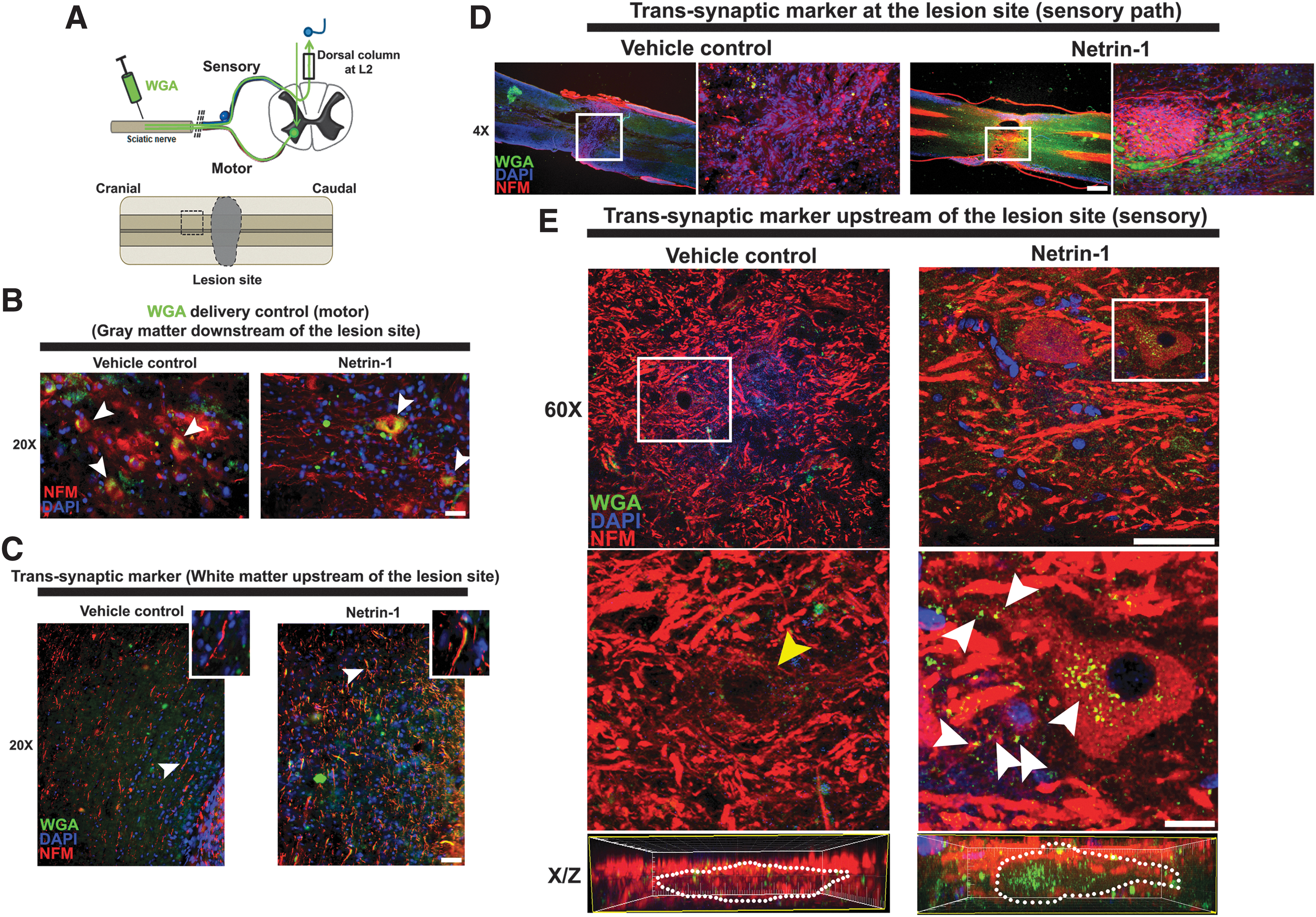
Transsynaptic interaction after complete spinal cord injury (SCI). (
In addition, we determined the sensory connection between the ascending axons from lumbar segments of the dorsal column with second order neurons localized in gray matter areas upstream of the lesion site (cartoon in Fig. 9A). Only the rats that were treated with netrin-1 presented a significant increase in the length of NFM-positive axons (p = 0.055) at the epicenter of the lesion, which were surrounded by green WGA clusters (Fig. 9D, white frames, and Supplementary Fig. S5).
Moreover, upstream of the lesion site, we observed neurons with a WGA fluorescent signal not only in their soma but also in their neurites (Fig. 9E, white arrowheads shows WGA clusters). Further, in the orthogonal confocal (X/Z) view, we detected a marked accumulation of green WGA inside of the neurons. In contrast, we did not find green WGA signals in the neurons localized upstream of the lesion site in rats treated with vehicle.
This functional reconnection would explain the locomotor recovery observed in rats with SCI after treatment with netrin-1. Reconnection would allow motor information to reach the hindlimb muscles through the sciatic nerve. This is supported by the observation that axons from the sciatic nerve presented a significant preservation of neurofilament polymerization in netrin-1 treated rats (p < 0.001) (Supplementary Fig. S6).
Recovery of locomotor activities responds to neural reconnection of regenerated axonal tract that crosses the lesion site
To test whether locomotor recovery was the result of axonal regeneration across the lesion or the result of spinal circuit plasticity below the lesion site, we performed a retransection of the spinal cord in Netrin-1 treated rats 22 days after the initial injury and in the same anatomical location. We observed a complete loss of locomotor activities previously recovered in Netrin-1 treated rats (Fig. 10A).
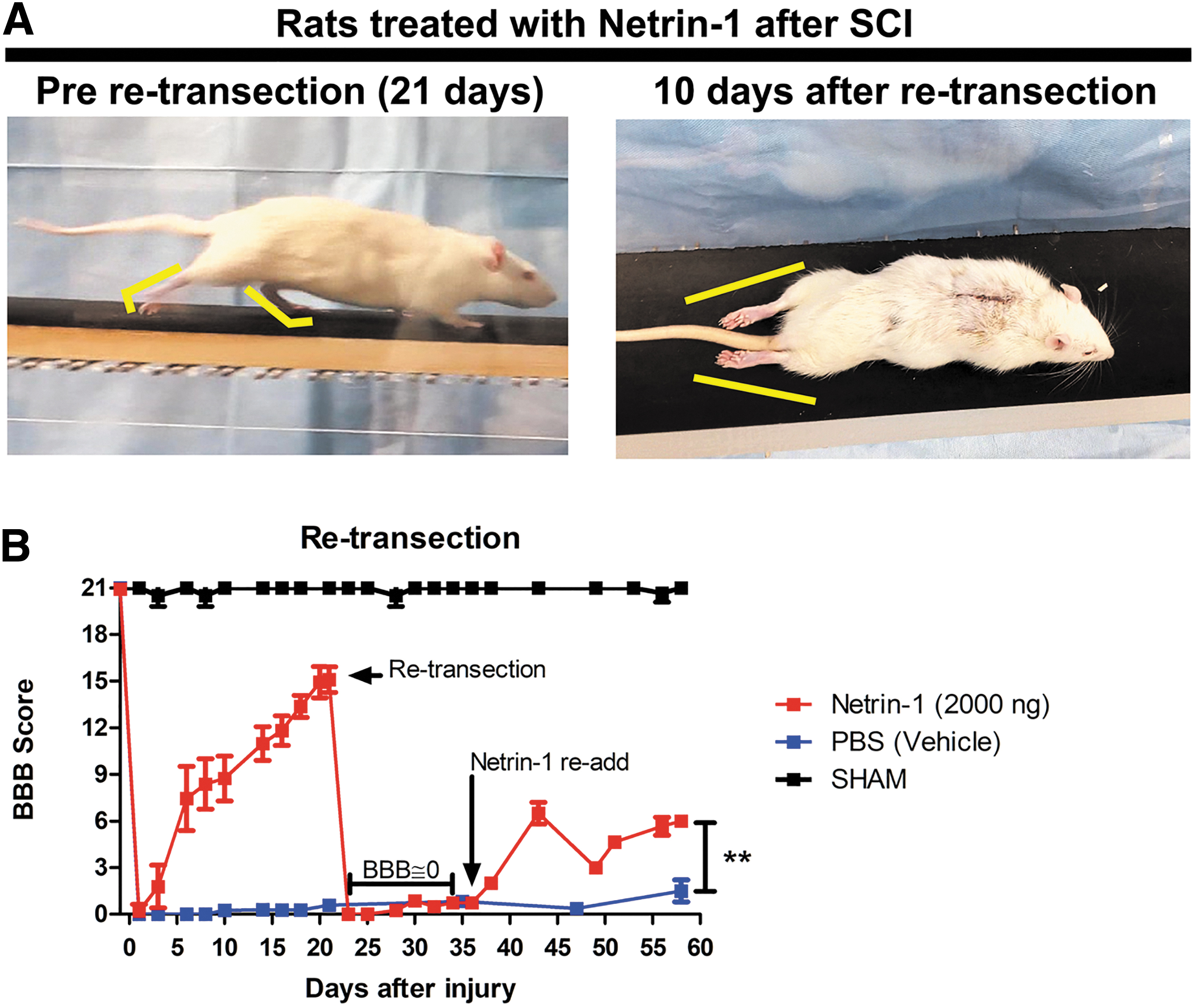
Retransection of spinal cord after locomotor recovery. (
Accordingly, the BBB score dropped to zero and remained at this level for 15 days. To further confirm the regenerative properties of netrin-1, 2000 ng of netrin-1 were reapplied in situ 15 days post-retransection (37days after initial injury). We found a slight but significant increase in the BBB score values 21 days after treatment compared with vehicle treated controls (p < 0.001) (Fig. 10B). No spontaneous locomotor recovery was observed in vehicle treated rats during the course of the experiment (60 days). These findings suggest that the locomotor recovery observed in netrin-1 treated rats was because of regeneration of the axonal tract across the lesion site, allowing the reconnection of spinal projecting neurons, rather than because of a compensatory response below the lesion site.
Discussion
The present study was designed to assess the effect of netrin-1 on axonal regeneration after SCI. Netrin-1 was selected as treatment based on its
The model is ideal to study axonal regeneration of long ascending and descending axonal tracts in the absence of sprouting from spared axons. As expected after an anatomically complete thoracic lesion, we observed a complete loss of locomotor function coinciding with the interruption of all motor cortex projections to lumbar segments and ascending sensory projections. 36
Our results show that netrin-1 treatment administered in situ at the time of injury induced a significant recovery of voluntary locomotion 21 days after injury. In the BBB locomotor rating test, netrin-1 treated rats reached the late stage of recovery, consistent with coordinated stepping. This observation was confirmed with the ladder rung test, where treated animals presented a significant reduction in the footfall errors. Finally, the step length and hindlimb rotation in treated animals showed no significant difference with sham operated controls. The regain of voluntary hindlimb movement suggests that lumbar circuits recovered their supraspinal inputs. The fact that no spontaneous recovery was observed in vehicle treated rats indicates this recovery is mediated by netrin-1.
Restoration of the locomotor function after anatomically complete SCI requires axon regrowth across the lesion site to reach the white matter and spare lumbar gray matter below the lesion and form new connections with local circuits. 37 Spontaneous axonal regrowth is impaired in adults, however.
It is important to distinguish between the regeneration of injured axons and axonal sprouting from noninjured axons. 38 In this regard, it is important to highlight that in our model of complete transection injury, ascending and descending axons crossing the lesion are indicative of regenerative growth rather than sprouting, because there are no spared axons potentially sending collaterals. Although some sprouting is observed during spontaneous SCI recovery, sectioned axons fail to regrow after complete transection injuries without therapeutic intervention. This failure is proposed to have multiple causes: on one side, regenerative capacity is decreased in mature axons, and on the other side, the injured spinal cord is a hostile environment for axonal growth, with augmented presence of inhibitors and absence of facilitators. 39
A marked axonal regeneration was observed at the epicenter of the lesion in netrin-1 treated rats. This was reflected in an increased number, length, and sprouting of propiospinal axons. In addition, transected descending and ascending axonal tracts regenerated crossing the lesion site. In particular, our tracing experiments showed regenerating axons in the CST, which is the most important descendent projection in the control of voluntary movement in humans. 40 Finally our retransection experiments confirmed that the recovery of voluntary locomotion was dependent on axon regeneration at the lesion site and ruled out the possibility of a compensatory response in the spinal execution centers below the lesion site. 36,41
Axonal regeneration alone, however, is not sufficient to achieve the recovery of locomotor function observed in netrin-1 treated rats after SCI. Another important component in neuronal connectivity is the establishment and maintenance of synaptic connections. 42,43 Regain of function requires the reconnection of these regrown axons from the sensory-motor cortex with relay circuits at lumbar segments. This process entails the regeneration, sprouting, and reconnection of supraspinal and propiospinal axons with spinal neuronal targets. 44,45 Therefore, the cellular mechanism implicated in the locomotor recovery could involve axonal regeneration and concomitant reconnection with preserved or newly formed intraspinal relay circuits. 23,36,46
Our findings support this mechanism because we observed new transsynaptic interactions between regenerated descending axons and neurons localized at lumbar segments and regenerated ascending axons and second order neurons localized in gray matter upstream of the lesion site. The formation of these new connections could also be responsible for preventing transsynaptic degeneration that occurs as a consequence of the loss of inputs by motor neurons below the lesion. 47 –49
This assumption is supported by our findings that the surface of motor neurons at the lumbar segment from netrin-1 treated rats after SCI was surrounded by axons from the white matter area and pre-synaptic clusters of synapsin-1. Together with the formation of new connections between the spinal circuits and supraspinal centers, inputs from sensory afferents that remain intact below the lesion become an essential source of control in the production of motor behaviors after SCI. 36
The regenerative potential of mature axons in the central nervous system is still under debate. A very recent work by Poplawski and colleagues 17 shows that cortical neurons revert to an immature or regenerative transcriptional state after SCI. This regenerative state is defined by the upregulation of genes involved in cellular growth (cell protrusion, cytoskeleton dynamics, and sprouting), and the downregulation of growth inhibitory pathways.
Of particular interest in the context of the present study is the upregulation of the NTN1 gene and its receptor Unc5 in the late regenerative state (21 days after injury). Interaction between secreted netrin-1 and UNc5 allows the development of the correct trajectory of CST axons to reach the final targets at different segments of the spinal cord. 18 Mutations in this gene promote an aberrant projection of CST axons to the dorsolateral funiculus and motor alterations. 50 Interestingly although axons spontaneously transition to a regenerative embryonic-like state after SCI, 17 this is not enough to trigger full regeneration.
Moreover, recapitulations of dedifferentiated states are observed after SCI in the expression pattern of NFM in axons. There is an intent to recap, without success, the embryonic axon growth program. 51 The NFM labels are prominently localized on growing axons, being these neurofilaments are critical for caliber regulation and axonal cytoskeletal architecture. 52 In Netrin-1 treated rats, we observed axonal regrowth, with NFM expression. Therefore, we hypothesize that Netrin-1 treatment could successfully enhance the reactivation of the axonal growth transcriptional program.
Besides, we observed that netrin action is not limited to promoting or enabling axonal growth, but also to promoting tissue preservation/protection and minimizing tissue loss and the spread of initial tissue damage. Therefore, Netrin-1 administration provides a supportive growth-promoting environment to these axons. At anatomical and histological levels, netrin-1 treatment reduced the size of the lesion after SCI.
The increase in the size of the lesion after SCI is directly related to the secondary damage caused by the inflammatory response being enhanced by recruitment and guidance of proinflammatory macrophages to the injury site, 53 which is triggered by chemokine C-C motif ligand 2 (CCL2) secretion from astrocytes. 54 Macrophage activation and recruitment has dual and opposing effects in lesion development, improving tissue cleanliness on one side, but increasing the size of the injury on the other side. 3,55 –57
It is well documented, however, that netrin-1 prevents CCL2 secretion and macrophage activation. 58 This could explain the reduced number of macrophages observed in our netrin-1 treated animals. Preventing macrophage infiltration could be favorable for repair, not only by limiting tissue damage and loss, but also macrophages are known to contribute to axonal dieback. Long-distance retraction of injured axons coincides with the infiltration of macrophages, whose phenotypes transition from anti-inflammatory to proinflammatory in response to myelin debris. 59 Direct contact of macrophages with the dystrophic endings of insulted axons is considered to be essential to this process, 60 because the depletion of infiltrating macrophages reduces axonal dieback after SCI. 61
In line with the neuroprotective role of netrin-1, the treatment prevented axonal dieback from both ascending and descending axonal tracts. Axonal dieback results in the formation of nonregenerative axonal structures at the tip of axons, called retraction bulbs. In addition, it increases the gap that axons need to bridge to reach the other side of the lesion, and increases the chances of axons growing in inappropriate directions, posing an additional challenge to regeneration.
Conclusions
Netrin-1 has several and independent biological functions that could be redundant regarding promoting locomotor recovery. In summary, our findings provide proof of-concept evidence of axonal regeneration that can be achieved across a complete sectioned spinal cord with a concomitant neuronal target reconnection, below and above a lesion, promoting a full recovery of locomotor function previously lost by SCI. For this reason, netrin-1 treatment could be in the future a potential agent to restore locomotor function in humans with SCI.
Footnotes
Acknowledgments
I would like to thank to Dr. Horacio Romeo (BIOMED-UCA-CONICET) for the stereotaxic surgery equipment; Dr. Pablo Lopez (INIMEC-CONICET-UNC) for WGA tracer and GFAP antibody, technicians Ana Uceda and Alejandra Sgariglia (German Hospital) for assistance with animal care, Scott Lyman for critically reading the manuscript. I am grateful to Dr. Paola Bianchimano from The Mount Sinai Hospital for her helpful insights. This work is dedicated to the memory of Prof. Dr. Jorge Toblli.
Authors' Contributions
HRQ conceived, coordinated, and supported the study. HRQ designed and performed all experiments. HRQ performed the formal analysis. HRQ wrote, reviewed, and edited the manuscript.
Funding Information
This study was supported by a grant from Florencio Fiorini Foundation (FFF-2016) to HRQ and by personal funds of HRQ.
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Video S1
Supplementary Video S2
Supplementary Video S3
Supplementary Video S4
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.