
Abstract
Neuroimmune dysfunction is thought to promote the development of several acute and chronic complications in spinal cord injury (SCI) patients. Putative roles for adrenal stress hormones and catecholamines are increasingly being recognized, yet how these adversely affect peripheral tissue homeostasis and repair under SCI conditions remains elusive. Here, we investigated their influence in a mouse model of SCI with acquired neurogenic heterotopic ossification. We show that spinal cord lesions differentially influence muscular regeneration in a level-dependent manner and through a complex multi-step process that creates an osteopermissive environment within the first hours of injury. This cascade of events is shown to critically involve adrenergic signals and drive the acute release of the neuropeptide, substance P. Our findings generate new insights into the kinetics and processes that govern SCI-induced deregulations in skeletal muscle homeostasis and regeneration, thereby aiding the development of sequential therapeutic strategies that can prevent or attenuate neuromusculoskeletal complications in SCI patients.
Introduction
Long-term management of patients with traumatic spinal cord injury (SCI) is still considered a therapeutic challenge, mainly because of the broad range of secondary complications that can worsen the functional prognosis and/or alter quality of life. 1,2 One such complication is neurogenic heterotopic ossification (NHO), which are spontaneous formations of mature bone in the periarticular muscles of paralyzed joints; 3 NHO affects up to 20% of SCI patients. 4 –7 The pathological pathways that lead to NHO are still poorly understood, and hence it is unclear whether they are shared with various other known etiologies of heterotopic ossification (HO), 8 that is, genetic mutations, 9 non-neurological trauma, and surgery- and/or burn-induced lesions. 10,11
NHO is known to result from a multi-step process with gradual mineralization and bone maturation. 12 –14 The first clinical symptoms are typically noticed 3–8 weeks after the neurological insult when interstitial edema becomes large enough to create visible swelling, 15 –17 most frequently around the hip. 18,19 However, this soft-tissue infiltration has been related to earlier periarticular abnormalities, mainly increased vascularity that is detectable on bone imaging as early as 17 days after SCI. 20 Serial sections of mature HO specimens, collected through excision surgeries, confirm that the first stages in the histopathological sequence leading to HO formation are lymphocytic infiltration and structural changes of microvasculature. 21 Nevertheless, investigation of the preosseous phase has been hindered by both technical difficulties and ethical concerns around invasive muscle biopsies for this patient population, and the exact kinetics of the early phase of NHO development therefore remain mostly undefined.
One of the current views is that pro-osteogenic factors (such as bone morphogenetic proteins, transforming growth factor-β1, and/or substance P), deemed to be released into the systemic circulation from damaged neurons, 22,23 may activate progenitor cells with osteogenic potential inside affected muscles. 24 The local drivers of this activation are still not understood, and whether or not they are related to traumatic myofibrillar lesions in patients is also unknown. Although acquired HO is commonly conceptualized as a pathological tissue-repair process, 25,26 there is currently no clinical evidence that this also applies to neurogenic HO.
In this study, we aimed to elucidate both when and how a central neurological insult negatively interferes with the muscular regeneration process that is induced by cardiotoxin (CDTX) injection. Our previous work already reported the histological aspect of NHO in C57BL/6J mice, which is characterized by the presence of scattered mineralized necrotic materials demarcated by osteoid matrix deposits between regenerating myofibers. 27 We have also highlighted an exacerbated local innate immune response, uncovering a key role for macrophages and oncostatin M in the ectopic formation of calcified bone-like nodules within affected limbs. 28 –30 What has remained unclear, however, is how and/or why the coexistence of an SCI derails the muscle regeneration process and causes NHO. We hypothesized that the priming effect of SCI takes place within the first hours after injury through the acute dysregulation of neuroimmune and hormonal systems, either synergistically or sequentially. To test this, we first studied how the timing of SCI and CDTX relate to each other in order for NHO to develop. Next, we explored the different local and systemic neurogenic and/or hormonal signals that are triggered by the neurological injury to assess and describe their roles in NHO formation.
Methods
Mice and housing
Adult (5–8 weeks of age) female C57BL/6J mice were obtained from the Animal Resources Centre (Perth, Western Australia, Australia). All mice were allowed to adjust to their new environments for at least 1 week before surgery. They were housed (≤5 per cage) in ventilated microisolator cages, layered with paper bedding, on a 12-h light/dark cycle at a constant temperature (20°C ± 2°C) and humidity (50% ± 20%), with unlimited access to food pellets and water. All experimental procedures were approved locally by the animal ethics committees of The University of Queensland (#050/17 and #449/18) and conducted in accordance with the Australian Code for the Care and Use of Animals for Scientific Purposes, as well as the ARRIVE (Animal Research: Reporting of In Vivo Experiments) guidelines, 31 with regard to randomization (order of surgery, treatment allocation), blinding (surgery and analysis), and reporting.
Spinal cord injury and animal care
Complete spinal cord transection at either the T2 or T9 level was performed as previously described, 32 through a posterior approach of the spine after paraspinal muscle dissection. With gentle traction, the interlaminar space was identified and dissected to expose the cord. Transection was conducted using a microblade and hemostasis obtained through gentle compression with a cotton swab. A similar surgical approach was followed for T9 spinal cord contusion injuries, but with the addition of a laminectomy. SCI was inflicted using the Infinite Horizon impactor device, delivering a 100-kilodyne force directly onto the exposed spinal cord. 33 The surgical site was then closed, followed immediately by intramuscular injection of CDTX (TXL8102, 0.26 mg/mL in sterile phosphate-buffered saline [PBS], 75 μL per hindlimb; Latoxan, Portes lès Valence, France).
For this, the lower hindlimb was shaved and the skin gently scrubbed with betadine. With the mice placed prone, the hindlimb was held in an extended position between the thumb and index finger. Next, a 0.5-mL ultrafine syringe (30-gauge needle; Becton Dickinson, Franklin Lakes, NJ), loaded with CDTX solution, was inserted mid-hamstring using a posterior approach and with an ∼45-degree angle to the horizontal plane. Correct intramuscular placement was confirmed through firm resistance when gently pulling back the syringe plunger; great care was taken at all times to not injure and/or create contact with the femoral periosteum. The CDTX solution was then slowly injected, with perception of an immediate swelling of the injected muscle confirming correct placement. Variations between the timing of injection and SCI were applied, where relevant, as per experimental design (see Fig. 1A).
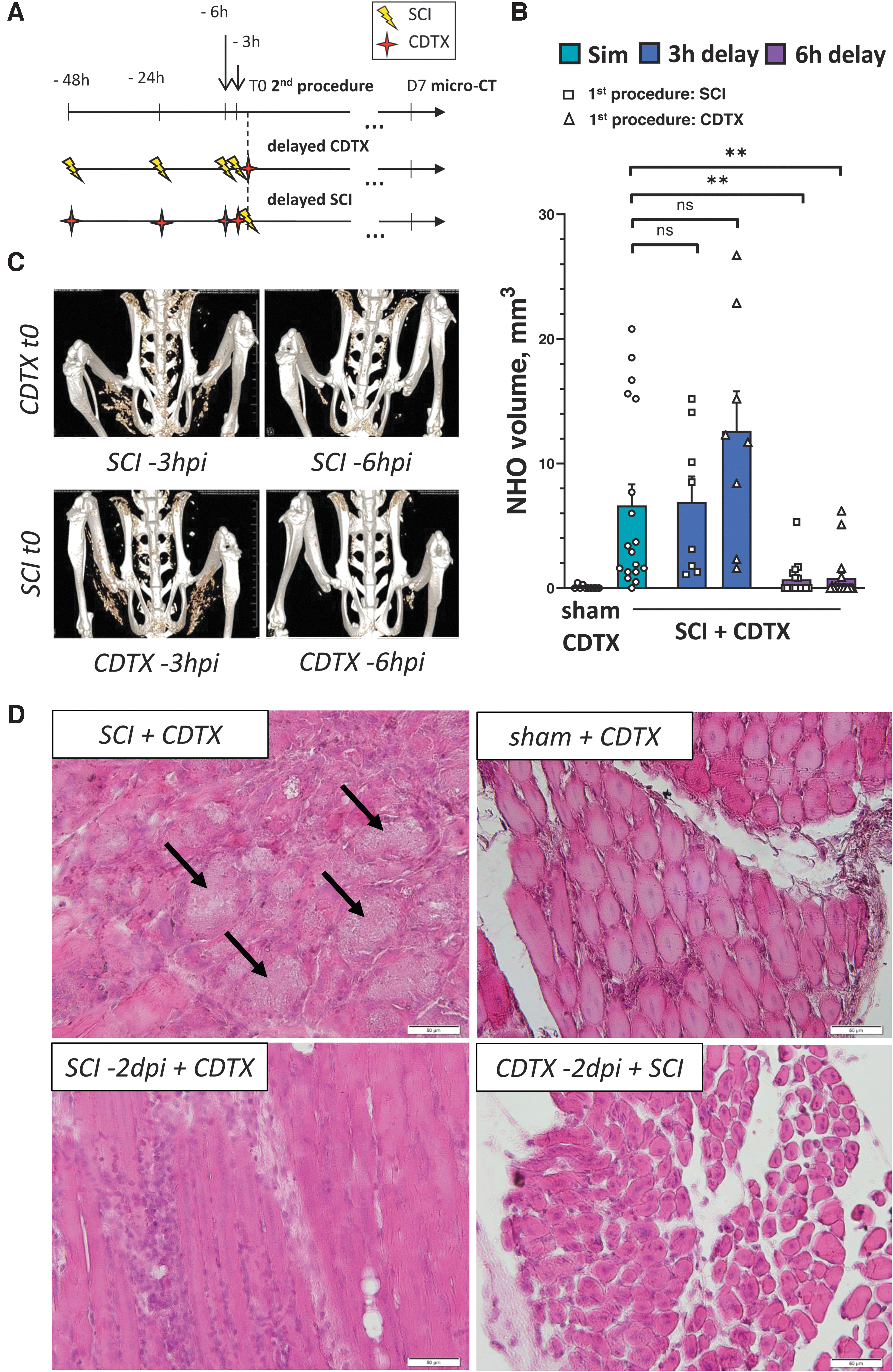
NHO formation in CDTX-injected SCI mice requires a synergy between the coordinated effects elicited by spinal cord transection and muscular injury. (
Mice received a single injection of buprenorphine (0.5 mg/kg subcutaneous) in Hartmann's solution (Baxter) for analgesia post-surgery and were kept overnight in a recovery chamber at 30°C and ∼40% humidity. Sham animals received a surgical dissection of paraspinal muscles and posterior ligaments, but without SCI. Mice with transection injuries were also given 125 mg/L of ciprofloxacin in their drinking water as a prophylactic treatment throughout the course of the experiment to prevent bladder infections. Post-operative care otherwise included manual bladder expression twice a day, which was combined with monitoring of body weight, motor function, and clinical signs for pain or distress until the experimental end-point.
Adrenalectomy and vascular filling
Mice were anesthetized by intraperitoneal (i.p.) injection of xylazil and zoletil (20 and 50 mg/kg, respectively). A bilateral retroperitoneal approach was used to identify the adrenal glands at the cranial end of the kidney, and these were then enclosed with ring forceps to obtain vessel hemostasis and excised. Abdominal wall incisions were closed with a 5.0 absorbable suture and sterile wound clips for the skin. Sham-operated mice were subjected to adrenal-gland exposure and surgical-site closure. Seven days post-adrenalectomy, all mice underwent SCI and a CDTX injection. To minimize the risk of hemodynamic shock and lethal hydroelectrolyte abnormalities in adrenalectomized mice, intravenous injections of Gelofusine (100 microliters per mouse) were administered 1 h before SCI as previously described. 34 Post-operative monitoring and care included daily surveillance for clinical signs of dehydration, i.p. injection of Hartmann's solution, and salt supplementation by the drinking water (1% NaCl). 35
Pharmacological treatments and chemical sympathectomy
(±) Propranolol hydrochloride (P0884, 10 mg/kg in sterile water; Sigma-Aldrich, St. Louis, MO) was injected i.p. The first dose was administered 2 days before SCI surgery, and treatment was continued throughout the experimental time point until euthanization and μCT evaluation at 7 days post-injury (dpi); treatment controls received matching volumes of sterile water only.
Substance P receptor blocker L-733,060 hydrochloride (1145; Tocris Bioscience, Bristol, UK) was dissolved into PBS and injected i.p. (10 mg/kg) 30 min before surgery and then again at 3 h after surgery. Control animals received matching volumes of PBS.
For chemical sympathectomy, the neurotoxin, 6-hydroxydopamine (6-OHDA; H4381; Sigma-Aldrich), was administered by i.p. injection at both 4 (100 mg/kg) and 2 days (250 mg/kg) before surgery to selectively kill peripheral noradrenergic neurons 36 ; controls received vehicle injection only. All mice were then subjected to spinal cord transection and CDTX injection as per the standard protocol, followed by euthanization and μCT evaluation at 7 dpi.
Micro–computed tomography and quantification of neurogenic heterotopic ossification volumes (bone volumes)
An Inveon pre-clinical micro–computed tomography (μCT) scanner (Siemens, Erlangen, Germany) was used for post-mortem sample analysis. CT images were acquired through an X-ray source with the voltage set to 80 kV and the current set to 250 μA. All scans were performed using 360-degree rotation (1-degree steps), with a medium-high magnification and a binning factor of 2; the exposure time was 2000 ms. Projection images were reconstructed using Inveon Acquisition Workstation software (IAW version 2.4; Siemens), using a three-dimensional (3D) Feldkamp algorithm with an isotropic voxel dimension of 27 μm. 3D images of the lower parts of the mouse skeleton were reconstructed from the scans using the μCT system software package. Quantitative assessments of ectopic mineralized tissue volumes were performed through manual definition of the region of interest, excluding normal bone. Identical thresholding values were applied for all analyzed scans.
Histology
At 7 days post-surgery, mice were euthanized by CO2 asphyxiation. Hindlimbs were fixed in PBS with 4% paraformaldehyde, decalcified in 5% ethylenediaminetetraacetic acid, and embedded in paraffin. Next, 5-μm sections were cut (Leica RM2245; Leica Biosystems, Buffalo Grove, IL), deparaffinized, and then rehydrated for staining with hematoxylin and eosin. Slides were viewed using an Olympus BX50 microscope with an attached DP26 camera and imaged using Olympus CellSens standard 1.7 imaging software (Olympus Corporation, Tokyo, Japan).
Enzyme-linked immunosorbent assays
To quantify plasmatic catecholamines and corticosterone, blood samples were harvested in heparinized tubes by cardiac puncture under isoflurane anesthesia at 3, 6, and 24 h after surgery using exactly the same conditions for all groups to minimize variation; naïve controls were used for baseline measurements. Blood samples were centrifuged twice at 1000g at 4°C for 10 min. Blood plasma was then harvested and stored at −80°C until further processing.
To assess the impact of SCI on circulating substance P levels, blood samples were collected through retro-orbital eye bleeds from different cohorts of mice under brief methoxyflurane anesthesia at 45 and 90 min as well as 6 and 24 h after surgery. Blood samples were centrifuged, processed, and stored as described previously. Commercially available enzyme-linked immunosorbent assay kits were used as per the manufacturer's instructions to measure norepinephrine/epinephrine (KA1877; Abnova Corporation, Taipei City, Taiwan), corticosterone (KA6129; Abnova), and substance P (MBS009083) in each sample.
Messenger RNA extraction for reverse-transcriptase polymerase chain reaction analysis in total muscle
Hindlimb muscles were harvested from euthanized mice at the specified time points after surgery and snap-frozen in liquid nitrogen. Frozen samples were homogenized using a TissueRuptor (Qiagen, Hilden, Germany) directly in TRIzol (Life Technologies, Carlsbad, CA) and RNA isolated from the aqueous phase after chloroform separation. Reverse transcription for complementary DNA (cDNA) generation was performed using the SensiFAST™ cDNA Synthesis kit (Bioline, London, UK) as per the manufacturer's instructions. Analysis of mRNA expression was carried out using the Taqman Fast Advanced Master Mix and primer/probe sets (ThermoFisherScientific, Waltham, MA): Adrb1 (Mm00431701_s1), Adrb2 (Mm02524224_s1), and RUNX family transcription factor 2 (Runx2; Mm00501584_m1) on a ViiA 7 Real-Time PCR System (Life Technologies), using 20 sec at 95°C, then 40 cycles of 95°C (1 sec) and 60°C (20 sec). Results were normalized relative to β-2 microglobulin and glyceraldehyde-3-phosphate dehydrogenase mRNA expression.
Statistical analyses
GraphPad Prism software (GraphPad Software Inc., La Jolla, CA) was used for data visualization and statistical analyses. A two-sided Mann-Whitney U test was used to detect differences in bone volumes (BVs) and protein concentrations between experimental conditions. One-way analysis of variance (ANOVA) with Tukey's multiple comparison test or non-parametric Kruskal-Wallis with Dunn's multiple comparison test were used to detect differences between groups, except for reverse-transcriptase polymerase chain reaction (RT-qPCR) experiments where ANOVA with Dunnett's multiple comparisons test was used on delta threshold cycle values. All data sets are presented as specified in the main text and/or figure legends. Appropriate sample sizes were determined/confirmed by both a priori and post hoc power analysis, with the level of power (1 – β) set at 0.80 and α < 0.05.
Results
Neurogenic heterotopic ossification formation in mice results from a synergy of the coordinated effects elicited by spinal cord injury and muscular injury
In the original mouse model, NHO was inducible with full penetrance when intramuscular CDTX injection was combined during the same procedure with a transection of the lower thoracic (T9) spinal cord. 28 Intramuscular injection of CDTX itself is a well-known experimental and commonly used method to study muscular regeneration. Through direct contact, the toxin induces a cascade of events, from selective myofiber fragmentation and recruitment of leukocytes within hours of treatment, to the progressive transformation of myotubes into myofibers after several days as the damaged muscle regenerates. Although it is clear that the presence of a central neurological insult can derail the muscular regeneration program into ectopic osteogenesis, the pathological underpinnings driving this effect are still unclear. In our previous work, we already showed that NHO formation does no longer occur if CDTX is injected 10 days after a complete transection of the spinal cord. 27 We therefore hypothesized that the SCI effect was temporary and first sought to better define the kinetics of its interaction with the muscular regeneration process.
To determine by how much either the muscular or neurological injury could be delayed while still triggering NHO formation, C57BL6/J mice underwent a two-stage surgery, starting with either a mid-thoracic spinal cord section or CDTX injection in both hamstrings, followed by the second procedure (i.e., CDTX injection for SCI mice or vice versa) 3, 6, 18, 24, or 48 h later (Fig. 1A); the time point of the second procedure was always defined as t0 and HO formation quantitatively assessed 7 days later using in vivo micro-CT imagery. We found that a delay of 3 h between injuries yielded comparable NHO BVs relative to controls (i.e., mice where both injuries were combined in one procedure [Sim]), irrespective of whether either the SCI or CDTX injection was conducted first (6.90 ± 5.84 mm3 [mean BV ± SD] in SCI-3hpi + CDTXt0 [n = 8] and 12.64 ± 8.92 mm3 in CDTX-3hpi + SCIt0 [n = 8] vs. 6.64 ± 7.18 mm3 in SCI t0 + CDTXt0 controls [n = 18]; p > 0.05; Fig. 1B,C).
However, introducing longer delays (up to 6 h) between procedures resulted in a significant reduction (>6-fold) in average NHO volume compared to controls where both injuries were inflicted simultaneously (0.71 ± 1.36 mm3 [average BV ± SD] in SCI-6hpi + CDTXt0 [n = 16] and 0.79 ± 1.82 mm3 in CDTX-6hpi + SCIt0 [n = 18]; p < 0.0029). A further delay of either injury to 18, 24, or 48 h resulted in a complete absence of NHO formation (Fig. 1D and data not shown).
Taken together, these results demonstrate that muscle and spinal cord injuries must take place within a 3-h time window of each other to trigger NHO formation, and hence that the initiating effect of SCI in our model is very acute and temporary. Given that this effect requires one to be precisely coordinated with the muscular regeneration process in order to obtain osteogenic conditions, it likely specifically targets a recruited and/or activated cellular subtype in the regenerating microenvironment. Considering the short time window that was identified, we next explored both endocrine and neurogenic mechanisms as possible initiating factors of NHO because of the transitory, but powerful, signals that they produce in response to stressful stimuli. 37,38
Lesion-level–dependent changes in catecholamines influence neurogenic heterotopic ossification bone volumes in spinal cord injury mice
Stress-induced signals, in particular catecholamines, were first examined as possible candidates mediating the acute transient effect of SCI on NHO formation. Catecholamines are normally released upon activation of the sympathetic nervous system and play vital roles in maintaining cardiovascular and metabolic homeostasis. 39 Consequently, acute autonomic dysfunction in spinal-cord–injured patients can lead to neurogenic shock within 2 h post-trauma. 40,41 Catecholamines are also known to be potent modulators of innate and adaptative immune cells. 42,43 This includes strong anti-inflammatory effects in macrophages through beta-2 adrenergic receptor (ADRB2), 44 that is, the very cells that are known to play a key role in NHO formation. 28 We therefore hypothesized that abnormal systemic levels of catecholamines after SCI may alter the function of immune cells recruited into the injured muscle and hence derail the regeneration process. To directly test this, we combined CDTX injection as per the originate model with a high-level thoracic (T2) spinal cord transection to decentralize the adrenal gland.
Quantification of epinephrine, norepinephrine, and corticosterone 24 h after surgery confirmed a dramatic reduction of circulating catecholamine levels in T2 SCI mice compared to both their T9 counterparts and naïve controls (Fig. 2A). 32 We further observed significantly elevated epinephrine levels in the circulation of T9 SCI mice with CDTX injection that were not present at earlier time points (3 and 6 hours post-injury; see Supplementary Fig. S1). The corticosterone response was otherwise found to be more acute, with significantly increased levels already being observed in both T2 and T9 SCI mice, as early as 3 h after injury (Supplementary Fig. S1). In contrast to catecholamines, no lesion-level–dependent differences in corticosterone concentrations were observed at the peak (24 h post-injury) between T2 and T9 SCI mice (Fig. 2A).

Catecholamines influence NHO bone volumes in SCI mice. (
With respect to NHO, bone volumes in T2 SCI mice with CDTX injection were significantly decreased compared to their T9 counterparts (mean BV ± SD: 1.2 ± 1.4 vs. 9.7 ± 3.7 mm3, respectively; p = 0.0012) at both 7 and 21 days post-injury (Fig. 2B and data not shown). A key role for adrenergic signals was further corroborated by performing bilateral adrenalectomies 1 week before our standard procedure (i.e., T9 SCI with CDTX injection). We confirmed that catecholamines and corticosterone were undetectable in plasma of adrenalectomized mice (data not shown). Despite the setup of a vascular filling protocol, we experienced a 66% mortality rate after the SCI procedure in adrenalectomized mice, and only 3 (out of n = 9) survived until 7 dpi. Nevertheless, none of these mice developed NHO (Fig. 2C).
To assess whether acute adrenergic signaling influenced NHO formation, we performed a chemical sympathectomy by injecting the catecholaminergic neurotoxin, 6-OHDA, 4 and 2 days before SCI surgery. This procedure effectively depletes adrenergic nerves and chromaffin cells in the adrenal medulla. 36,45,46 Chemical sympathectomy before SCI did indeed reduce NHO volumes 2.4-fold (Fig. 2D).
We next evaluated the expression of β-adrenergic receptors in our model. β2-adrenoreceptors (ADRB2) are the predominant adrenoceptor (sub)type expressed in skeletal muscle (90%), and they have also been previously identified as regulators of muscle regeneration after myotoxic injury. 47 Yet putative roles for other β-adrenergic receptor subtypes and/or isoforms remain unclear. RT-qPCR analysis revealed a 10- to 50-fold increase in Adrb1 mRNA expression in CDTX-injected muscles of T9 and T2 SCI mice at 1 dpi (p < 0.002; Fig. 2E). Expression of Adrb2 was trending higher (∼6-fold) in T2 SCI mice with CDTX injection, but this did not reach statistical significance compared to other groups (p > 0.05); no changes in Adrb1 (and Adrb2) expression were observed at earlier time points (Supplementary Fig. S2A). Given these observed patterns of expression, and to directly confirm a role for catecholamines in our model, we next evaluated the effect of the non-selective β1/β2 antagonist, propranolol, on ectopic bone formation in CDTX-injected T9 SCI mice.
Consistent with our earlier findings in both T2 SCI and adrenalectomized T9 SCI mice, presenting with profoundly reduced and undetectable blood catecholamine levels, respectively, micro-CT evaluation at 7 dpi showed a significant reduction in NHO volumes in propranolol-treated mice (2.7 ± 2.5 [average BV ± SD] vs. 5.6 ± 3.5 mm3 in vehicle-treated controls; p = 0.0136), and to a level that was similar to that observed in T2 SCI mice (Fig. 2F).
To assess whether the observed effects of adrenergic signaling could be caused by reduced activity and/or differentiation of osteogenic progenitors, we quantified Runx2 mRNA expression (a marker of early osteogenic differentiation) in CDTX-injected muscles of both T2 and T9 SCI mice. We found that Runx2 mRNA expression was significantly upregulated (∼20-fold; p < 0.0001) in CDTX-injected muscles of SCI mice, with no influence of lesion level (Fig. 2G). Taken together, these results indicate that adrenergic signaling contributes to the multi-step process leading to ectopic osteogenesis and NHO formation after SCI, most likely through activation of ADRB1 in injured muscle. However, the window during which adrenergic signals most likely do so (i.e., at ∼1 dpi) is outside of the critical time period during which SCI presence can derail the muscle regeneration process. Cell-fate decisions toward osteogenic differentiation thus are taken before and/or not influenced by lesion-level–dependent differences in catecholamine production/release.
Neurogenic heterotopic ossification forming mice display an immediate systemic release of substance P after injury
Having determined that level- and/or condition-dependent differences in the neuroendocrine stress response modulating NHO volumes occurred mostly past the established critical time window, we next aimed to identify earlier triggers that could be responsible for initiating NHO. Substance P (SP) appeared to be a relevant candidate, considering that it is released both peripherally from nociceptive nerve terminals under inflammatory conditions and also in the dorsal horn cells of the injured spinal cord within hours of insult. 48 Consistent with this, we indeed found that substance P levels in plasma already peaked between 45 and 90 min post-injury (mpi) and, importantly, in NHO-forming mice only (Fig. 3A). We corroborated this result in the contusion injury model, which better mimics the human condition (Fig. 3B). An ∼3-fold increase in SP plasma levels was found at 90 mpi in mice with a contusive SCI compared to those with a spinal cord transection (respective means ± SD: 1012 ± 230 vs. 295 ± 24 pg/mL). We then used our standardized micro-CT protocol to directly compare NHO formation across both models of SCI, revealing significantly greater BVs in contusion SCI mice compared to those with spinal cord transection (respective means ± SD: 31 ± 6 vs. 11 ± 9 mm3; p = 0.0159; Fig. 3C).
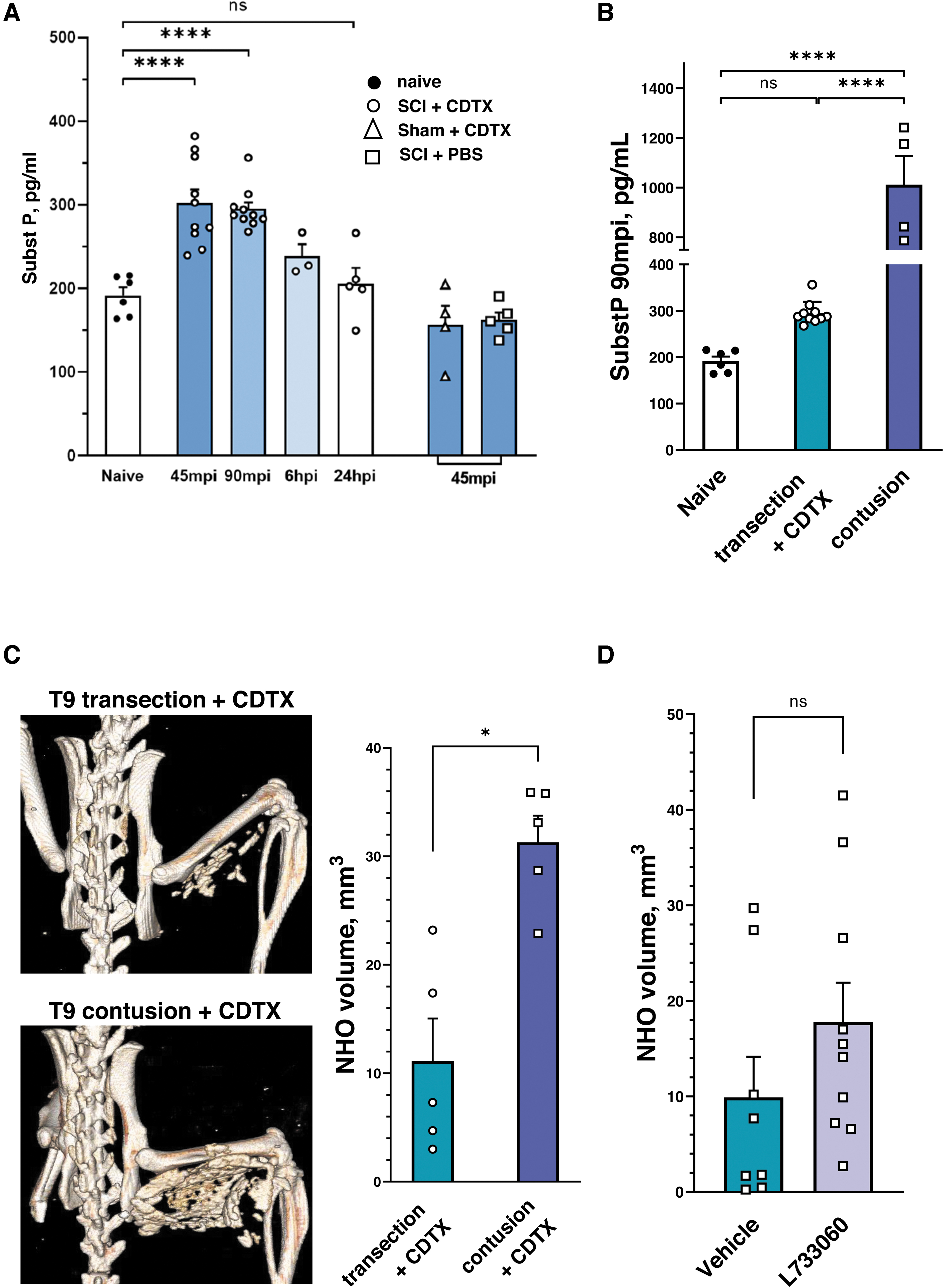
Acute substance P release in NHO-forming mice correlates with ectopic mineralized tissue volumes. (
Taken together, these results confirm a positive correlation between acute systemic release of SP after SCI and the average volume of NHO that is formed in CDTX-injected muscles. Guided by these findings, we last explored whether increased substance P signaling could be the initial step in NHO formation. For this, mice were administered the neurokinin-1 receptor (NK-1R) antagonist, (+)-LP 733,060, during the first 3 h after T9 SCI and CDTX injection. No differences were found, however, in NHO volumes between treated mice and controls (respective mean ± SD: 17.7 ± 13.0 vs. 9.9 ± 12.0 mm3; p = 0.1457; Fig. 3D), suggesting that substance P is either not the early trigger of ectopic bone formation or, alternatively, that it must act through mechanisms that are independent of NK-1R.
Discussion
Enhanced systemic stress and peripheral changes in immune function are thought to drive the onset of a range of acute and chronic complications in SCI patients. 49 –51 However, whether such SCI-associated factors also play a role in the aberrant regulation of cell-fate decisions leading to NHO remains to be clarified. In this study, we demonstrate for these effects in peripheral tissues to be polymodal and sequential. Indeed, our results reveal that the combination of low-level (T9) SCI and muscle injury in C57BL/6 mice triggers a significant release of circulating epinephrine at 1 dpi. This increase in catecholamine levels influences NHO volumes, as illustrated by the inhibitory effects of propranolol and chemical sympathectomy on NHO volumes. The catecholamine effect appears secondary, however, given that it does not drive the initiation of an osteogenesis program in muscle cells. We corroborated this further by showing that the local initiation of an osteogenic program critically requires both muscle injury and SCI to be coordinated within <6 h of each other. The unsuspected transience and rapid onset of this synergistic effect is correlated with increased SP levels in NHO-forming mice between 45 and 90 min after injuries.
Overall, these novel findings demonstrate that ectopic osteogenesis is initiated acutely after SCI and then secondarily regulated by inter-related and time-dependent phases, involving multiple and potentially redundant pathways. By providing a renewed focus on the specific features that characterize neurogenic HO in mice, we also emphasize that the cellular mechanisms driving ectopic osteogenesis in pre-clinical models are likely heterogenous. 8
One of the complexities in understanding ectopic osteogenesis in the NHO mouse model is that both SCI and CDTX injection individually induce generic local and systemic responses to tissue injury. 52 –54 This makes it challenging to dissect the specific factor(s) that drive and/or derail the muscular regeneration process toward ectopic osteogenesis under SCI conditions. Previous work already highlighted a decisive role for immune signals, in that depletion of monocytes/macrophages with clodronate-loaded liposomes dramatically reduced NHO volumes. 28 Whether and how SCI affects the function of infiltrating and/or resident phagocytic cells in NHO remains to be clarified. However, it is known that normal functioning of the immune system relies, in part, on a working sympathetic-neuroendocrine adrenal axis. 32
Consistent with both this and the previously established role for immune cells in ectopic osteogenesis, 28 we show that NHO formation, like other SCI sequalae, is also influenced by adrenergic signaling. Specifically, we found reduced NHO volumes in T2 SCI mice compared to their T9 counterparts, that is, those that either lack or have brain control over sympathetic outflow to the adrenal gland, respectively. We corroborated this by confirming that the levels of circulating catecholamines at 1 dpi in T2 SCI mice were far below those of naïve controls, 32 suggesting an impaired release by the decentralized adrenal glands and/or increased excretion, as has been reported for human SCI patients. 55 It is otherwise worth noting that both spinal cord transection models lead to either complete (T2) or severe (T9) damage to brain pathways that provide input into sympathetic pre-ganglionic neurons innervating the hindlimb, 56 yet only T9 SCI spurs ectopic bone formation. A systemic, rather than local, change in catecholamine levels therefore appears to be the likely adrenergic driver of NHO development/progression.
In contrast to catecholamines, glucocorticoid levels were not different between T2 and T9 SCI mice, indicating a similar stress response through the hypothalamic/pituitary/adrenal pathway irrespective of lesion level. A causal role for catecholamines in NHO formation after SCI was otherwise directly demonstrated through antagonism of adrenergic signaling, in that beta-blocker treatment in T9 SCI mice yielded a similarly negative regulation of ectopic osteogenesis compared to that observed in T2 SCI mice.
Given that lesion-level–dependent changes in circulating epinephrine were only detected at 24 h post-surgery and did not modify the abnormal expression of osteogenic marker Runx2, they are unlikely responsible for the initiation of an osteogenic program within injured muscle. The modulating action(s) of catecholamines on NHO volumes are therefore more likely mediated by an altered functionality of resident and/or recruited immune cells within the injured muscles of SCI mice. The ability of myeloid cell populations to misguide progenitor cell differentiation during aberrant tissue repair is indeed increasingly recognized, 57,58 and the influence of catecholamines on immune cell activation, proliferation, and/or apoptosis is also well established. 44
Consistent with this, both innate and adaptive immune cells are known to express various adrenergic receptors. ADRB2 has been the most extensively studied receptor subtype to date, and the evidence shows that it mediates macrophage polarization and cytokine production. 59 Beta-adrenergic signaling also contributes to macrophage migration and phagocytic ability. 60 Upregulation of ADRB1 within muscles of NHO-forming mice, as was observed here at 1 dpi, is particularly intriguing because it specifically arose from the cumulative effects of SCI- and CDTX-induced muscle injury. Being the most marginally expressed ADRB isoform in both myocytes and immune cells, its specific functions in muscle homeostasis and repair are yet to be clarified. 61 Further studies on ADRB1 are therefore warranted, both in relation to a putative role in NHO formation and also as to what signals drive its upregulation under the circumstances where SCI and muscle injury are combined within 6 h of each other.
The critical time window during which SCI and muscle injury must temporally coincide in order for ectopic osteogenesis to occur demonstrates, for the first time, that the priming effect of SCI in relation to NHO is not permanent, at least not in this model. It also shows that, as a comorbidity, SCI itself is only capable of derailing the muscular regeneration process early after CDTX injection, thereby making it important to understand the actions of this toxin. Cardiotoxin γ is a protein kinase C inhibitor that leads to pore formation in plasma and mitochondrial membranes, cell swelling, and fragmentation without affecting the extracellular matrix. 62 Although its precise mode of action remains incompletely understood, 63 it reportedly affects membrane calcium-binding sites and/or calcium release. 64 Whereas any such deregulation of Ca2+ homeostasis in itself is not sufficient to cause NHO in compromised myofibers, it may very well contribute to the progressive mineralization of myonecrotic debris within injured muscles under SCI conditions, 27 that is, where the effects of CDTX, SCI-induced neuropeptide release and metabolic changes are combined.
In the present study, we identified an early blood release of substance P after SCI that occurs within the established critical time window. Moreover, we found that this SP peak was 3 times higher and positively correlated with the significantly increased NHO volumes in contusion SCI mice. Beyond its well-established role in regulating neurogenic inflammation 65,66 and inducing HO in mice, 67 substance P can also trigger mitochondrial reactive oxygen species (ROS) production. 68,69 This finding is of interest given that the functioning of Ca2+ channels in skeletal muscle is modulated by mitochondrial respiration. 70 Although we were not able to block the acute/immediate initiating step(s) leading to ectopic osteogenesis through short-term NK-1 antagonism within the first 3 h post-injury, there is substantial evidence that substance P can also act through other receptors/mechanisms. 71,72 NK-1-independent roles for substance P could therefore be explored further in future experiments. Central nervous system lesions are otherwise also known to cause a shift in muscle fiber phenotype, 73,74 where mitochondria-dense slow and fatigue-resistance oxidative fibers convert to a fast fatigable glycolytic type with increased expression of fast sarco(endo)plasmic reticulum Ca2+-ATPase (SERCA1). 75
Increased sensitization of the mitochondrial permeability transition pore has otherwise also been linked to decreased oxidative capacity in the SCI population 76 and to deregulated Ca2+ efflux mechanisms. 77 These changes, alone or in combination, may further contribute to metabolic decline and aberrant release of Ca2+ in skeletal muscle after SCI. 78 In summary, how SCI-induced neuropeptide release, metabolic, and/or structural changes contribute to the CDTX effect in our mouse model of NHO warrants further investigation. We postulate that the disruption of calcium sequestration in injured and denervated myofibers is one of the immediate/acute initiating and decisive steps in ectopic osteogenesis under SCI conditions. This then sets the scene for impaired debris clearance and the persistence of necrotic calcified nodules that trigger an osteo-inductive microenvironment in which deregulated immune cells can secondarily promote and/or further drive NHO formation (Fig. 4).
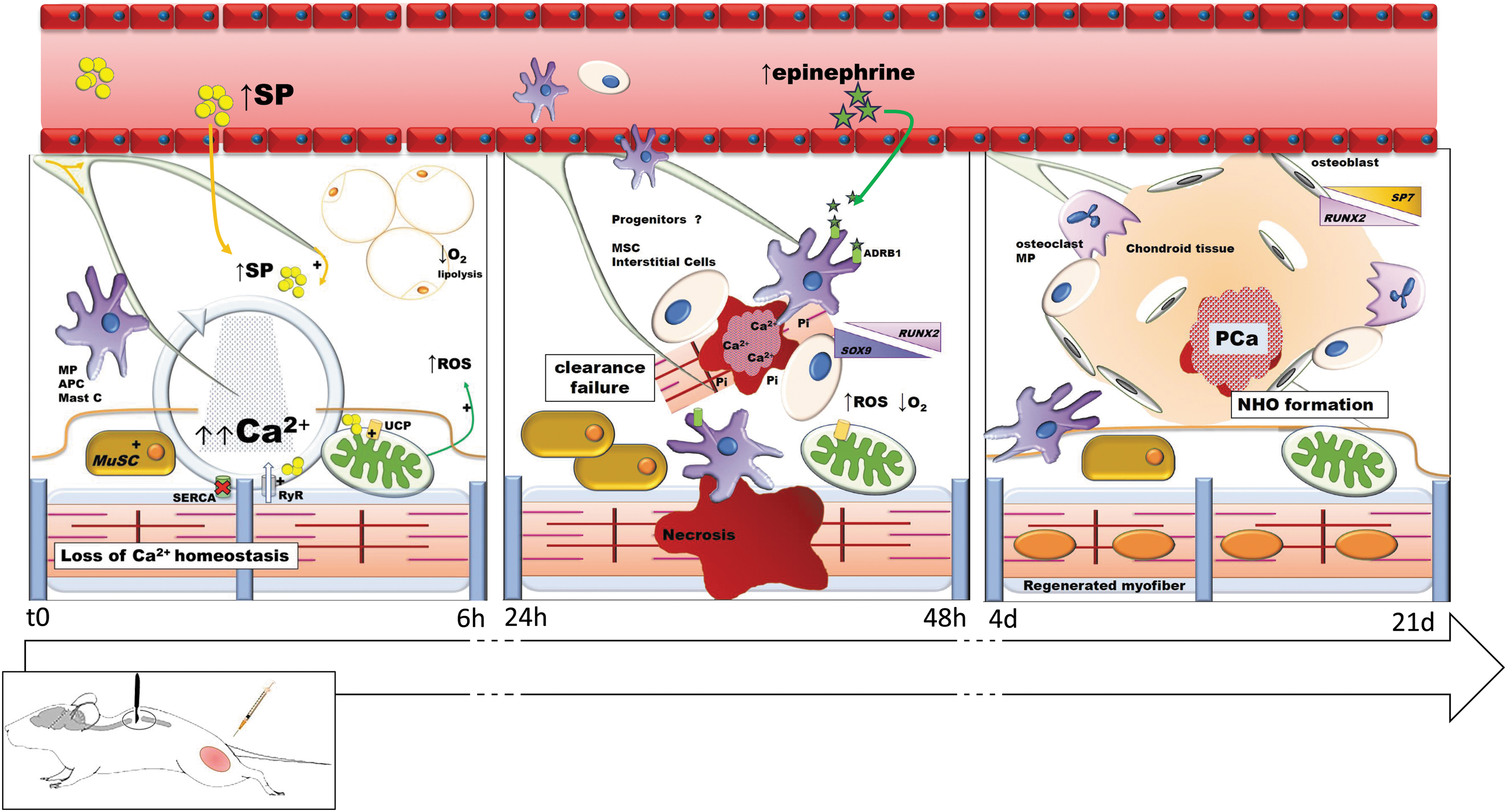
Graphical representation of sequenced SCI-induced dysregulations impairing muscular regeneration and leading to NHO formation in mice. MP, macrophage; APC, antigen-presenting cell; Mast C, mast cells; SP, substance P; ROS, reactive oxygen species; MuSC, muscle stem cells ; SERCA, sarco/endoplasmic reticulum Ca2+-ATPase; RyR, ryanodine receptors; MSC, mesenchymal stem cells; Pi, inorganic phosphate; PCa, calcium phosphate; NHO, neurogenic heterotopic ossification; SCI, spinal cord injury. Color image is available online.
Overall, this work highlights the multitude of physiological changes that occur as a result of SCI and how these can sequentially influence peripheral tissue homeostasis, including in the context of muscle injury and repair. By elucidating the temporal kinetics and interconnections between the various factors and/or priming events that occur after SCI and can drive NHO, we unlock new avenues for further research as well as the development of specifically timed combination therapies to better manage acute SCI and reduce the incidence of secondary complications in at-risk patients.
Footnotes
Acknowledgments
The authors acknowledge the assistance of UQBR staff with animal husbandry and sample collection, the Preclinical Imaging Facility at the Translational Research Institute, which is supported by Therapeutic Innovation Australia (TIA), an Australian Government Initiative through the National Collaborative Research Infrastructure Strategy (NCRIS) Program, as well as the facilities, scientific, and technical assistance of the National Imaging Facility, a National Collaborative Research Infrastructure Strategy (NCRIS) capability, at The University of Queensland's Centre for Advanced Imaging.
Authors' Contributions
C.D., F.G., M.J.R., and J.P.L. conceived the study and designed experiments. C.D., H.W.T., M.C., I.K., and M.J.R. performed experiments and/or assisted with data acquisition. C.D. and M.J.R. wrote the manuscript. All authors contributed to data interpretation, provided critical feedback and revisions, and also approved of the final manuscript.
Funding Information
Experiments were supported, in part, by Ideas Grant 1181053 (to J.P.L., H.W.T., and F.G.) from the Australian National Health and Medical Research Council (NHMRC) and by the Mater Foundation. All work conducted in the laboratory of M.J.R. was made possible by SpinalCure Australia. J.P.L. was supported by Research Fellowship 1136130 from the NHMRC, and C.D. by a Research Fellowship from L'Institut Servier (France).
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.