
Abstract
Incomplete spinal cord injury (SCI) may result in muscle weakness and difficulties with force gradation. Although these impairments arise from the injury and subsequent changes at spinal levels, changes have also been demonstrated in the brain. Blood-oxygen-level dependent (BOLD) imaging was used to investigate these changes in brain activation in the context of unimanual contractions with the first dorsal interosseous muscle. BOLD- and force data were obtained in 19 individuals with SCI (AISA Impairment Scale [AIS] C/D, level C4–C8) and 24 able-bodied controls during maximal voluntary contractions (MVCs). To assess force modulation, participants performed 12 submaximal contractions with each hand (at 10, 30, 50, and 70% MVC) by matching their force level to a visual target. MVCs were weaker in the SCI group (both hands p < 0.001), but BOLD activation did not differ between SCI and control groups. For the submaximal contractions, force (as %MVC) was similar across groups. However, SCI participants showed increased activity of the ipsilateral motor cortex and contralateral cerebellum across all contractions, with no differential effect of force level. Activity of ipsilateral M1 was best explained by force of the target hand (vs. the non-target hand). In conclusion, the data suggest that after incomplete cervical SCI, individuals remain capable of producing maximal supraspinal drive and are able to modulate this drive adequately. Activity of the ipsilateral motor network appears to be task related, although it remains uncertain how this activity contributes to task performance and whether this effect could potentially be harnessed to improve motor functioning.
Introduction
Spinal cord injury (SCI) has dramatic consequences for the affected individuals. The loss of structural integrity of the spinal cord affects bidirectional communication to and from the brain to peripheral effectors 1 and the loss of voluntary control of muscles is one of the most prominent impairments. Most of the injuries are classified as incomplete (∼ 66% 2 ), and approximately half of the individuals with SCI (54%) in the United States are discharged from hospital with a cervical injury (45% of the individuals has an injury at C4–C72). Consequently, many individuals with SCI are challenged by weakened hand muscles and one of the highest priorities for quadriplegics is to regain arm–hand function. 3,4
In able-bodied individuals, voluntary control of hand muscles is characterized by a series of distinctive features. First, a relatively large proportion of the motoneurons innervating hand muscles receive direct (i.e., monosynaptic) input from corticospinal fibers. 5,6 Second, most motor units are recruited at relatively low force levels (below 50% of maximal voluntary force [MVC] 7 –9 ) and increasing force output beyond this level relies on rate control. In comparison, in biceps brachii, the recruitment of additional motor units continues up to 88% maximal voluntary contractions (MVC). 7 Third, motoneurons innervating distal muscles, which include the intrinsic hand muscles, rely more heavily on synchronized input for their activation. 10,11
Marked changes in motor unit behavior have been observed in hand muscles weakened by SCI. Rate modulation in the thenar muscle was significantly reduced, which resulted in low maximal firing rates in motor units affected by SCI (9 Hz compared with 34 Hz in able-bodied individuals 12,13 ). Further, motor unit recruitment continued across an expanded force range (up to 85% MVC 12 ). Although it is evident that SCI impairs the transmission of descending supraspinal drive to the target motoneuron pools located below the level of injury, 1 much less is known about the contribution of supraspinal factors on the reduced firing rates. Indirect measurements obtained using neurophysiological techniques suggest that individuals with SCI are still capable of generating high levels of supraspinal drive. 14,15 However, only a few studies have focused on changes in brain activation during voluntary contractions after incomplete cervical SCI. 16,17 Although some studies have shown increased cortical activation during motor tasks, 18 –20 none have quantified or controlled for reduced force output.
Here, we use blood-oxygen-level dependent (BOLD) imaging to assess changes in cortical activation during voluntary contractions with a hand muscle following chronic cervical SCI. To answer the questions of whether individuals with incomplete SCI are still capable of generating maximal supraspinal output and whether they are capable of modulating this output, muscle force and cortical activation were measured during brief maximal and submaximal contractions with the first dorsal interosseous muscle.
Methods
Study population
Nineteen individuals with SCI (age 29–65 years, 4 females) and 24 age- and sex matched able-bodied controls participated in the study. Demographic information is presented in Table 1. Experimental procedures were approved by the medical ethical committee of the University Medical Center Groningen and all participants provided written informed consent.
Demographics and Injury Characteristics
Data shown as number (percentage) or mean/median (range).
SCI, spinal cord injury; AIS, American Spinal Injury Association impairment scale.
Magnetic resonance imaging (MRI) acquisition
Scans were acquired using a 3.0 T Philips Intera Achieva MRI scanner (Philips Medical Systems, Best, the Netherlands) equipped with an 8- (n = 16) or 32-channel (n = 27) SENSE head coil. Functional images were obtained using an echo-planar imaging (EPI) sequence (echo time = 29 ms; repetition time = 2 sec; flip angle = 82 degrees; 39 slices; 3.5 mm slice thickness; field of view 224 mm; matrix size 64 x 64; transverse slice orientation). A T1-weighted anatomical scan was obtained with the following characteristics (echo time = 30 ms; repetition time = 9 ms; flip angle = 8 degrees; 170 slices; 1 mm slice thickness; field of view = 256 mm; matrix size 256 x 256; transverse slice orientation).
Paradigm
Experimental setup and procedures were based on a previous study. 21 Participants lay supine in the scanner with a force-transducer in each hand. Their arms were extended and supported at the wrist using foam pads. Their head was immobilized using foam padding. All participants were familiarized with the experimental tasks during a practice session that took place at least 1 week prior to scanning. 14
Participants performed two motor tasks involving isometric index finger abductions (Fig. 1). During task 1 (MVC task), participants generated six MVCs alternatingly with the left and right hand. During task 2 (submax task), participants performed six sets of four contractions at 10, 30, 50, and 70% MVC (three sets per hand, three contractions per target level). Participants received feedback of the task and their force level on a screen that was visible via a mirror on top of the head coil. Participants were instructed to match their force level to a horizontal cursor indicating the target level and to hold their force as steady as possible; the target level was randomized within each set. All contractions were sustained for 10 sec followed by 50 or 40 sec rest for the MVC and submax task, respectively.
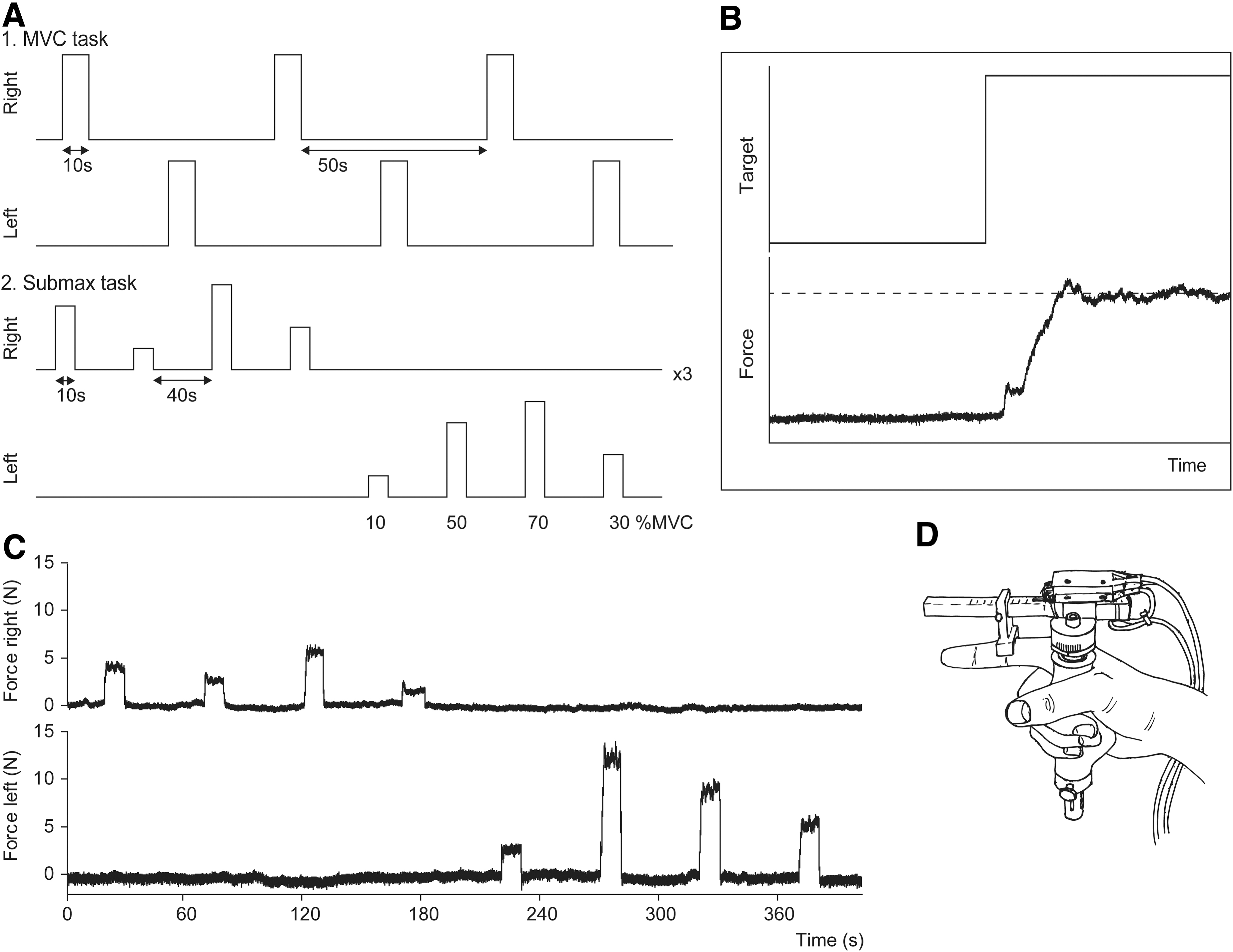
Experimental tasks and apparatus.
Force
Index finger abduction force was recorded using MR-compatible force transducers 22 (Fig. 1D). Participants wrapped digits 3–5 and their thumb around the handgrip while extending their index finger. The transducer was manipulated by the experimenters to align the horizontal bar parallel to the index finger and the C-shaped connector was positioned over the proximal interphalangeal joint. Participants' hands were taped to the transducers to maintain hand position. Force signals were amplified and sampled (500 Hz) using a 1401 micro interface and a computer equipped with Spike2 software (version 7.12, Cambridge Electronic Design, Cambridge, UK).
Statistical Analysis
Force data were analyzed in Spike2. Maximal force was determined for each participant during the MVC task. For the submaximal contractions, mean force was determined for the plateau phase of each contraction (2 sec after onset until 1 sec before relaxation) and expressed as a percentage of MVC. Onset and duration were determined for all muscle contractions (both tasks) to model the BOLD responses in the functional MRI data.
Statistical analysis of the force data was performed in RStudio (R version 3.4.3). MVCs were compared between groups using analyses of variance (ANOVAs), correcting for sex. Mean force level (as %MVC) during the submaximal contractions was compared across groups using multilevel regression. Model intercepts and slopes (for target level) were allowed to vary randomly for each participant. To assess differences between SCI and control participants, group and the interaction effect between group and force level were included in the model. After inclusion of additional fixed or random effects, the change in the Akaike information criterion (AIC) was used to assess whether the more complex model explained significantly more variance (decrease in AIC >2). Model residuals were examined graphically for normality and heteroscedasticity, and if required, the dependent variable was transformed to meet these criteria. To test robustness of the final models, the models were re-estimated on a trimmed data set (by excluding data points with scaled residuals >2 for the corresponding model). During the unilateral contractions, subjects also showed small associated contractions of the contralateral (non-target) muscle. This force was compared across groups using Mann–Whitney U tests per condition.
MRI data were analyzed using SPM12 (Wellcome Trust Centre for Neuroimaging, Institute of Neurology, London, UK,
Whole-brain analysis
General linear models (GLMs) were constructed for both tasks by defining contractions with the left and right hand as separate conditions (using the onsets and durations obtained from the force data). For the submax task, mean force level (as %MVC) was included in the GLM as a parametrically modulated regressor (orthogonalized polynomial expansion up to first order). Motion parameters from the realignment step were used to calculate framewise displacement between scans, and motion censoring was applied if framewise displacement exceeded a threshold of 0.9 mm. 23 Further, participants with root mean square of the motion parameters >1.5 mm (across an entire run) were excluded from further analysis. The realignment parameters were not included in the GLMs. The models were convolved with the canonical hemodynamic response function, and the data were high pass filtered with a cut-off of 128 and 275 sec for the MVC and submax task, respectively. The first level contrast images for each participant were used in a second level analysis; two-sample t tests were used to compare activation between SCI and control groups for the effect of “task” (both tasks) and “force” (submax task). Clusterwise inference was used (p < 0.05 familywise error [FWE] corrected, with a cluster forming threshold of p < 0.001 uncorrected and volume >20 voxels).
Region of interest (ROI) analysis
A ROI analysis was performed to assess the relationship between force modulation and activation in cortical motor areas. A total of 12 (bilateral) ROIs were obtained by intersecting anatomical ROIs of the cortical motor regions (human motor area template 24 ) with the group maps obtained from the MVC task (p < 0.05, FWE corrected). A GLM was constructed with eight conditions (four force levels × two hands). Beta estimates were extracted for each ROI and condition. To account for potential differences in localization of peak activity between individuals, the mean value was calculated from the top 25% voxels in each ROI. Multilevel regression was used to compare ROI activation between groups following same statistical procedure as described. Here, model intercepts and slopes (for mean force) were allowed to vary randomly per participant. Fixed effects were included for mean force, group (SCI), and the interaction between group and force. For ipsilateral M1, AIC scores were used to compare models including the mean force of the target versus models including non-target muscle.
Results
The data of one SCI and one control participant were excluded from further analysis because of excessive motion during both of the experimental tasks. Injury characteristics and/or demographics of the remaining SCI (n = 18) and control participants (n = 23) are provided in Table 1. For one SCI participant, all data obtained from the left hand were discarded because of incorrect task performance. MRI data were excluded because of excessive head motion during the MVC task (one SCI participant, right hand) and submax task (two SCI participants, both hands). A summary of the head motion analysis is provided in Supplementary Tables S1 and S2.
MVC task
As expected, SCI participants were weaker than controls as indicated by lower MVCs for the left (30.7 ± 12.3 vs. 50.3 ± 13.8 N, F1,37 = 24.6, p < 0.001) and right hand (19.4 ± 8.3 vs. 37.4 ± 8.0 N, F1,38 = 54.3, p < 0.001; Fig. 2). BOLD-activation during the MVCs with and right hand are shown in Figure 3. For the right hand, control participants demonstrated higher activation in a cluster in the right caudate nucleus (Fig. 3C). For the left hand, BOLD-activation did not differ across groups.
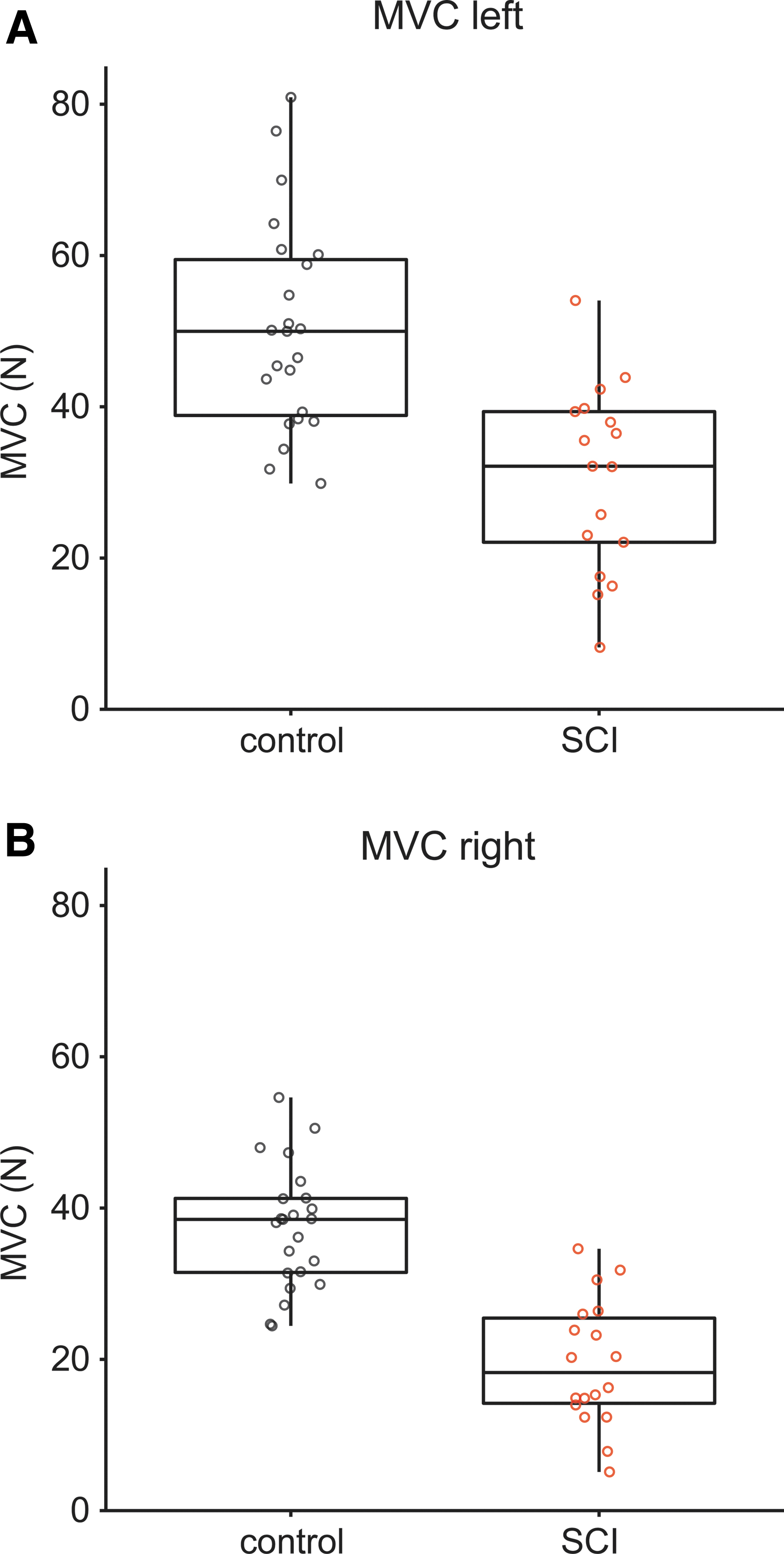
Maximal voluntary contraction (MVC)
Blood-oxygen-level dependent (BOLD) activation during the maximal voluntary contraction (MVC) task with the right hand. Voxels showing significant (p < 0.05, familywise error [FWE] corrected) BOLD activation for the main effect of task for MVCs with the right hand in
Submax task
On average, mean force (as %MVC) did not differ across groups (Fig. 4). However, a (small) interaction effect between group (SCI) and target force level was observed for the left hand (β = -0.028, t = -2.304, p = 0.027). Associated force of the non-target hand was significantly higher in the SCI group for the 70% condition with the right hand only (p = 0.009; Fig. 4D).
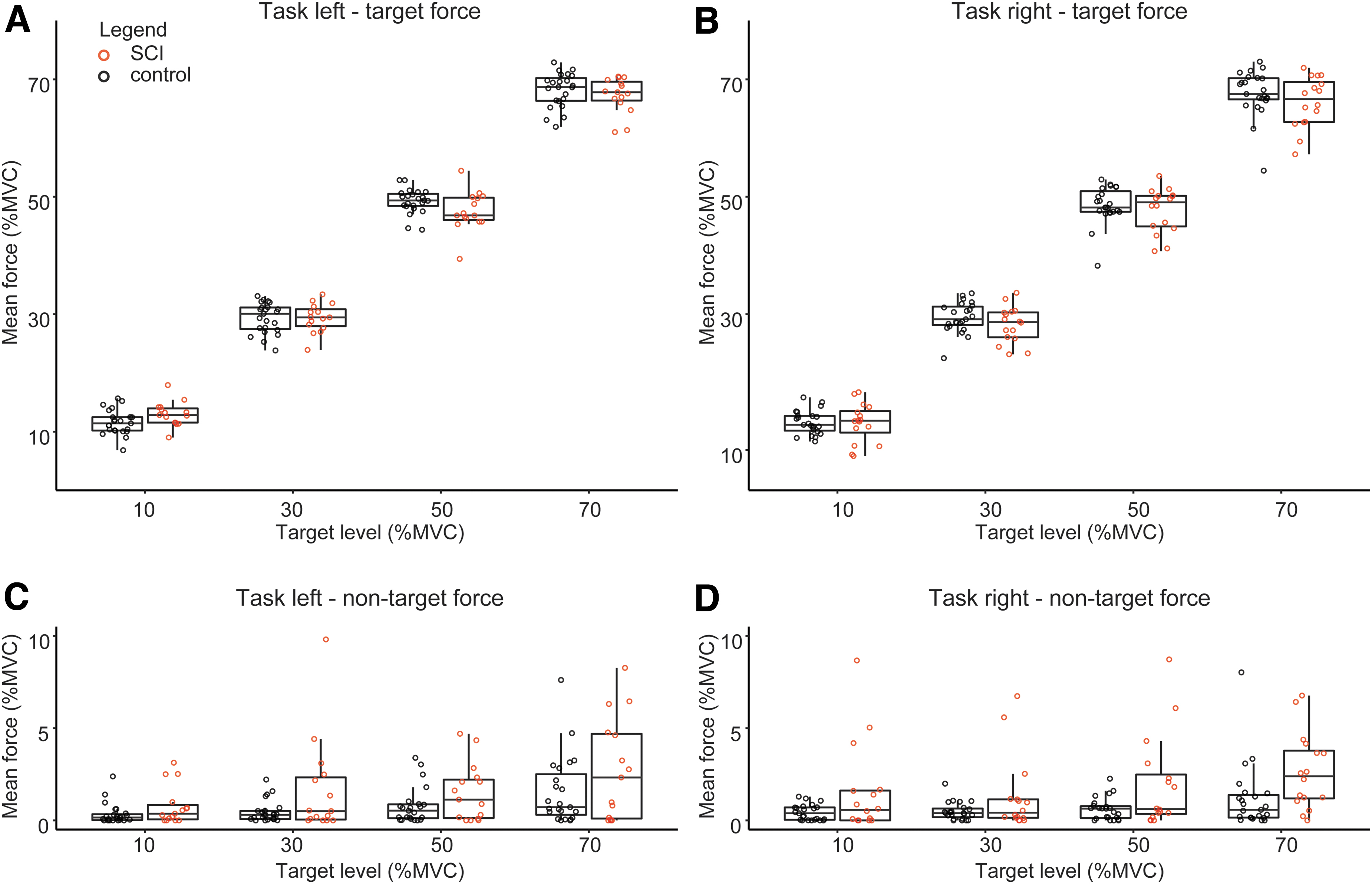
Force during the submax task. Mean force per target level during the submax task for the left
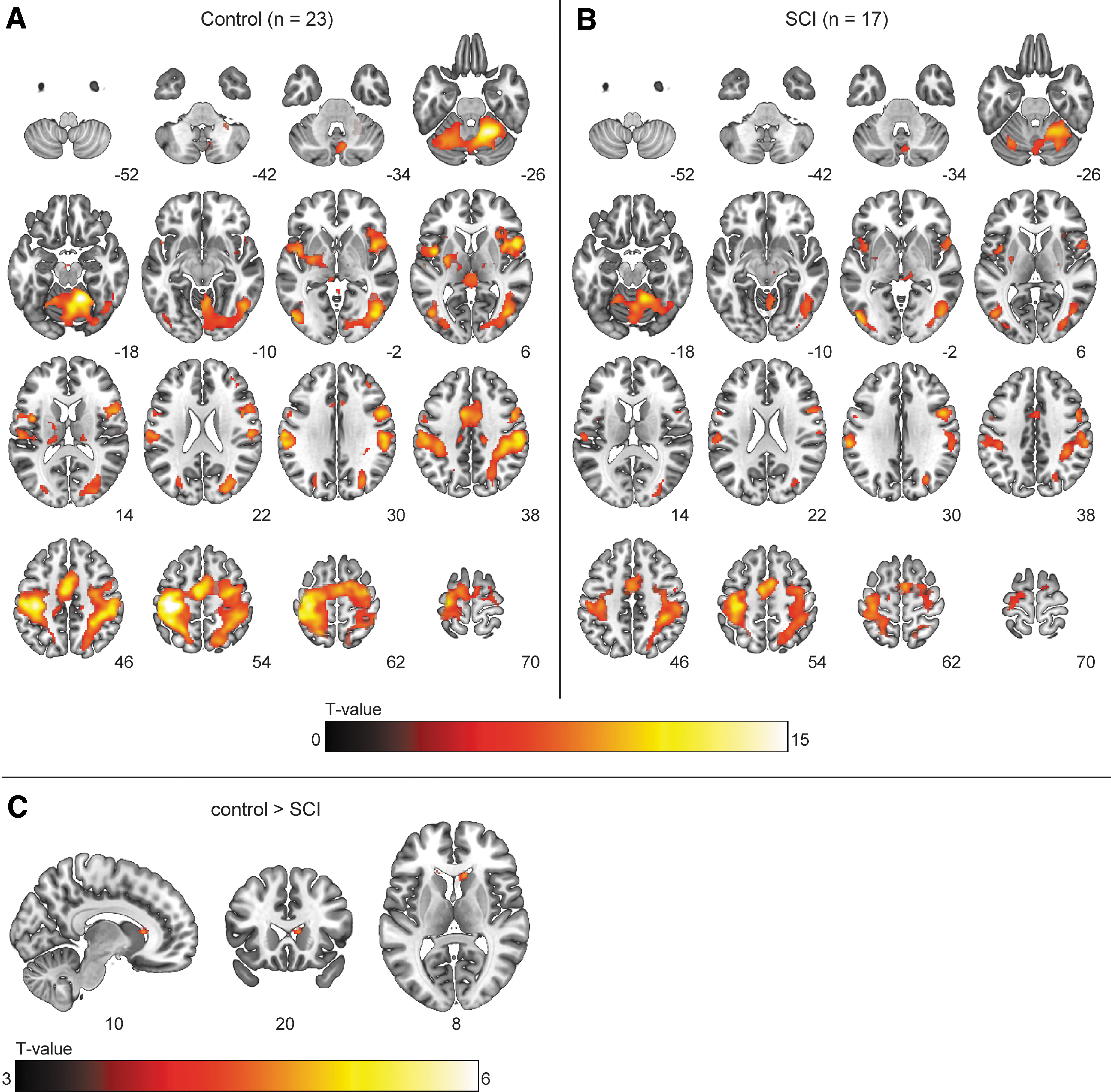
For the effect of “task,” whole-brain analysis demonstrated BOLD activation within the motor network in both groups (Fig. 5). In SCI participants, increased BOLD activation was found in a cluster spanning the ipsilateral M1, S1, supplementary motor area (SMA), and pre-motor cortex, as well as in clusters (predominantly) in the contralateral cerebellum (Fig. 6). For the effect of “force” (i.e., activation corresponding to the parametric covariate), BOLD activation was observed in the contralateral sensorimotor cortex and ipsilateral cerebellum; activation did not differ between groups.
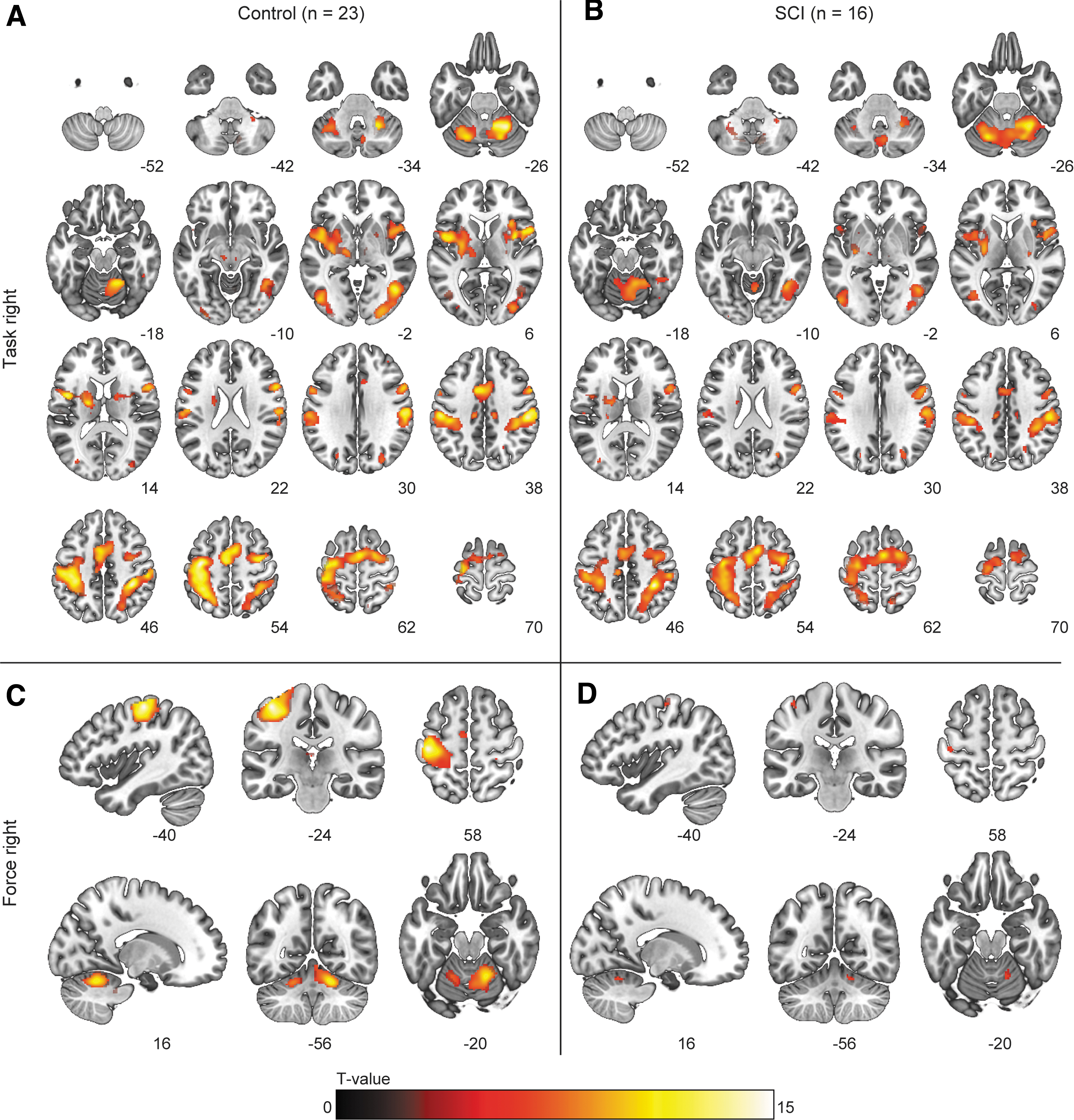
Blood-oxygen-level dependent (BOLD) activation during the submax task with the right hand. Voxels showing significant (p < 0.05, familywise error [FWE] corrected) activation for the main effect of “task” in the
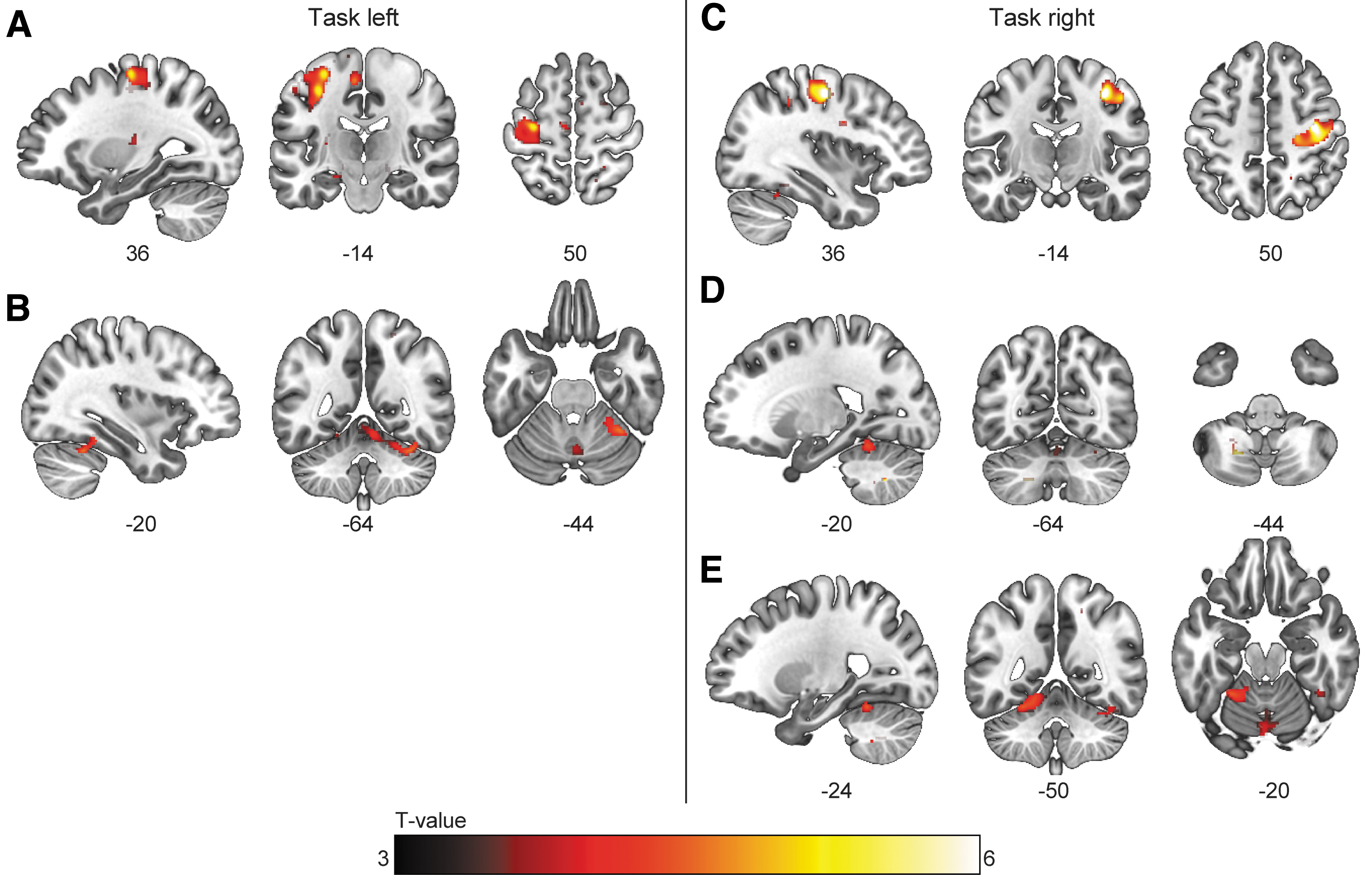
Blood-oxygen-level dependent (BOLD) activation during the submax task with the right hand. Clusters showing significantly (p < 0.05 familywise error [FWE] corrected at cluster level, κ > 20) higher activation in spinal cord injury (SCI) participants than in the control group for the effect of “task” during the submaximal contractions. For the left hand, BOLD activity was higher in
The results of the ROI analysis are summarized in Figure 7. All motor regions (contra- and ipsilateral) demonstrated a significant increase in BOLD activation with increasing force level (all p < 0.001). Modulation of the BOLD activity with increasing force was the strongest in the contralateral M1 and S1 (both hands) and contralateral SMA (right hand). Activity was significantly higher in the SCI group in all regions (all p < 0.05), except for the right S1 (both hands) and the left pre-SMA and S1 (right hand). No interaction between group and force was observed in any of the regions indicating similar modulation of the BOLD signal with force across groups. BOLD activation in the ipsilateral M1 was explained better by the force of the target hand than by the associated force of the non-target hand (task left: Δ AIC = 18.5, task right: Δ AIC = 23.6).
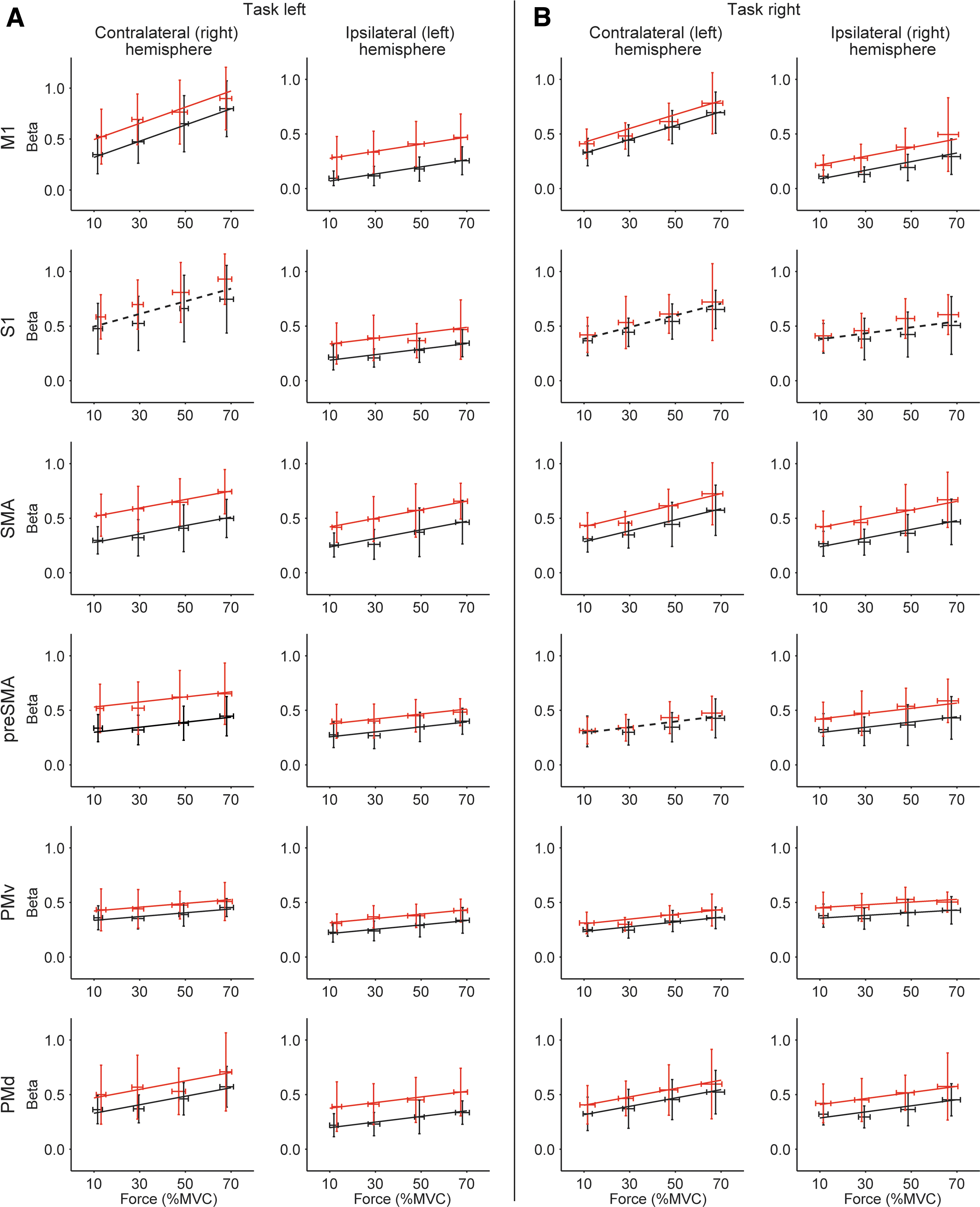
Multi-level models for the region of interest (ROI) analysis. Panels show the regression lines obtained for the multi-level analysis of the ROI betas for the submax task with
Discussion
Performance of index finger abductions at various force levels by individuals weakened by SCI was characterized by increased brain activation in both the contra- and ipsilateral motor networks, compared with age-matched controls. The increased BOLD-activation was more strongly related to the effect of task than to force gradation. Interestingly, brain activation did not differ between SCI and control groups during maximal voluntary contractions.
Individuals with SCI were able to adequately modulate supraspinal drive
Although SCI participants generated significantly weaker maximal forces than the control participants, minimal differences in BOLD activation were observed between the groups during the MVCs. This finding supports neurophysiological data 14,15 suggesting that individuals with incomplete SCI remain capable of maximally increasing their supraspinal drive. As expected, force gradation was associated with increased activation in the contralateral motor cortex, pre-motor areas, and ipsilateral cerebellum, 25 –27 but no differences between the groups were observed. This result was confirmed by the ROI analysis, which demonstrated the strongest effect of force level on BOLD activation in the contralateral M1, S1, and SMA. Thus, our data suggest that individuals with SCI are still capable of modulating their supraspinal drive in order to increase force output, and are able to do so adequately.
Increased ipsilateral activation in SCI participants
Although force gradation was not associated with different activation patterns between the groups, we observed increased activity in the ipsilateral M1, S1, SMA, and pre-motor cortex, as well as in the contralateral cerebellum in SCI participants, which was related to task performance. This finding was supported by the ROI analysis, which demonstrated higher intercepts in the regression models for the SCI group. Thus, SCI participants showed increased BOLD-activation that was related to index finger abduction but not to force gradation.
The relevance of activation in ipsilateral motor areas while performing unimanual motor tasks remains a topic of debate. 28,29 It could be a cross-activation phenomenon reflecting reduced inhibition of the ipsilateral cortical areas 30 with little functional consequence; however, there are several studies to suggest that ipsilateral activation may be functionally relevant for motor performance. First, neuronal firing rates recorded from the motor cortex of monkeys showed activation during both contra- and ipsilateral arm movements, 31,32 and the patterns of cortical activation could predict muscle activity of either the contra- or ipsilateral arm. 31 Further, a virtual lesion of the ipsilateral motor cortex, induced by repetitive transcranial magnetic stimulation, resulted in timing errors of muscle activation during a hand grip task. 33 Finally, following hemisection of the corticospinal tract in monkeys, the hand representation in the ipsilateral M1 decreased. 34 In the present study, BOLD activity in the ipsilateral M1 was better associated with task-related muscle activation in the target hand than with the associated force of the non-target hand. Combined evidence suggests a role for the ipsilateral motor cortex during movement preparation. 31
In addition to muscle weakness, SCI poses difficulties for force gradation, 1 making matching submaximal forces more difficult. Because task precision has also been shown to be associated with increased activation in the ipsilateral M1, 35 it is possible that increased difficulty underlies the ipsilateral activation. Alternatively, the activation of the ipsilateral motor areas in the present setting could be related to the hand grip force necessary to hold the force transducer during the index finger abductions. Ipsilateral projections from the primary motor cortex to neurons in the reticular formation have been described, 36 and ipsilateral reticulospinal projections to spinal motoneurons are known to be involved in power grip, but less in fine finger movements after spinal cord injury. 37 This also explains why the increased ipsilateral activity was associated more strongly with the effect of task than with force modulation.
Because ipsilateral cortical activation is often increased in populations with reduced muscle activation – including incomplete SCI, 18 –20 multiple sclerosis, 38,39 and stroke, 40 as well as in elderly individuals 41 – better understanding of this ipsilateral activation is relevant for both motor control and rehabilitation purposes. Following SCI, the extent of ipsilateral activation has been shown to be associated with injury severity, defined by the degree of spinal cord atrophy, 20 and ipsilateral activation has been shown to decrease with time post-injury. 18
Conclusion
We show that individuals following incomplete cervical SCI remain capable of producing maximal supraspinal drive and are able to modulate this drive adequately. The changes in motoneuron (firing) behavior in hand muscles, which occur following SCI, are therefore more likely because of reduced supraspinal input received by the motoneurons. As a consequence of damage to descending tracts where they pass the lesion site, axons that synapse onto the motoneuron pools located downstream from the injury show disruption and demyelination. 1,42 Thus, the supraspinal drive that reaches these motoneuron pools is diminished and there is loss of synchronicity of the synaptic input. 42
Further, we found that task performance in individuals with SCI was accompanied by increased ipsilateral activation, which is related to the output of the task-performing hand. It remains uncertain how this activation contributes to the task and whether this effect can be harnessed to improve motor functioning.
Footnotes
Acknowledgments
We thank Tjalling Nijboer for his technical support, and Anita Sibeijn-Kuiper and Judith Streurman for operating the MR scanner. We also thank all our participants for taking part in this study.
Funding Information
Grants were provided by the Junior Scientific Masterclass of the University Medical Center Groningen, the National Institutes of Health (N01-NS-3-2351), and The Miami Project to Cure Paralysis.
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.