
Abstract
Post-traumatic epilepsy (PTE) is acquired epilepsy after traumatic brain injury (TBI). Despite the availability of more than 20 antiseizure medications (ASMs), there is no way at present to prevent epileptogenesis in TBI survivors, and many cases of PTE become drug-resistant. Importantly, the adverse effects of ASMs can significantly affect patients' quality of life. Mammalian models are commonly used for studying refractory PTE, but are expensive and laborious. Zebrafish models have become popular for studying epilepsy, but most focus on larvae, and there have been no reports to date of pharmacological screening in an adult zebrafish model of acquired epilepsy. Valid animal models are critical for understanding PTE and for developing novel therapeutics. The aim of the present study was to characterize the cognitive impairments of a zebrafish model of TBI that leads to the development of PTE. Using combined behavioral and electrophysiological approaches, we also characterized the pharmacological effects of the most commonly used ASMs to manage PTE (valproate, carbamazepine, and phenytoin). Zebrafish with PTE exhibited impairments in learning and memory, difficulty in decision making, and reduced social preference. Valproate and carbamazepine had a limited protective effect against behavioral seizures, and all three drugs failed to significantly reduce electrographical seizures. The negative impacts of TBI and ASMs in zebrafish parallel those observed in other animals, making the zebrafish model of PTE a promising high-throughput model of refractory and drug-resistant epilepsy.
Introduction
Traumatic brain injury (TBI) is a leading cause of chronic neurologic deficits, disabilities, and death, making it a major socioeconomic and health care burden. 1,2 Post-traumatic epilepsy (PTE) is a common complication of TBI that can begin up to decades after the initial injury. 3,4 There are no treatments to prevent epileptogenesis after TBI, and PTE is drug resistant in one-third of patients with PTE. 5
Valid animal models are critical for understanding PTE and developing novel therapeutics. The vast majority of PTE studies rely on rodents because of their high genetic homology with humans and similarities in seizure-related phenotypes. Because injury severity is one of the greatest predictors of PTE, 6 however, high death totals are unavoidable in such experiments. 7 Moreover, fewer than 50% of rodents with severe TBI develop epilepsy after a year with a low seizure rate (<1 per day). 8
Zebrafish are amenable to quantitative measures of TBI and PTE outcomes. Using zebrafish can be less expensive and allows for higher-throughput experiments, which makes a zebrafish model of PTE a good complementary model to rodent studies. We previously established a zebrafish model of PTE induced by closed-head and brain-specific TBI by delivering short pulses of high-intensity, focused ultrasound energy (pHIFU), which induces stress forces and cavitation in tissues. 9,10 The zebrafish PTE model mimics many clinical aspects of human PTE, including blood–brain barrier disruption, molecular changes, and motor deficits similar to those observed in humans. Most importantly, zebrafish with severe TBI developed spontaneous, unprovoked, and recurrent seizures, the hallmark of epilepsy. 10
Psychosocial impairment is one of the most debilitating consequences in survivors of TBI and is worsened by the concomitant development of epilepsy. 11 Patients with PTE experience more severe psychosocial dysfunction than patients with TBI alone. 12,13 In addition, antiseizure medications (ASMs) are associated with a high risk of adverse effects such as neurobehavioral, neuropsychological, and cognitive impairments (Table 1). 14 –17 Models utilizing adult zebrafish allow for investigation of cognitive outcomes because they exhibit broader and higher-order behavioral responses than zebrafish larvae. 14
Neuropsychological Side Effects of Anticonvulsants
In this article, we describe the effect of ASMs on behavioral and electrophysiological seizures in the zebrafish PTE model. In addition, we tested the effect of ASMs on the cognitive ability of zebrafish with severe TBI using two different T-maze paradigms to examine social preference and learning and memory.
Methods
Animals
All procedures were conducted under guidelines established by the Animal Care Committee at St. Michael's Hospital in accordance with the standards set by the Canadian Council on Animal Care. Male and female AB strain zebrafish (6–12 months old) were maintained under a 12:12-h light:dark cycle and kept in tanks at 25°C and pH 6.8–7.0.
Induction of PTE
The TBI and subsequent PTE were induced in zebrafish by using a pHIFU system, as described previously. 10 Briefly, anesthetized zebrafish were secured in restraint; the right telencephalon was aligned to the pHIFU focal zone and was subjected to three consecutive bursts of acoustic pressure of 3MPa. Because of the nature of the non-invasive injury procedure, we used naïve animals as controls. Zebrafish were then moved to an anesthetic-free recovery tank, righting reflex time was measured, and deaths or hemorrhages were noted.
Drug reagents
Individual solutions of sodium valproate (VPA), carbamazepine (CBZ), and phenytoin (PHT) (Sigma-Aldrich, Canada) were prepared daily, each dissolved in dimethyl sulfoxide (DMSO) and diluted in filtered room-temperature water to achieve a final DMSO concentration of 1% w/v.
Toxicological evaluation
We established the maximum tolerated concentration (MTC) of ASMs to determine experimental concentrations. Zebrafish were incubated with different concentrations of ASMs for 24 h in complete darkness, after which the following signs of toxicity were assessed: bleeding, loss of posture, body deformation, escape response, slow heartbeat, and death. 18 The MTC of ASMs was defined as the highest concentration that resulted in no signs of toxicity in any fish in a group (n = 5 per group) after 24 h of exposure.
We used these values to assess their immediate effects in zebrafish with PTE. At 14 days post-injury (dpi), injured zebrafish were randomly selected and assigned to each drug group (n = 9). Differences in seizure rate and duration, number of interictal epileptiform discharges (IEDs), learning and memory, decision making, social interaction, locomotor changes were evaluated, and pre- and post-treatment results were compared.
Behavioral seizure scoring
We evaluated the development of epilepsy at 3, 7, and 13 dpi. Zebrafish were individually transferred to a novel 2-L tank and video-recorded for 30 min. Video files were scored blindly and manually by a researcher using previously validated behavioral seizure scales. 10 At 14 dpi, the same fish from the 13 dpi evaluation were treated with ASMs for 2 h and behavioral seizures were scored again.
Electrophysiology
At 21 dpi, zebrafish from the naïve, TBI-only, and various treatment groups underwent electrophysiological recordings. Fish from the treatment groups were immersed in the drugs for 2 h before the recording session. For electrophysiological recordings, fish were anesthetized, and their craniums were delicately removed. A chloride-coated silver wire was placed on the supraneural spine as a reference electrode, 19,20 and a NaCl-filled glass electrode was placed on the right telencephalon as described previously. 10 Field potential signals from the right telencephalon were recorded using an Axopatch 200B amplifier (Molecular Devices) in the frequency range of 0.1–55 Hz. Recorded signals were analyzed blindly and manually by two researchers using Clampfit 11.0.3 (Molecular Devices).
T-Maze
The T-maze was made of white, non-transparent acrylic and consisted of a starting zone (5 cm × 12.5 cm) and two long arms, each made up of a corridor (14.5 cm × 8.5 cm) and a reservoir (10 cm × 8.5 cm). The maze was filled with water to a depth of 10 cm. Each group was habituated in the T-maze for 30 min before testing to minimize stress from procedural novelty.
To test learning and memory, fish were fasted for 24 h to increase their appetite drive. Using a procedure adapted from Colwill and colleagues, 21 we made the two arms visually distinct: the rewarding environment was covered with a blue sleeve, and the punishing environment was covered with a red sleeve (Fig. 1C). As soon as a fish entered the blue arm, it was immediately transferred to a blue container and rewarded with food. When a fish entered the red arm, it was stroked persistently with a string on a soft plastic rod for 30 sec to develop passive avoidance behavior. This procedure was performed in three learning trials separated by 10-min intervals. In a fourth trial (test), choice accuracy (the rewarding environment was the “correct” choice) and latency to enter an arm were recorded manually.
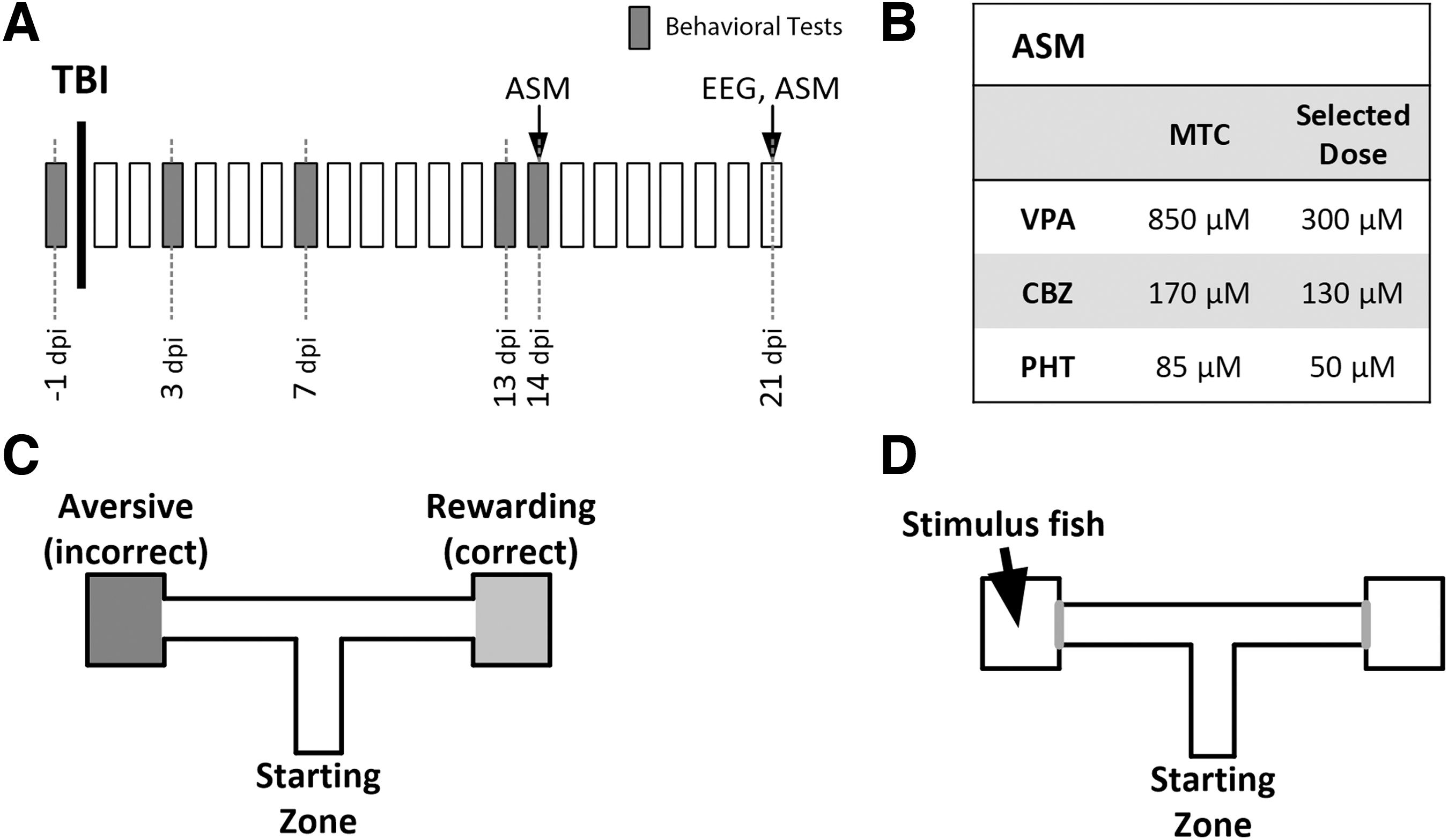
Experimental design. Timeline. Behavioral seizure monitoring and cognitive tests were performed in naïve animals one day before traumatic brain injury (TBI) and repeated at 3, 7, 13, and 14 days post-injury (dpi). At 14 dpi, antiseizure medications (ASMs) were administered and behavioral monitoring was repeated; the results were compared with those from 13 dpi. Electrographical recording was performed at 21 dpi (
To evaluate social interaction, we blocked each reservoir with a transparent door and placed five stimulus fish in one of the reservoirs (Fig. 1D). Test zebrafish were gently introduced individually to the starting zone and videotaped for 5 min before being gently netted and transferred back to their home tank. Fish behavior was scored using automated tracking software (ToxTrac) 22 to assess interaction time, total distance, average speed, and exploration rate.
To assess the effect of ASMs, injured zebrafish were randomly assigned a drug treatment (VPA, CBZ, or PHT). Cognitive ability was assessed before and after drug administration.
Statistical analysis
Data were analyzed with GraphPad Prism. Discrete data are reported as percentages and were compared using the Fisher exact test. Continuous data are reported as mean ± standard error of the mean (SEM) and tested for normality using a combination of Anderson-Darling, D'Agostino & Pearson, Shapiro-Wilk, and Kolmogorov-Smirov tests and then compared using parametric (paired t test, one-way analysis of variance [ANOVA]) or non-parametric tests (Wilcoxon matched-pairs rank test, Kruskal-Wallis test) as approriate.
Because this study was not designed to compare effectiveness of the various treatments against each other, continuous data from electrophysiology recordings were compared for each treatment group versus the naïve group. Differences in T-maze latencies and social interaction times were compared between naïve and injured fish at various time points. For all comparisons made in regard to effect of ASMs, the results in untreated versus treated fish were compared using paired t tests. p < 0.05 was considered significant.
Results
Maximum tolerated concentrations of ASMs and drug exposure
The MTCs for VPA, CBZ, and PHT were established as 850 μM, 170 μM, and 85 μM, respectively. These MTCs were higher than the doses previously reported to suppress PTZ-induced seizures. 23,24 For the remaining experiments, we selected concentrations lower than the MTCs but higher than those reported in PTZ studies, because our MTC tests were performed in non-injured, healthy animals. The doses we selected for VPA, CBZ, and PHT were 300 μM, 130 μM, and 50 μM, respectively (Fig. 1B).
Seizure development after severe TBI
Spontaneous myoclonic-like behavioral seizures were observed in 81–100% of injured zebrafish (Fig. 2A), and more convulsive, tonic-clonic–like seizures were observed in 44–82% of fish over the first 13 dpi (Fig. 2B), with increasing proportions at later times. The number of seizures was significantly greater at all time points compared with the naïve group, in which no seizures were detected (Fisher exact test: myoclonic: p = 1E-4 for all time points, tonic-clonic: p = 0.027, 5E-4, and 1E-4).

Zebrafish with severe traumatic brain injury (TBI) develop behavioral seizures that respond well to sodium valproate (VPA) but not carbamazepine (CBZ) or phenytoin (PHT). The TBI led to a significant percentage of fish developing spontaneous behavioral myoclonic-like seizures (
Erratic movements, defined as stage 2 seizures in our previous study, 10 are prodromes of seizures in zebrafish. We compared the occurrence of erratic movements and seizures in the untreated state (13 dpi) versus treated state (14 dpi) using the Fisher exact test. Before treatment, all injured fish showed erratic movements. The rate of occurrence of erratic movements decreased significantly only after exposure to VPA (from 100% of fish to 50%, p = 0.038), which also suppressed myoclonic seizures (from 100% to 38%, p = 0.013) and tonic-clonic seizures (from 75% to 13%, p = 0.013).
A CBZ exposure suppressed tonic-clonic seizures with the greatest effect (from 78% to 11%, p = 0.008), but did not change the rate of erratic movements or myoclonic seizures (p = 0.500, and 0.103, respectively) The PHT-treated group showed no significant changes after treatment in erratic movements, myoclonic or tonic–clonic seizures (p = 1.0, 0.500, and 0.167, respectively) (Fig. 2C–E).
Electrographical seizures
Electroencephalographical recordings from the naïve, TBI-alone, VPA-treated, CBZ-treated, and PHT-treated groups were performed at 21 dpi to confirm the occurrence of spontaneous seizure activity. To compare the effects of ASMs on ictogenesis after TBI, we quantified the number of IEDs and seizure duration. The IEDs were defined as sharp spikes that lasted <3 sec, whereas discharges of >3 sec were considered ictal. No naïve animals exhibited electrographical seizures, while 71% of the TBI group showed at least one seizure during the 15-min recording.
There was no significant decrease in the proportion of fish having seizures after treatment (Fig. 3B). There were 6.60 ± 2.18 IEDs and no ictal discharges detected in the naïve animals. The VPA-treated group showed a mild but not statistically significant increase in the number of IEDs and seizure duration compared with naives (p = 0.170, and 0.072, respectively). The number of IEDs in the CBZ-treated group also demonstrated a trend toward an increase (p = 0.076), and seizure duration was significantly increased (p = 0.045). The PHT-treated group had significantly more IEDs and longer seizures compared with the naïve group (p = 0.006, and 0.023, respectively). There were no statistically significant differences in the number of IEDs or the duration of seizures, however, between any drug-treated groups and the TBI-alone group (Fig. 3C, D).
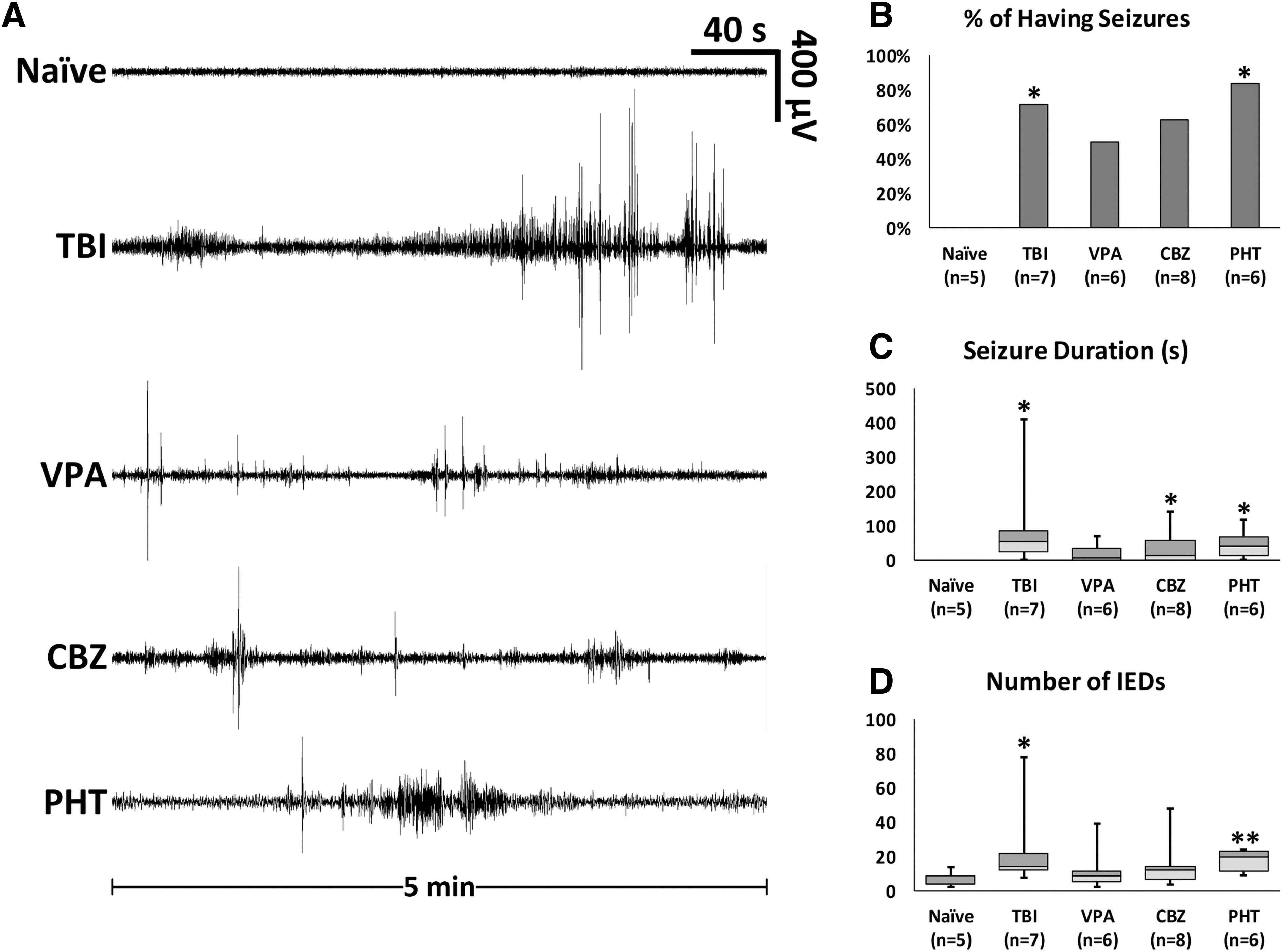
Spontaneous electroencephalographical seizures in zebrafish with post-traumatic epilepsy. Representative epochs from naïve fish, fish with traumatic brain injury (TBI), and fish with TBI after drug treatment. Recordings performed at 21 days post-injury (dpi) (
Cognitive ability of zebrafish with PTE
Of naive fish, 67% made correct choices in the T-maze, which was higher than the rates at 3 and 13 dpi in the TBI-alone group, but not statistically significant (27%, p = 0.067 and 37%, p = 0.101, respectively) (Fig. 4A). Interestingly, all naïve fish entered at least one reservoir (correct or incorrect) during the trial, but 40% of fish at 3 dpi (p = 0.037) and 32% of fish at 13 dpi (p = 0.049) did not enter either reservoir during the trial (Fig. 4B). Data for latency to enter a maze arm were not normally distributed, and although the Kruskal-Wallis test did not reveal a significant difference between latencies to enter a maze arm (p = 0.335), naïve fish took an average of 75.56 ± 12.39 sec to enter either arm. This value was 65% higher at 3 dpi (124.13 ± 17.36 sec), before decreasing slightly at 13 dpi (103.54 ± 11.57 sec) (Fig. 4C).
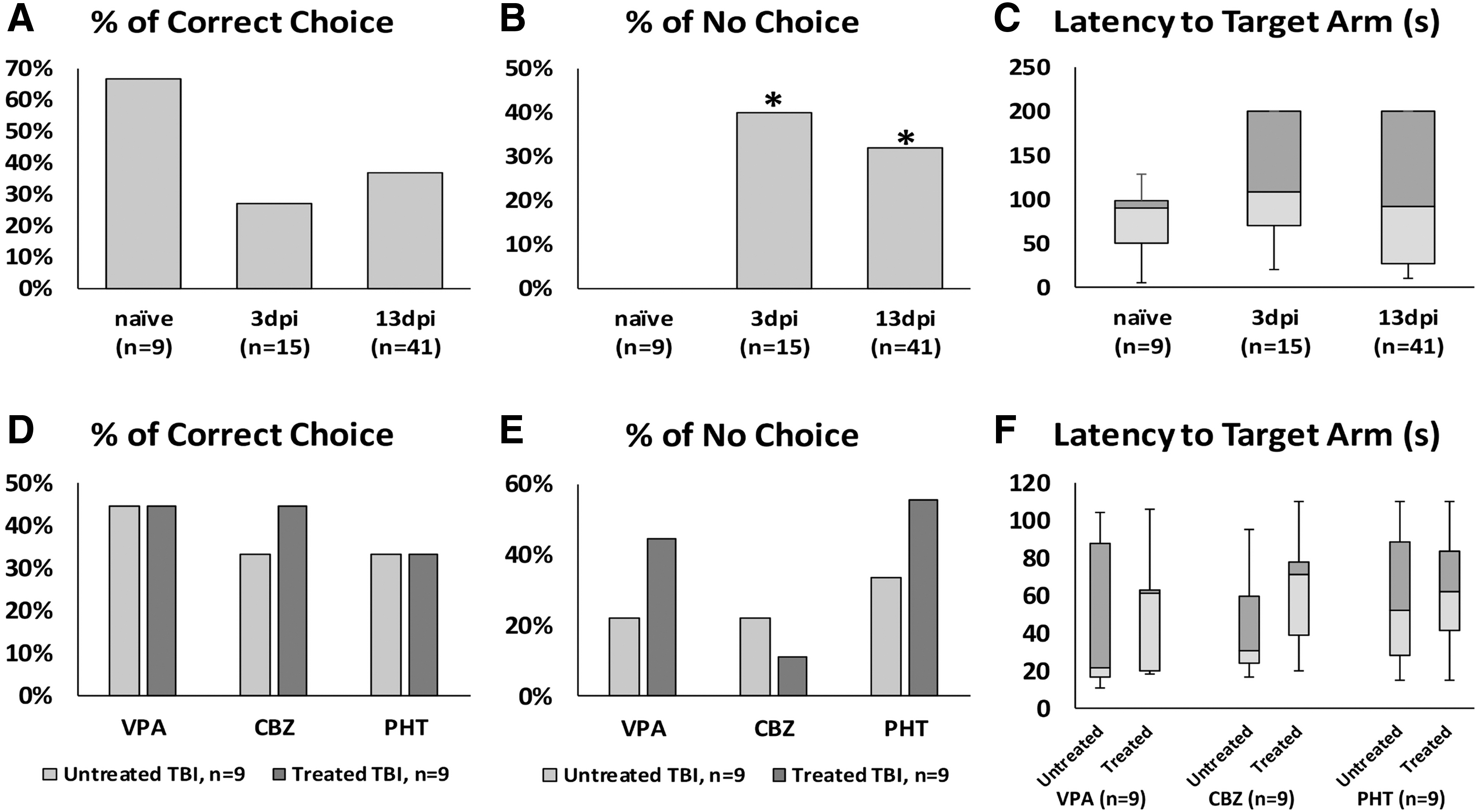
Effect of severe traumatic brain injury (TBI) and antiseizure medications (ASMs) treatment on learning, memory, and decision making in the T-maze task. There were no significant effects on learning (
Cognitive effects of ASMs in a zebrafish model of PTE
There were no statistically significant differences in correct choices, no-choice, or latency to respond (Fig. 4D,E,F) in any drug-treated groups versus TBI-alone. After drug administration, however, all three groups showed some changes in no-choice events during the trial (VPA: from 22% to 44%; CBZ: from 22% to 11%; PHT: from 33% to 56%).
Decreased social interaction after TBI
Data for social interaction time (time spent near the stimulus fish) were normally distributed and were significantly different when compared with a one-way ANOVA (F = 5.960 (2, 64), p = 0.0042). The interaction time remained stable until 3 dpi, but significantly dropped at 13 dpi (Fig. 5A, Tukey multiple comparisons test p = 0.0099). Paired t tests were used to compare social interaction time in untreated versus treated fish. Social interaction times were increased in all three ASM-treated groups, but the only statistically significant difference was for PHT: VPA, from 102.22 ± 32.93 sec to 154.53 ± 29.72 sec (p = 0.235); CBZ, from 130.31 ± 19.42 sec to 200.89 ± 30.41 sec (p = 0.071); PHT, from 104.79 ± 31.28 sec to 267.23 ± 9.51 sec (p = 0.001).
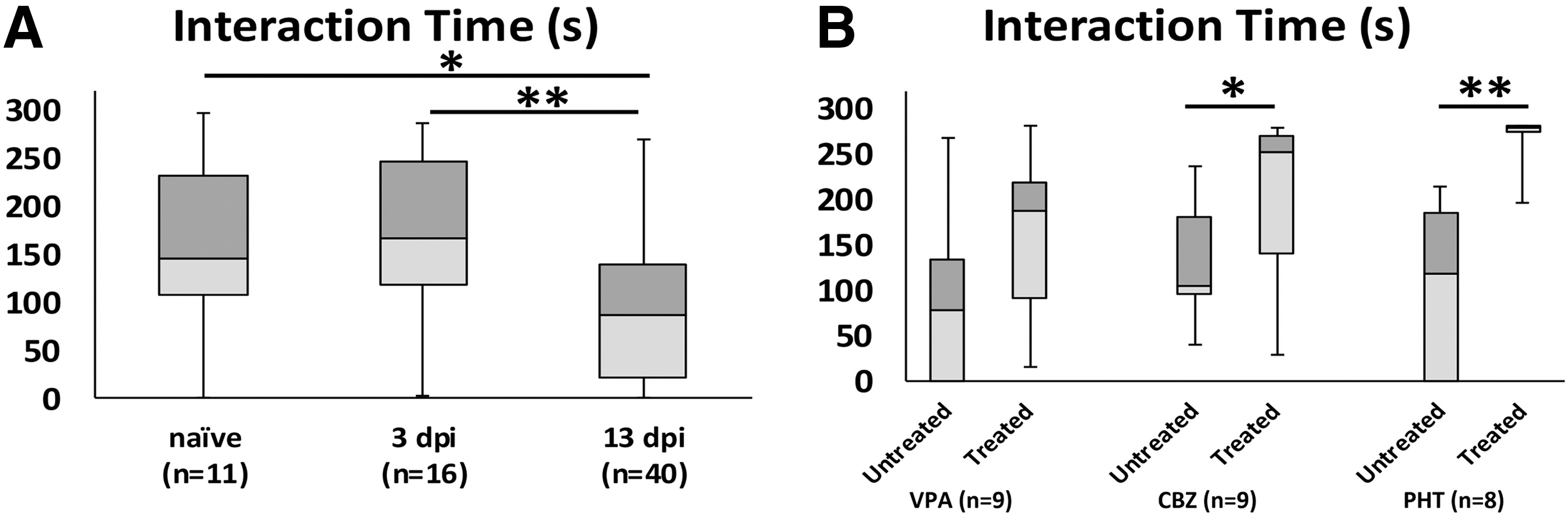
Social preference reduced two weeks after traumatic brain injury (TBI).
We also compared locomotor behavior changes during the social interaction test. The TBI groups did not show changes in total distance traveled, average speed, or exploration rate at 13 dpi (Fig. 6), nor did ASMs affect the total distance traveled or average speed. The CBZ-treated group showed a decrease in exploration rate (p = 0.029), while VPA and PHT had no significant effect (p = 0.986 and p = 0.299, respectively).

Locomotor activity and spontaneous exploration in the T-maze. Traumatic brain injury (TBI) had no effect on the total distance traveled, the average swim speed, or the exploration rate in the T-maze compared with naïve controls (
Discussion
Zebrafish-based paradigms are becoming increasingly popular in behavioral and pharmacological studies of neurological disorders because they offer economical and high-throughput approaches. 25 –30 Several studies have investigated ASM effects on acute seizures in adult zebrafish. 23,31,32 The present study is the first to assess the effect of ASMs in an adult zebrafish model of acquired epilepsy. We evaluated cognitive changes in zebrafish after severe TBI and assessed the immediate effects of three commonly used ASMs—VPA, CBZ, and PHT—on seizure suppression and cognitive behavior. These ASMs are often effective in managing early post-traumatic seizures, but do not prevent the development of PTE. 15,33,34
After sustaining severe closed-head injury, all zebrafish developed myoclonic-like behavioral seizures, and by 13 dpi, 81% had developed tonic-clonic-like behavioral seizures observable during 30 min of monitoring. This rate of seizure development after TBI aligns with our previous report. 10 It should be noted that the incidence of PTE in the zebrafish model is higher than what is seen clinically. 10 While this does not perfectly recapitulate the human condition, it may allow for mechanistic and treatment studies to be performed more readily than in other models.
After a 2-h treatment with VPA, CBZ, or PHT, only the VPA-treated group showed a significant reduction in all seizure-related behaviors—erratic movements, myoclonic seizures, and tonic-clonic seizures. The VPA-treated group also had similar electrographical findings compared with the naïve group. No significant changes in electrographical seizures, however, were observed with any of the ASMs compared with TBI alone, despite the fact the drugs we used are known to be effective in drug-induced seizure models.
We had selected drug concentrations lower than our MTCs, but higher or the same as reported in drug-induced acute seizure studies using zebrafish (Fig. 1B). 18,23,24,32,35 The discordance between behavioral and electrographical seizures was not necessarily unexpected, because patients with TBI can have subclinical/non-convulsive seizures while receiving ASMs, 36 and studies using other models of zebrafish seizures have reported subclinical seizures without behavioral correlate. 37
Our findings suggest this model could serve as a valuable tool in studying refractory epilepsy and developing treatment strategies. On exposure to the T-maze, injured zebrafish were significantly more likely to have “no-choice” events, defined as the failure to enter either reservoir within the 5-min time limit, while no-choice outcomes were not seen with any naïve zebrafish. Not choosing either arm indicates learning deficits and indecision 38 ; the behavior is also observed in animals with phenylketonuria and after exposure to the hallucinogen lysergic acid diethylamide. 38,39 There was also a trend for a decreased rate of correct choices at 3 dpi, starting to recover to naïve levels by 13 dpi. Together, these results demonstrate zebrafish with TBI experience learning deficits.
There were no differences in locomotor behavior in the T-maze after TBI, and we have similarly shown zebrafish with severe TBI do not exhibit significant motor changes at 14 dpi in a novel-tank test, 10 indicating poor performance in learning and memory tasks was not because of physical disability. Somewhat surprisingly, treatments with VPA, CBZ, and PHT did not affect T-maze performance. Our results suggest zebrafish with TBI have deficits in learning and decision making that are not further affected by treatment with ASMs.
As expected, severe TBI in our zebrafish model induced significant social deficits at 13 dpi, consistent with previous reports of psychosocial deficits after TBI in other animal models. 11,40,41 We did not find any difference in social behavior immediately after a 2-h acute exposure to VPA or CBZ, whereas there was a positive effect of PHT exposure as evidenced by increased social interaction. Our results for VPA are consistent with previous reports showing chronic exposure in zebrafish reduces sociability, whereas acute exposure (up to 7 h) does not affect social behavior. 42 We could not find any reports on the effects of CBZ and PHT exposure on zebrafish behavior, but preliminary evidence indicates these drugs may enhance social behavior, 43,44 consistent with our findings for PHT.
This is the first study of neuropsychological impairment and ASM response in a zebrafish model of TBI and PTE, which is also the first acquired epilepsy model in adult zebrafish exhibiting drug-resistant and refractory characteristics. A limitation of the study is the neuropsychological effect of ASMs was not tested in naïve animals. In addition, we only used a single concentration of each drug, and only studied immediate effects. Further ASM screening with a range of doses and exposure times, perhaps including other drugs effective in focal epilepsy (e.g., levetiracetam, oxcarbazepine), may provide more insight into the complex neuropharmacological profile of this epilepsy model. In addition, we have not tested the effect of any therapies on the process of trauma-induced epileptogenesis.
While there are currently no anti-epileptogenic therapies to prevent PTE, pre-clinical studies have identified promising targets. 45,46 Future studies should investigate whether candidate antiepileptogenic therapies administered shortly after TBI can prevent the development of recurrent, spontaneous seizures seen in this zebrafish model.
Conclusion
We validated a zebrafish model of severe brain-specific, closed-head TBI, which results in the development of drug-resistant epilepsy in less than 2 weeks. The negative impact on neuropsychological behaviors after TBI with or without ASMs parallels its effects on other mammalian models, including humans. Our data strongly support the utility of this model in studying PTE.
Footnotes
Funding Information
This work was supported by Canadian Institutes of Health Research (MFE-164732, S.-J.C.; PJT-148507 A.Y.R), National Research Foundation of Korea (NRF-2019R1A6A3A03032940, S.-J.C.), and Toronto General & Western Hospital Foundation (A.Y.R).
Author Disclosure Statement
No competing financial interests exist.