
Abstract
Traumatic brain injury (TBI) is defined as brain damage produced by an external mechanical force that leads to behavioral, cognitive, and psychiatric sequelae. The basolateral amygdala (BLA) is involved in emotional regulation, and its function and morphology are altered following TBI. Little is known about potential sex-specific effects of TBI on BLA neuronal function, but it is critical for the field to identify potential sex differences in TBI effects on brain and behavior. Here, we hypothesized that TBI would produce sex-specific acute (1 h) effects on BLA neuronal activation, excitability, and synaptic transmission in adult male and female rats. Forty-nine Wistar rats (n = 23 males and 26 females) were randomized to TBI (using lateral fluid percussion) or Sham groups in two separate studies. Study 1 used in situ hybridization (i.e., RNAscope) to measure BLA expression of c-fos (a marker of cell activation), vGlut, and vGat (markers of glutamatergic and GABAergic neurons, respectively) messenger RNA (mRNA). Study 2 used slice electrophysiology to measure intrinsic excitability and excitatory/inhibitory synaptic transmission in putative pyramidal neurons in the BLA. Physiological measures of injury severity were collected from all animals. Our results show that females exhibit increased apnea duration and reduced respiratory rate post-TBI relative to males. In male and female rats, TBI increased c-fos expression in BLA glutamatergic cells but not in BLA GABAergic cells, and TBI increased firing rate in BLA pyramidal neurons. Further, TBI increased spontaneous excitatory and inhibitory postsynaptic current (sEPSC and sIPSC) amplitude in BLA neurons of females relative to all other groups. TBI increased sEPSC frequency in BLA neurons of females relative to males but did not alter sIPSC frequency. In summary, lateral fluid percussion produced different physiological responses in male and female rats, as well as sex-specific alterations in BLA neuronal activation, excitability, and synaptic transmission 1 h after injury.
Introduction
Traumatic brain injury (TBI) is defined as brain damage produced as a consequence of an external mechanical force 1 that leads to behavioral, cognitive, and physical consequences. 2,3 TBI severity can be classified as mild, moderate, or severe. Clinical severity is based on different factors such as post-traumatic amnesia, loss of consciousness, and the Glasgow Coma Scale score in which patients are classified on the basis of clinical symptoms that are mild (score: 13-15), moderate (score: 9-13), or severe (score: <8). 4 Most TBIs are classified as mild, accounting for over 75% of all cases. 5 TBI is often referred to as a “silent epidemic” due to its high incidence, the lack of awareness of its impact, and the delayed evidence of its sequelae. 6 Frequent causes of TBI are falls, being struck by/against an object, traffic accidents, assaults, 7,8 sport-related activities, 9 and military combat-related activities. 10 TBI is a pathological process that leads to structural and functional brain damage. 11 While primary damage occurs at the moment of impact and results from mechanical forces that permeate brain tissue and disrupt brain function, secondary damage evolves over days to months and involves a series of molecular, biochemical, and cellular events that lead to tissue damage, cell death, and atrophy. 12 -14
The brain areas most vulnerable to TBIs are the frontal and temporal lobes, 15,16 the latter of which contains the amygdala, a brain region critical for stress processing and emotional memory. In addition to behavioral and cognitive problems, individuals with TBI exhibit increased risk for developing psychiatric disorders such as depression, anxiety, and substance use disorders. 17 -19 Pre-clinical work in our lab has shown that TBI, induced using the lateral fluid percussion model, increases alcohol self-administration 20 and anxiety-like behavior in adult male Wistar rats. 21 Additionally, clinical studies report sex differences in psychiatric disorders after TBI. For example, in one study, more women than men met the criteria for depression, post-traumatic stress disorder and other anxiety disorders, while more men than women met the criteria for substance use disorders. 22 These data highlight the need to investigate the neurobiological bases underlying potential sex differences in brain injury effects, as well as the potential impact of these sex differences on therapeutic strategies.
Focal and diffuse pre-clinical models of TBI produce morphological and functional changes in the basolateral amygdala (BLA), including altered excitability, 23,24 greater dendritic branch complexity, 25,26 and upregulation of N-methyl-D-aspartic acid or N-methyl-D-aspartate receptors. 27 These changes have been reported in rats during both early (1 day) 25 and late (11 days) phases of the post-TBI recovery period 24 using a lateral fluid percussion model. Hyperexcitability occurs when there is an imbalance between excitation and inhibition, in some cases as a result of disrupted GABAergic inhibitory control. 28 Hyperexcitability in the amygdala is a characteristic feature of affective disorders such as anxiety 29 and fear disorders, 30 and neuronal activation, as measured by fos protein or c-fos mRNA expression, is elevated in brain areas related to emotional behaviors (e.g., amygdala sub-regions) in rats after various stimuli. For example, increases in c-fos expression are observed in the BLA of rats after exposure to inescapable stress, 31 lithium chloride injections, 32 and aversive light in an open field. 33
Prior work examining TBI effects on amygdala structure and function has used either male or female rodents (mostly male), with no comparison between the two sexes, which is an important omission given the sexually dimorphic responses to TBI. 34 Here, we carried out two studies that aimed to test the acute (1 h) effects of TBI on BLA neuronal activation, as measured by c-fos, and on BLA neuron excitability and synaptic transmission, as measured by slice electrophysiology, in adult male and female Wistar rats. We hypothesized that TBI would produce sex-specific effects on neuronal activation and excitability, as well as synaptic transmission in BLA pyramidal neurons. Our results show that mild TBI produces sex-specific effects in the BLA and suggest that post-TBI neuropathology differs in males and females.
Methods
Animals
A total of 49 Wistar rats, male (n = 23) and female (n = 26), 3-4 months old at the time of surgery (Charles Rivers Laboratories, Raleigh, NC) were pair-housed under conditions of controlled temperature and humidity and exposed to a 12-h reverse light-dark cycle. Standard rat chow food and water were provided ad libitum. Upon arrival, animals were allowed to acclimate to the colony room for 1 week. All procedures were approved by the Institutional Animal Care Use Committee of the Louisiana State University Health Sciences Center (LSUHSC; New Orleans, LA) and were conducted in accordance with the National Institute of Health (NIH) guidelines.
Surgical procedure
Animals underwent craniotomy (2 mm posterior to bregma and 3 mm lateral to the midline; corresponding to the somatomotor cortex [SMC] area) before TBI, as previously described. 35,36 SMC was selected because previous studies in our laboratory have demonstrated that the amygdala is highly sensitive to lesions in this area, resulting in alteration of BLA-related behaviors such as anxiety-like behavior 21 and alcohol consumption. 20 Also, prior biochemistry experiments reveal that TBI in SMC increases pGLUR1S845 and pGRS232 levels in CeA 2 weeks after injury 21 and decreases 2AG levels in BLA 1 h after injury. 24 Animals were anesthetized (4% isoflurane in oxygen 2 L/min) for 4 min and then they were placed in a stereotaxic frame (model 900; Kopf Instruments, Tujunga, CA) with anesthesia (2-3% isoflurane in oxygen 1 L/min) through a nose mask. After the skull was exposed through an incision in the midline, a craniotomy (5 mm in diameter) was performed. A Luer Loc connector was positioned directly over the craniotomy and secured with cyanoacrylate glue. Dental cement (Lang Dental Manufacturing, Wheeling, IL) was applied around the female Luer Loc connector and allowed to harden prior to the Luer being filled with saline and capped. Animals were administered topical bupivacaine on the surgical area. Animals were recovered under temperature-controlled conditions before being returned to their individual home cages.
Traumatic brain injury (lateral fluid percussion)
Lateral fluid percussion injury (LFP) was performed as previously described by our laboratory. 35,36 This model is one of the most widely used in pre-clinical studies that reproduce the histopathology associated with TBI in humans. 37 Three days after craniotomy, animals were randomly assigned to TBI or Sham group. Animals were anesthetized with isoflurane (4% induction and 2-3% maintenance) and positioned in a stereotaxic frame, the female Luer Loc was connected to the LFP system (Fluid Percussion Injury, Model 01-B, Custom Design and Fabrication, Virginia Commonwealth University) by pressure tubing. The occurrence of mild, moderate, and severe TBI in rats using lateral fluid percussion has been described as 1.8-2.2 atm (26-32 psi), 2.4-2.8 (35-41 psi), and >2.8 atm (41 psi), respectively. 38,39 Anesthesia was lowered to 1% and after 1 min, a mild fluid percussion (26-32 psi) was delivered. Given that female animals in Study 1 had worse physiological response to TBI, we reduced the range of psi in females for Study 2, (26-28 psi), yet kept it within the range of mild TBI. 38,39 Animals were then monitored for signs of apnea and respiratory rate (breathing per minute). Sham animals underwent an identical procedure, but no lesion was produced. For Study 1, animals were sacrificed 1 h after TBI, by decapitation under deep isoflurane anesthesia, and brains were excised and frozen in methyl butane on dry ice and stored at -80°C until further processing. For Study 2, 1 h after TBI, animals were intracardially perfused with an N-methyl-D-glucamine (NMDG) based artificial cerebrospinal fluid (aCSF) under deep isoflurane anesthesia prior to brain excision.
Study 1
RNAscope assay
A total of 24 rats were used in this experiment RNAscope in situ hybridization is a method that allows visualization of single mRNA molecules in the cell through the use of specifically designed probes and a hybridization-based signal amplification system. 40 We used RNAscope markers for vGat and vGlut to “phenotype” the cells in BLA. Transcripts for vesicular glutamate and vesicular GABAergic transporter are reliable markers of glutamatergic and GABAergic neurons, respectively, and are detectable in the cell body. Rat brain coronal slices 10 μm thickness were obtained using a cryostat. Sections from 1.56 to 3.36 mm posterior to bregma 41 were collected and mounted onto Super Frost Plus (Fisher Scientific) slides. To ensure that the entire structure was sampled for our quantification, we divided the BLA into three portions, anterior-posterior (AP): anterior (-1,56, -2.04), middle (-2.16, -2.76), and posterior (-2.92, -3.36) using a rat brain atlas. 41 One slide from each depth was taken for quantification. RNAscope Multiplex Fluorescent Kit® (Advanced Cell Diagnostics, Newark, CA) was used according to the user manual for fresh frozen tissue.
Briefly, brain tissue was fixed in chilled (4°C) paraformaldehyde in 1 × phosphate-buffered saline (PBS; Fisher Scientific) for 15 min. Sections were dehydrated in ethanol (50%, 70%, 100%, 100%) for 5 min each. After dry, 2-4 drops of warmed protease 3 solution were placed on each slice for 45 min at room temperature. Slides were washed with agitation in 1 × PBS (2 × 2 min) and were covered with 80-100 μL of the probe for 2 h at 40°C in a HybEZ oven. Probes used in this experiment were c-fos (C1; Rn-Fos, Cat #403591), a marker of cell activation, vesicular glutamate transporter (vGlut; C2; Rn-Slc17a7, Cat # 317001), a marker of glutamatergic neurons, and vesicular GABA transporter (vGat; C3; Rn-SLc32a1, Cat # 424541) a marker of GABAergic neurons. Probes were diluted according to manufacturer's instructions. The C2 and C3 probes were mixed 1:50 with the C1 probe. Slides were removed from the oven and incubated in 5 × saline-sodium citrate buffer for 15 min; 2-4 drops of amplifier reagents were placed on the tissue followed by washes in wash buffer (2 × 2 min each). Incubation time was 40 min for Amp1, and 20 min for both Amp2 and Amp3.
Then, sections were incubated (20 min) with fluorescently labeled probes by selecting specific combination of colors associated with each channel green (Alexa 488 nm), orange (Atto 550 nm) and far red (Atto 647 nm). Amp4 Alt A was used to detect triplex c-fos, vGlut and vGat RNAs in green, orange, and far red, respectively. Slides were cover-slipped using a mounting medium containing 4′,6-diamidino-2-phenylindole (DAPI; ProLong Gold, Life Technologies Carlsbad) and left to dry at 4°C until visualization in the microscope. Negative controls were included and processed identically to ensure that there was no background noise and that the tissue was appropriately prepared. Universal negative control with a probe targeting DapB gene from a soil bacterium (Bacillus subtilis strain SMY) was used (RNAscope 3-plex Negative Control Probe, Cat # 320871).
Imaging processing and cell quantification
Images were captured with the ZEISS AxioScan.Z1 slide scanner and analyzed with Zeiss Zen Lite software. Negative controls were quantified first. For each cell, we counted the total pixels of the fluorescent signal (fluorescent dots) assuming that each pixel represents a single molecule of mRNA. The mean plus 3 standard deviations was used as threshold. The selection criterion for c-fos, vGlut, and vGat positive cells were ≥14, ≥ 11, and ≥10 positive pixels, respectively, after adjusting the threshold using Zeiss Zen Lite software.
Study 2
A total of 25 rats were used in this experiment. Electrophysiology brain slice preparation and recording were performed as previously described. 24,42,43 Briefly, brains were removed and sliced (Leica VT1200-S vibratome, Leica, Buffalo Grove, IL) in room temperature aCSF containing (in mM): 92 NMDG, 2.5 KCl, 1.25 NaH2PO4, 30 NaHCO3, 20 HEPES, 25 glucose, 2 thiourea, 5 Na-ascorbate, 3 Na-pyruvate, 0.5 CaCl2, 10 MgSO4; pH is titrated to 7.3-7.4 with concentrated hydrochloric acid 44 Coronal slices containing BLA were then transferred to a holding chamber (containing the same NMDG solution as slicing) in a water heat bath kept at 37°C. After 12 min, slices were removed from the holding chamber in the heat bath and allowed to rest at room temperature in a chamber containing holding solution for 45 min before the start of electrophysiological recordings. The holding solution consisted of (in mM): 92 NaCl, 2.5 KCl, 1.25 NaH2PO4, 30 NaHCO3, 20 HEPES, 25 glucose, 2 thiourea, 5 Na-ascorbate, 3 Na-pyruvate, 2 CaCl2, 2 MgSO4.
At the time of recording, slices were transferred to the slice chamber in an electrophysiology rig in a recording aCSF kept at 30-32°C by an in-line heater. The recording solution consisted of (in mM): 119 NaCl, 2.5 KCl, 1.25 NaH2PO4, 24 NaHCO3, 12.5 glucose, 2 CaCl2, 2 MgSO4. Whole–cell recordings were performed using borosilicate glass micropipettes (3-7 MOhm) filled with internal solution containing (in mM): 130 K-gluconate, 10 HEPES, 10 Na2-phosphocreatine, 4 MgCl2, 4 Na2-ATP, 0.4 Na-GTP, 3 ascorbic acid, 0.2 EGTA (pH 7.25, 290-295 mOsm). Pyramidal neurons on the side ipsilateral to the injury were identified and targeted for recordings based on cell body size and pyramidal-like somatic morphology. 45,46
Recordings were collected as previously described. 24,42 Electrical signals were amplified and digitized by a Multiclamp 700B amplifier (Molecular Devices, San Jose, CA). Recordings were sampled at 10 kHz and low pass filtered with a cutoff of 4 kHz. Signals were further filtered offline in Matlab with a cutoff of 2 kHz. Recordings were excluded if the series resistance was above 30 MOhm or the spike height, recorded in current clamp mode, was less than 60 mV. Synaptic properties were assessed by quantifying amplitude and rates of spontaneous excitatory and inhibitory postsynaptic currents (sEPSCs and sIPSCs). When whole–cell recording configuration was established, the voltage was clamped at -70 mV (for sEPSCs) or -50 mV (for sIPSCs) and 5 min of spontaneous postsynaptic currents (PSCs) were recorded. Spontaneous postsynaptic events were detected in these recordings by thresholding rapid excursions in inward current (@ -70 mV holding potential) or outward current (@ -50 mV holding potential); the average event amplitude and mean frequency over the 5 min recording period were quantified.
Excitatory to inhibitory (E/I) amplitude and frequency ratios were calculated as the ratio of mean parameters from neurons in which both recordings were collected. Estimated quantities related to intrinsic excitability included: resting membrane potential (RMP), input resistance, voltage sag, action potential firing in response to depolarization and action potential firing gain. Rebound response to hyperpolarization-activated inward current, known as the voltage sag, was calculated as ΔV2 / ΔV1, where ΔV1 is the difference between pre-step baseline voltage (-70 mV) and the steady-state voltage during step current injection, and ΔV2 is the difference between current step steady-state voltage and the minimum voltage during current injection. Voltage sag 47,48 is reflective of subthreshold current mediated by hyperpolarization-activated cyclic nucleotide–gated channels, also known as Ih . Input current steps of varying current amplitudes were delivered and the membrane voltage response to these current step inputs was recorded. Action potentials during each current step were counted and used to construct a firing rate-input (FI) curve. The FI curve gain was quantified by calculating the slope of the curve between 100 and 500 pA.
Statistical analysis
All data are expressed as mean ± standard error of the mean (SEM). In Study 1, injury pressure and apnea were analyzed with a two-tailed t-test, and respiratory rate with two-way analysis of variance (ANOVA), factors (Injury: Sham vs TBI; and Sex: male vs female). In Study 2, injury pressure and apnea were analyzed with a two-tailed t-test, and respiratory rate with two-way ANOVA, factors (Injury: Sham vs. TBI; and Sex: male vs. female). RNAscope data were analyzed with two-way ANOVA. Significant interactions were followed up using post hoc analysis with Tukey's multiple comparisons test. Statistical analyses were performed with GraphPad Prism 7 software (Graphpad Software Inc., La Jolla, CA). Electrophysiological data and statistical analysis were performed in Matlab (MathWorks, Natick, MA). Statistical comparisons of electrophysiological parameters were performed using two-way ANOVA, factors (Injury: Sham vs. TBI; and Sex: male vs. female). In the case of significant ANOVA result for at least one factor or interaction effect, post hoc comparisons were performed using Tukey's honest significant difference. For the firing rate input current curves, we performed an additional three-way repeated measures ANOVA where input current was the repeated measures within-subjects factor. 49
In the Sham male group, there are five rats with an average of six neural recordings per rat (N = 30 neurons). In the Sham female group, there are seven rats with an average of 4.7 neural recordings per rat (N = 33 neurons). In the TBI male group there are six rats with an average of 5.2 neural recordings per rat (N = 31 neurons). In the TBI female group there are seven rats with an average of 5.1 neural recordings per group (N = 36 neurons). Statistical analysis performed on electrophysiological data was done considering each recording parameter as an independent measurement (i.e., N = # of neurons). Neural properties exhibit significant heterogeneity, even within cell class and neural property heterogeneities can be important for determining neural circuit function. 50,51 One concern, however, is that collecting multiple samples from the same animal may result in pseudo-replication under certain conditions—specifically, when the intra-group variance of individual animal averaged parameters (animal means) is large relative to the individual data point variance within an animal there is an increased chance of obtaining type I errors (false positives). 52 One tool to assess this risk is calculating the intraclass correlation coefficient (ICC), which quantifies the relative mean discrepancy within groups. 53 Here, we calculate the ICC for every electrophysiological parameter that we found a significant two-way ANOVA effect.
Results
TBI effects on physiological responses in male and female rats
Immediately after TBI, animals were tested for physiological parameters including apnea and respiratory rate. Injury pressure was also recorded. Given that the range of psi in female animals of Study 2 was lower than the psi range in Study 1, we compared physiological responses in both studies to determine whether there was any significant difference in the injury response in rats between the two studies. In male animals, a two-tailed t-test revealed that males in both studies received injuries of equivalent pressure (t = 0.2282, p = 0.8241; Fig. 1A). A two-tailed t-test revealed that males in both studies had a similar duration of apnea (t = 0.4781, p = 0.6429; Fig. 1B). A two-way ANOVA revealed that there was no main effect of injury (F1,19 = 0.7297, p = 0.4036) nor a main effect of study (F1,19 = 0.7297, p = 0.4036) in the respiratory rate of male animals between Study 1 and Study 2, and there was no injury × study interaction effect (F1,19 = 1.605, p = 0.2205; Fig. 1C). In females, a two-tailed t-test revealed that females in both studies received injuries of equivalent pressure (t = 0.4348, p = 0.6729; Fig. 1D). A two-tailed t-test showed that there were no significant differences in the length of apnea in female animals in the two studies (t = 1.148, p = 0.2775; Fig. 1E). A two-way ANOVA showed a main effect of injury (F1,21 = 35.65, p = < 0.0001) but not a main effect of study (F1,21 = 1.728, p = 0.2028), and also an injury × study interaction effect (F 1,21 = 5.162, p = 0.0337) on respiratory rate (Fig. 1F). TBI animals from Study 1 exhibited fewer breaths per minute compared with sham animals of both studies. TBI animals from Study 2, had fewer breaths per minute compared with sham animals in Study 1.

Traumatic brain injury (TBI) effects on physiological parameters. Comparison by Study. There were no significant differences in the psi of males between Study 1 and Study 2 (p = 0.8241;
When we compared results by sex in Study 1, a two-tailed t-test revealed that males and females received injuries of equivalent pressure (t = 2.029, p = 0.0699; Fig. 2A). A two-tailed t-test revealed that female animals had significantly greater duration of apnea compared with males (t = 3.929, p = 0.0028; Fig. 2B). To determine the magnitude of physiological effects after normalizing for injury psi, we calculated the apnea/psi ratio; a two-tailed t-test showed increased apnea in female animals compared with males (t = 4.249, p = 0.0017; Fig. 2C). A two-way ANOVA showed a main effect of injury (F1,20 = 25.09, p = < 0.0001), a main effect of sex (F1,20 = 23.77, p = < 0.0001) , and an injury × sex interaction effect (F1,20 = 14.21, p = 0.0012) on respiratory rate. Post hoc comparisons showed that TBI female animals took fewer breaths per minute compared with all other groups (Fig. 2D).
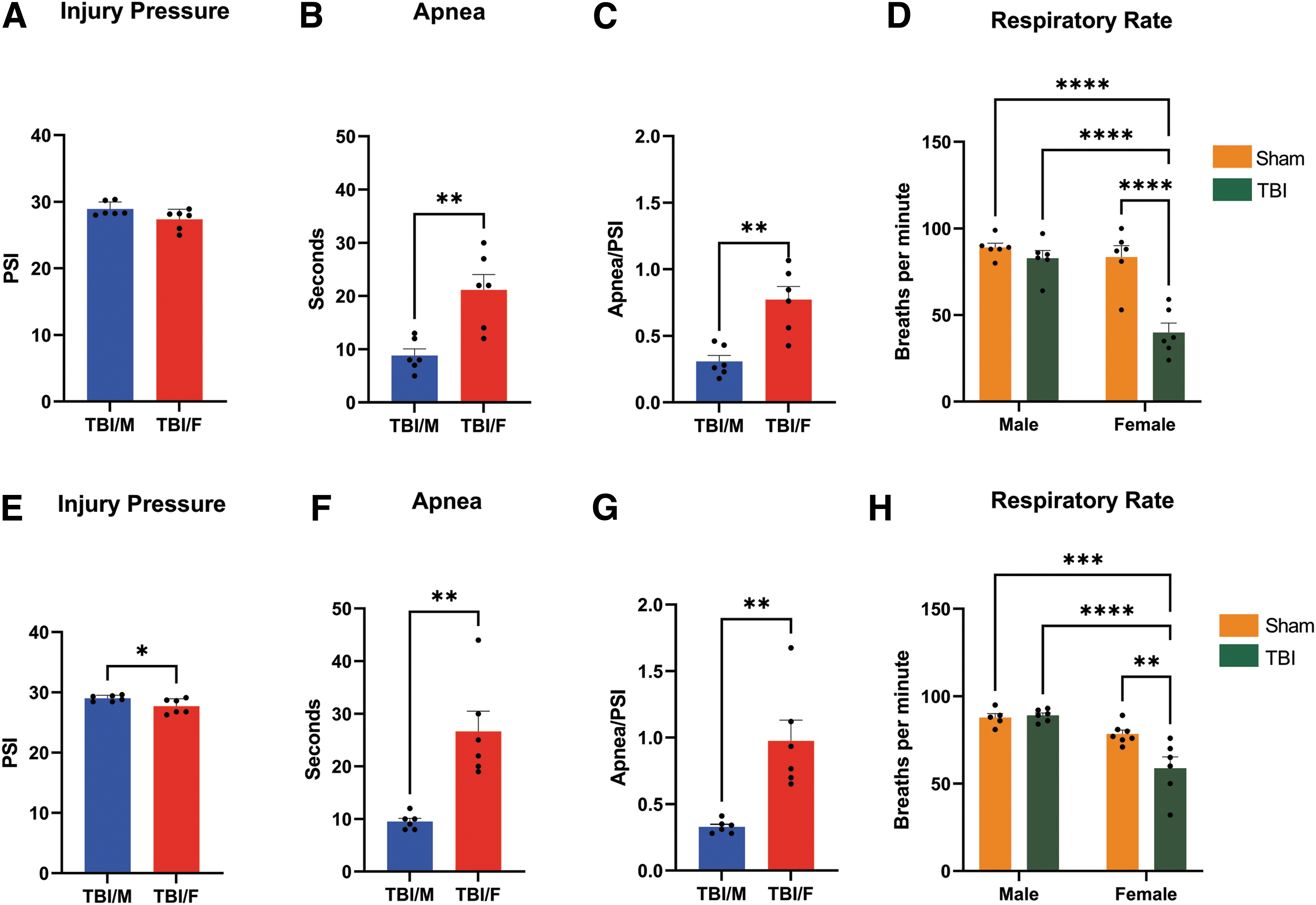
Traumatic brain injury (TBI) effects on physiological parameters. Comparison by sex. In Study 1, there were no significant differences between male and female rats in injury pressure received (p = 0.0699;
In Study 2, a two-tailed t-test revealed that males received a higher pressure compared with females (t = 2.389, p = 0.0386; Fig. 2E). A two-tailed t-test revealed that females had significant greater duration of apnea compared with male animals (t = 4.428, p = 0.0013; Fig. 2F). Again, we calculated the apnea/psi ratio, and a two-tailed t-test showed increased apnea in female animals compared with males (t = 4.088, p = 0.0022; Fig 2G). A two-way ANOVA showed a main effect of injury (F 1,20 = 6.196, p = 0.0217), a main effect of sex (F 1,20 = 28.62, p = < 0.0001), and an injury × sex interaction effect (F1,20 = 7.918, p = 0.0107) on respiratory rate. Post hoc comparisons showed that TBI female animals took fewer breaths per minute relative to all other groups (Fig 2H).
TBI increases glutamatergic BLA neuron activation
RNAscope was performed to test TBI effects on activation of glutamatergic and GABAergic neurons in the BLA (Fig. 3A) of adult male (sham: Fig. 3B1-3B4; TBI: Fig. 3C1-3C4), and female (sham: Fig. 3D1-3D4; TBI: Fig. 3E1-3E4) Wistar rats. Table 1 presents average vGlut+, vGlut+c-fos+, vGat+, and vGat+c-fos+ cell counts. A two-way ANOVA revealed a main effect of injury (F1,20 = 18.51; p = 0.0003) but not a main effect of sex (F1,20 = 2.802, p = 0.9958), nor an injury × sex interaction effect (F1,20= 1.148, p = 0.2966) on the percentage of c-fos–positive glutamatergic neurons in the BLA ipsilateral to the injury (Fig. 3F). A two-way ANOVA revealed a main effect of injury (F1,20 = 9.455, p = 0.0060) as well as main effect of sex (F1,20 = 8.337, p = 0.0091) but not an injury × sex interaction effect (F1,20 = 1.152, p = 0.2959) on the percentage of c-fos-positive glutamatergic neurons in BLA contralateral to the injury (Fig. 3G). When we analyzed GABAergic neurons, a two-way ANOVA revealed no main effect of either injury (F1,20 = 3,154, p = 0.0910) or sex (F1,20 = 0.4024, p = 0.5330), and no injury × sex interaction effect (F1,20 = 3,988, p = 0.0596) on the percentage of c-fos-positive GABAergic neurons in the BLA ipsilateral to the injury, (Fig. 3H). A separate two-way ANOVA revealed no main effect of injury (F1,20 = 0.8512, p = 0.3672) but a main effect of sex (F1,20 = 10.28, p = 0.0044) on the percentage of c-fos positive GABAergic neurons in BLA contralateral to the injury. There was not an injury × sex interaction effect (F1,20 = 2.303, p = 0.1448) on the percentage of c-fos positive GABAergic neurons in BLA contralateral to the injury (Fig 3I).
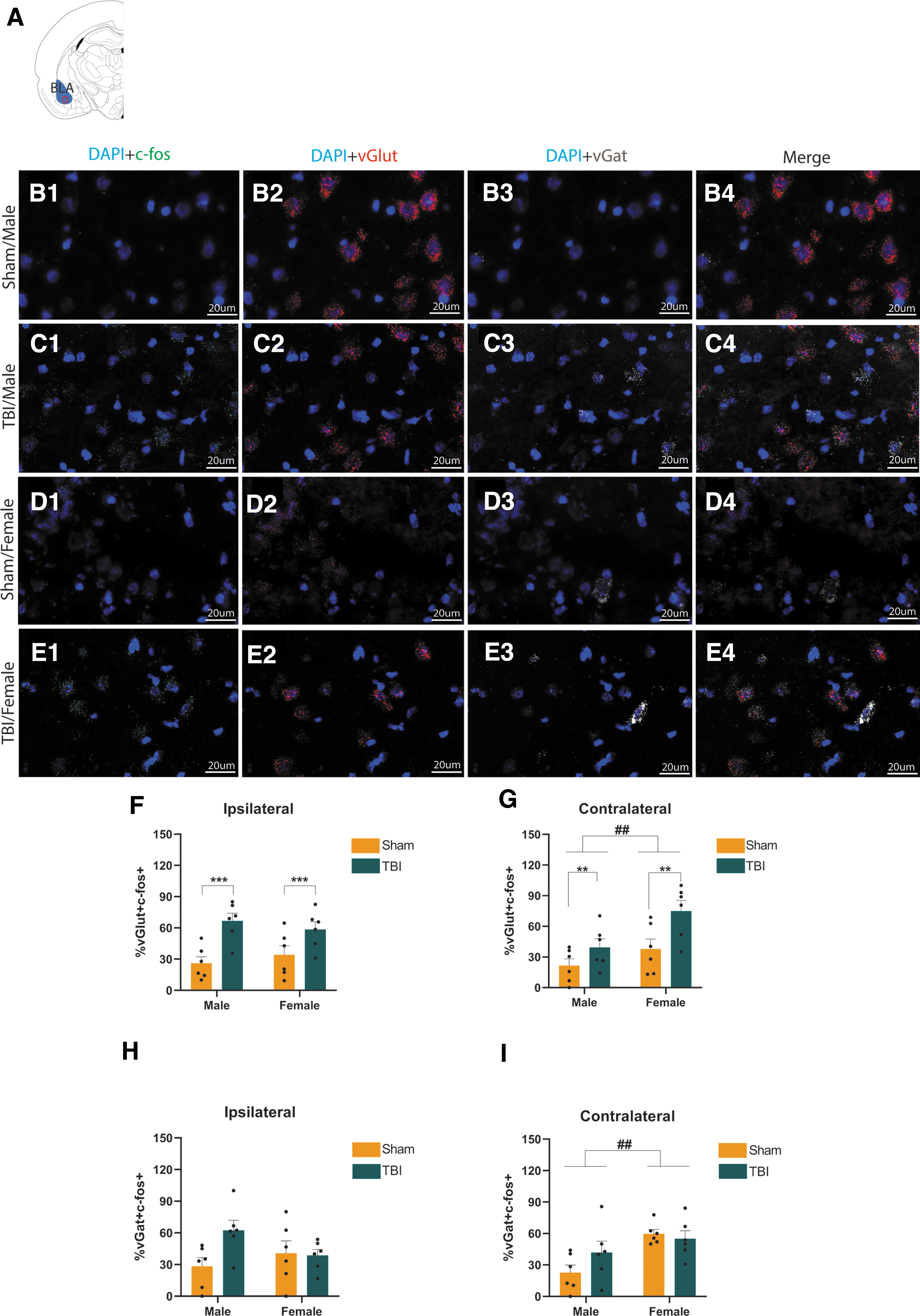
c-Fos expression in basolateral amygdala (BLA) glutamatergic and GABAergic neurons 1 h after traumatic brain injury (TBI). Representation of the BLA target area
41
RNAscope Data for Male and Female Rats
Data expressed as a mean ± standard error of the mean.
vGlut, vesicular glutamatergic transporter; vGat, vesicular GABAergic transporter; TBI, traumatic brain injury.
TBI alters intrinsic excitability of BLA pyramidal neurons in males and females
To test the prediction that TBI increases neuronal excitability in BLA pyramidal neurons, adult male and female Wistar rats in Study 2 were sacrificed 1 h post-TBI for whole-cell patch-clamp slice electrophysiology. Measures of input resistance (Rinput), resting membrane potential [RMP], sag, and firing rate were recorded. A two-way ANOVA revealed neither a main effect of injury (F1,122 = 0.01, p = 0.91) or sex (F1,122 = 0.34, p = 0.56), nor an injury × sex interaction effect (F1,122 = 1.47, p = 0.23) on RMP (Fig. 4A). Similarly, a two-way-ANOVA revealed no main effect of injury (F1,126 = 0.24, p = 0.63) or sex (F1,126 = 2.42, p = 0.12), nor an injury × sex interaction effect (F1,126 = 0.53, p = 0.47) on Rinput (Fig. 4B). Figure 4C shows the characteristic rebound voltage excursion seen in some neurons following hyperpolarization caused by negative current steps, known as voltage sag. A two-way ANOVA revealed a main effect of injury (F1,126 = 7.02, p = 0.0091) and sex (F1,126 = 12.27, p = 0.0006) but not an injury × sex interaction effect (F1,126 = 2.45, p = 0.12) on voltage sag recorded from hyperpolarization voltage responses in current-clamp mode. Post hoc pairwise comparisons revealed higher sag in TBI males compared with all other groups (Fig. 4D). Figure 4E shows membrane voltage responses to current step inputs.
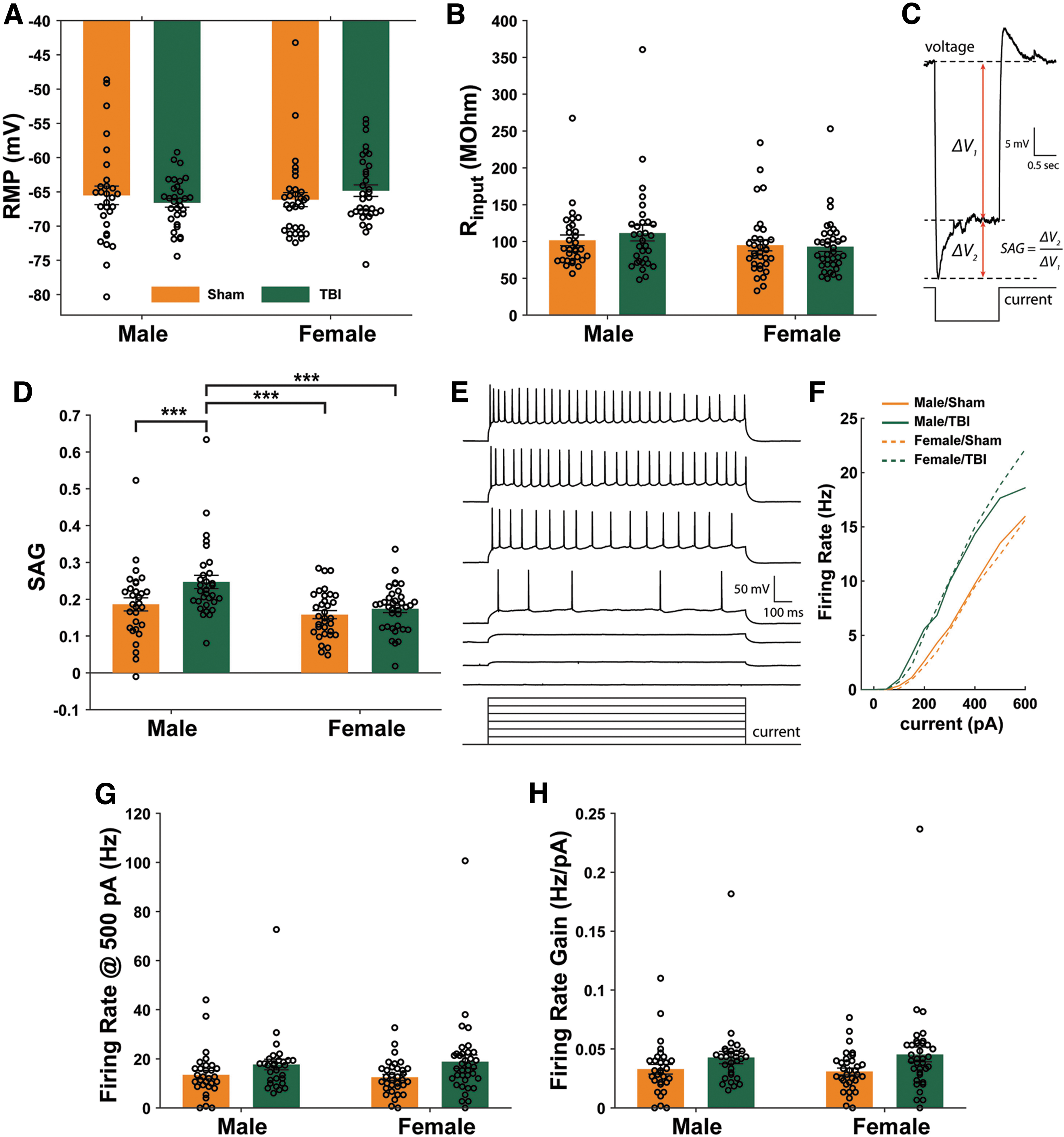
Traumatic brain injury (TBI) effects on basolateral amygdala neuron excitability 1 h after TBI. There was no effect of either injury (p = 0.91) or sex (p = 0.56) on the resting membrane potential (RMP) of recorded neurons
Figure 4F shows the FI curves constructed from the number of action potentials during each current step. The slope of this curve between 100 and 500 pA was used to calculate FI gain. A two-way ANOVA revealed a main effect of injury (F1,125 = 6.36, p = 0.013) on firing rate measured at 500 pA injection, but there were no post hoc pairwise differences between male and female groups (Fig. 4G). The ICC for firing rate was 0.065. A two-way ANOVA revealed a main effect of injury (F1,125 = 6.03, p = 0.015) on firing rate gain measured between 100 and 500pA, but again, this effect was apparent in males and females, with no post hoc differences between individual groups (Fig. 4H). The ICC for firing rate gain was 0.001.
TBI alters synaptic transmission in BLA pyramidal neurons in females
To test whether TBI alters synaptic transmission in the BLA of adult male and female Wistar rats, the frequency and amplitude of sEPSCs and sIPSCs were measured in BLA pyramidal neurons. Figure 5A shows representative traces of postsynaptic currents recorded at -70 mV and -50 mV in the same neuron, used to extract sEPSCs and sIPSCs, respectively. A two-way ANOVA revealed a main effect of injury (F1,117 = 8.93, p = 0.0034) and sex (F1,117 = 5.47, p = 0.021), as well as an injury × sex interaction effect (F1,117 = 5.65, p = 0.0191) on sEPSC amplitude. The ICC for sEPSC amplitude was 0.001. Post hoc pairwise comparisons revealed that sEPSC amplitude for BLA pyramidal neurons from TBI females was larger than for neurons recorded from animals in all other groups (Fig. 5B). A two-way ANOVA revealed a main effect of sex (F1,109 = 7.83, p = 0.0061), as well as an injury × sex interaction effect (F1,109 = 13.64, p = 0.0003) on sIPSC amplitude. The ICC for sIPSC amplitude was 0.001. Post hoc pairwise comparisons showed that sIPSC amplitude for BLA pyramidal neurons from TBI females was larger than for neurons recorded from animals in all other groups (Fig. 5C).
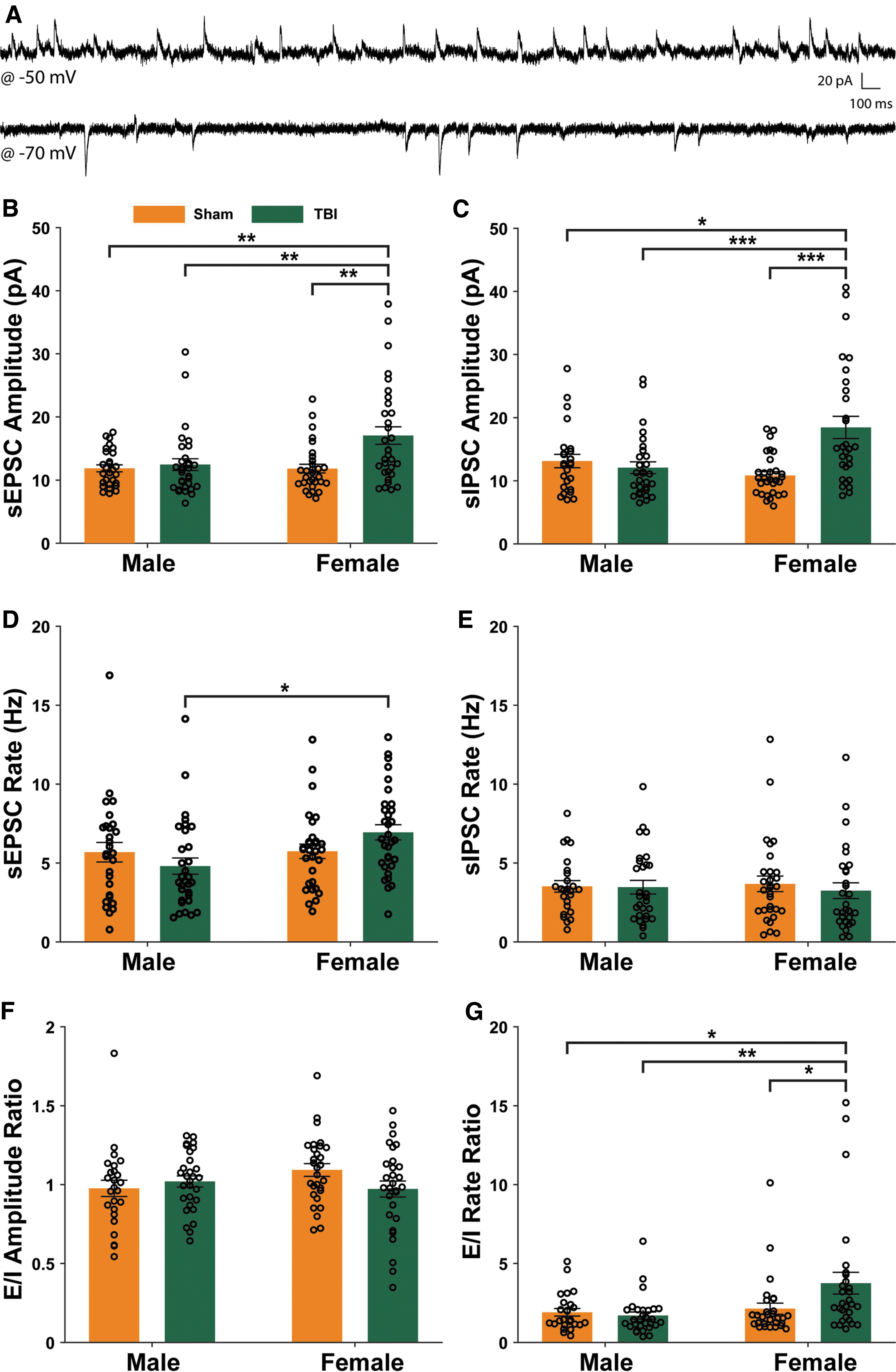
Traumatic brain injury (TBI) effects on synaptic transmission in basolateral amygdala pyramidal neurons 1 h after TBI. Representative traces of spontaneous postsynaptic current (sPSC) recordings from the same neuron with the voltage clamped at @ -50 mV or @ -70 mV during different recording periods
A two-way ANOVA showed a main effect of sex (F1,117 = 4.53, p = 0.0355), as well as an injury × sex interaction effect (F1,117 = 4.05, p = 0.0464) on sEPSC frequency. The ICC for sEPSC frequency was 0.119. Post hoc pairwise comparisons revealed that sEPSC frequency for BLA pyramidal neurons from TBI females was larger than for neurons recorded from TBI male rats (Fig. 5D). Finally, a two-way ANOVA revealed no effect of injury (F1.109 = 0.28, p = 0.60) or sex (F1.109 = 0.0, p = 0.95) on sIPSC frequency (Fig. 5E).
To examine the excitatory-inhibitory balance on a cell-by-cell basis, we calculated E/I balance of spontaneous post-synaptic current (sPSC) amplitude and frequency. A two-way ANOVA revealed no main effect of injury or sex on the excitatory to inhibitory ratio (E/I) of sPSC amplitudes (Fig. 5F), but there was a main effect of injury (F1,108 = 6.96, p = 0.0095), as well as an injury × sex interaction effect (F1,108 = 4.43 p = 0.038; Fig. 5G) on the E/I ratio of sPSC frequencies. The ICC for E/I frequency ratio was 0.042. Post hoc comparisons revealed that female animals had a higher E/I ratio of sPSC frequency compared with males and Sham female rats.
Discussion
We tested the hypothesis that TBI in the SMC has sex-specific effects on neuronal activation, as well as intrinsic excitability and synaptic properties of neurons in the BLA 1 h after injury. Our results show TBI increased apnea and reduced respiratory rate in females compared with males, suggesting exaggerated physiological responses to TBI in females, even when we normalized these outcomes to fluid delivery pressure. In Study 2, females received a lower range of psi injuries relative to Study 1, but in both studies, the psi used for both sexes was within the bounds of what is considered mild TBI in the literature (psi = 26-32). 38,39 When we compared the physiological response of females in both studies, there were no significant differences in the psi or apnea, which support the notion that females in both studies received similar mild TBI injuries. Further, we observed exaggerated physiological disturbances and neuronal changes in females after injury (relative to males), and these effects are unlikely to be explained by the lower range of psi delivered to females in Study 2. Female animals weighed less than male animals, but it is not clear whether this difference in body weight leads to altered physiological response to injury relative to males. We chose to match animals for age rather than body weight to avoid injury interactions with developmental stages. It is noteworthy that the lateral fluid percussion model compromises the brainstem 54 ; therefore, one may speculate that a smaller animal, receiving a mild TBI may be more susceptible to apnea regulated by the brainstem.
Sex affects TBI outcomes, 55 with an apparent discrepancy in the direction of sex differences reported in clinical and pre-clinical studies. In human studies, worse outcomes are reported in women compared with men: women exhibit greater deficits after TBI, recover more slowly, and report symptoms more frequently than men. 56 –59 However, results from pre-clinical studies seem to be more variable. In a recent systematic review, the authors found that among 43 animal studies, 44% of studies reported that females had better outcomes than males, whereas 14% reported that females had worse outcomes, and 28% reported no sex differences. 60 Nevertheless, results in pre-clinical studies seem to depend on variables such as the type of TBI model used, the specific outcome measured, and time of testing or sacrifice. For example, 55% of controlled cortical impact (CCI) studies reported that females have better outcomes, while only 40% of lateral fluid percussion studies reported that female animals have better outcomes. 60 Lateral fluid percussion is a mixed model of focal and diffuse injury. 61,62 Diffuse injury results in widespread damage in the brain that affects axons, vasculature, and hypoxia. 63 It is possible that female animals are more vulnerable to this type of injury than males.
Our results show that TBI results in activation of BLA neurons. Specifically, we found that TBI increases activation of glutamatergic cells, evidenced by an increase in the percentage of glutamatergic BLA neurons with co-localized c-fos (a marker of neuronal activation) in both sexes, with no effect on c-fos expression in GABAergic cells in either sex. BLA neurons are approximately 80-90% glutamatergic, with a smaller percentage of GABAergic interneurons, and glial cells. 64 The functional balance between glutamatergic and GABAergic neuron activity is fundamental for normal BLA function. TBI transiently disrupts the cell membrane, leading to changes in ionic distribution and alterations in membrane potential. 65 In the acute phase following TBI (∼1 h) there is a large increase in glutamate release, 65 which plays a role in the pathophysiology of TBI given that excess glutamate release produces excitotoxicity leading to neuronal damage and death.
Our results demonstrate that TBI alters intrinsic excitability parameters, such as firing rate and sag in BLA neurons. Prior work from our group suggests that increases in BLA excitability after TBI is sustained for at least 11 days in female rats. 24 Specifically, we previously reported that action potential firing gain in response to injected current was higher in BLA neurons from female rats that received moderate TBI. Our current findings indicate that 1 h after injury, firing rate and firing rate gain are higher in BLA neurons from injured male and female rats relative to sham controls. TBI also increased the amplitude of sEPSCs and sIPSCs in female rats relative to injured males and relative to sham controls. Finally, our results show that TBI increased the frequency of sEPSCs in females compared with males. We also observed TBI-induced increases in the E/I ratio of sPSC rates in BLA pyramidal neurons recorded from females. Altogether, these changes suggest that mild TBI produces rapid changes in the intrinsic and synaptic properties of BLA neurons, and that TBI induces pre- and post-synaptic alterations in synaptic transmission in the BLA, although it remains to be determined how cortical TBI produces these effects in the BLA.
Several studies have shown that TBI alters neuronal excitability, but the underlying mechanisms are not completely understood. It is also not clear how TBI produces rapid (e.g., 1 h as in the current study) and lasting (e.g., 11 days as in prior work from our group 24 ) alterations in synaptic function in the amygdala—it is possible that rapid and persistent mechanisms may be the same or different. It is possible that TBI alters BLA function via mechanical forces. For example, two mechanical phenomena leading to primary injury are direct contact and propagation of stress waves from the site of injury through the brain and movement of the brain due to inertial or acceleration loading. 66 The lateral fluid percussion model uses rapid propagation of a pressure wave through fluid into the open skull (craniotomy) 67 that results in temporary displacement of the brain relative to the inner surface of the skull, and mechanical deformation. 61 It is possible that those mechanical forces directly impact the amygdala and trigger neuropathological alterations that are manifested in the acute and chronic phases after injury. One potential limitation of this study is the fact that all electrophysiological data were collected only ipsilateral to the side of the lesion; this also precludes comparison between the two studies performed here.
Pre-clinical studies have reported alterations in cortical excitability after injuries induced by focal cortical compression, 68 lateral fluid percussion, 69,70 or weight drop 71 at different time-points. For example, using a multi-electrode array to record neural activity in the primary somatosensory cortex to vibrissa deflection, Ding and colleagues 68 found a period of neuronal inhibition immediately after the injury onset followed by increased neuronal excitation in the granular layer, 2 h following injury. Measuring firing rate 24 h after TBI, Johnstone and colleagues 71 found in vivo hypo-excitation in cortical neuronal response in response to simple and complex whisker deflections from the supragranular cortical layer through sub-granular layer IV. In addition, those authors reported increased spontaneous firing rate of neurons in cortical layers IV and V. Alwis and colleagues 72 found that 8-10 weeks after TBI, there was hyper-excitation in response to sensory input in the supragranular cortical layers which was more marked after complex stimuli. Finally, using a controlled cortical impact model, Almeida-Suhett and colleagues 23 found decreased inhibitory signaling in the BLA 7 days after injury suggesting increased excitability in this structure following TBI. Based on these results, we speculate that alterations in neuronal excitability in cortical areas likely play a role in altered excitability of neurons in the BLA, a region that does receive (apparently sparse) inputs from SMC (the site of injury in this study).
It is possible that primary and secondary injuries both play a role in the physiological changes that we observed in our model of TBI 1 h after injury. Secondary injury typically starts minutes to hours after impact and involves damage to the blood–brain barrier (BBB), excitotoxicity, neuroinflammation, oxidative stress, axon degeneration, and cell death. 73 Neuroinflammation is a major factor in TBI pathophysiology that initiates after impact, 74 persists up to 17 years after injury in humans, 75 and involves immune cells, microglia, cytokines, and other inflammatory mediators some of which increase cellular excitability. 76 Further, studies using lateral fluid percussion have shown BBB disruption 3 to 60 min post-TBI, with effects on various brain regions in the lesioned hemisphere. 77 Increases in BBB permeability allow brain infiltration of leukocytes (e.g., neutrophils and monocytes) 78 that secrete chemokines, reactive oxygen species, and cytokines that contribute to prolonged inflammation. 79 Experimental models of TBI have demonstrated elevated levels of cytokines in the brain: for example, interleukin-1β, a pro-inflammatory cytokine produced by microglia, astrocytes and neurons, is upregulated in rodent models as early as 1 h post-TBI, 80,81 and the same pattern of effects is observed for tumor necrosis factor α. 82 –84 Proinflammatory cytokines can alter the gating properties of voltage-gated ion channels, leading to hyperexcitability and changes in synaptic transmission. 85 TBI-induced inflammation may increase neuron excitability both at site of injury (e.g., SMC in this study) and in sub-cortical brain regions (e.g., BLA), and ongoing work in our laboratory is focused on examining this possibility.
In conclusion, results from this study show sex-specific effects of TBI on physiological parameters and BLA neuronal activity 1 h following mild TBI. In both males and females, mild TBI produced increases in the nuclear expression of a neuronal activation marker (i.e., c-fos) in glutamatergic but not GABAergic neurons in the BLA, and also increased firing rate in BLA pyramidal neurons. However, only females exhibited increases in synaptic transmission 1 h after TBI. Future studies will examine potential sex differences in TBI effects on the activity and activation of BLA input circuitry at early and late post-injury time points, as well as the role of these changes in mediating post-TBI behavioral outcomes.
Footnotes
Acknowledgments
We would like to thank Dr. Marcus Weera and Dr. Maria Elena Secci for technical assistance for the RNAscope study.
Authors' Contributions
AJ-S was responsible for performing experiments and collecting data, data analysis, and writing/editing the manuscript. JWM, was responsible for performing experiments and collecting data, data analysis, and writing/editing the manuscript. ZFS was responsible for performing experiments and collecting data. SE was responsible for conceptualization and study design. PEM was responsible for conceptualization, study design, writing/reviewing/editing the manuscript, and funding acquisition. NWG was responsible for conceptualization, study design, writing/reviewing/editing the manuscript, and funding acquisition.
Funding Information
Support for this study was provided by National Institute on Alcohol Abuse and Alcoholism, NIH/ NIAAA research grant 1R01AA025792-01A1. This work was also supported in part by a Merit Review Award #I01 BX003451 (to NWG) from the United States Department of Veterans Affairs, Biomedical Laboratory Research and Development Service.
Author Disclosure Statement
NWG holds shares in Glauser Life Sciences, Inc. SE is on the Scientific Advisory Board of Avanos. These activities have no relation to any of the work presented in this paper. No competing financial interests exist for AJ-S, JWM, ZFS and PEM.