
Abstract
The inflammatory response associated with traumatic spinal cord injury (SCI) contributes to locomotor and sensory impairments. Pro-inflammatory (M1) macrophages/microglia (MϕMG) are the major cellular players in this response as they promote chronic inflammation resulting in injury expansion and tissue damage. Fatty acid-binding protein 4 (FABP4) promotes M1 MϕMG differentiation; however, it is unknown if FABP4 also plays a role in the etiology of SCI. The present study investigates whether FABP4's gene expression influences functional recovery following SCI. Analysis of quantitative polymerase chain reaction data shows a robust induction of FABP4 messenger RNA (mRNA; >100 fold) in rats subjected to a T9-T10 contusion injury compared with control. Western blot experiments reveal significant upregulation of FABP4 protein at the injury epicenter, and immunofluorescence analysis identifies that this upregulation occurs in CD11b+ MϕMG. Further, upregulation of FABP4 gene expression correlates with peroxisome proliferator-activated receptor γ (PPARγ) downregulation, inactivation of Iκβα, and the activation of the NF-κB pathway. Analysis of locomotor recovery using the Basso-Beattie-Bresnahan's locomotor scale and the CatWalk gait analysis system shows that injured rats treated with FABP4 inhibitor BMS309403 have significant improvements in locomotion compared with vehicle controls. Additionally, inhibitor-treated rats exhibit enhanced autonomic bladder reflex recovery. Immunofluorescence experiments also show the administration of the FABP4 inhibitor increases the number of CD163+ and liver arginase+ M2 MϕMG within the epicenter and penumbra of the injured spinal cord 28 days post-injury. These findings show that FABP4 may significantly exacerbate locomotor and sensory impairments during SCI by modulating macrophage/microglial activity.
Introduction
Meta-inflammation following spinal cord injury (SCI) results in impaired axonal regeneration and functional recovery. Accumulating evidence demonstrates the release of pro-inflammatory free fatty acids (FFAs) following SCI promotes M1-MϕMG differentiation and deters anti-inflammatory (M2) presence. 1 –7 Studies using chimeric animal models to ascertain the role of macrophages or microglia in SCI demonstrate a marked difference in their function and distribution within the injured spinal cord. 8,9 High-resolution imaging data show pro-inflammatory macrophages residing in the gray matter promote axonal degeneration and injury expansion, resulting in locomotor and sensory deficits. 9 –13 The activity of M1 microglia in the white matter, however, induces neurotoxic A1 astrocyte differentiation through the secretion of interleukin (IL)-1α and tumor necrosis factor (TNF)α, resulting in the death of axotomized neurons and differentiated oligodendrocytes. 8,9
Current U.S. Food and Drug Administration–approved standard of care for the treatment of SCI seeks to modulate inflammation by using the immunosuppressant methylprednisolone. Notably, this course of treatment is not shown to improve patients’ locomotor or sensory function and has been associated with higher rates of complications, which indicates that suppressing inflammatory activity does not improve functional outcomes. 14 Additionally, there is still significant debate in the field regarding the effect of eliminating macrophage or microglial activity following SCI and whether this is detrimental or beneficial to recovery. 8,15 -17 In contrast, peripheral nerve injury (PNI) data demonstrate that, instead of ablation, immunomodulatory factors that prompt M2 differentiation of MϕMG can facilitate tissue repair, remodeling, membrane reconstruction, and also prevent the onset and progression of neuropathic pain. 18 –21 Considering the promising outcomes of MϕMG immunomodulation in PNI, further research is needed to elucidate the potential benefits of modifying the activity of these cells to promote recovery after SCI. 11,22 –25
Fatty acid-binding protein 4 (AFABP or aP2) is a cellular transporter of long-chain FFAs that modulates the inflammatory activity and differentiation of macrophages and microglia. 26 -28 FABP4 regulates lipid signal transduction in the nucleus through its binding and transport of pro-inflammatory n-6 polyunsaturated fatty acids (n-6 PUFAs). 29 -31 Importantly, pro-inflammatory lipid stimulation increases the expression of FABP4 during microglial-mediated neuroinflammation and in macrophages of atherosclerotic plaques. 28,32 In the context of atherosclerosis, elevated FABP4 expression initiates a feedback loop with c-Jun N-terminal kinase (JNK) and activator protein-1 (AP-1), leading to the activation of pro-inflammatory pathways and the release of TNF-α, IL-1β, and IL-6. 32,33 Moreover, studies investigating the effects of obesity on microglial activation reveal FABP4 upregulation promotes NF-κB pathway activation by inhibiting the uncoupling protein 2–redox mechanism, resulting in the dysfunction or death of hypothalamic neurons. 28
In contrast, FABP4-deficient MϕMG exhibit altered pro-inflammatory responsiveness with reduced NF-κB activity and decreased production of TNF-α, inducible nitric oxide synthase, IL-1β, and IL-6. 17,18,28,32 Additionally, FABP4 inhibition in macrophages enhances peroxisome proliferator-activated receptor γ (PPARγ) receptor activity and elicits an anti-inflammatory cellular response which promotes anti-inflammatory M2 differentiation. 26,28,29,34 In fact, clinical studies in patients with SCI list FABP4 as a biomarker of recurrent pressure injury and deep tissue injury risk, and highlight the preventative potential of modulating this protein both locally and systemically to avoid these co-morbidities. 35 While the effects of FABP4 upregulation in metabolic diseases have been well studied, the outcomes resulting from FABP4 over-expression or inhibition in the progression of spinal cord injury are unknown. The present study examines the gene expression of FABP4 following SCI to determine whether it may play a role in recovery.
Methods
Animals
All animal studies were performed in compliance with the Loma Linda University School of Medicine regulations and institutional guidelines consistent with the National Institutes of Health Guide for the Care and Use of Laboratory Animals. The present study used 220-225 g female Sprague-Dawley rats (n = 70; Charles River Laboratories, Portage, MI) housed in individual cages on alternating 12-h light/dark cycles with food and water ad libitum.
Spinal cord contusion injury and BMS309403 treatment
Two different cohorts were used in this study. In the first cohort, rats were anesthetized, and spinal cord injury was generated using the Multicenter Animal Spinal Cord Injury Study (MASCIS) Impactor as previously described. 36 -38 Animals received a moderate contusion injury at T9-T10. The MASCIS software values for compression and compression rate ensured injury severity and reproducibility. Only animals with a compression value of 0.397 ± 0.0303 and a compression rate of 1.924 ± 0.2134 were included in the study. Sham controls only received a T9-T10 laminectomy. For both sham and injured groups, 5 mm lesion epicenter segments were harvested 1, 3, 7, 14, and 28 days post-injury (dpi). A minimum of three animals were used for each time-point (see figure legends for n at each time point).
The second cohort of animals was injured as described above and concomitantly treated with the FABP4 small molecule inhibitor BMS309403, which targets the fatty acid-binding pocket. 39 A small subcutaneous pocket over the sacral vertebrae was created to house a primed osmotic mini-pump (DURECTTM, Cupertino, CA). The pump was previously filled with 200 μL of 100 μM BMS309403 solution (inhibitor), or 100 μM ethanol solution (vehicle) dispensed to the epicenter at a mean flow rate of 0.25 μL/h. Surgeries, pump preparation, and pump placement were conducted in the manner previously reported. 36 Behavioral studies were conducted 7, 14, 21, and 28 dpi. The Crede's maneuver was performed twice per day to express retained urine from the bladder of all injured rats in the study. Autonomic bladder recovery was assessed by measuring expressed urine volume (in mL) of vehicle and inhibitor-treated rats for 9 days. Bladder function was restored once expressed urine volume was ≤0.5 mL. From this cohort, 7 mm lesion epicenter segments were harvested 28 dpi. Post-operative care and euthanasia for all animals were conducted as previously reported. 36,37
Quantitative real-time polymerase chain reaction
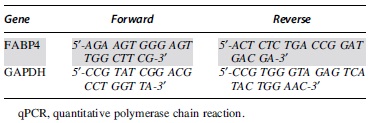
Complementary DNA was amplified by real-time polymerase chain reaction (PCR) using primers for the detection of FABP4 and glyceraldehyde 3-phosphate dehydrogenase (GAPDH; Table 1), which served as the internal control. Amplification was conducted using the CFX96 Real-Time PCR Detection System (Bio-Rad Laboratories, Hercules, CA). Relative gene expression levels were normalized to control (median sham).
qPCR Gene Expression Analysis Primers
Western blot
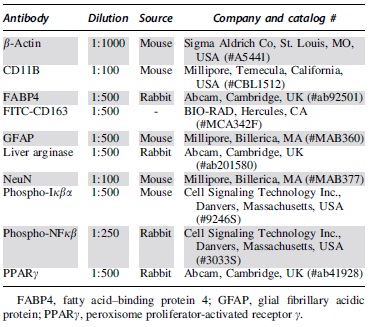
Western blots were conducted as previously described. 3,36,40 Briefly, membranes were incubated overnight with rabbit anti-FABP4, anti-rabbit PPARγ, anti-rabbit P-NF-κB, or mouse anti-P-Iκβα and mouse anti-β-actin (Table 2). Secondary anti-rabbit 800 and anti-mouse 680 antibodies (LI-COR® Biosciences, Lincoln, NE) were applied to the membranes for detection. Finally, membranes were imaged using the Odyssey® Infrared Imaging System (LI-COR Biosciences). Relative protein levels were normalized to β-actin, and fold change was assessed as the level of FABP4 protein over control (median sham). Image Studio™ Lite Software was used for protein quantification (LI-COR Biosciences).
Western Blot and Immunofluorescence Analysis Antibodies
Behavioral evaluation of spontaneous locomotion
The Basso-Beattie-Bresnahan (BBB) 22-point locomotor scale was used to assess spontaneous open-field locomotion 7, 14, 21, and 28 dpi. 41 Before testing, animals were acclimated to the open-field environment, and baseline values were taken. Testing sessions were recorded and evaluated as previously described. 36,37 Two blinded observers scored the degree of locomotor function and recovery. Post hoc transformation of scores was conducted to improve the scale's metric properties and reduce scale-introduced variability. 42
Catwalk analysis
The CatWalk Gait Device (Noldus Information Technology Inc., Leesburg, VA) was used to analyze the walking patterns of animals during conditioned locomotion. 43 -45 Each animal's baseline performance was acquired before the injury. The locomotor recovery of BMS309403-treated rats and vehicle controls was assessed after injury at 7, 14, 21, and 28 dpi. Average values for baseline and time-points were gathered for analysis. The Catwalk software (ver. 20.6) classified front paws and hind paws by adjusting intensity thresholds. Significant differences in regularity index, average speed, cadence, maximum variation, and step variation are presented as indicators of locomotor recovery.
Histology
Spinal cord epicenters were prepared as previously described. 36 -38 Longitudinal sections of 20 μm thickness were double-labeled with rabbit anti-FABP4 and mouse anti-CD11b (monocytes, macrophages/ microglia) or with mouse anti-NeuN (neurons) or mouse anti-glial fibrillary acidic protein (GFAP; activated astrocytes; Table 2). Rabbit anti-FABP4 was also used to quantify levels of FABP4 protein in the second cohort. Mean fluorescent intensity was calculated from each animal's averaged fluorescence intensity values. Additional slides from this cohort were used to stain MϕMG using anti-mouse FITC-CD163 and rabbit anti-liver arginase (Table 2). Sections were incubated with the appropriate combination of Alexa Fluor 488 conjugated donkey anti-rabbit antibody, Alexa Fluor 594 conjugated donkey anti-mouse (1:1000; Invitrogen, Carlsbad, CA). Cell nuclei were stained using Invitrogen™ Prolong™ Gold Antifade Mountant with DAPI. Sections were imaged using the confocal laser scanning microscope (FV1000, Olympus or LSM710; Carl Zeiss GmbH, Jena, Germany) or fluorescence microscope (BZ9000; Keyence Corporation, Osaka, Japan). A minimum of three sections were analyzed per animal. Images were analyzed and prepared for publication using ImageJ software (National Institutes of Health, Bethesda, MD).
Statistical analysis
All statistical analyses were performed using Prism Software (GraphPad Software Inc., San Diego, CA). Outliers were identified using the Grubbs’ method. All data are presented as mean ± standard error of the mean. Differences were considered statistically significant at p < 0.05.
Results
FABP4 gene expression is significantly upregulated following SCI
The first series of experiments used quantitative polymerase chain reaction analysis to determine FABP4 mRNA levels in SCI. The data show a modest increase of FABP4 mRNA 1 dpi and a robust 50-100 fold induction at 3 (**p = 0.004), 7 (****p < 0.0001), 14 (*p = 0.017), and 28 dpi (***p = 0.0005; Fig. 1B). Basal expression of FABP4 mRNA was observed in sham animals. As mRNA upregulation is not always translatable to protein, 46 we conducted Western blot analysis to ascertain the spatiotemporal gene expression of FABP4. We found that FABP4 protein levels in the injury epicenter were induced 20-40-fold (Fig. 1C, 1D) at 7 (****p < 0.0001) and 28 dpi (*p = 0.0191) compared with control.

Time-course messenger RNA (mRNA) and protein analysis show significant gene upregulation of fatty acid-binding protein 4 (FABP4) at the injured spinal cord epicenter.
FABP4 gene expression in CD11b+ macrophages/microglia following SCI
Double staining of 7 dpi tissue was used to determine FABP4 expression within CD11b+ MϕMG, and GFAP+ activated astrocytes or NeuN+ neurons in the ventral gray matter (VGM). 47 The double immunostaining data of sham tissues confirms a previous report 48 suggesting a significantly low or undetectable basal expression of FABP4 in normal tissues in the central nervous system. In contrast, the data for this time-point indicate only CD11b+ macrophages/microglia, not neurons or activated astrocytes, have elevated FABP4 protein expression (Fig. 2A-C).
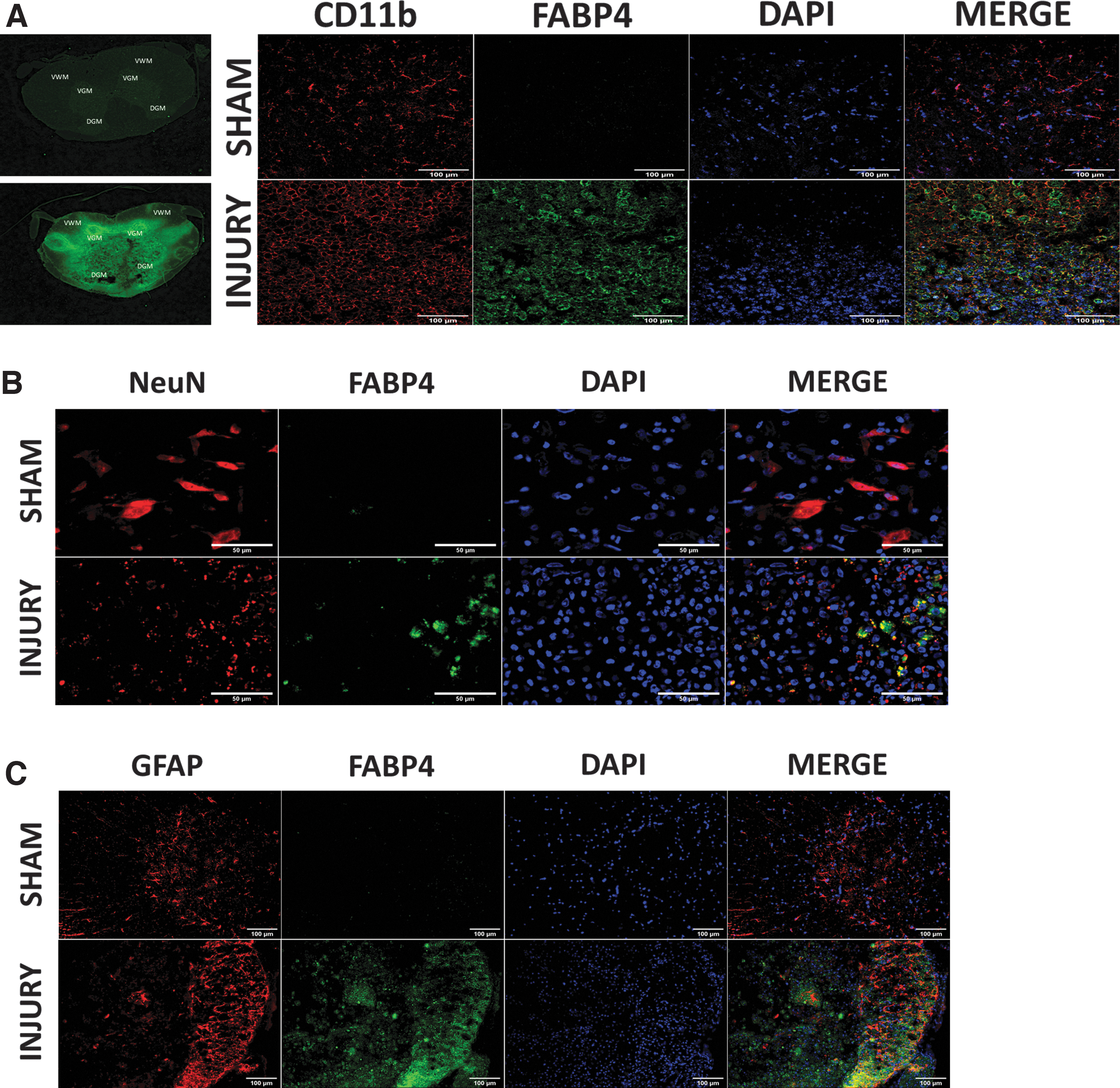
Fatty acid-binding protein 4 (FABP4) expression in CD11b+ macrophages/microglia following spinal cord injury.
Increased FABP4 expression is correlated to PPARγ regulation and levels of phosphorylated NF-κB and Iκβα
Temporal analysis of PPARγ regulation following injury shows significant differences in protein expression between sham and injury (*p = 0.0482), particularly between 7 dpi and 7 (**p = 0.0054) day sham. Interestingly, there were also significant differences between PPARγ upregulation 7 dpi and robust downregulation 3, 14, and 28 dpi (**p = 0.0028, **p = 0.0029, and *p = 0.0159 respectively; Fig. 3A, 3C). Moreover, levels of phosphorylated (activated) NF-κB were significantly increased in injured animals (***p = 0.0011) particularly 7 (**p = 0.0056) and 28 dpi (**p = .0056). Interestingly, activation of NF-κB was concurrent with increased phosphorylation (inactivation) of Iκβα in injured animals compared with sham (***p = 0.0008) at both 7 (**p = 0.0049) and 28 dpi (**p = 0.0049; Fig. 3B, 3C).
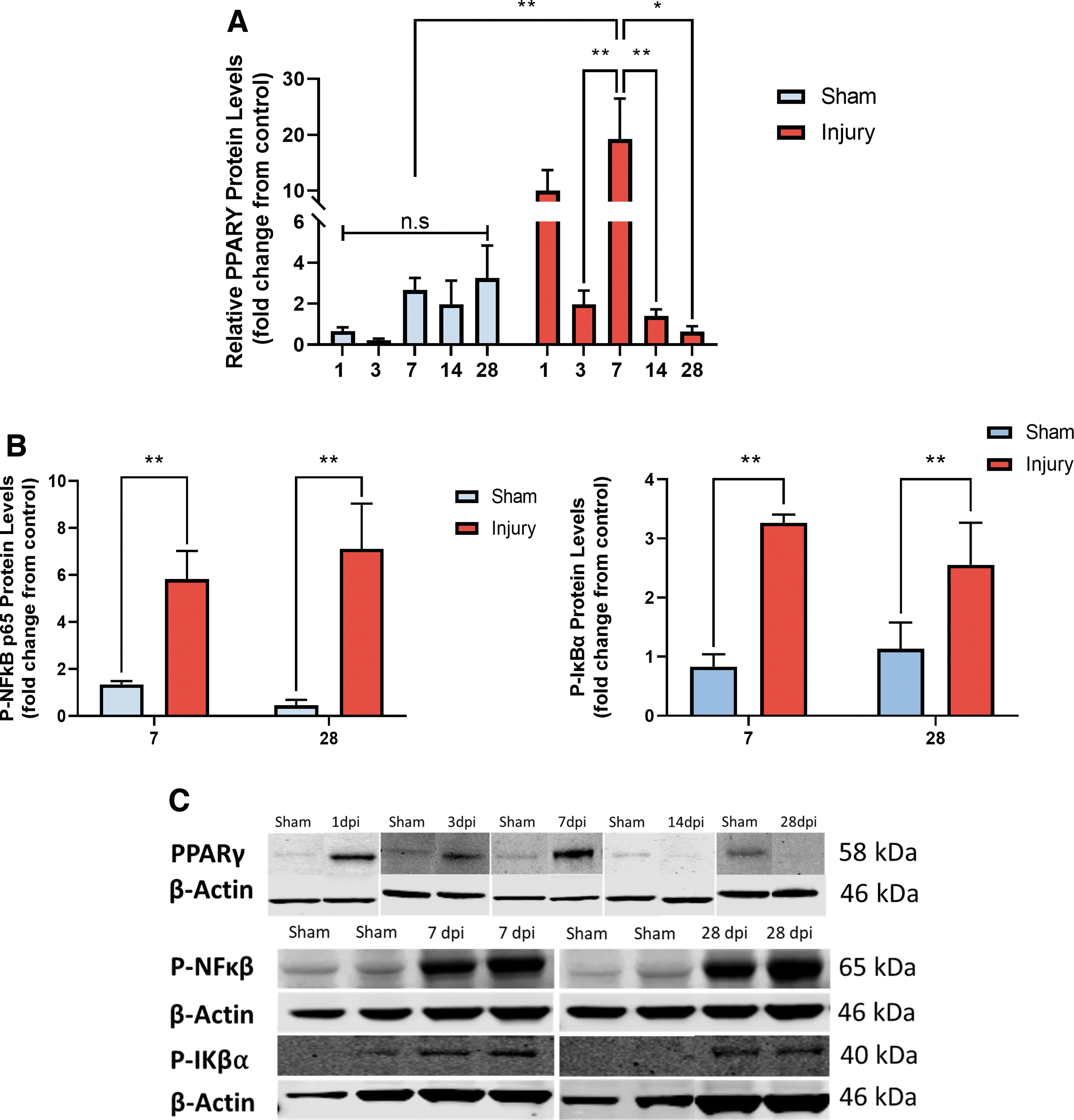
Increased fatty acid-binding protein 4 (FABP4) levels correlate with time-points of significant peroxisome proliferator-activated receptor γ (PPARγ), P-Iκβα, and P-NF-κB regulation.
Administration of FABP4 inhibitor BMS309403 promotes locomotor recovery following spinal cord injury
The inhibitory efficacy of BMS309403 was validated through immunofluorescence. The data indicate that administration of BMS309403 significantly downregulates FABP4 protein expression in treated animals (Fig. 4B, 4D). Administration of BMS309403 did not affect levels of FABP5 (Fig. 4C, 4D), a protein also expressed in both macrophages and neurons following SCI. 36
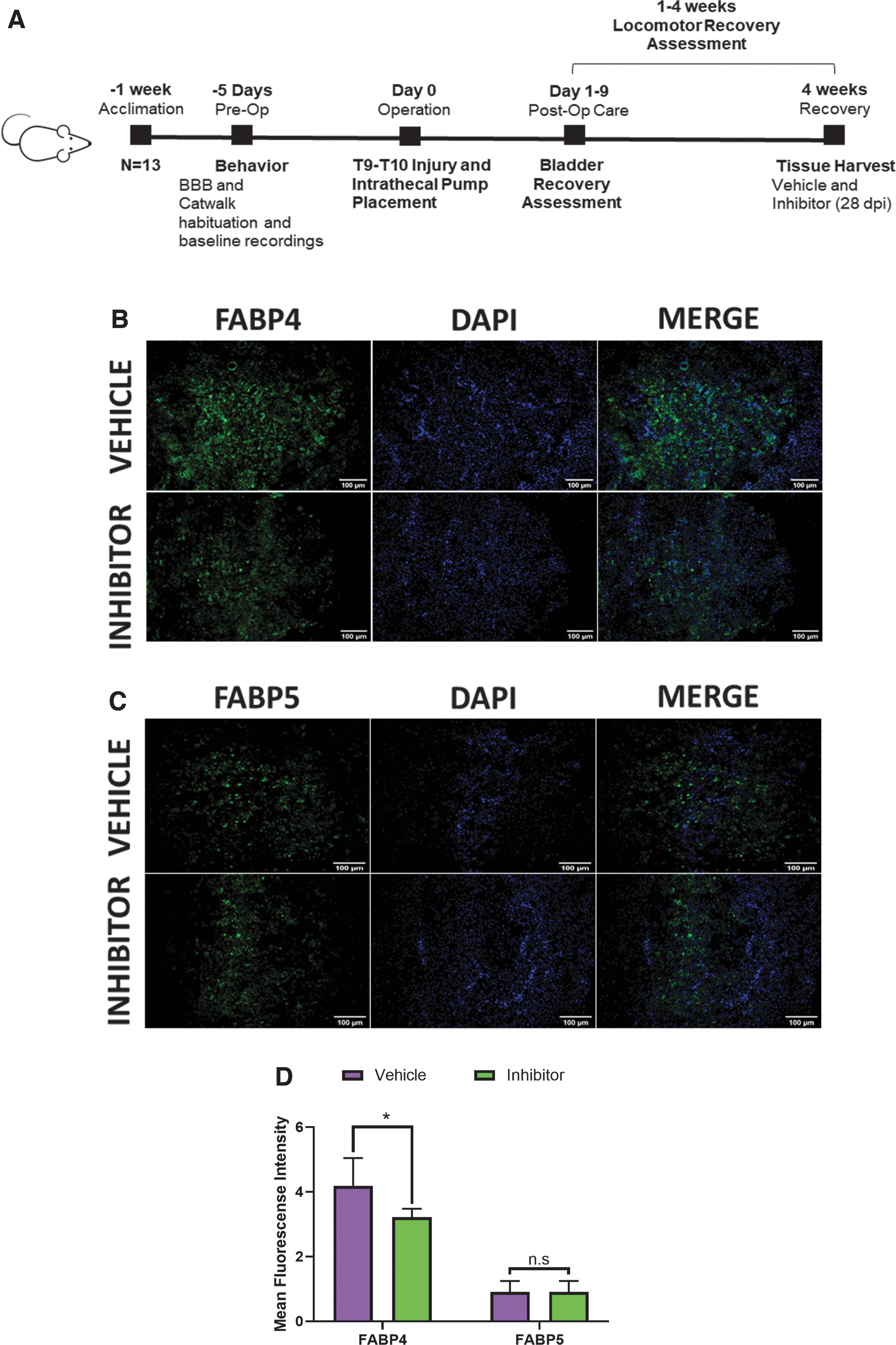
Treatment with BMS309403 small molecule inhibitor downregulates fatty acid-binding protein 4 (FABP4) expression in rats undergoing spinal cord injury.
Differences in spontaneous open-field locomotion of treated and control groups were determined using the BBB locomotor behavior assessment. The data show a significant enhancement in the recovery of SCI rats treated with BMS309403 compared with animals treated with the vehicle 7 dpi (*p = 0.0184; Fig. 5A). Because of the ordinal nature of the BBB locomotor scale, scores that are not commonly assigned can disproportionately increase variance. 42 To reduce potential variability introduced by the scale, results from the original 22-point scale were transformed. 42 The transformed data show an overall significant difference between the groups (*p = 0.0487) and confirmed the observed difference 7 dpi (*p = 0.0174; Fig. 5B).

Basso-Beattie-Bresnahan (BBB) locomotor behavior assessment shows administration of fatty acid-binding protein 4 (FABP4) inhibitor BMS309403 promotes locomotor recovery following spinal cord injury.
The Catwalk Gait Analysis Device (CatWalk) was used to determine changes in dynamic and static parameters of locomotion. The CatWalk delivered a comprehensive and reliable readout of recovery as it was specifically developed to provide automated parameters for rat locomotion and improve the evaluation and detection of interlimb coordination following SCI. 43,45,49 We found that parameters of hindlimb coordination such as regularity index (14 dpi, *p = 0.0465; Fig. 6A) and average speed (*p = 0.0195 overall and *p = 0.0257 at 28 dpi; Fig. 6B) were significantly higher in treated rats versus vehicle. Inter-paw coordination of the hindlimbs was also improved in treated rats as indicated by the cadence (Fig. 6D), a measure of the number of steps per second (overall *p = 0.0127; 28 dpi *p = 0.011). Strikingly, the data for maximum variation (Fig. 6C) indicates rats treated with the inhibitor displayed more consistent and uninterrupted movement of their hindlimbs when compared with vehicle both overall (****p < 0.001) and 28 dpi (*p = 0.0272). Finally, because of improved speed and reduced step variation, the run duration (Fig. 6E) for treated rats was lower than that of control (overall **p = 0.003; 28 dpi *p = 0.0185).

Catwalk analysis of locomotion in rats treated with fatty acid-binding protein 4 inhibitor show inhibition promotes significant locomotor recovery.
Autonomic bladder recovery is drastically accelerated in FABP4 inhibitor-treated rats
Residual urine volume in rat bladders was collected for 9 days after injury and the administration of BMS309403 or vehicle. When compared with controls, the amount of residual urine volume in inhibitor-treated rats was significantly decreased overall (*p = 0.022) and at 1 (*p = 0.037) and 3 dpi (*p = 0.01; Fig. 7A). Further, unpaired t-test analysis indicates inhibitor-treated rats also had a significantly faster rate of autonomic bladder recovery (4.75 days) compared with vehicle-treated rats (8 days; *p = 0.0282; Fig. 7B).

Fatty acid-binding protein 4 inhibition promotes autonomic bladder recovery.
Number of M2 CD163 and liver arginase–positive macrophages/microglia is significantly increased in the epicenter and penumbra of inhibitor-treated rats
We conducted immunofluorescence experiments using membrane marker CD163 and intracellular marker liver arginase—both of which have been well characterized as M2- associated proteins 50 -52 —to determine the effect of FABP4 inhibition on M2 MϕMG numbers. The data indicate a significantly increased number of M2 MϕMG (*p = 0.0196) in the epicenter of inhibitor-treated rats (Fig. 8A, 8C). Interestingly, more of these anti-inflammatory cells were also observed in the penumbra of inhibitor-treated spinal cord tissue (*p = 0.0166) compared with vehicle (Fig. 8B, 8D).

Fatty acid-binding protein 4 inhibition increases the number of CD163 and liver arginase double-positive cells in both the injured spinal cord epicenter and penumbra.
Discussion
The present study reports the robust upregulation of FABP4 mRNA and protein following spinal cord injury. This upregulation is observed in CD11b+ MϕMG but not in neurons or astrocytes. Further, increased FABP4 gene expression is correlated with downregulation of PPARγ, inactivation of Iκβα, and activation of NF-κB. Moreover, FABP4 inhibition increases the number of CD163+ liver arginase+ M2 macrophages/microglia in the injury epicenter and penumbra 28 dpi. Notably, the administration of BMS309403 stimulates locomotor recovery and reduces the severity of autonomic loss.
FABP4 promotes M1 macrophage/microglia differentiation
Following SCI, lipid accumulation in the macrophage cytoplasm impairs cholesterol efflux regulatory protein ATP-binding cassette A1 (ABCA1) and promotes M1 macrophage differentiation and the M2 to M1 macrophage switch. 53 Notably, FABP4 binding of pro-inflammatory lipids activates the IKK-NF-κB and JNK-AP-1 pathway and promotes pro-inflammatory MϕMG activity. 10,18,26,27,32,34,53 –57 Recent studies in ischemic stroke demonstrate the upregulation of FABP4 prompts M1- MϕMG differentiation resulting in neurological deficits. 27 Moreover, studies show FABP4 binding of specific unsaturated long-chain fatty acid ligands also activates its nuclear translocation signal and allows it to regulate PPARγ transcriptional activity and deter M2 differentiation. 58 Accumulating evidence demonstrates that FABP4, which at low levels supports PPARγ activity by shuttling ligands to the nucleus, can at high levels also prompt PPARγ ubiquitination, proteasomal degradation, and downregulation which ablate the receptor's anti-inflammatory properties. 59 Moreover, through its inhibition of the PPARγ-liver X receptor α (LXRα)-ATP-binding cassette A1 (ABCA1), FABP4 is also able to deter M2 macrophage differentiation. 34,55 Our study suggests a similar modulation of PPARγ by FABP4 as its upregulation during the acute phase of injury is subsequently downregulated at times when high levels of FABP4 are sustained. Interestingly, our study also shows PPARγ downregulation was accompanied by inactivation of Iκβα and activation of the NF-κB pathway, a process known to initiate pro-inflammatory MϕMG differentiation. 27,28,60 -62
FABP4 inhibition increases the number of M2 macrophages/microglia in the injured epicenter and penumbra
Infiltration of hematogenous macrophages supports axonal regeneration following peripheral nerve injury (PNI). 63 In this context, M1 macrophages active during Wallerian degeneration are replaced by pro-regenerative M2 macrophages. 63 Delays in the switch from M1 to M2 lead to impairments in functional recovery. 63 In contrast, the population of M2 MϕMG in the injured spinal cord is significantly reduced after 1 week and overtaken by an M1 pro-inflammatory phenotype directly associated with axonal dieback and retraction. 10,53 Additionally, M1 microglial activation during the acute injury phase induces neurotoxic A1 astrocyte differentiation. 64 The detrimental outcomes resulting from M1 MϕMG activity highlight the importance of identifying key factors that could promote their anti-inflammatory differentiation and allow for regeneration.
Several studies have shown that the inhibition of FABP4 polarizes MϕMG from the M1 to the M2 state and promotes recovery. 26,27,55,57 Remarkably, our data corroborate these findings and demonstrate that FABP4 inhibition increases the number of CD163+ and liver arginase+ M2 MϕMG in the injury epicenter and penumbra. Notably, the presence of M2 macrophages/microglia 28 dpi affirms that the inhibition of FABP4 is sufficient to modulate their differentiation and suggests its potential use as a novel therapeutic target to promote the M1 to M2 MϕMG switch and allow for regeneration following SCI.
FABP4 inhibition promotes functional outcomes following SCI
Blood-derived macrophages in PNI adopt the transcriptome of pro-regenerative endoneurial macrophages, whereas the transcriptome of macrophages in the injured spinal cord mostly resembles that of foam cells in atherosclerotic plaques. 65,66 The granulomatous infiltration of lipid-filled M1 macrophages from the subarachnoid space into the spinal cord induces peripheral tissue necrosis, promotes scar tissue formation and deters axonal regeneration. 16 Moreover, the persistent activity of M1 macrophages hinders inflammatory resolution and impairs regeneration at the epicenter while also promoting cytotoxicity in the penumbra and worsening clinical outcomes.
Importantly, our study demonstrates inhibition of FABP4 increases the number of M2 macrophages/microglia and creates a more permissive environment for recovery. In this pro-regenerative environment, we observed improvements in base of support, gait, and inter-paw coordination as well as decreased variation in movement. These changes are displayed by significant differences in regularity index, average speed, cadence, and the decrease in run duration and step variation. Additionally, we found FABP4 inhibition ameliorates autonomic bladder dysfunction during the acute phase of SCI by lessening urinary retention in the bladder and enhancing the rate of voiding reflex recovery.
Overall, our findings suggest that FABP4 is a promising and novel therapeutic target for promoting recovery after SCI through its modulation of macrophage and microglial activity.
Footnotes
Acknowledgments
Special thanks are extended to Dr. Susan Gardner, Dr. Kimberly Payne, Dr. Alfonso Duran, Dr. Carlos Casiano, Dr. William Langridge, Dr. Salvador Soriano, Dr. Michael Kirby, Dr. Melinda Soeung, and Ms. Lorena Salto for their invaluable help.
Funding Information
Research reported in this publication was supported by National Institutes of Health awards R25GM060507 and P20MD006988.
Author Disclosure Statement
No competing financial interests exist.