
Abstract
Therapies are limited for pediatric traumatic brain injury (TBI), especially for the very young who can experience long-term consequences to learning, memory, and social behavior. Animal models of pediatric TBI have yielded mechanistic insights, but demonstration of clinically relevant long-term behavioral and/or cognitive deficits has been challenging. We characterized short- and long-term outcomes in a controlled cortical impact (CCI) model of pediatric TBI using a panel of tests between 2 weeks and ∼4 months after injury. Male rats with CCI at postnatal Day (PND) 10 were compared with three control groups: Naïve, Anesthesia, and Craniotomy. Motor testing (PND 25–33), novel object recognition (NOR; PND 40-50), and multiple tasks in water maze (WM; PND 65-100) were followed by social interaction tests (PND 120-140). Anesthesia rats performed the same as Naïve rats in all tasks. TBI rats, when compared with Naïve controls, had functional impairments across most tests studied. The most sensitive cognitive processes affected by TBI included those that required fast one-trial learning (NOR, WM), flexibility of acquired memory traces (reversals in WM), response strategies (WM), or recognition memory in the setting of reciprocal social interactions. Both TBI and Craniotomy groups demonstrated increased rates of decision making across several WM tasks, suggesting disinhibition of motor responses. When the TBI group was compared with the Craniotomy group, however, deficits were detected in a limited number of outcomes. The latter included learning speed (WM), cognitive flexibility (WM), and social recognition memory. Notably, effects of craniotomy, when compared with Naïve controls, spanned across multiple tasks, and in some tasks, could reach the effect sizes observed in TBI. These results highlight the importance of appropriate control groups in pediatric CCI models. In addition, the study demonstrates the high sensitivity of comprehensive cognitive testing to detect long-term effects of early-age craniotomy and TBI and provides a template for future testing of experimental therapies.
Introduction
Traumatic brain injury (TBI) is the leading cause of long-term disability in children and young adults in the United States, making it a significant public health problem. 1 Following moderate-to-severe TBI, children can have functional deficits that persist for years and even into adulthood. 1,2 These deficits can alter developmental trajectories in the maturing brain, leading to psychosocial, cognitive, and behavioral abnormalities. Animal models of pediatric TBI have yielded mechanistic and therapeutic insights into developmental brain injury, but many have been limited in their ability to measure functional deficits with translational relevance to those seen clinically. 3,4 In addition, some studies have evaluated only acute deficits early after injury, without identifying longer term or sustained behavioral deficits. This could be particularly important after developmental brain injury, as some deficits may only be detected at long-term times into adulthood. 5
Another reason for these limitations may be that some pre-clinical TBI studies have only measured outcomes in a single domain, such as motor testing using rotarod or beam walking, or spatial learning with Morris water maze testing. Detection of sustained deficits following developmental brain injury could be improved by utilizing a comprehensive battery of functional testing that measures outcomes in many domains and includes different types of tests for each domain. 6 The importance of evaluating a broad panel of outcome tests in pediatric pre-clinical models of TBI is highlighted in a recent review by Semple and colleagues. 7
The choice of control animals in developmental TBI models is another factor that may lead to variability of results and affect sensitivity of a study design. For example, some studies use animals with sham craniotomy as a main control group to delineate the effects of TBI. 7 It is possible that either the craniotomy or anesthetic exposure could influence behavioral outcomes as the developing brain may be especially sensitive to such insults. 8,9 If such effects are present, the window of opportunity for detection of the effects of TBI might be diminished. Further, if Naïve rodents are used as the only control group, the effects observed after brain injury are likely to include a combination of effects from other factors, with the craniotomy being one of the major ones. One could argue that the sensitivity of these different insults to ameliorating effects of therapeutics might be different. Such questions, however, cannot be addressed in studies with a limited number of control groups. Implementation of studies with multiple control groups, in addition to a larger number of animals and longer time needed to complete the experiments, is critically dependent on whether the protocols are available that would be sensitive enough to demonstrate long-term functional outcomes of not only TBI, but also craniotomy and/or anesthesia.
In the current study, we compared a controlled cortical impact model (CCI) of pediatric TBI using three control groups (Naïve, Anesthesia only, and Craniotomy only). We aimed to test a panel of tasks to characterize multiple behavioral and cognitive domains relevant to the impairment seen in pediatric TBI. We measured functional outcomes in the motor domain (gait analysis, foot fault), learning and different types of memory (novel object recognition, multiple protocols for reference and episodic-like memory in water maze), and social interactions (three chamber test, social recognition).
Methods
Study and group design
The study was approved by the Animal Care and Use Committee of the Johns Hopkins University School of Medicine (Baltimore, MD) and in compliance with the National Institutes of Health (NIH) guidelines. Surgeries were conducted at the postnatal Days 10-11 (PND) in Sprague Dawley rats. Only male pups were used in this study. Rats were randomly assigned to four different treatment groups: Naïve controls (n = 14), Anesthesia only group (n = 12), Craniotomy only group (n = 17), and TBI (n = 17). The study design is shown in Figure 1. After weaning from dams, rats were housed in standard rat cages and kept in groups of three per cage for up to 2 months of age, and, later, in groups of two per cage. After the end of behavioral testing, rats were anesthetized, sacrificed and their brains harvested. Detailed methods and procedures can be found in the Supplementary Methods.

Timeline of experimental procedures. Traumatic brain injury (TBI) was performed on the left hemisphere (see details in the “Methods” section) at post-natal Day 10 (PND 10). To assess for possible TBI induced motor deficits, Foot Fault and Gait Analysis tests were performed at PND 25-26 and PND 27-P30, respectively. Novel object recognition was conducted at PND 40-50 to test recognition memory. Then, a number of water maze protocols were employed to test for spatial reference memory as well as episodic-like and working memory at PND 65-100. Finally, possible alterations in social interactions were tested at PND 120-140 using social recognition and social motivation tasks. Color image is available online.
Controlled cortical impact (CCI) injury
TBI was performed using a controlled cortical impact (CCI) device, as previously described. 10 -12 Injury was produced using a 3-mm metal impactor tip with a depth of penetration of 1.5 mm, at velocity of 5.5 ± 0.3 m/sec, and a duration of deformation of 50 msec. At the completion of surgery, isoflurane was discontinued. When the rats emerged from anesthesia, they were returned to their cages with their littermates and mother.
The rats assigned to the Craniotomy group underwent identical surgeries, with the exclusion of the CCI. The rats assigned to the Anesthesia group received isoflurane for the same length of time as the TBI and Craniotomy groups (30 min) without any surgical interventions. Representative brain images from each of the groups are presented in Supplementary Figure S1 demonstrating the extent of brain damage in the TBI group and preserved brain tissue in the Craniotomy group, as expected from our previous studies. 10 -12
Behavioral testing
Behavioral testing started when rats reached 25 days of age in the order described below (Fig. 1). The battery of tasks included tests to access multiple domains affected in children with TBI: locomotor function and coordination, different types of learning and memory, and social interactions (social recognition and social motivation). ANY-maze software (Stoelting Co, Wood Dale, IL) was used to record data in all tasks unless noted otherwise.
Foot fault test
The test was conducted at PND 25-26, 2 weeks after surgery. Animals were placed on an elevated, horizontal metal grid with a mirror underneath for video recording of the paw placement. The number of foot slips for each limb over 5 min was analyzed during video playback.
Gait analysis
A runway test was conducted at PND 27-33 using a 1 m long enclosed glass runway using Gait Scan software (Clever Sys, Reston, VA). The following parameters were analyzed: foot pressure, foot angles, run speed and distance, step base, step length, and whole-body length.
Novel object recognition (NOR)
NOR test was conducted at PND 40-50. Rats were habituated to the box (60 × 60 cm) twice a day 10 min each for 2 consecutive days. In Session 1, two similar objects from LEGO® were placed in the box and rats were allowed to investigate the objects for 5 min. On the next day, session 2 was conducted when one of the objects was replaced with a new one, and rats were allowed to investigate the objects for 5 min.
Morris water maze
Morris water maze tasks were performed at PND 65-100 as described before 13 with modifications (see the Supplementary Methods for detailed descriptions of the protocols).
Platform and straight swim pre-training
Rats were first pre-trained in a straight swim task to familiarize them with procedural aspects of testing in the water maze, to decrease effects of stress from swimming and to reveal possible changes in motor abilities that could interfere with cognitive testing. One rat from the Anesthesia group was excluded due to slow swimming related to a recent leg injury. The physical health of this rat was normal for all preceding tasks; therefore, it was not excluded from the analyses of those tasks. No treatment-related differences were observed in this task (data not shown).
Classic Morris water maze (MWM) protocol
Learning in this task results in formation of long-lasting spatial reference memory. 13,14 The duration of the task was 2 days. For each trial, the rat was placed in a 200-cm pool in a randomly predetermined quadrant other than the quadrant containing the platform. Inter-trial intervals were approximately 20 min. Training trials were 60 sec long and probe trials were 35-45 sec long. At the end of the probe trials, the collapsed platform was returned to its raised position to maintain the same response-reinforcement contingency as in the platform trials. 15,16 The probe trials conducted at the beginning and the end of a daily session were used to assess memory following short (30 min) and long delays (∼24 h).
Repeated reversals in water maze (MWM-Reversals)
In contrast to the reference memory task (MWM), the reversal task challenged episodic-like memory as rats need to keep changing their memory representation for the environment by discriminating the information by “what,” “where,” and “when” categories. 14,17,18 This task was performed similarly to the MWM except the platform location was changed daily for three consecutive days.
Radial Water Maze (RWM)
After completion of the MWM-Reversals task, a radial maze enclosure was placed into the same pool and the rats were required to find the hidden platform in one of the eight arms of the Radial Water Maze. Similar to the repeated reversals, the position of the platform was changed daily. For 4 days, each rat performed six 90 sec-long trials started in a randomly predetermined arm.
Radial Water Maze with cued and hidden platform
After the RWM task, rats were trained for 2 additional days in a more challenging protocol. In addition to trials with a hidden platform (as in the previous stage of RWM), trials in which the position of platform was prompted by a highly visible cue were introduced. Positions of the cued platform were changed between each trial. Eight daily trials switched between two types of trials: from a trial with the cued platform to a trial with the hidden platform. The position of a hidden platform was changed daily.
Social interaction testing
Social recognition
At PND 120-130, social recognition memory was tested based on the habituation-dishabituation protocol. 19,20 In an orientation trial, rats were placed in an open field (40 × 60 cm) and allowed to explore the new environment for 5 min. Then, three habituation trials were conducted in which the test rat was introduced to the same young male pup (PND 25) and allowed to interact for 2 min per trial. In the fourth dishabituation trial, the test rat was introduced to a novel PND 25 male rat pup with an expectation of activation in social interactions.
Three-chamber social motivation test
Social motivation 20,21 was tested at PND 130-140. In an orientation trial, a test rat was placed in the middle part of a social motivation chamber (50 × 80 cm) and allowed to explore the new environment for 10 min. Trial 1 and 2 represented the stage of habituation when a new rat pup (PND 25) was placed in one of the containers repeatedly, and the test rat was allowed to investigate for 5 min per trial. Trial 3 represented the stage of discrimination when yet another pup was placed in the second container, and the test rat could explore both pups simultaneously for another 5 min.
Statistical analysis
The data were analyzed using the statistical package STATISTICA 13 (TIBCO Software Inc., Palo Alto, CA) and a minimal level of significance p < 0.05. The data were analyzed using mixed design analysis of variance (ANOVA) with Treatment (Naïve, Anesthesia, Craniotomy, TBI) as an independent factor and repeated measure(s) (RM; trials, blocks, arms, etc.). The least significant difference post hoc test was applied to significant Treatment or Treatment × RM interactions to evaluate differences between four sets of means: Naïve versus Anesthesia; Naïve versus Craniotomy; Naïve versus TBI; and Craniotomy versus TBI. Two-tailed t-test was used to analyze differences from chance levels. The details of statistical results including F, df, and post hoc tests are presented in Supplementary Tables S1-S6. Data in figures represent means ± standard error of the means unless otherwise noted.
Results
Locomotor function in foot fault test and gait analysis
The assessment of locomotor function started in a foot fault test conducted 2 weeks after TBI when rats reached the age of 25-26 days (PND 25-26; Fig. 1). Rats from the Anesthesia group were not significantly different from Naïve controls in any of the measures. The Craniotomy group demonstrated more errors than Naïve controls for the rear paw on the side contralateral to craniotomy (right rear paw, p < 0.0001). TBI rats had more misplacements than Naïve rats for both rear paws (Fig. 2A; Supplementary Table S1). For the front paws, both Craniotomy and TBI groups showed a notable tendency making more missteps as compared with Naïve rats; however, these differences did not reach significance.
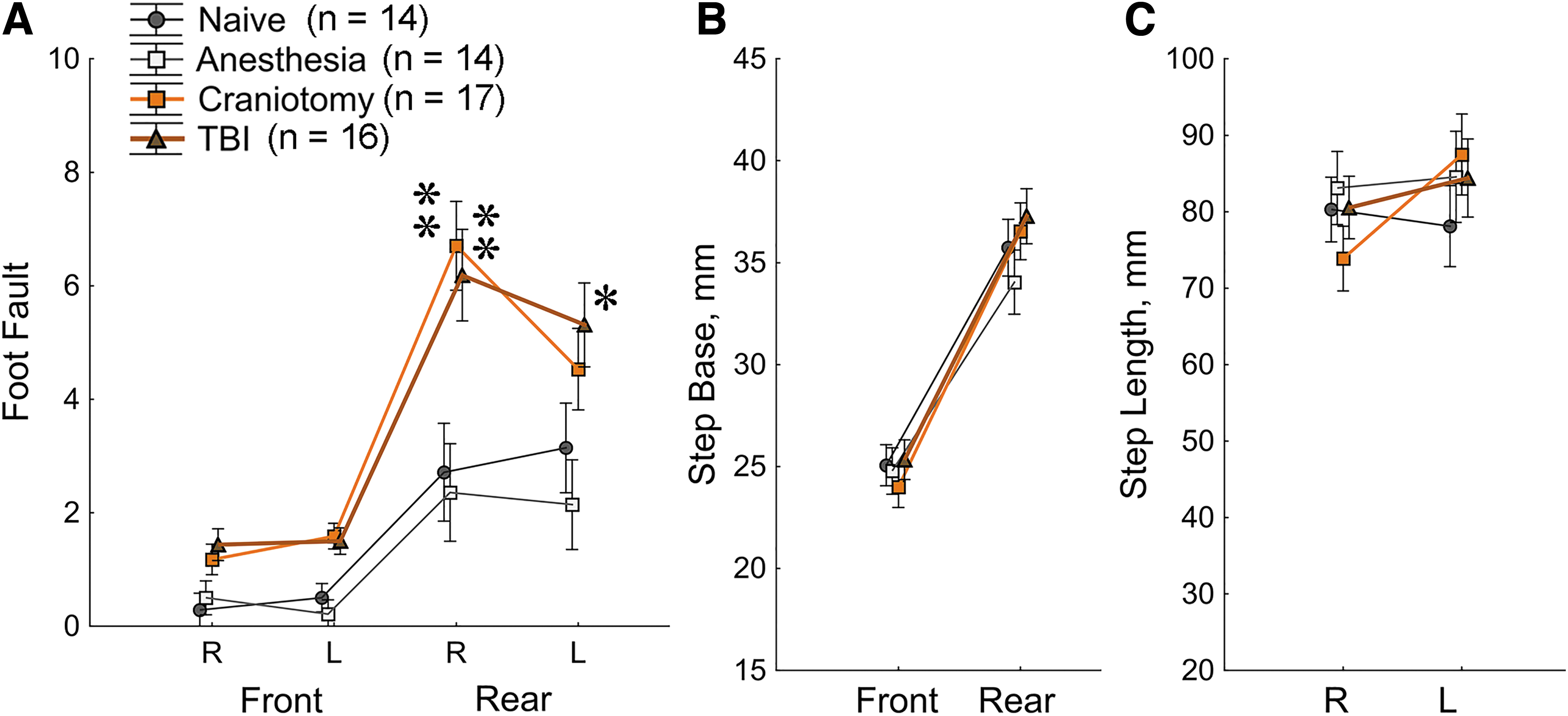
Traumatic brain injury (TBI) and Craniotomy rats show deficits in the Foot Fault test
The gait analyses were conducted in a runway test in the 3rd week after TBI when rats reached age of PND 27-33. Different variables including foot pressure, foot angles, run speed and distance, foot spacing, and whole-body length where analyzed, and no significant treatment-related changes were found. Results for the width of step base and step length are shown in Fig. 2B, 2C. As expected, a significant effect of front versus rear paws was observed for the variable of step base (Fig. 2B) with the rear paws having wider step than the front paws (Supplementary Table S1).
Object recognition memory
The object recognition memory was tested in PND 40-50 rats at Week 5 after surgeries. In Session 1, no significant differences in preferences investigating two identical objects were found in any of the treatment groups (Fig. 3B; Supplementary Table S2). After a 24-h delay, Session 2 was conducted when one of the objects was exchanged for a novel one (Fig. 3A, 3B). As expected, rats from the Naïve control group investigated the new object significantly longer than the previously presented object (Fig. 3B). Rats from the Anesthesia group demonstrated significant preferences to the new object, similar to the naïve control group. However, Craniotomy and TBI groups showed no preferences between the new and old objects (Fig. 3A, 3B; Supplementary Table S2), implicating deficits in the object recognition memory.

Traumatic brain injury (TBI) and Craniotomy rats show deficits in the Novel Object Recognition (NOR) test.
Analysis of distances travelled during object recognition testing showed that Craniotomy and TBI groups travelled longer distances in Session 2 as compared with Session 1 (Fig. 3C). Thus, in addition to significant deficits in object recognition memory, Craniotomy and TBI groups demonstrated behavioral activation during presentation of the novel object.
Reference memory in a classic Morris water maze
Cognitive testing in a classic version of the Morris water maze started at Week 10 after surgeries when animals reached approximately 11 weeks of age (Fig. 4A-C). This task assays spatial reference memory for a hidden platform that stays in the same position across trials and days. 14,16 As training progressed, latency to find the platform decreased in all treatment groups (ANOVA, effect of trial p < 1.0E-6; Supplementary Table S3). There were no significant between-group differences in swim speed (data not shown). Despite the training-related improvements in the escape latencies, rats from the TBI group required significantly longer time than Naïve controls to find the platform (ANOVA, effect of Treatment, p < 0.012; Fig. 4A). Craniotomy group was not different from Naïve controls in this measure if averaged across the training (Fig. 4A insert); however, in contrast to Naïve and Anesthesia groups, an overnight delay significantly increased latency to find the platform in Craniotomy rats (Fig. 4A). This data indicated that spatial learning abilities in Craniotomy group were relatively preserved but deficits occurred in long-term retention of platform location.

Traumatic brain injury (TBI) and Craniotomy rats demonstrated deficits in the Reference
To assay the development of spatial preferences, the probe trials were conducted, in which the platform was initially lowered and then made available after a variable interval (40-45 sec). The good performance in the probe trials required not only spatial memory for the location of the platform (as in the training trials), but also persistence in looking for the platform in the appropriate location. This complexity led to longer dynamics of learning in the probe trials. Specifically, in Naïve controls, learning the spatial location of the platform (as judged by the escape latency in the training trials) reached the asymptotic performance after the first 4-5 trials, whereas the appropriate spatial preference was observed only at the end of the 2nd day of testing (Probe 3; Fig. 4B). All treatment groups reached spatial preferences above chance level (Probe 3; Fig. 4B, 4C). Nevertheless, TBI and Craniotomy groups showed less efficient performance than Naïve controls (Fig. 4C; p < 0.005). In addition, testing of long-term reference memory after a 24-h delay demonstrated the loss of spatial preferences in the TBI and Craniotomy groups (Fig. 4E, Previous Platform for Reversal 1).
Episodic-like memory in Morris water maze
During the next 3-day long training, the platform position was changed daily, while the trial schedule as well as spatial cues around the water maze were kept the same. TBI rats showed longer latencies finding the platform as compared with Naïve controls (ANOVA, effect of treatment p < 0.013; Fig. 4D). Probe trials conducted at the end of each reversal indicated that all treatment groups were able to show spatial preferences to the new position of platform (Fig. 4F). These data indicated that deficits in spatial reversal learning observed during training trials in TBI rats did not prevent forming new spatial memories. This was documented in the probe trials with a short 30-min-long delay (Fig. 4F). However, these memories were not retained, as TBI rats demonstrated no spatial preferences when tested in the probe trials after a long over-night delay (Fig. 4E). Importantly, in contrast to TBI, Naïve controls showed significant preferences to the platform location of the previous day of testing (Fig. 4E). This data implicated that performance of Naïve controls in this task required not only learning the new platform but inhibiting visits to the old platform location as well. In the course of repeated reversals, the preferences of Naïve controls to the previous platform decreased to the chance level (Fig. 4E). In the last reversal, when learning the new platform position in Naïve controls was not complicated by interferences with memory of the previous platform location, their performances in the final probe trial were highly efficient (Fig. 4F) and similar to those in the reference memory task (Fig. 4B, 4C). This stage of repeated reversals revealed significant deficits in TBI and Craniotomy groups, as both groups demonstrated less accurate spatial preferences than in Naïve controls (Fig. 4F). Anesthesia group showed performances statistically indistinguishable from Naïve controls.
Episodic-like memory tested in the Radial Water Maze
Similar to reversal training in MWM, episodic-like memory in the Radial Water Maze (RWM) was tested by changing the location of the platform daily. During the 1st day of training (Fig. 5A), Naïve and Anesthesia groups demonstrated a one-trial learning; namely, the level of their performances in the latency and number of errors reached asymptotic levels already at the 2nd training trial (Fig. 5B, 5E; Supplementary Table S4). Craniotomy group showed a mildly delayed learning as they reached their asymptotic performance in the latency to find the platform at the 4th trial. TBI rats were not able to demonstrate the one-trial learning and improved incrementally up to the 4th trial.

The Radial Water Maze task revealed deficits in spatial learning and memory in traumatic brain injury (TBI) rats.
Starting from the 2nd day of testing, rats were required to learn a new position of the platform. In the orientation trial (Trial 1), rats from all groups visited the previous platform location demonstrating normal long-term spatial memory (Fig. 5G, 5H, Platform 1). Independent of the treatment condition, about 30% of errors in the orientation trial were made due to repeated visits to the previous platform location (Fig. 5H, Trial 1). Interestingly, the orientation trial in TBI rats was significantly shorter than in Naïve controls (Fig. 5C, Trial 1) while the number of errors was not different (Fig. 5F, Trial 1). Indeed, the rate of arm choices (the number of arm choices per sec; Fig. 5D) was significantly increased in TBI rats as compared with control. Craniotomy rats showed significantly higher rates of arm choices in the orientation trial as well (Fig. 5D, Trial 1).
After the orientation trial, TBI rats showed significantly longer latency and more errors to find the platform than the Naïve controls. Anesthesia and Craniotomy groups demonstrated performance comparable to controls. When memory for the platform was assessed after a 24 h-long delay (in Trial 1 of the next testing day), TBI rats made more errors before reaching the previous day's platform location than controls (Fig. 5G, Platforms 2-4). Despite this impairment, TBI rats visited the previous platform location more often than expected from a chance level across all 6 trials of testing (Fig. 5H). These data indicate that the deficit seen in TBI rats after the 24-h delay is not due to an impairment in long-term memory, but likely due to a deficit in its immediate recall during Trial 1. When given more time/trials, TBI rats demonstrated their spatial preferences to the previous platform. In fact, the inability of TBI rats to inhibit such preferences after the orientation trial might be a reason for their less efficient performance than in control groups.
Spatial and goal-directed strategies in the Radial Water Maze
All groups have learned the RWM task creating conditions for a more challenging test. At the next step of testing, a new type of trials was introduced in which the position of platform was prompted by a highly visible cue at the end of the arm (Fig. 6). Positions of the cued platform were changed between trials. Eight daily trials switched between two types of trials: from a trial with the cued platform to a trial with the hidden platforms. Thus, this stage of testing required acquisition of spatial memory for a new hidden platform while, if the cue is present, responding by a goal-directed behavior toward the cued platform.
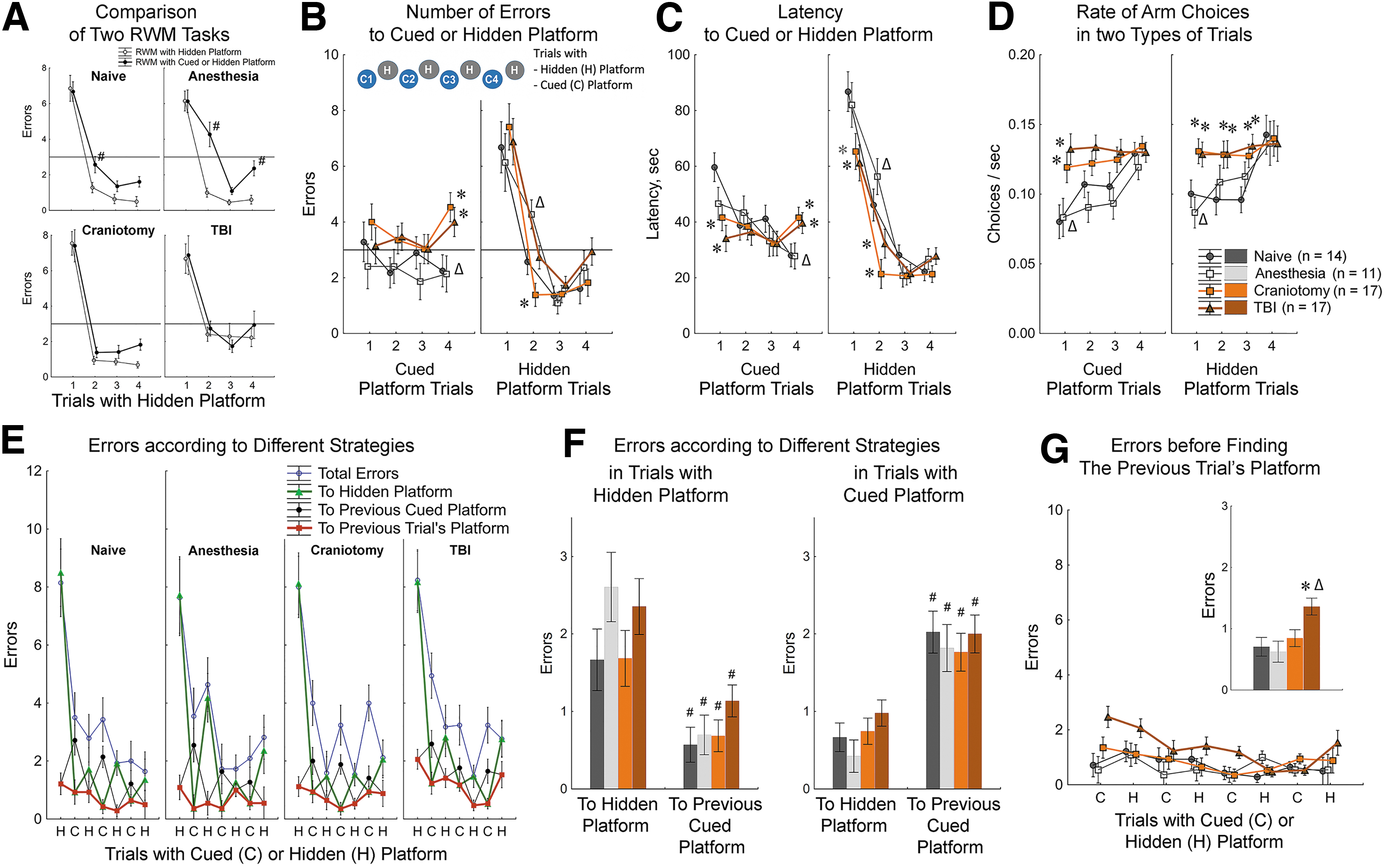
Strategy competition in the Radial Water Maze (RWM) revealed cognitive impairments in traumatic brain injury (TBI) and Craniotomy rats.
The new challenging protocol showed that introducing the cued trials impaired learning the hidden platform. In particular, performances of Naïve and Anesthesia groups in the spatial trials were significantly worse when compared with their performances in the previous standard protocol (Fig. 6A). These impairments were most obvious in the 2nd trial (Fig. 6A). The standard Radial Water Maze protocol demonstrated that the 2nd trial was critical to dissociate differences in the speed of learning between treatment groups. With the new protocol, the between-group differences in the second trial were readily apparent as well. However, in this more challenged condition, the differences between treatment groups were represented by a dissociation between best performances of Craniotomy group and the worst performances of the Anesthesia group (Fig. 6B, 6C, Hidden Trial 2). Interestingly, Craniotomy group had dramatically fewer number of errors in spatial trials, but significantly more errors in the cued trials than Anesthesia group (Fig. 6B, Cued Trial). These data implied that better performance of Craniotomy rats in spatial trials came at the expense of performance in the cued trials. In the Anesthesia group, the cued trial performance was similar to the Naïve controls, however their attempts to adequately respond to cued platform might have deteriorated their ability to learn the location of new hidden platform.
Analysis of decision rates (number of arm choices per second) yielded the clear dissociation between high rates in TBI and Craniotomy groups and low rates in Naïve and Anesthesia controls (Fig. 6D). This dissociation was reminiscent of that observed in the orientation trials in the standard Radial Water Maze (Fig. 5D, Trial 1). In the more complex protocol, the between-group differences were observed in the orientation trials for both cued and hidden platforms. In addition, Naïve controls continued to show low decision rate in most of the trials with hidden platform. It is noteworthy that the dissociation of Naïve and Anesthesia groups from Craniotomy and TBI groups in the decision rate was independent of the actual success of those decisions. For example, the low rate of decision in Anesthesia rats was observed despite of their problematic performance in the spatial trials. Craniotomy rats demonstrated the high rate of arm choices in the trials with successful decisions (the spatial trials with hidden platform) and in the trials with less successful choices (the cued trials). These data indicated that the high decision rates observed in Craniotomy and TBI groups might be reflective of their inability inhibiting the action before making a decision.
The analysis of the order of choices within each trial revealed a highly organized behavior (Fig. 6E). Namely, during the cued trials all rats visited the hidden platform before going to the cued platform (Fig. 6F). During the trials with hidden platform, all rats visited the arm with the previous cued platform before going to the correct arm (Fig. 6F). Entering the correct arm and contacting a platform did not necessarily result in climbing on the platform, as animals continued to engage in orientation behavior and visited other arms before their final choice. These observations can be reconciled if in the beginning of every trial the choice was directed to the location of the platform in the previous trial. Indeed, the analyses of such choices revealed that visiting the previous trial's platform location was the 1st or 2nd choice for most of the animals (Fig. 6E, 6G). TBI group had followed this strategy as well, however, made significantly more errors finding the previous platform location than Naïve controls (Fig. 6G). This deficit in following spatial strategy coincided in TBI rats with the impairment in learning goal-directed behavior as they demonstrated long latencies and substantial number of errors in the final cued trials (Fig. 6B, 6C).
Social interactions
Social recognition task (Fig. 7A-D)
All treatment groups demonstrated robust habituation to the new cage environment, and no differences were observed in novelty-induced exploratory activity (Fig. 7B; Supplementary Table S6). Then, social recognition memory was tested using a habituation-dishabituation paradigm. Analyses of the latency to initiate the social contact revealed that TBI rats took significantly longer for their first social approach as compared with Naïve and Anesthesia groups (Fig. 7C). In accordance with this observation, analyses of duration of social investigation in the first 30 sec of each of the tests showed that TBI rats spent less time interacting with novel rats. This was true for both the 1st trial (when the first novel rat was presented) and the 4th trial (when the second novel rat was presented; Fig. 7D). In terms of social recognition at the dishabituation phase, TBI rats were the only treatment group that failed to significantly increase social engagement when the novel rat was presented (Fig. 7D). These observations were supported by significant effects of Treatment, Trial and Treatment × Trial interaction (ANOVA; Supplementary Table S6). In addition, analyses of total time of social investigation in this task revealed that Craniotomy group accumulated longer duration of social interactions than Anesthesia and TBI group. The size-effect of this effect was small (Fig. 7D).

Deficits in social interactions were revealed in traumatic brain injury (TBI) rats in reciprocal social recognition
Three-chamber social motivation task
In this task, no treatment-related differences were detected (Fig. 7E-G). During the 1st two trials, rats from all treatment groups showed significant preferences investigating the enclosure with social object (a young rat) as compared with a non-animated enclosure. During the 3rd trial, when a novel rat was present in addition to the familiar one, all groups showed clear preferences investigating the novel rat (Fig. 7F, 7G).
Data integration from different tasks
Multiple tasks are necessary to systematically characterize the effects of brain injury on behavioral and cognitive domains. However, as outcomes from different tasks are represented on different scales, such studies bring an additional complexity in reconciling the multitude of behavioral measures and delineation of relative sensitivity of different tasks/measures. To address this issue, we calculated effects sizes for a set of variables drawn from all tasks that yielded significant main effect of group (ANOVA; Supplementary Tables S2-S6). The effect sizes were calculated using Hedges q statistic as described (Fig. 8). 22 Comparisons between Naïve and TBI groups yielded variables with medium to large effect sizes in all tasks (Fig. 8A, 8C). Maximum effect sizes were observed for the time spent around the platform during the last reversal in the Morris water maze (q = 1.50) and for the number of errors in the Radial Water Maze (q = 1.57). The same two variables were among the most sensitive to differentiate Naïve versus Craniotomy (q = 1.16) and Craniotomy versus TBI (q = 1.54) groups (Fig. 8C). In addition, dissociations of Naïve versus Craniotomy groups were particularly strong for the variables of foot faults (q = 1.17), motor activation in Novel Object Recognition task (q = 1.20), and errors to visible platform in the Radial Water Maze (q = 1.07).
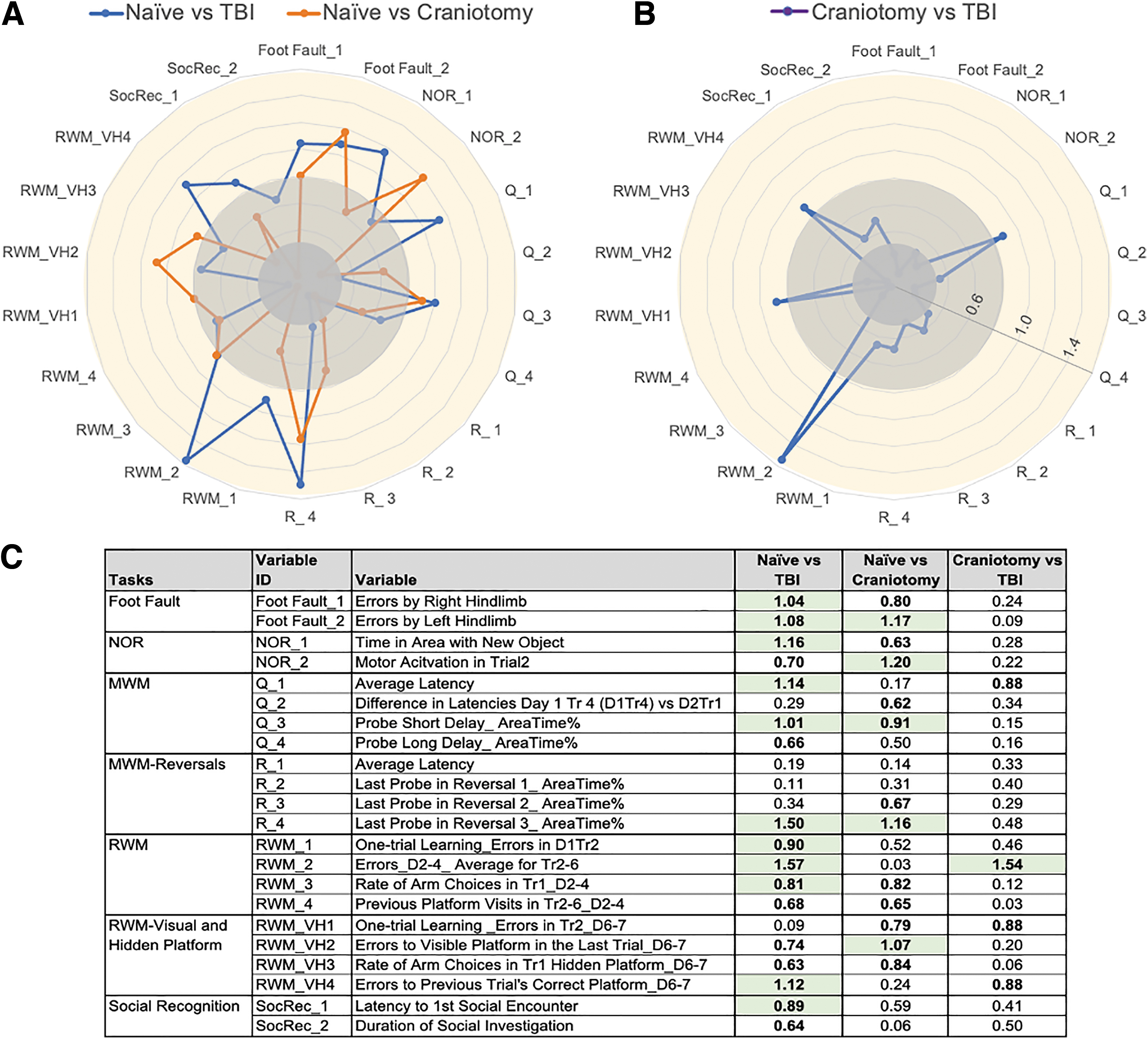
Effect sizes of traumatic brain injury (TBI) depend on the choice of control group (Naive or Craniotomy).
When controlling the false discovery rate (FDR = 0.05), we noticed that Naïve versus TBI comparisons, which satisfied the Benjamini-Hochberg p value, had effect sizes q > 0.80 (Fig. 8A, 8C). For Naïve versus Craniotomy and Craniotomy versus TBI comparisons, variables that satisfied the FDR had bigger effect sizes, q > 0.90 and q > 1.50, respectively (Fig. 8C).
Discussion
In this study, we conducted a detailed systematic evaluation of cognitive sequelae of pediatric TBI in concert with assessment of deficits in sensorimotor coordination and social domains. Several control groups were included to differentiate possible effects of pediatric trauma involving damage to neural tissue from the effects of craniotomy and anesthesia. A structured set of multiple cognitive tasks with increasing levels of complexity allowed for robust interpretations of the most vulnerable cognitive domains (Fig. 8; Supplementary Table S7).
Foot fault test but not gait analysis is sensitive to the effects of TBI and Craniotomy
TBI was modeled using one of the most widespread protocols, controlled cortical impact (CCI) with the impact directed at the sensorimotor cortex, so it was important to assess the extent of functional motor deficits. Analyses of locomotor functions at the 3rd week after trauma revealed significant deficits in both Craniotomy and TBI groups (Fig. 2; Fig. 8C). The impairments were easily detectable in the foot fault test but not in gait analyses. In contrast to the latter, the foot fault test requires integration of visual and proprioceptive information to construct prediction and then timely execute appropriate foot placement. Although we did not analyze the extent of functional or histological tissue damage in this study, the behavioral data suggest that the requirement of complex sensorimotor integration might make the foot fault test more sensitive to deteriorating effects of TBI and craniotomy than sensorimotor reflexes supporting normal gait. In all rats, the number of missteps were higher for hind paws than for front paws, implicating that correct prediction of placement is more difficult for hind paws. Not surprisingly, the effects of TBI and craniotomy were most significant for the number of hind paws missteps. It is important to note that despite significant differences observed in the Foot Fault test, the lack of gross motor abnormalities in gait analysis, motor activity and swim speed indicated that the deficits observed in cognitive tasks are not likely to be explained by sensorimotor impairments.
The presence of foot fault deficits without gait alterations is consistent with the limited data in developmental TBI models. To our knowledge, only one other study employed foot fault testing in a pediatric TBI model. Ajao and colleagues showed increased foot faults at post-injury Days 7 and 60 after CCI in PND 17 rats. 23 The pediatric TBI studies using gait analysis have been performed primarily in piglets, demonstrating sustained deficits out to 12 weeks after CCI in 4-week-old piglets, 24,25 and correlating with injury severity. 26 One group evaluated a panel of sensorimotor tests for one month after CCI in PND 17 rats, and concluded that gait analysis was not suitable for measuring long-term recovery following CCI in immature rats, due to lack of motivation in performing the task during development, and interference in the signal by male rat testes after puberty. 6 This study also raised concerns with the use of grid walking for foot fault analysis, due to the lack of scalability with growth of the animals.
NOR task is highly sensitive to the effects of TBI and Craniotomy
Testing of object recognition memory revealed that TBI and craniotomy resulted in significant deficits observed as a lack of exploratory preferences for a novel object (Fig. 3; Fig. 8C). The impairments in novel object recognition coincided with significant motor activation in these groups, implying that detection of novelty was preserved; however, discrimination of the source of novelty was impaired. The object recognition task is based on a natural motivation of rodents to explore novelty and has been shown to require normal functioning of the perirhinal cortex and hippocampus. 27,28 Significant deficits in the TBI group indicate that an early-life damage of the sensorimotor cortex may result in long-term secondary changes in the integrity of other cortical areas such as perirhinal cortex and hippocampus. These effects are consistent with a secondary (delayed) cell loss in this model, particularly in the hippocampus. 29 The presence of object recognition deficits in the Craniotomy group suggests that deteriorating functional outcomes can be observed even if the injury is limited to dura matter.
Similar deficits in novel object recognition have been demonstrated by our group and others, using the CCI model in immature rats injured at PND 10-21. 11,30,31 Importantly, post-injury impairments in object recognition were prevented by several multi-mechanistic, neuroprotective treatments, including erythropoietin, 32 aceytyl-L-carnitine, 31 progesterone, 33 docosahexaenoic acid, 30 MK-801, 34 and the 20-HETE inhibitor, HET0016. 11 In addition, a decrease in novel object recognition in pediatric TBI models appears to relate to injury severity in closed head injury models, 35 with repeat TBI events leading to worsening memory impairments. 36
Multi-stage Morris water maze is sensitive to the effects of TBI and Craniotomy
To further analyze TBI-related cognitive deficits, we employed a battery of tasks based on a motivation to escape from water. These tasks were based on similar procedural approaches (such as a hidden escape platform, similar inter-trial intervals, different start location for every trial, etc.) facilitating between-task comparisons. In contrast to the novel object recognition task, in the water maze tasks information about objects (spatial cues and escape platform[s]) had to be integrated into a spatial map of the environment with critical roles of hippocampus-entorhinal cortex and hippocampus—prefrontal cortex networks. 37 -39 Importantly, the task complexity increased as training progressed starting from testing spatial reference memory to testing features of episodic-like memory that require use of “where” and “when” categories in an integrated and flexible manner. 40 The latter stages have increased sensitivity to hippocampal dysfunctions. 14,41
In the Morris water maze, the TBI rats demonstrated significant deficits in measures of both, reference and episodic-like memories (Fig. 4; Fig. 8C). In addition, they showed delayed learning and spent longer time to reach the hidden platform(s) than the other groups. Despite training-related improvements in performance, the deficits of the TBI rats in finding the platform persisted up to the end of the training. As start locations varied every trial, these observations suggest that TBI rats could have difficulties re-orienting a spatial map as quickly as controls. The performance of the TBI rats in the probe trials is consistent with this interpretation as they showed significant deficits in spatial preferences as compared with Naïve controls; nevertheless, their preferences were significantly higher than a chance level.
In addition to the TBI, the Craniotomy group had deficits in the Morris water maze as well. However, the nature of craniotomy-induced deficits seems different than in the TBI group. Craniotomy rats were not impaired in latency to reach the platform demonstrating normal spatial learning and the ability to re-orient the spatial map to varied start positions. Nevertheless, the Craniotomy rats demonstrated deficits in the probe trials similar to the TBI rats. Thus, in the Craniotomy group, the deficits in the probe trials coexisted with the normal performance in the training trials and could reflect a poor acquisition of a waiting response for the platform rather than an impairment in spatial memory.
In addition, in the reference memory task, the Craniotomy rats performed poorly after an overnight delay, indicating an impairment in memory retrieval. This deficit was temporal and disappeared after additional training as the Craniotomy group demonstrated normal retrieval visiting previous platform locations during Repeated Reversals and Radial Water maze testing. The deficits were not debilitating, as performance of the TBI rats was above the chance level. In addition, the low accuracy of spatial exploration in the probe trials observed after craniotomy was not consistent with primary deficits in spatial memory. As poor performance in the probe trials can reflect not only impaired spatial memory but a response strategy that is not compliant with persistent presence in the vicinity of the platform, utilization of an additional spatial task that does not require acquisition of such response (like RWM) might be helpful in delineating deficits specific to spatial memory.
The Morris water maze deficits we found are comparable to those seen in other pediatric CCI studies in rodents. Adelson and colleagues found a significant Morris water maze deficit in both PND 7 and PND 17 rats, compared with craniotomy controls, with a graded response based on severity of injury. 42 Interestingly, one study evaluated long-term water maze deficits at 3-6 months following PND 17 CCI in rats. They did not find differences in cued or spatial learning between craniotomy controls and CCI rats. However, they did see that injured rats had fewer entries into the target area, and the cumulative distance traveled in spatial learning was negatively correlated with the number of target area entries in the probe trials. 29 Several studies have utilized water maze testing to demonstrate improved behavior with neuroprotective treatments after CCI, such as buspirone, 43 COX-2, minocycline, 44 ketogenic diet, 45 and flumazenil. 46 In contrast, some studies did not show improvements in Morris water maze deficits with well-studied treatments such as progesterone 47 and COX-2 inhibition. 48 Further, some developmental TBI studies did not detect any water maze deficits in their injured animals, when compared with craniotomy controls. These findings speak to the importance of the use of appropriate age-matched control groups, with inclusion of a naïve control group.
Similar to the adult CCI literature, many studies to date in immature rats have used only sham craniotomy controls. Schober and colleagues did include both craniotomy and naïve controls in water maze testing after CCI in PND 17 rats, with the two control groups performing similarly on hidden platform testing. 49 Other important factors that may affect variability of findings in water maze testing are the age at injury and testing 42,50 as well as specifics of water maze testing paradigms utilized. 29,51
Radial Water Maze is sensitive and differentiates to the effects of TBI and Craniotomy
Although the Radial Water Maze has been used in adult rodents after TBI 52,53 and after hypoxia-ischemia, 54,55 to our knowledge, there are no published studies of Radial Water Maze in immature rats after CCI, indicating that utilization of this task in the future research may increase sensitivity of study design to effects of TBI and efficacy of treatments. In our study, the rats were trained in the Radial Water Maze, in which most procedural aspects of the task were the same as in the preceding repeated reversals in the Morris water maze. For example, the location of hidden platform was changed daily, and the start position was varied between trials. However, the swimming area was structured by a radial maze enclosure, which allowed for the outcome measures that did not require a waiting response to the platform as in the previous tasks. In this setting, the TBI rats demonstrated delayed learning but, at the end of the 1st training session, they were as efficient finding the platform as control rats. Importantly, when memory for the platform was tested after an overnight delay, the performance of the TBI rats was similar to controls as well. These data documented that in the Radial Water Maze, the TBI rats were able to learn and remember the 1st platform location. Their performance, however, deteriorated when the platform location started to be switched daily.
Similar to the repeated reversal in the Morris water maze, the TBI rats were deficient in finding a new platform location up to the end of the daily sessions. After an overnight delay, the TBI rats were not able to locate the previous day's platform as efficiently as Naïve controls; however, when given more time/trials, the TBI rats demonstrated their spatial preferences to the previous platform. Importantly, the inability of the TBI rats to inhibit such preferences after the orientation trial (Trial 1) might be a reason for their underperformance. Similar deficits inhibiting previous platform visits were detected in the Craniotomy rats.
The Radial Water Maze task showed that an early-life injury to the somatosensory cortex resulted in impairments of episodic-like memory, observed as difficulties with an immediate recall and deficits inhibiting responses based on previous memory traces (Fig. 5; Fig. 8C). The latter deficit also was observed after pediatric craniotomy. It is difficult to conclude whether preserved reference memory of the TBI rats in the Radial Water Maze is due to task-related differences or additional training received during previous tasks. Irrespective of these possibilities, demonstration of specific impairments in episodic-like but not reference memory in the TBI rats suggests that the former is more sensitive to deteriorating effects of TBI.
New RWM protocol with multiple alternative choices uncovers disinhibition in TBI and Craniotomy
Because rats are able to show more sophisticated spatial learning than mice, 56,57 we explored the additional effects of TBI in more challenging conditions. To this end, trials with cued platform were introduced between hidden platform trials allowing for the analyses of competition between spatial (hippocampus-dependent) and goal-directed (caudate-dependent) strategies. As switching between these strategies requires medial prefrontal cortex (mPFC) activity, 58,59 this task was particularly suited to challenge mPFC interactions with hippocampus and nucleus Caudatus. The analysis of strategies employed in this task revealed that rats from all treatment groups memorized the platform location from the previous trial.
For example, in a hidden platform trial, the animals visited first the location of cued platform from the previous trial despite the fact that the cue was no longer present. This behavior indicated that the rats continued to use previously acquired spatial strategy independently of the type of trials (with hidden or cued platform). This behavior was consistent with a one-trial learning demonstrated by Naïve control, Anesthesia and, partially, Craniotomy rats at the preceding stage of the Radial Water Maze. In the new task, the rats revealed an amazing fit of their cognitive abilities, in that they were able to learn one-trial associations seven times in row. As expected from the previous tasks, the TBI rats were significantly impaired in their abilities to form one-trial spatial associations (Fig. 6; Fig. 8C). It is important to note that such spatial strategy was no longer adaptive for the new task; however, it was not easily abandoned. In the uncertain situation of multiple platform locations, the performances of Craniotomy and Anesthesia groups were clearly dissociated. Both groups had difficulties balancing spatial and goal-directed strategies. In the Anesthesia rats, the attempts to follow the cue coincided with delayed learning of the hidden platform location, whereas in the Craniotomy rats the spatial strategy remained dominant resulting in the best performance in the hidden platform trials at the expense of success in the cued platform trials. Low efficacy in the cued platform trials was one of the most sensitive measures dissociating Craniotomy rats from Naïve controls. The TBI rats were also impaired adopting goal-directed strategy leading to significant deficits in the cued trial learning.
Testing in a situation of multiple alternative choices revealed that TBI as well as Craniotomy rats had significantly higher rate of arm choices than Naïve and Anesthesia groups. Similar observations were made for the orientation trials at the previous stage of the Radial Water Maze. The higher rates of choices in the TBI and craniotomy groups suggest possible impairment inhibiting immediate motor responses. Comparison of deficits observed across multiple tasks revealed that TBI- and craniotomy-induced changes in rate of choices had big effect sizes supporting (Fig. 8C) biological significance of this behavioral phenotype.
Social recognition but not motivation is affected by TBI
Finally, we tested the long-term effects of early-age TBI on social interactions using two tasks (Fig. 7; Fig. 8C)—social recognition 19,60 and three-chamber social motivation 21 —that assess overlapping as well as differing aspects of social behaviors. Both of these tasks require preserved levels of social motivation to ensure appropriate performance. The protocols utilized for both of these tasks included stages of habituation in which animals were presented the opportunity to interact repeatedly with the same social stimulus. The later stages, however, differed between the tasks. In the social recognition task, a dishabituation stage required the animal to recognize a novel social stimulus using memory representations of the previous social stimulus. In contrast, the social motivation task included a stage of discrimination, in which a novel and already familial social stimuli were presented simultaneously reducing requirement for stable social memory traces.
Another important between-task difference was that only the social recognition task was based on reciprocal interactions of free roaming animals. The social recognition task proved to be more sensitive to the effects of TBI. In particular, TBI rats took longer to initiate the 1st social encounter and spent less time investigating novel social stimulus. As TBI rats did not show any changes in the level of social investigation in the social motivation task, the TBI-induced social deficit seemed to be limited to a setting with higher-risk reciprocal interactions. An additional TBI-related deficit was revealed at the stage of dishabituation when TBI rats failed to reactivate social behaviors with a novel conspecific. This finding could be interpreted as an impairment in social recognition memory, which would be consistent with TBI-induced impairments in memory documented in the previous tasks. However, considering the deficit in social interactions in the 1st trial, the alternative explanation could suggest the primary TBI effect on social motivation.
Recent developmental TBI studies have demonstrated deficits of social recognition and social motivation after injury. Using CCI in PND 21 mice, Semple and colleagues found that injury-induced social deficits emerged during development. When tested in adulthood, injured mice had a lack of preference for sociability in the three-chamber task, and a lack of preference for social novelty that were not seen when tested in adolescence. 5 In follow-up studies, this group also demonstrated important age-at-injury and sex-dependent effect of these post-injury sociability changes, with younger age and male sex having greater deficits. 61,62 Another study showed impairments in both social novelty and sociability in adolescence, following CCI in immature (PND 14) rats. 63 Similar to our results, but using a closed-head injury model of moderate severity in PND 11 rats, Runyan and colleagues showed social novelty recognition impairment, but did not find sociability deficits using 3-chamber testing at 4 and 8 weeks after CCI in both male and female rats. 60 Further, treatment with intranasal oxytocin before behavioral testing reversed the injury-related social recognition deficits. These studies, along with our results, would suggest that social interaction testing is an important component in defining post-injury behavioral profiles following TBI in pediatric models. Further, the translational relevance of social interaction testing after TBI, and its sensitivity to treatment therapies, support inclusion of this testing in future pediatric TBI studies.
Limitations of the study
There are a number of limitations that were not addressed in the current study. All experiments were carried out using only male rats, leaving possible sex-related differences untested. We were also unable to pretest the rats prior to injury, as they are developmentally unable to perform most of the tasks prior to CCI on postnatal Day 10. In addition, behavioral testing was performed sequentially rather than in parallel, which could limit direct comparison between tested outcomes, as they were tested at different times during maturation and in adulthood. Possible effects of enrichment must also be considered in this longitudinal study. The same animals were tested in a lot of tasks and handled from a very young age. The combination of these procedures can be viewed as cognitive and behavioral enrichment. Enrichment is known to improve inhibitory control, behavioral performance and decrease the likelihood of detecting cognitive deficits. 64,65 From this point of view, the enrichment could underestimate the effects of TBI and other treatments in this study. However, it has also been shown that cognitive benefits of enrichment can be higher in normal controls than in experimentally affected groups. 66 The latter consideration would have opposite expectations for the effect of enrichment, which can be specifically tested using combination of longitudinal and cross-sectional design. These questions will require further investigation.
Summary
Using detailed systematic analyses of learning and memory in multiple tasks/protocols, we demonstrated long-term cognitive and behavioral effects of pediatric TBI. Despite remarkable neuronal loss known to occur in the juvenile CCI model long-term, 29 the deficits in most cognitive tasks were not debilitating, a testament to the high adaptive capacities of the developing brain. The most sensitive cognitive processes affected by TBI included those that require fast one-trial learning and flexibility of acquired memory traces or response strategies. These processes are known to require multiple cortical and sub-cortical regions (such as prefrontal cortex, perirhinal cortex, dorsal striatum) 14,41,58,59 that are not affected by mechanical force of the CCI, however, are very likely to be involved in the long-term sequelae of pediatric TBI. The TBI effects observed in this study were compared not only with Craniotomy group, but also with Naïve and anesthesia controls. This experimental design allowed delineating additional deficits due to early-life craniotomy and anesthesia.
Very limited deficits observed in the Anesthesia group is consistent with a recent report on relatively safe profile of isoflurane used in this study 67 ; however, the effects of anesthesia might be more pronounced with prolonged exposure and different anesthetics. Considering that craniotomy did not involve any experimental damage to neural tissue, the long-term cognitive impairments were impressive as some of the effect sizes were comparable to those in the TBI rats. These remarkable effects of pediatric craniotomy suggest that in studies that use the craniotomy group as the only control group, multiple effects of TBI might be missed. Craniotomy-induced cognitive impairments were more readily detectable in more complex tasks indicating that these effects can also be missed if study design incudes only simple tasks with low cognitive demands. Our study demonstrates that inclusion of multiple control groups allows for better characterization of long-term effects of TBI on cognition. Noteworthy, the lack of Naïve controls in study design, a practice that is unfortunately widely accepted in the field, prevents addressing mechanisms of craniotomy-related deficits, which may be more responsive to treatments.
In conclusion, we identified short- and long-term deficits between post-natal Days (PND) 25 and PND 140 and found that TBI rats, when compared with naïve controls, had impairments across most tests studied. When TBI group was compared with Craniotomy as a control, the deficits were detected in a limited number of cognitive outcomes related to learning speed and cognitive flexibility. Notably, effects of craniotomy, when compared with Naïve controls, spanned across multiple tasks, and sometimes reached the effect sizes comparable to that of TBI. Peri-operative anesthesia exposure alone did not produce deficits. These results highlight the importance of selection of appropriate control groups in pediatric CCI models. In addition, the results demonstrate that performing a wide array of comprehensive cognitive testing improves the ability to detect deficits following developmental TBI.
Footnotes
Authors' Contributions
N.E.D. conducted behavioral experiments and handled data. T.P and O.C. helped with behavioral testing. M.S. performed sham and CCI surgeries. R.C.K. conceptualized the study and helped with organization, equipment, and funding. C.L.R. planned and organized the study and acquired funding support. A. S. designed and supervised behavioral experiments, analyzed and visualized the data. N.E.D., R.C.K., C.L.R., and A.S. wrote the manuscript.
Funding Information
This study was supported by funding from National Institute of Neurological Disorders and Stroke (R01 NS092747) to C.L.R. A.S. was also supported by RF1 A055974.
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Methods
Supplementary Figure S1
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
Supplementary Table S6
Supplementary Table S7
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.