
Abstract
Repeated subconcussive head impacts are frequently experienced by athletes involved in competitive sports, such as boxing. The objective of the present study was to investigate the changes in working memory performance and memory retrieval-related neural oscillations in boxing athletes who experienced repeated subconcussive head impacts. Twenty-one boxing athletes (boxing group) and 25 matched controls (control group) completed a modified visual working memory task, and their continuous scalp electroencephalography (EEG) data were collected simultaneously. The behavioral measures and retrieval-related low-frequency neural oscillations were analyzed at each working memory set size in both groups. Subjects in the boxing group showed a reduced mean accuracy, diminished capacity estimates, and slower reaction time at demanding set sizes, and a marginally increased intra-individual coefficient of variation (ICV) for overall set sizes. Additionally, decreased event-related frontal theta synchronization, parieto-occipital alpha desynchronization, and frontal low beta synchronization were observed in the boxing group, suggesting underlying working memory dysfunction for efficient neurocognitive resource employment, inhibition of distracting stimuli, and post-retrieval control in the boxing group. Moreover, a negative correlation was found between frontal beta synchronization and reaction time for most set sizes in both groups. The present study was the first to reveal the underlying working memory deficits caused by the cumulative effects of boxing-related subconcussive head impacts from the perspective of behavior and EEG time-frequency oscillations. Joint analysis of EEG low-frequency oscillations and the innovative task with multiple challenging load conditions may serve as a promising way to detect concealed deficiencies within working memory processing.
Introduction
Sports-related head impacts are usually experienced by individuals participating in a variety of confrontational sports competitions, 1,2 but they are only taken seriously by those experiencing them when the severity of the injury is clearly diagnosed as a concussion. Therefore, most of the points of interest in previous studies on sports-related mild traumatic brain injury (mTBI) were limited to concussions. 3,4 However, the severity of trauma for the majority of sports-related head impacts is at the subconcussive level. 5,6 The management of concussions and subconcussive injuries differs in that most of those who experience subconcussive injuries will not leave competition for systematic assessment and treatment. More importantly, after an initial exposure to mTBI, those experiencing them are more vulnerable to subsequent second and multiple exposures, 7,8 and many recent studies have demonstrated that repeated exposure to mTBI could significantly affect the behavioral performance and the structural integrity of the brains of those exposed. 9 –11 Hence, the underlying neurophysiological mechanism of the negative consequences of repeated subconcussive head impacts is worthy of in-depth investigation.
Boxing can be fully considered to be a classical collision sport that is directly associated with repeated subconcussive head impacts. Boxing athletes are subjected to repeated blows to the head in daily training and competitions, which provides an appropriate opportunity for researchers to investigate the neurophysiological mechanism underlying naturalistic exposure to repetitive subconcussive impacts. Recent research suggested that enhanced corticomotor inhibition, changed neural resource recruitment strategy, and deteriorated cognitive performance were detected in amateur boxing participants after they experienced three sparring bouts. 12 Additionally, a post-career study of boxing athletes suggested that the years of sparring and professional bouts were strongly correlated with diffusion tensor imaging (DTI) metrics of corpus callosum subregion white matter and fine motor dexterity, which also demonstrated that significant microstructural disruption and executive dysfunction as well as depressive symptoms can serve as distinct characteristics of boxing sequelae. 13 Moreover, a subset of individuals in boxing populations develop a series of neurodegenerative diseases, such as chronic traumatic encephalopathy (CTE) and dementia pugilistica. 14,15 Therefore, both short-term and long-term negative influences caused by boxing-related head impacts are conspicuous, but the underlying neural mechanisms are far from clear. Previous studies have performed several meaningful explorations of the neuroelectrophysiological characteristics of related conditions.
The low-frequency neural oscillations (theta, alpha, and beta bands) was the focus of previous research and was demonstrated to be the main frequency band that could be used to effectively distinguish mTBI characteristics. 16,17 Significantly suppressed alpha and beta desynchronization and enhanced theta desynchronization were reported in those who had sustained repeated mTBIs while they performed a task-switch paradigm, suggesting a striking change in neural communication for those individuals. 18 In addition, neural oscillation combined with neuroimaging research revealed that theta-band power reflecting cognitive control was closely associated with subthreshold structural aberrations in those with mTBI; moreover, the degree of phase synchronization for theta oscillation immediately after injury could effectively predict the level of symptom recovery. 19 Additionally, previous research was not limited to independent frequency bands, and an innovative combined frequency oscillation study was proposed. The theta-to-gamma relative power spectral density (PSD) ratio was employed to study mTBI, and athletes with a history of mTBI exhibited a significantly decreased frontal theta-gamma frequency ratio and degenerative behavioral measures on the task of visuomotor control, indicating that the specific frequency ratio could serve as a sensitive method to reveal the neurophysiological characteristics of sports-related mTBI for athletes with underlying deficits in executive and control function. 20 Moreover, recent research revealed a dose-dependent relationship in asymptomatic collegiate water polo athletes between low-frequency oscillation (combined delta and theta band)-based functional connectivity and repeated head impacts; individuals with the greatest head impact exposure exhibited significant changes in whole-brain functional connectivity, which was associated with the dysfunction of inhibition control and information processing. 21
Therefore, the aforementioned studies indicated that electroencephalography (EEG) low-frequency neural oscillations, especially theta-band oscillations, can be considered among the most sensitive characteristics for those with mTBI and repeated subconcussive exposure. However, to date, few studies have explored the neurophysiological mechanism of boxing-related repeated subconcussive head impacts with EEG time-frequency oscillation technology, especially in the domain of working memory.
Working memory is a core cognitive ability for information maintenance, retrieval, and manipulation within a short time, 22 and the capacity of working memory is limited and varies among individuals 23,24 ; moreover, prior studies have demonstrated that the behavioral measures and processing course of working memory are susceptible to damage from mTBI. 25,26 Therefore, a modified Sternberg (visual working memory) task was employed in the current research. For this task, a total of six difficulty levels (from set size 2 to set size 7) were used to precisely evaluate the dynamic change in working memory processing. Moreover, in addition to the individual's behavioral markers for reaction time (RT) and mean accuracy, the intra-individual coefficient of variation (ICV) and Cowan's K estimates could be calculated and analyzed by this task, to assess intra-individual variability and the working memory capacity for each subject in each set size. 27,28 Importantly, real-world images selected from open data sets 29 were adopted as the memory array for this task, which could pertinently reflect the real-world visual working memory conditions and could prevent subjects from using unsuitable memory tactics of certain phonological loops. Accordingly, this modified task was employed in the present study to better evaluate the processing capability of working memory for both boxing athletes and matched controls.
To date, there have been no published reports investigating the accumulative effects of repeated subconcussive exposure on working memory in boxing athletes by employing the EEG time-frequency analysis method. Therefore, in the present study, we combined a modified visual working memory task with multiple load effects and event-related time-frequency oscillation technology to explore the working memory characteristics and corresponding neuroelectrophysiological mechanisms underlying changes in athletes with repeated subconcussive exposures. We hypothesized that both the behavioral measures and time-frequency indicators could be modulated by the gradually intensified working memory demands for the two populations. A reduced mean accuracy and slower reaction time were predicted for boxing athletes within a certain set size, and the ICV and Cowan's K estimates were expected to show deteriorated effects for athletes as well. Moreover, with respect to the EEG time-frequency analyses for working memory processing, the specified regions of interest (ROIs) of low-frequency neural oscillations for event-related synchronization (ERS) and event-related desynchronization (ERD) were expected to show negative changes; specifically, the boxing group was anticipated to have attenuated power of frontal theta ERS and parieto-occipital alpha ERD compared with the control group, and slightly detectable changes in frontal beta ERS power were also expected for the boxing group within a certain set size. Further, significant correlations were expected between the neural oscillatory power of specific ROIs and the behavioral measures.
Methods
Subjects
Twenty-one qualified subjects for the boxing group were recruited from Wuhan Sports University, and 25 subjects for the control group were recruited from the surrounding colleges. The athletes in the boxing group had to meet the following inclusion criteria: (1) subjects had been engaged in boxing for >1 year and had experience in provincial or national championships, (2) subjects were between 17 and 25 years old, (3) subjects had undergone brain magnetic resonance imaging (MRI) (1.5 T) examinations to ensure that there were no physical abnormalities in the brain, and (4) subjects had to be right-hand dominant. The subjects were excluded if they met the following exclusion criteria: (1) subjects had been previously diagnosed with concussion or/and moderate to severe craniocerebral trauma, (2) subjects had a history of neurological or/and psychiatric conditions, (3) subjects were taking medicine, and (4) subjects had other conditions that are well known to impair cognitive function (e.g., chronic renal or hepatic conditions). Subjects in the control group had to meet the following inclusion criteria: (1) subjects were all non-athletes, (2) subjects had no history of repeated subconcussive head impacts or/and concussion, (3) subjects had no history of combat sports (e.g., boxing, kickboxing, or wrestling) and specified contact sports (e.g., football, ice hockey, and rugby), and (4) subjects met the same criteria for ages, brain MRI, and right-hand dominance as those in the boxing group. The control group had the same exclusion criteria as the boxing group. Moreover, all the subjects for both groups were male, the education levels of the two groups were matched, and normal color vision and normal or corrected-to-normal vision were required for subjects in both populations. Subjects completed the Mini-Mental State Examination (MMSE) before performing the working memory task for the purpose of screening general cognitive function. In addition, Raven's Standard Progressive Matrices (RSPM) was performed to assess the intelligence quotient (IQ) for each subject in both groups. Informed consent was obtained from each subject, and the study was approved by the Ethics Committee of the General Hospital of PLA Central Theater Command.
Working memory paradigm and procedures
A modified Sternberg task 30 was conducted by using E-prime 2.0 software (Psychology Software Tools, Inc., Sharpsburg, PA). The visual working memory task was implemented in a quiet, air-ventilated, and temperature-controlled room. The memory elements were displayed in a 3 × 3 cm array against a gray background (RGB: 128 128 128) on a 17-inch Dell monitor. Subjects sat relaxed, and the distance between the eyes and the monitor was ∼70 cm, which resulted in a visual angle of 2.46 degrees.
The schematic procedure of the modified visual working memory task is shown in Figure 1. The procedure includes six blocks in total, and each block was set to a specified working memory set size according to the number of real-world images in the memory array; that is, set size 2 required subjects to remember two images, and set size 3 required subjects to remember three objects. In a similar fashion, the difficulty gradually increased to the top load: set size 7. For a specific trial (taking set size 3 as an example), the procedure started with the presentation of a white cross for 750 ms (jittered between 500 and 1000 ms); consecutively, the memory array of three images was presented for 1000 ms, and the subject was asked to try their best to remember the images. Then, after a delay of 3000 ms, a probe of one image was presented, and the subject was instructed to distinguish whether the probe was included in the previous memory array (congruent trial) or not (incongruent trial) within 2000 ms; subjects' responses were instantly recorded as they clicked specified buttons to reflect their decisions. The probability of congruent and incongruent trials was equivalent (50% chance for each block) in each set size. The inter-stimulus interval (ISI) was 1000 ms, and a rest period of 90 sec was scheduled after each block was finished.

The procedure for the modified Sternberg task. The working memory phase and duration are labeled above and below each segment of the figure. According to the number of images presented in the memory array, the task had a total of six blocks, which correspond to six set sizes (set size 2 – set size 7). Here, the example shows three images in the memory array; namely, set size 3. ISI, interstimulus interval; msec, millisecond. Color image is available online.
Prior to formal acquisition of neuroelectrophysiological information, each subject was questioned to verify that he or she was acquainted with each element within the memory array. This verification was performed to ensure that each element would not cause obvious discomfort or have particular significance to the subject, to prevent undesired electrophysiological signals from being elicited. Subsequently, there was a preparative section for the actual operation of the task, with 60 trials that consisted of different set sizes, to help subjects familiarize themselves with the procedure. Feedback about the accuracy and RT of each trial was presented on the monitor for the preparative section, but the feedback was not presented in the formal procedure. The formal procedure was started when the mean accuracy exceeded 90%. There were six blocks within the modified task, and each block consisted of 60 trials; therefore, each subject was required to complete 360 trials in total. The entire experimental procedure took ∼1.25 h for each subject.
EEG acquisition
When the subjects performed the visual working memory task, synchronized EEG digital data were recorded with a 64-channel array elastic cap by employing Ag-AgCl electrode sensors, and the eegoTM amplifier (Germany) served for the acquisition of electrophysiological signals. The distribution of electrode sensors complied with an extended 10/20 international system. The continuous EEG recordings were referenced online to the CPz sensor, and the ground electrode was set to the AFz sensor. Signals from all sensors were digitized with a sampling rate of 1000 Hz, and the online EEG data were subjected to a bandpass filter of 0.3 ∼ 30 Hz (24 dB/octave, zero phase). The impedance of all sensors remained <5 kΩ during EEG acquisition.
Behavioral analyses
The mean accuracy and RT were calculated for each set size and each group, respectively, and all trials in which the performance was incorrect were ruled out for the precise calculation of RT and oscillatory power. Additionally, trials with an RT of <100 ms or >2000 ms were excluded to prevent pre-judgment and delayed reactions. The ICV was evaluated based on the mean (M) and standard deviation (SD) of the RT for each set size and each subject separately; thus, the ICV could reflect the intra-individual variability of behavioral measures. The computational formula for the ICV was as follows: ICV = SD (RT)/M (RT). 27 Moreover, Cowan's K estimate was the quantitative measure of an individual's capacity for visual working memory, which was calculated by the following formula: K = N × (H - FA). 28 The N within the formula indicated a specific set size, H was the hit rate (proportion of correct reactions to the incongruent trials), and FA reflected the false alarm rate (proportion of incorrect reactions to the congruent trials); therefore, this formula fully took into account the bias of lucky guesses and was appropriate for the modified working memory task with multiple load conditions and greater difficulty.
EEG data analyses
The offline EEG data for all subjects were processed via the EEGLAB toolbox. The preprocessing procedure of the offline data was performed with a high-pass filter for 0.01 Hz, a low-pass filter for 40 Hz, and a notch filter for 50 Hz. All EEG signals were further re-referenced to the mean of bilateral mastoid sensors (M1 and M2). Consecutive EEG data were segmented into epochs of 3000 ms in length that occurred from 1000 ms pre-stimulus to 2000 ms post-stimulus relative to the onset of retrieval. The apparent artifacts and the channels with strikingly abnormal signals (bad channels) within the segmented EEG data were identified by visual inspection. Then, trials with uncorrectable artifacts were excluded, each bad channel was spherically interpolated, and the upper limit for the number of interpolated channels was set to 10% (six bad channels) for each EEG data. Independent component analyses (ICAs) were conducted to distinguish and correct the artifacts of horizontal and vertical ocular movements, myoelectric activity, and other unwanted components. Moreover, based on the fact that only the trials with correct reactions could be included for further processing, the consequence was that there were only three relatively less challenging working memory load sets, including set sizes 2, 3, and 4, with sufficient correct trials (> 40 trials for each subject). Hence, this research investigates the time-frequency oscillations in individuals in both groups in response to just three set sizes.
Time-frequency decomposition was conducted for the approved artifact-free signals by employing customized scripts based on MATLAB (MathWorks, Inc.). Additional time points were attached to perfect the edge effect. Then, the short-time Fourier transform (STFT) with the Han window function was performed to obtain a time-frequency power calculation for each segmented epoch. The window length was set to 0.2 sec, and the frequency band ranging from 1 to 30 Hz was analyzed. Then, time-frequency power was averaged across epochs for each set size as the primary power data. Additionally, the averaged power for the time window prior to the onset of retrieval (-800∼-200) was calculated as the baseline power, which was subtracted from primary power data for baseline correction. However, to exhibit the relatively high frequency oscillations (beta band) more clearly, the baseline correction was adjusted by dividing by, rather than subtracting, the baseline power. Previous studies have demonstrated that frontal and parieto-occipital sensors are scalp ROIs for working memory. 31 –35 Moreover, the grand average for all the channels and subjects was calculated to construct the whole-brain time-frequency plot for each group, which indicates that the power of theta and alpha frequency bands was of conspicuous prominence. Additionally, the power of the low beta band was identified distinctly in the control group rather than the boxing group. Therefore, the present study investigated the difference for individuals in both populations in specified time-frequency ROIs: the power of theta band and low beta band oscillations were calculated within the time window for 200 ∼ 500 ms and 700 ∼ 1400 ms in the frontal region (average of sensors F1, F2, and Fz), respectively, and the power of alpha band oscillation was evaluated within the time window for 400 ∼ 800 ms in the parieto-occipital region (average of sensors PO3, PO4, and POz).
Statistical analysis
An independent t test was performed to analyze the homogeneity of the demographic and general characteristics between the athletes and controls. To test the hypothesis about the behavioral results in this research, boxing athletes were expected to have decreased behavioral performance compared with that of controls, and a two-way repeated-measures analysis of variance (ANOVA) (group factors [athletes vs. controls] × condition factors [set sizes 2, 3, and 4]) was implemented to assess the mean accuracy, RT, ICV, and Cowan's K estimate. For the EEG data of event-related neural oscillations, a two-way repeated-measures ANOVA (group factors [athletes vs. controls] × condition factors [set size 2, 3, and 4]) was performed to evaluate the time-frequency power for each ROI (theta ERS, alpha ERD, and low beta ERS) in each brain region (frontal and parieto-occipital regions); specifically, the theta and low beta ERS were calculated for the frontal region, and the alpha ERD was measured for the parieto-occipital region. Bonferroni correction was conducted for post-hoc comparisons. Greenhouse–Geisser correction was used to modify the sphericity assumptions. Pearson correlation was performed to evaluate the associations amonog behavioral measurements, general characteristics, and the neural oscillatory power of time-frequency ROIs.
Results
Demographic and general information results
The demographic and general characteristics of the two groups are presented in Table 1. There was no significant difference in the demographic measurements between boxing athletes and controls, suggesting that the homogeneity of the subjects in the two groups was suitable for classification and further comparison. In addition, the MMSE and RSPM results demonstrated no significant differences between athletes and controls, indicating that there were no pronounced differences in general cognitive function and estimated IQ for subjects in either group.
Demographic and General Characteristics of Boxers and Controls (Means ± SD)
SD, standard deviation; BMI, body mass index; MMSE, Mini-Mental State Examination; RSPM, Raven's Standard Progressive Matrices.
Behavioral results
The indicators of behavioral measurements, including mean accuracy, RT, ICV, and Cowan's K estimate, were detected for both groups in each working memory set size, and the results are displayed in Figure 2. For the results of mean accuracy, the analysis revealed significant main effects of group, F(1,44) = 7.18, p < 0.05,

The results of the behavioral measures.
For the RT results, the two-way repeated-measures ANOVA revealed significant main effects of group, F(1,44) = 9.92, p < 0.01,
For the ICV results, the analyses indicated significant main effects of set size, F(5,220) = 9.07, p < 0.001,
For Cowan's K estimate of working memory, the analyses revealed significant main effects of group, F(1,44) = 7.22, p < 0.05,
Time-frequency analysis results
The time-frequency power of each ROI, which consisted of theta ERS, alpha ERD, and low beta ERS, was calculated for each working memory set size in both groups. The grand average whole-brain time-frequency analysis and grand average event-related neural oscillation as well as corresponding topographical maps are displayed in Figures 3 –5.
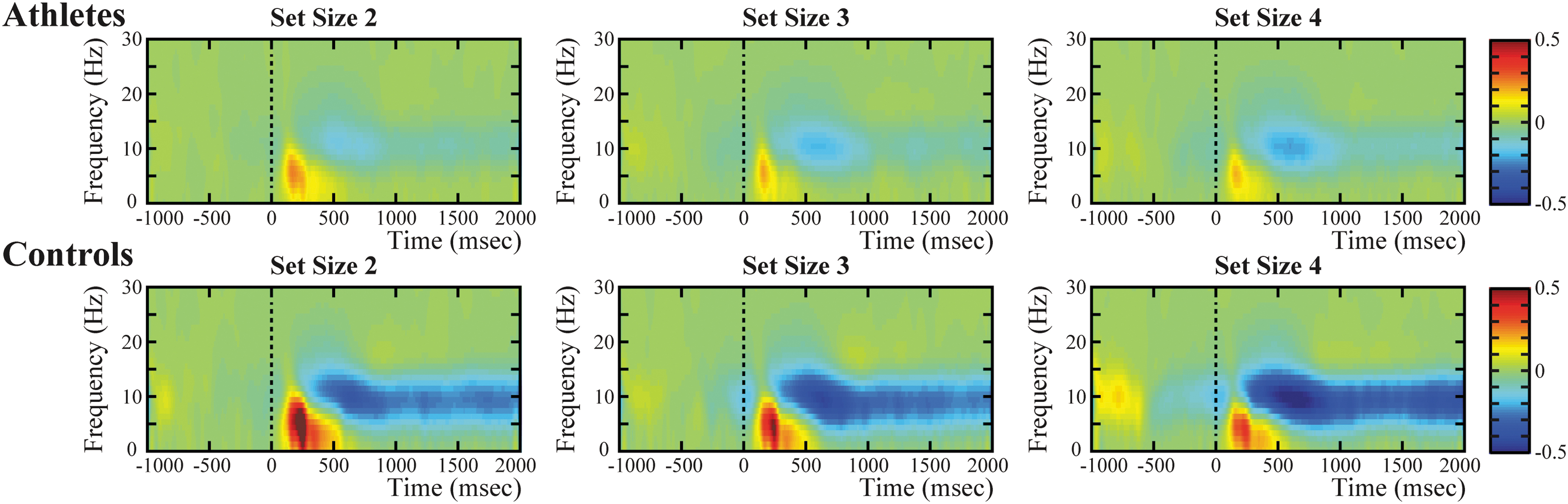
Whole-brain time-frequency analysis at each set size in both groups. The figures exhibit a striking synchronized theta oscillation and desynchronized alpha oscillation in both groups, as well as a slightly pronounced synchronized low beta oscillation only in controls. Msec, millisecond. Color image is available online.

Frontal theta ERS.
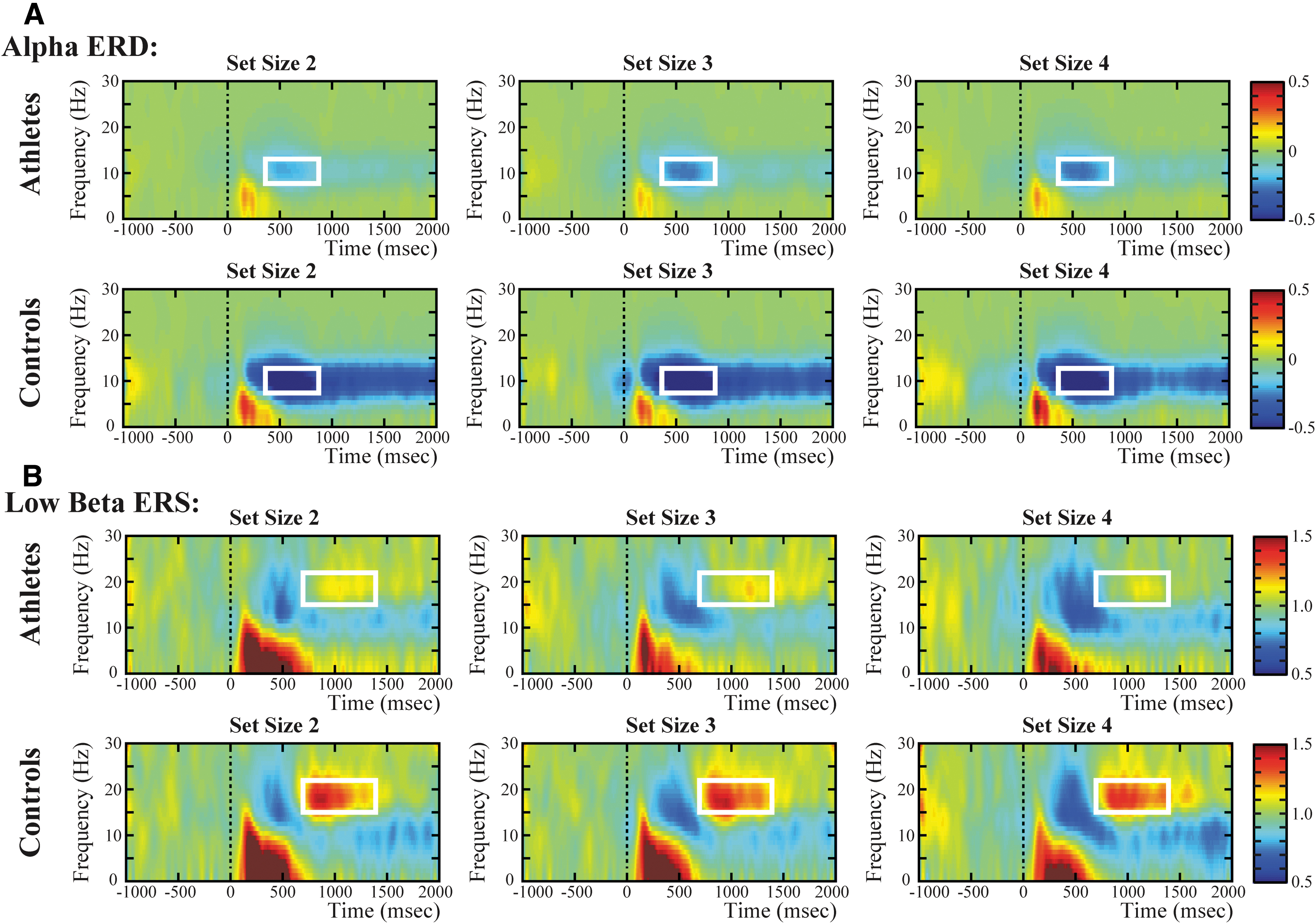
Parieto-occipital alpha ERD and frontal low beta ERS.
The grand average of the whole-brain time-frequency power, which was averaged across all 64 sensors and all the subjects for each group, was calculated to define specific ERS and ERD in memory retrieval processes (Fig. 3). The analysis showed that a striking synchronized neural oscillation (ranging from 2 to ∼9 Hz) and a salient desynchronized neural oscillation (ranging from 7 to ∼13 Hz) were induced in subjects in both the boxing and control groups; additionally, a slightly pronounced synchronized neural oscillation (ranging from 14 to ∼23 Hz) was detected in controls rather than athletes. To precisely explore the neural oscillatory changes related to working memory retrieval, based on previous research conclusions and combined with the present findings, our study further refined the time-frequency ROI as theta ERS and low beta ERS in the frontal region (average of the three frontal sensors: F1, Fz, and F2) and alpha ERD in the parieto-occipital region (average of the three parieto-occipital sensors: PO3, POz, and PO4).
For the theta ERS in the frontal region, the included ROI of theta synchronization was set to the average over 4 ∼ 7 Hz and across 200 ∼ 500 ms after retrieval onset. A significant main effect of group, F(1,44) = 10.27, p < 0.01,
For the alpha ERD in the parieto-occipital region, the included ROI of alpha desynchronization was set to the average over 8 ∼ 12 Hz and across 400 ∼ 800 ms after retrieval onset. Analysis suggested a significant main effect of group, F(1,44) = 4.92, p < 0.05,
For the low beta ERS in the frontal region, the included ROI of low beta synchronization was set to the average over 15 ∼ 22 Hz and across 700 ∼ 1400 ms after retrieval onset. Analysis revealed a significant main effect of group, F(1,44) = 5.64, p < 0.05,
Correlation analyses results
Pearson correlation analyses were performed to illuminate the relationship among each behavioral indicator, general characteristics, and each time-frequency power of the specified ROI for each group. Analyses demonstrated that the time-frequency power of low beta ERS in the frontal region was significantly correlated with RT for subjects in the boxing group in set size 2 (r = -0.47, p < 0.05) and set size 4 (r = -0.51, p < 0.05). Moreover, the frontal low beta ERS was negatively correlated with RT as well for subjects in the control group, with marginal significance in set size 2 (r = -0.39, p = 0.057), and set size 4 (r = -0.44, p < 0.05) (Fig. 6). This result indicated that the diminished power of low beta ERS was associated with the delayed reaction time in working memory retrieval for subjects in both the boxing and control groups. No other significant correlation was revealed between the time-frequency power of frontal theta ERS or parieto-occipital alpha ERD and behavioral measures or general characteristics for either population (for detailed data, see Supplementary Tables S1–S4).
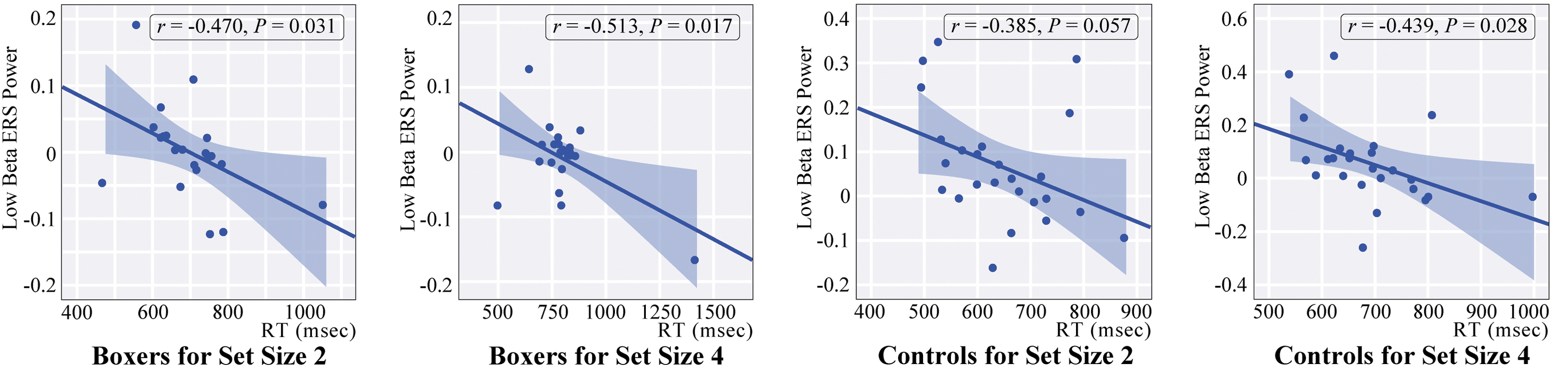
Correlations between the time-frequency power of low beta ERS and reaction time in both groups. ERS, event-related synchronization; RT, reaction time; msec, millisecond. Color image is available online.
Discussion
The present study aimed to investigate the accumulative effects of repeated subconcussive exposure on working memory in boxing athletes by employing EEG event-related neural oscillation technology. Real-world images were adopted as the memory array for the modified visual working memory task, which was successfully performed to assess the behavioral and electrophysiological characteristics of subjects in both the boxing and control groups. Regarding the results of behavioral measures, the current study found that athletes showed lower mean accuracy, delayed RT, and decreased Cowan's K estimate in relatively demanding set sizes than matching controls; additionally, marginally increased ICV was found for athletes compared with controls. For the time-frequency analysis, compared with the outcomes of controls, athletes revealed significantly lower induced theta ERS power in the frontal region for all set sizes. Notably, the theta power in set size 2 was significantly enhanced compared with set sizes 3 and 4, suggesting a load-dependent tendency toward theta band oscillation for both populations. Further, the time-frequency power of alpha ERD in the parieto-occipital region exhibited attenuated change (less desynchronization) in the athletes than in controls for all set sizes. Moreover, the conspicuously enhanced time-frequency power of low beta synchronization in the frontal region was revealed for the controls rather than the athletes. Crucially, significant negative correlations were found between frontal low beta power and RT in both groups, demonstrating that the decreased power of low beta ERS was correlated with delayed reaction speed. In brief, the athletes showed degenerated behavioral measures and weakened characteristics of event-related time-frequency oscillations when performing the modified task, indicating that the athletes had underlying abnormalities in the neurocognitive processing of working memory.
In terms of the behavioral measures, contrary to our expectation, the athletes with repeated subconcussive exposure exhibited comprehensive degeneration of working memory indicators, including mean accuracy, RT, and Cowan's K estimate, in relatively demanding set sizes. Therefore, the outcome of the present study was different from that of prior studies for soccer players with repeated subconcussive exposure, 36,37 which only showed a reduction in memory accuracy but without an increase in reaction times. Moreover, based on the balance theory between response accuracy and response time. 38 the concussed individuals in the current study could be considered to have sustained more serious damage to working memory than the soccer players in previous studies. Additionally, by means of the modified task with multiple challenging set sizes, the present results suggested that the behavioral differences between the two groups existed only in relatively challenging set sizes. This load-dependent damage effect for athletes may be closely associated with a decrease in working memory capacity. Pertinently, the capacity of an individual's working memory is finite; most individuals could maintain approximately four simple stimuli within a limited time. 24,28 For the present study, as a reliable and reasonable capacity estimate of working memory, Cowan's K estimate revealed that set size 5 was the memory storage ceiling for subjects in both populations, suggesting that the individuals failed to hold more than five real-world images within the working memory system, which essentially correlated with the findings of previous studies that used simple stimuli as a memory array; additionally, the compromised working memory capacity of the athletes in the present study may represent a prominent feature of boxing-related subconcussive injuries. Moreover, the ICV results showed that the athletes had slightly increased intra-individual variability for behavioral performance compared with controls, which was consistent with previous research conclusions for the go/no-go task for individuals with boxing-related head impact exposure. 39 Weakened stability of behavior indicated that neurocognitive function manipulated by the frontal lobe was significantly compromised by the cumulative effect of repeated subconcussive head impacts, 27 whereas the underlying neuroelectrophysiological evidence could contribute to further explaining the differentiated behavioral performances for both populations.
EEG theta band oscillation most likely originates from the hippocampus and the distributed functional networks involving the hippocampus, diencephalon, and neocortex, 40 which are closely associated with the operation of the central executive for working memory 41 and can reflect the capability of cognitive control and the efficiency of neural resource utilization. 42,43 The present study revealed that the boxing athletes had significantly decreased retrieval-related theta synchronization power for all set sizes compared with matching controls, indicating that the athletes had limited cognitive control abilities and had less efficient employment of neurocognitive resources. Pertinently, the affected cognitive control and processing efficiency exactly corresponded to the reduced mean accuracy and delayed RT within their behavioral performance. Moreover, frontal theta oscillation was significantly sensitive to subthreshold microstructural aberrations following mTBI, 19 which suggested that the attenuated power of frontal theta synchronization could be considered not only a time-frequency indicator of mTBI but also a consequence of impaired neural microstructure that resulted from repeated subconcussive exposure. Additionally, our results found that the time-frequency power of theta ERS significantly attenuated with increasing set sizes in both populations. This descending trend indicated a load-dependent effect for frontal theta ERS and suggested that the participants had increased exertion of limited neurocognitive resources and an overdraft of cognitive control capabilities with increasing working memory loads. Further, the descending trend for theta ERS power corresponded to the increasing tendency of frontal theta oscillation observed during the working memory maintenance period, 31 which further demonstrated that the frontal theta oscillation had specific load-dependent sensitivity for the different phases of working memory processes. Meanwhile, the theta oscillation induced by healthy individuals with low working memory capacity was shown to be more susceptible to being affected by challenging working memory loads than individuals with high capacity working memory, 42 indicating that the high working memory load effect could affect theta oscillation sensitively, especially for those with weakened working memory capacity. Pertinently, based on previous research, real-world images could reflect more complicated representations than the simple memory stimuli (such as numbers or colors) within the traditional working memory task 44 ; therefore, the current modified Sternberg task with real-world stimuli represented more challenging loads and could effectively detect the differences in theta ERS for athletes with potential working memory deficits. Consequently, the power of theta synchronization may be one of the sensitive measures associated with impaired cognitive control and processing efficiency for individuals who experienced sports-related subconcussive exposure.
In parallel, alpha oscillation is an indispensable component for the exertion and control of working memory, which interacts with theta oscillation to promote the efficiency of working memory processing. 32,33 The time-frequency power of alpha oscillations within the posterior cortical region has been closely correlated with the suppression of distracting and irrelevant information, 32,45 and with the inhibition of neurocortical excitability. 46,47 Importantly, the inhibition of distracting stimuli reflected by alpha oscillation serves to preferentially devote resources to task-related information and facilitate effective processing for working memory. 34 In the present study, the athletes had significantly diminished parieto-occipital power of alpha ERD for all set sizes compared with the controls, indicating that the cognitive processing efficiency for working memory retrieval was affected negatively in athletes with repeated subconcussive exposure. Similar to the findings of the present study, the sensitivity of alpha oscillation was also represented in repeated concussions; asymptomatic athletes with multiple concussed impacts showed a significant reduction in event-related alpha activity, and notably, the magnitude of the alterations in alpha power was closely related to the number of concussions. 48 Therefore, alpha oscillations could be thought to be significantly sensitive to repeated mTBI and were regulated by the cumulative effect of head impacts. Additionally, by employing a similar task, decrements in alpha ERD and theta ERS were found during retrieval in elderly subjects (mean age 65), 35 suggesting that the weakened power of alpha ERD within the modified working memory task may represent underlying degenerative changes in neurocognitive function. Moreover, one of the most representative indicators for aging-related degenerative changes was the attenuated event-related potential (ERP) P300, 49 which represents top-down modulated attentional resource allocation 50 and has suppressed changes in those with mTBI as well. 51 Interestingly, the magnitude of the P300 component was significantly correlated with the power of event-related alpha desynchronization. 48,52 Consequently, the cumulative effect of repeated head impacts on alpha ERD elicited by the visual working memory task could serve as a significant characteristic of the underlying degenerative neuroelectrophysiological damage.
Inconsistent with the event-related theta or alpha oscillations that were frequently noted, the novel finding within the current study was the results associated with working memory-related frontal low beta synchronization. The representation of cognitive function for beta oscillation was unlikely to be accurately described by a specific theory. The frontal beta oscillation elicited by the working memory task was thought to be associated with post-retrieval control operations, and to perform within a comprehensive frontoparietal network for post-retrieval control. 53 Additionally, it is related to the function of stopping retrieval and erasing working memory information when a trial was finished, to better prepare for the upcoming trials (stop and clear) 54 and to prevent unwanted information from entering into specific memory processes (protection). 55 Thus, based on this theory, our results showed a significantly decreased power of frontal low beta synchronization for subjects in the boxing group, indicating that the athletes with hypoactive beta ERS have attenuated capabilities of efficient “stop, clear, and protection” and reduced efficiency of recovery from event-related neural oscillations, which, consequently, may increase confusion in the working memory processes. Pertinently, these affected functions for post-retrieval control for each single trial may deteriorate monolithic behavioral performance in the working memory task, which could also explain the significant negative correlation between the time-frequency power of retrieval-related low beta ERS and reaction speed for most working memory set sizes in the present study. Meanwhile, this significant correlation for the two groups indicated a constant relationship between electrophysiological and behavioral characteristics in working memory retrieval processing for the modified Sternberg task, and the frontal low beta ERS power could serve as an important indicator that reflects the working memory processing ability of healthy subjects as well. In addition, previous studies have demonstrated that beta oscillations are mainly generated in the cortex, basal ganglia, or interactions and communications within the cortico-basal ganglia network. 54,56 Pertinently, the frontal cortex and white matter tracts are highly susceptible to damage from mTBI, 57,58 which could cause disrupted communication flow between the cortex and basal ganglia and cause a corresponding reduction in frontal beta oscillations. 59
Therefore, it can be concluded that the fundamental reason for the affected low beta synchronization and correlated behavioral deficits was closely associated with the underlying damage within the frontal cortex and basal ganglia-related white matter integrity caused by repeated subconcussive exposure.
Based on several limitations, the effects within the present work should be interpreted with caution. First, the electrophysiological information collected in the present study was based on scalp sensors, which cannot accurately locate the specific affected neuroanatomical areas associated with boxing-related injuries. Second, the degree of subconcussive exposure sustained by the athletes failed to be exactly quantified, although the years engaged in boxing, which was the accepted indicator in similar studies, 39,60 was specified as an indicator of the degree of exposure in this work. Pertinently, a customized cap with inertial sensors may be an ideal way to quantify the magnitude of each blow in future studies. 21 Third, there were insufficient trials in each set size for both groups, especially in the most challenging set sizes, although the design of this modified task is the result of comprehensive consideration of the task duration, number of set sizes, and sample sizes. In addition, adding an athlete group with no history of concussive and repeated subconcussive head impacts on the basis of the current study may be an ideal research protocol. More neuropsychological assessments and behavioral measures as well as longitudinal studies using the present results as the baseline should be considered in future studies.
Conclusion
Evidence for the differences between boxing athletes and controls in behavioristics and event-related neural oscillations was detected while the subjects performed a modified Sternberg task with memory arrays of real-world images. The athletes with repeated subconcussive exposure exhibited deteriorated behavioral measures in challenging set sizes and showed marginally increased intra-individual behavioral variability. Significantly diminished frontal theta ERS, parieto-occipital alpha ERD, and frontal low beta ERS power were found in the athletes, indicating that the athletes had deteriorated capability for efficient neurocognitive resource employment, inhibition of distracting stimuli, and post-retrieval control. These changes could result in a significant reduction in the efficiency of working memory processing. Moreover, the frontal low beta ERS was negatively correlated with RT, suggesting that the efficiency of recovery from event-related neural oscillations could also modulate the behavioral performance of working memory. Distinctly, the results of attenuated retrieval-related low-frequency neural oscillation could be attributed to the accumulative effects of repeated subconcussive head impacts sustained by the boxing athletes. Therefore, joint analysis of EEG event-related low-frequency oscillations and the modified task with challenging load effects may be a promising method to detect concealed deficiencies within working memory processing for athletes with related conditions.
Ethics Statement
All procedures contributing to this study followed the Helsinki Declaration and were approved by the ethical committee of Wuhan School of Clinical Medicine, Southern Medical University (China) (approved number: [2020]041-1). The protocol was fully explained to all participants, and written informed consent was obtained before the procedures.
Footnotes
Acknowledgment
We are grateful for the technical support received from the Department of Psychology, Southern Medical University, and for the contributions by participant recruitment from Wuhan Sports University.
Funding Information
This work was supported by the National Natural Science Foundation of China (81571049), the Chinese PLA technology innovation project (CLB18J042), and the Fujian Health Research Talent Training Project (2018-1-61).
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.