
Abstract
Traumatic spinal cord injury (TSCI) is a devastating traumatic disease of the central nervous system, which leads to refractory loss of motor and sensory function. So far, there is no effective treatment for TSCI. Recently, however, nano-sized exosomes from various spinal cord cells have shown great prospects in the treatment of various diseases, including TSCI. Microglia are one of the components of the spinal cord microenvironment. Anti-inflammatory microglia (M2) have been shown to inhibit inflammation and promote the functional recovery of spinal cord after TSCI. However, the role micro RNAs (miRNAs) in exosomes derived from M2 microglia in the treatment of TSCI is unclear. In this study, we investigated whether M2 microglial exosomes (M2-Exos) could better promote the functional behavior recovery of mice with TSCI than M0 microglial exosomes (Exos). Compared with Exos, M2-Exos were found to have a better effect in promoting the recovery of functional behavior, promoting axon regeneration and reducing the level of pyroptosis of spinal cord neurons after TSCI. Through a series of experiments, we also confirmed that miR-672-5p is the most critical miRNA associated with M2-Exos, and that its targeting gene is AIM2. M2-Exos rich in miR-672-5p could inhibit the AIM2/ASC/caspase-1 signaling pathway by inhibiting AIM2 activity, so as to inhibit neuronal pyroptosis and finally promote the recovery of functional behavior in mice with TSCI. In conclusion, our study suggests that the application of M2-Exos may be a promising treatment strategy for TSCI.
Introduction
Traumatic spinal cord injury (TSCI) is an incurable traumatic disease of the central nervous system (CNS), which leads to limb paralysis and even death. 1 To date, there is an estimated 23 million people afflicted with TSCI worldwide, 2,3 with the mortality rate of TSCI patients with quadriplegia as high as 20%. 4 The spinal cord microenvironment is where normal cellular metabolism and functional activities, including the regulation of cytokines, ion concentration, pH value, oxygen concentration, and blood supply, occur.5.6 The pathophysiological mechanism of TSCI is complex, but its fundamental mechanism is the imbalance of the spinal cord microenvironment and death of nerve cells in the injured area. 7,8 Reversing the imbalance in the spinal cord microenvironment and inhibiting nerve cell death are important directions for the study of TSCI therapy.
Microglia are not only important glial cells in the spinal cord microenvironment, but also key immune cells. Studies have shown that microglia increased at Days 3 and 7 post-TSCI, and the inflammation peaked at Day 7. 9 -11 Taken together, the data indicated that activation of microglia was an important factor in the acute (within 3 days) and subacute (4-14 days) phases after TSCI. 7,8,11,12 After TSCI, resting microglia (M0) are activated. According to different spinal cord microenvironment states, microglia undergo various morphological and functional changes, and finally polarize into pro-inflammatory microglia (M1) and anti-inflammatory microglia (M2), both of which are in a state of dynamic balance. 13 -15 M1 secrete pro-inflammatory cytokines, including tumor necrosis factor (TNF)-α, interleukin (IL)-1β, and IL-6. 16 M2 secrete anti-inflammatory cytokines, including IL-4, IL-10, and transforming growth factor β. 17,18 M1 inhibit, while M2 promote, neurogenesis. 19 Therefore, focusing on the role of M2 in promoting neurogenesis and inhibiting inflammation is of great significance for the treatment of TSCI.
Neurons are the key cells of the spinal cord that conduct information throughout the body, and are also the core component of the spinal cord microenvironment. 5 Programmed cell death (PCD) of neurons after TSCI leads to the loss of individual motor sensory function. 20 Pyroptosis, a newly identified form of pro-inflammatory PCD, is widely involved in the immune response of CNS diseases. 21 Pyroptosis includes the caspase-1-mediated inflammatory corpuscle pathway and caspase-4/5/11-mediated inflammatory corpuscle pathway. 21 -23 After TSCI, it was shown that a variety of pattern recognition receptors (PRRs) were activated and apoptosis associated speck like protein (ASC) and caspase-1 precursor were recruited to assemble inflammatory corpuscles. 24
Next, caspase-1 precursor was cleaved into activated caspase-1, which promoted IL-1β/18 precursor conversion to IL-1β/18, eventually leading to pyroptosis. 23 Common PRRs include nucleotide binding oligomerization domain like receptors (NLRs) and absent in melanoma 2 like receptors (AIM2), NLR-protein 1 (NLRP1) and AIM2 are expressed in neurons. 25 Recent studies have shown that AIM2 inflammatory bodies are activated in neurons of patients with CNS injury. 26 Inhibition of AIM2 expression can reduce pyroptosis of spinal cord neurons induced by ischemia-reperfusion. 27 Further, inhibiting the expression of AIM2, and thus inhibiting neuronal pyroptosis, is an important method to reduce secondary spinal cord injury (SSCI) and improve spinal cord function after TSCI.
Exosomes are nanoscale membrane vesicles secreted by cells and originate from the regulation process of endocytosis fusion efflux. 28 Exosomes are one of the important ways of intercellular signal transduction and paracrine signaling. 29 Exosomes have cell type dependent specific proteins, including CD9, CD63, CD81, and TSG101. 28,29 Studies have shown that exosomes from various cell sources can inhibit the spinal cord microenvironment inflammatory response after TSCI, reduce PCD levels, 30 inhibit the activation of M1 and pro-inflammatory reactive astrocytes, 31 and promote the transformation of microglia from M1 to M2, 32 all of which reduce the damage of TSCI and promote the recovery of spinal cord nerve function. It was found that M1-derived exosomes promoted pyroptosis of renal epithelial cells after acute renal injury, while M2-derived exosomes had the opposite effect. 33
Here, we speculate that microglia can regulate neuronal PCD after TSCI, especially inflammation related pyroptosis, by secreting exosomes. At present, studies on the treatment of TSCI with exosomes from different cell sources, and studies on the relationship between microglia and neuronal cells, are increasing gradually. However, there are few studies on the treatment of TSCI and the regulation of neurons by M2-derived exosomes. Therefore, we conducted an in-depth mechanistic study on M2-derived exosome regulation of the spinal cord microenvironment and its potential as a treatment for TSCI.
Recent studies have focused on the potential mechanisms of exosomal contents, including RNA and proteins, in the treatment of various diseases. 34,35 Studies have shown that exosome-derived micro RNA (miRNA) can regulate target cells. 35 However, the potential mechanism by which M2-exosome (M2-Exos) derived miRNAs regulate TSCI remains unknown. In this study, we extracted the exosomes of M0 and M2 to study whether they have a potential role in the treatment of TSCI. Our results showed that M2-Exos promoted the recovery of spinal cord function, and that this effect was closely related to the regulation of the spinal cord microenvironment and inhibition of neuronal pyroptosis. Using miRNA microarrays, it was found that miR-672-5p was the most differentially expressed M0-exosome (Exos) derived miRNA and which could inhibit neuronal pyroptosis in vivo and in vitro.
Accordingly, experimental results indicated that knocking down miR-672-5p in M2-Exos (miRKD-M2-Exos) could eliminate the inhibitory effect of M2-Exos on neuronal pyroptosis after TSCI. Meanwhile, AIM2 was identified as a targeting gene of exosomal miR-672-5p and several gain- and loss of function studies were carried out to verify this interaction. In conclusion, we found that M2-Exos could inhibit the activity of AIM2 via miR-672-5p and further inhibit neuronal pyroptosis after TSCI, thereby regulating the AIM2/ASC/caspase-1 signaling cascade, to promote the functional recovery mice after TSCI. This experimental study reveals a potential mechanism of M2-Exos in the regulation of the spinal cord microenvironment, and indicates their potential as a therapeutic target for TSCI.
Methods
Reagents and antibodies
The IL-4 for stimulating microglia to polarize to M2 type was purchased from Sigma Aldrich (St. Louis, MO). The lipopolysaccharide (LPS) and adenosine triphosphate (ATP) for stimulating neuronal pyroptosis were purchased from Sigma Aldrich. In this study, the antibodies used for Western blot included anti-Alix (Abcam, Cambridge, U.K.), anti-CD9 (Abcam), anti-TSG101 (Abcam), anti-Calnexin (Abcam), anti-IL-1β (Proteintech, Wuhan, China), anti-IL-18 (Proteintech), anti-caspase-1 p20 (Santa Cruz Biotechnology, Dallas, TX), anti-β-actin (Abcam), anti-AIM2 (Proteintech), and anti-ASC (Proteintech). The antibodies used for immunofluorescence included anti-F480 (Abcam), anti-NF200 (Abcam), anti-glial fibrillary acidic protein (GFAP; Cell Signaling Technology, Beverly, MA), anti-NeuN (Abcam), anti-IL-1β (Proteintech), anti-IL-18 (Proteintech), and anti-caspase-1 p20 (Santa Cruz Biotechnology). The antibodies used for flow cytometry included CD11b/c (Biolegend, San Diego, CA) and CD206 (Biolegend). The secondary antibodies for Western blot included horseradish peroxidase binding anti-rabbit IgG (Thermo Fischer Scientific; former Savant, MA) and anti-mouse IgG (Thermo). The secondary antibodies of immunofluorescence included goat anti-rabbit Alexa Fluor 488 (Jackson ImmunoResearch, West Grove, PA), goat anti-mouse Alexa Fluor 594 (Jackson ImmunoResearch) and goat anti-rat Cy3 (Jackson ImmunoResearch). The enzyme-linked immunosorbent assay (ELISA) kits included IL-1β, IL-18, and TNF-α (R&D Systems, Emeryville, CA).
Cell culture
The extraction and culture of primary neurons were performed as per our previous methods. 36,37 Briefly, the cerebral cortex of C57BL/6 mice was extracted and digested with 0.25% trypsin ethylenediaminetetraacetic acid (EDTA) solution (Gibco-BRL, Grand Island, NY) and DNase (Sigma Aldrich), after which the digestion was terminated with 10% horse serum (Gibco). After centrifugation, filtration, and resuspension, the neurons were seeded on culture plates and cultured in neurobasal medium (Gibco) containing 2% B27 (Gibco), 0.25% penicillin+streptomycin (Sigma Aldrich) and 1% glutamine (Sigma Aldrich). Half the media was changed every 2 days and the cells were cultured until the 7th day.
Similarly, the extraction and culture of primary microglia were performed as per our previous methods. 38 The cerebral cortex of C57BL/6 mice was extracted, after which the tissue was cut into pieces, digested with 0.25% trypsin EDTA solution and DNase, and then be centrifuged. The supernatant was then filtered and after resuspending, the cells were seeded in culture bottles containing Dulbecco's Modified Eagle Medium basic medium (Gibco) containing 10% fetal bovine serum (FBS; Gibco) and 0.5% penicillin+streptomycin (Sigma Aldrich). The medium was changed every 2 days and the cells were cultured until the 14th day, after which the culture bottle was shaken to obtain the supernatant, where the microglia were present. The purity of the microglial culture was then determined by labelling the cells with anti-F480.
Exosome isolation and identification
The M0 were stimulated with 20 ng/mL IL-4 for 24 h and polarized into M2. M0 (CD11b/c+) and M2 (CD11b/c+ CD206+) were identified by flow cytometry. The media of M0 or M2 media was collected and centrifuged at 300 × g for 10 min; after the temperature had been adjusted to 4°C, media was centrifuged for a further 10 min at 2000 × g. After centrifugation, the supernatant was filtered through a 0.22 μm sterile filter (Steritop™ Millipore, Billerica, CA). The filtered supernatant was then added to the upper chamber of an Amicon Ultra-15 Centrifugal Filtration device (Millipore) and centrifuged at 4000 × g, until the volume of liquid in the upper chamber was reduced to about 200 μL. Then, the ultrafiltration supernatant was washed twice with phosphate-buffered saline (PBS) and filtered again to about 200 μL. Next, the supernatant was pipetted into a sterile Ultra-Clear™ tube (Beckman Coulter, Asphalt, CA) and centrifuged for 1 h at 100,000 × g and 4°C using an optima L-100 XP Ultracentrifuge (Beckman Coulter). The fraction containing the Exos and M2-Exos were recovered using an 18-G needle, then diluted in PBS and centrifuged at 4000 × g and 4°C until the final volume reached 200 μL. Exos and M2-Exos were stored in a refrigerator at -80°C or immediately used for subsequent experiments.
The diameter of Exos and M1-Exos were tested and analyzed with a nanosight LM10 System (Nanosight Ltd., Navato, CA). The morphology of Exos and M2-Exos was also observed with a transmission electron microscope (TEM; Tecnai 12; Philips, Best, the Netherlands). The expression of the exosome-specific markers Alix, CD9 and TSG101 were then determined by Western blot.
Exosomes uptake by neurons
Exos and M2-Exos were respectively diluted with PBS, after which 4 mg/mL Dil solution (Molecular Probes, Eugene, OR) was added. The dilution was then centrifuged for 1 h at 100,000 × g and 4°C to dislodge excessive dye. Next, exosomes were resuspended with PBS and washed three times. The Dil-labeled exosomes were co-cultured neurons for 24 h, after which the neurons were washed with PBS for three times and fixed with 4% paraformaldehyde (PFA). The uptake of Dil-labeled exosomes by neurons was observed on a laser confocal microscope (Stellaris STED, LEICA, Weztlar, Germany) and the fluorescence intensity of the Dil was measured with ZEN lite software (Carl Zeiss AG, Oberkochen, Germany).
Vector constructs, lentivirus production and cell transfection
We used lentiviral vectors (GenePharma, Shanghai, China) to construct a negative control with the LV2 empty lentivirus (miR-NCKD) and an LV2-mmu-miR-672-5p-inhibitor vector (miRKD), and used miR-NCKD and miRKD to infect microglia, respectively, with an appropriate multiplicity of infection. Then, we constructed a short hairpin RNA (shRNA) vector for lentivirus gene transfer targeting mouse AIM2 (shAIM2) by GenePharma (Shanghai, China), and constructed a scrambled lentiviral construct as the negative control (shNC). The shAIM2 and shNC were used to transfected microglia, respectively, with Lipofectamine 3000 reagent (Invitrogen, Carlsbad, CA).
Mouse TSCI model and experimental groups
The animal experiments in this study were approved by the animal ethics committee of Nanjing Medical University (Nanjing, China). As previously study described, 37 male mice (C57BL/6, 8-10 weeks old) were used to establish a TSCI model. Animals were anesthetized by continuous inhalation of isoflurane, after which the spinal cord at T10 was exposed using laminectomy and a 5 g rod was dropped at a height of 6.5 cm using a spinal cord impactor (68097, RWD, CA). Immediately after the weight had impacted the spinal cord, the muscles and skin were sutured. The bladders of each mouse were then discharged manually three times a day until the micturition reflex recovered.
Mice were randomly assigned into several groups. Immediately after establishing TSCI model, the mice were respectively injected with PBS (200 μL), Exos, M2-Exos, miR-NCKD-M2-Exos, or miRKD-M2-Exos (200 μL PBS diluted 200 μg exosomal protein) by tail vein injection.
Functional locomotor scores
The functional behavior of mice was evaluated using various scales and at different time-points after TSCI. Briefly, the Basso Mouse Scale (BMS) was used to quantified mice neurological function at 1, 3, 7, 14, 21, and 28 days post-TSCI. The BMS scores ranged from 0 (complete paraplegia) to 9 (normal function). Specifically, the gross motor capability and coordination of the mice in the groups was measured at 28 days post-TSCI with an accelerating rotator (parameter range: 0-40 rpm). Each mouse repeated the experiment twice with an interval of 20 min. The average value of descent latencies was taken as the experimental result. The motor function recovery of TSCI mice was evaluated by a swimming test.
In brief, the mice were trained to swim from one end of a transparent tank filled with water to the other and the hindlimb swing, hindlimb movement, forelimb dependence, trunk stability and body angle of mice were measured and quantified with the Louisville swimming scale (LSS). Each mouse was tested twice, and the average value of the LSS score was taken as the experimental result. Footprint analysis was performed as previously study described. 39 The mice forelimbs and hindlimbs were soaked in blue and red ink, respectively. When the mice ran at a constant speed, the stride length and width were measured and analyzed. Each mouse was evaluated by two independent examiners who were unaware of the treatment regimen.
Immunofluorescence staining
To obtain the injured spinal cord, the mice were anesthetized and perfused with 0.9% normal saline and 4% PFA through the heart. The injured spinal cord was then removed with surgical instruments and fixed overnight in 4% PFA. The samples were dehydrated in 15% and 30% sucrose solution, each for 1 day, after which the spinal cord was into 12 mm thick slices. After being permeabilized with 0.05% Triton X-100 (Sigma Aldrich) and then being blocked with 10% bovine serum albumin (BSA; Sigma Aldrich), the spinal tissues/cells were incubated with primary antibodies (anti-NF200, anti-GFAP, anti-caspase-1 p20, and anti-NeuN for spinal tissues; anti-caspase-1 p20, anti-IL-1β, anti-IL-18, and anti-NeuN for cells) overnight at 4°C, after which the tissues/cells were washed with PBS for three times incubated with secondary antibodies for 2 h at room temperature (RT). Finally, the tissues/cells were washed with PBS for three times again and the nuclei were stained with 4′,6-diamidino-2-phenylindole (Thermo Fischer Scientific). A fluorescence microscope (AxioVertA1 and ImagerA2) was used to observe fluorescence and Zen Lite software was used to measure the average expression strength of each protein in areas near the injury center.
Quantitative real-time polymerase chain reaction (qRT-PCR)
TRIzol® reagent (Invitrogen) was used to extract total RNA from cells and exosomes. A reverse transcription system (Toyobo, Osaka, Japan) was used to synthesize complementary DNA (cDNA) and an ABI 7900 rapid real-time PCR system (Applied Biosystems, Carlsbad, CA) was used to perform qRT-PCR, which utilized SYBR Green PCR master mix (Applied Biosystems, Foster City, CA). The messenger RNA (mRNA) expression levels of interest were normalized to the internal control (β-actin) and the 2−ΔΔCT method was used to evaluate the relative expression levels. The specific primers for miR-672-5p, miR-136-5p, miR-21-5p, miR-218-5p, miR-338-3p, U6, IL-1β, IL-18, caspase-1, AIM2, and β-actin were all purchased from RiboBio Co, Ltd. (Guangzhou, China). The primer sequences are listed in Supplementary Table S1.
ELISA
To evaluate the expression levels of pro-inflammatory cytokines including IL-1β, IL-18, and TNF-α in the injured mice spinal cord, we extracted the spinal cord tissues at 7 days post-TSCI, which were then placed into a homogenizer with liquid nitrogen. We then incubated the tissue with lysis buffer composed of 1 mM EDTA, 10 mM Tris pH 8.0,1 mM phenylmethylsulphonyl fluoride, 1% Triton X-100, 150 mM NaCl, and 5 μL/mL protease inhibitor 1 h at 4°C. The lysate was then centrifuged at 1500 × g and the respective cytokine expression levels in the supernatant were measured. Various ELISA kits (IL-1β, IL-18, and TNF-α) was used to measure the pro-inflammatory cytokines in the microglial culture medium and a plate reader was used to measure the optical fluorescence or density.
Western blot analysis
Total tissue or cell protein was treated with radioimmunoprecipitation assay lysis and extraction buffer (KeyGen Biotechnology, Nanjing, China). A BCA kit (Thermo Fisher Scientific) then was used to determine the protein concentration, after which equal amounts of protein were separated by SDS-PAGE and transferred to polyvinylidene fluoride (PVDF) membranes (Millipore). The PVDF membranes were blocked with 5% BSA (Sigma Aldrich) for 2 h at RT and then incubated with primary antibodies overnight at 4°C. After washing three times with tris buffered saline with Tween, the membranes were incubated with secondary antibodies for 2 h at RT. ECL reagent (Thermo Fischer Scientific) was used to visualize reacting bands and ImageJ (National Institutes of Health, Bethesda, MD) was used to semi quantitatively analyze the density of protein bands.
Exosomal miRNA microarray assay
Microglia treated with Exos and M2-Exos were detected by a miRNA microarray from OE Biotech Company (Shanghai, China). Three samples were taken from each group for testing.
The miRNA microarray for microglia treated with Exos and M2-Exos were performed by OE Biotech Company (Shanghai, China). Three samples were processed in each group and analyzed with an Agilent-Mouse miRNA microarray 21.0 (8 × 60 K, Design ID: 070155).
The Affymetrix miRNA 4.0 platform (Santa Clara, CA) was chosen to conduct the microarray analysis. According to the instructions, the samples were labeled, hybridized, washed and analyzed (Agilent Technologies Inc., Santa Clara, CA). Both upregulated and downregulated genes were used a fold change cut off value of ≥1.5 set to identify expressions of different miRNA.
Luciferase reporter gene detection
MiR-672-5p binding sequences containing the wild type (WT) or mutation (MUT) and sequences corresponding to the 3′-UTR of AIM2 mRNA were synthesized by GeneScript (Nanjing, China). All of the sequences were cloned into the FseI and XbaI restriction sites of the pGL3 luciferase control reporter vector (Promega, Madison, WI, USA). Then, the AIM2 3′-UTR reporter constructs (pGL3-WT-AIM2 and pGL3-MUT-AIM2) were generated. After inoculation in a 24 well plate and incubation for 24 h, microglia were transfected with miRKD or miR-NCKD. The microglia were transferred to a 96 well plate and co-transfected with pGL3-MUT-AIM2 or pGL3-WT-AIM2 3′-UTR (dose 100 ng). A Dual-Luciferase® Assay Kit (Promega) was used to determine the firefly and Renilla luciferase signals.
Statistical analysis
All experiments were repeated at least three times. The data are shown as mean ± standard deviation. We used SPSS 25.0 and GraphPad software 7.0 to conduct statistical analysis. The unpaired two-tailed Student's t-test was chosen to compare two groups, and one-way or two-way analysis of variance followed by the Tukey's post hoc test was chosen to compare more than two groups to calculate the p values. In this study, a value of p < 0.05 indicated statistical significance.
Results
Identification and comparison of Exos and M2-Exos
Microglia were extracted from the brain tissue of C57BL/6 mice. The microglia and culture purity were identified by immunofluorescence staining with F480 antibody (Supplementary Fig. S1A). M0 and M2 were identified by flow cytometry. M0 microglia were CD11b/c-positive and CD206-negative, M2 were CD11b/c-positive and CD206- positive (Supplementary Fig. S1B). Exosomes from M0 and M2 were extracted, respectively. TEM showed that Exos and M2-Exos had similar spherical morphologies, with diameters of approximately 120 nm (Fig. 1A). Exos and M2-Exos had similar particle size distribution curves, as confirmed by nanoparticle tracking analysis (Fig. 1B). Exos and M2-Exos showed no significant differences in mean diameter, total protein content, exosome quantities per mL or exosome quantities per mg of protein (Fig. 1C). Western blot analysis showed that Exos and M2-Exos expressed the exosome surface marker proteins Alix, CD9, and TSG101, but not the marker calnexin. There was no significant difference of Alix, CD9 and TSG101 expressions between Exos and M2-Exos (Fig. 1D, 1E). Meanwhile, exosomes labeled with Dil dye were co-cultured with neurons for 24 h. The presence of exosomes in the cytoplasm of neurons was detected by laser confocal detection, which confirmed that exosomes could be ingested by neurons (Fig. 1F).

Characterization and contrastive analysis of Exos and M2-Exos.
M2-Exos promote the recovery of functional behavior after TSCI
Previous studies demonstrated that bone marrow mesenchymal cell (MSC)–derived exosomes promoted the functional recovery of mice after injury. 30 In this study, we investigated whether microglia-derived exosomes could promote the functional recovery of mice after injury, and whether M2-Exos showed better results than Exos. Functional behavior assessments were performed at designated time points after TSCI as shown in Figure 2A. On Day 28 post-TSCI, the functional recovery of mice in walking, hindpaw placement and hindlimb movement were judged by BMS scoring. Results indicated that the M2-Exos group was better than the Exos group (p < 0.001; Fig. 2B). The recovery of hindlimb function and tail balance in the M2-Exos group was increased compared with that in the Exos group, as assessed by rotarod testing (p < 0.01; Fig. 2C). LSS analysis indicated that the score of M2-Exos group was better than the score of Exos group in hindlimb movement, hindlimb swing, forelimb dependence, trunk stability and body angle during swimming, which confirmed that the M2-Exos group had better recovery of motor function after TSCI (p < 0.001; Fig. 2D, 2E). Footprint analysis showed that compared with the Exos group, the ipsilateral hindlimb step length of the M2-Exos group was longer and the contralateral hindlimb step width was smaller, which again confirmed that the recovery of hindlimb motor function in the M2-Exos group was better than that in the Exos group (p < 0.05; Fig. 2F, 2G). In addition, the motor function recovery of the M2-Exos and Exos groups after TSCI was better than that of the PBS group.
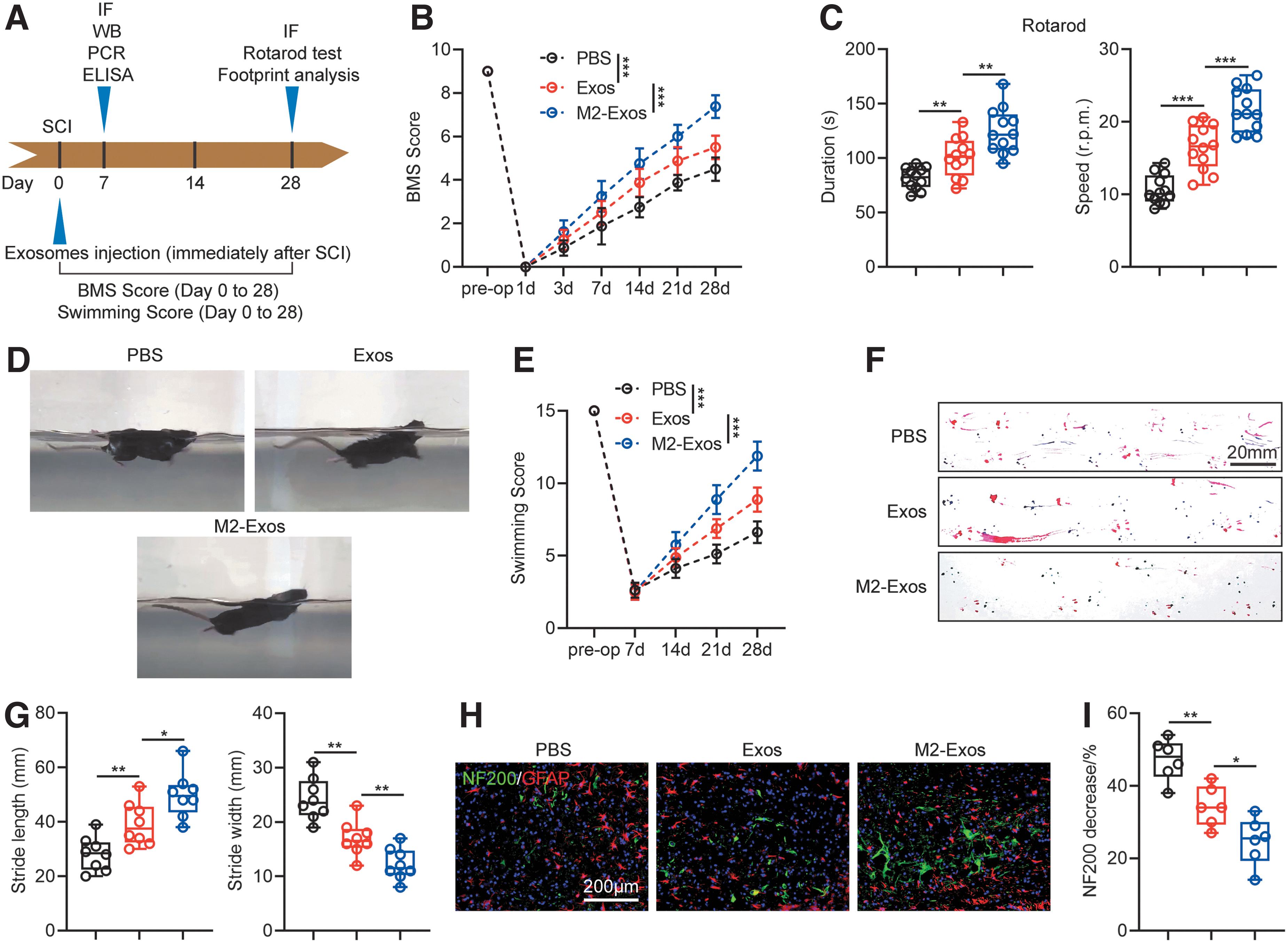
Administration of M2-Exos promotes better recovery of functional behavioral in traumatic spinal cord injury (TSCI) mice.
We further evaluated the morphology and density of neuronal axons in the central area of the TSCI to clarify the differences in neuronal functional recovery between Exos and M2-Exos groups from an anatomical point of view. Astrocytes were labeled with GFAP and neuronal axons were labeled with neurofilament-200 (NF200). The ratio of neuronal axon reduction in immunofluorescence of spinal cord was analyzed, and the results indicated that at 28 days post-TSCI, the number of NF200+ cells in the M2-Exos group was higher than that in Exos group, and the number of NF200+ cells in the two groups was higher than that in the PBS group (p < 0.05; Fig. 2H, 2I). Taken together, the experimental results show that M2-Exos are better than Exos in promoting hindlimb functional recovery and axon regeneration in mice after TSCI.
M2-Exos inhibit neuronal pyroptosis after TSCI in vivo
At 7 days post-TSCI, inflammation and pyroptosis levels in the spinal cord microenvironment reached a peak. The expression levels of caspase-1, IL-1β, and IL-18 in the groups were detected by qRT-PCR at 7 days post-TSCI. Results indicated that the expression levels of caspase-1, IL-1β, and IL-18 in spinal cord tissue of the Exos and M2-Exos groups were lower than those in the PBS group, and the degree of reduction in the M2-Exos group was lower than that in the Exos group (p < 0.05; Fig. 3A). Next, the expression levels of the pro-inflammatory cytokines IL-1β, IL-18, and TNF-α in the groups at 7 days post-TSCI were detected by ELISA. Results indicated that the expression levels of IL-1β, IL-18, and TNF-α in the Exos and M2-Exos groups were lower than those in the PBS group. Similarly, the reduction of levels in the M2-Exos group was lower than that in the Exos group (p < 0.05; Fig. 3B). Western blot was used to detect the expression levels of caspase-1 p20, IL-1β, and IL-18 in the spinal cord tissue of each group at 7 days post-TSCI. Similar to the PCR results, the expression levels of caspase-1 p20, IL-1β, and IL-18 in the Exos and M2-Exos groups were lower than those in the PBS group, and the degree of reduction in the M2-Exos group was lower than that in the Exos group (p < 0.01; Fig. 3C, 3D). The level of pyroptosis was determined by calculating the percentage of caspase-1 p20+ neurons in the total number of neurons. Immunofluorescence of spinal cord tissue after TSCI showed that, compared with the PBS group, the expression levels of caspase-1 p20 in the Exos and M2-Exos groups decreased, and the number of caspase-1 p20+/NeuN+ neurons, as a percentage of total NeuN+ neurons, also decreased. In addition, the pyroptosis of neurons in the M2-Exos group was lower than that in the Exos group (p < 0.001; Fig. 3E, 3F). In general, the results confirmed that M2-Exos could reduce pyroptosis of spinal cord neurons, and that this therapeutic effect was better than that of Exos.

Administration of M2-Exos following traumatic spinal cord injury (TSCI) inhibits neuronal pyroptosis in vivo.
M2-Exos inhibit neuronal pyroptosis in vitro
To determine whether M2-Exos have a therapeutic effect similar to that observed in vivo, prior to adding PBS, Exos, or M2-Exos, we first stimulated neurons with 1 μg/mL LPS for 5.5 h, and then stimulated neurons with 5 mmol/L ATP for a further 0.5 h. After about 24 h, the expression levels of caspase-1, IL-1β, and IL-18 in neurons were detected by qRT-PCR. The results indicated that Exos and M2-Exos reduced the expression of caspase-1, IL-1β, and IL-18 in neurons stimulated by LPS+ATP, and that the effect of M2-Exos was stronger than that of Exos (p < 0.01; Fig. 4A). The expression levels of IL-1β, IL-18, and TNF-α in neuronal supernatant were next detected by ELISA. The results indicated that both Exos and M2-Exos reduced the expression of IL-1β, IL-18, and TNF-α in neurons stimulated by LPS+ATP, and that the inhibitory effect of M2-Exos was stronger than that of Exos (p < 0.01; Fig. 4B).
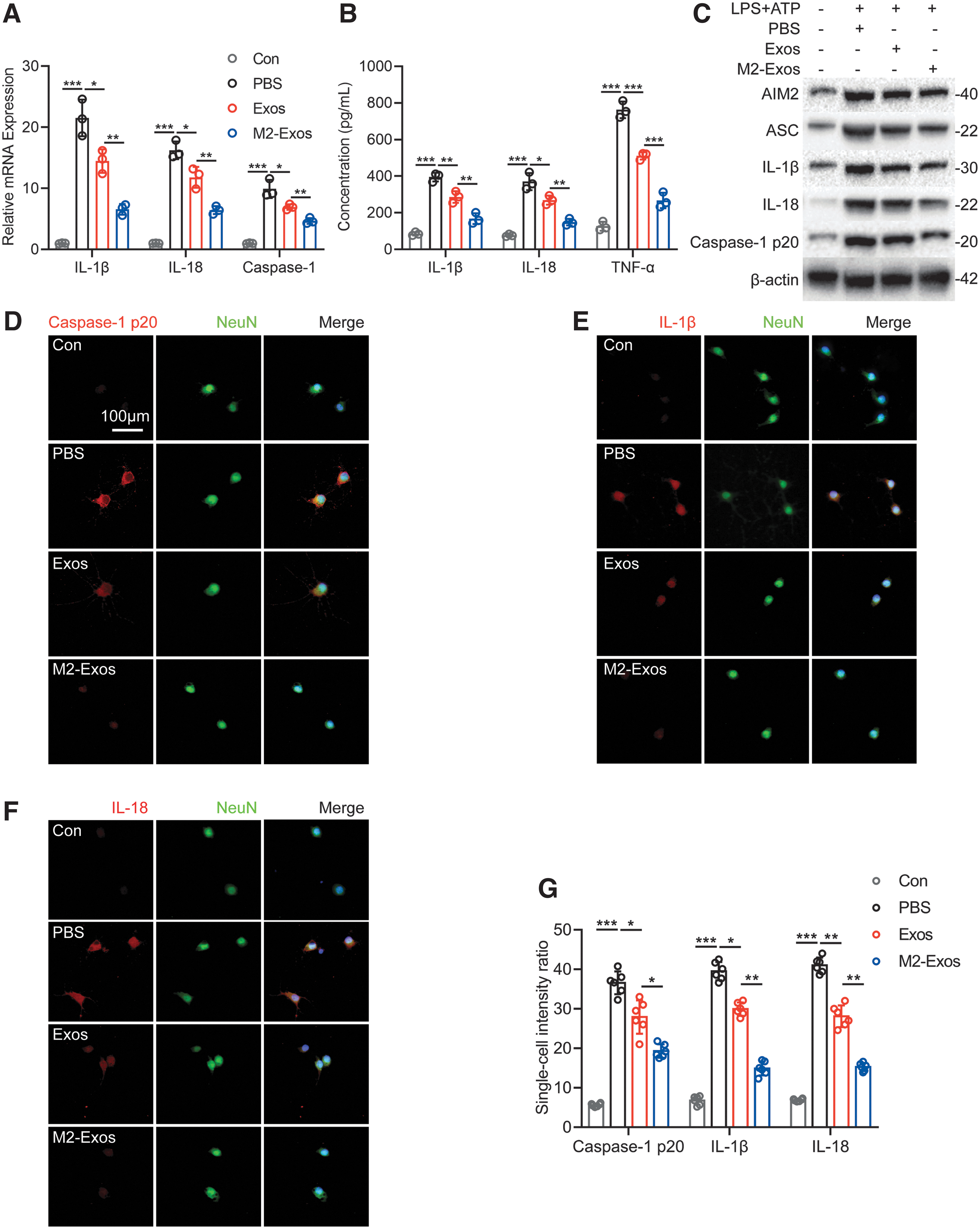
M2-Exos inhibits neuronal pyroptosis in vitro.
Total protein from neurons was extracted and detected by Western blot. It was found that after LPS+ATP stimulation, the expression of pyroptosis related proteins AIM2, ASC, caspase-1 p20, IL-1β, and IL-18 in neurons increased, while Exos and M2-Exos inhibited the expression of these proteins, especially M2-Exos (Fig. 4C). Immunofluorescence of neurons after LPS+ATP stimulation showed that Exos and M2-Exos inhibited the expression of caspase-1 p20, IL-1β, and IL-18, while M2-Exos had a clearer inhibitory effect (p < 0.05; Fig. 4D-G). According to the above experimental results in vitro, we confirmed that M2-Exos inhibited neuronal pyroptosis and protected neurons in vitro, results that were consistent the experimental results in vivo.
MiR-672-5p is upregulated in M2-Exos and transferred to neuron by exosomes
The above experimental results indicated that M2-Exos had a better effect on promoting the recovery of spinal cord function than M0 Exos, and could effectively inhibit neuronal pyroptosis. Previous studies have shown that miRNAs are functional components contained in exosomes and play an important role in cell communication and functional regulation. 40 Therefore, we further isolated RNA from exosomes of M0 and exosomes of M2 and performed a microarray profile analysis of miRNAs derived exosomes. The miRNAs microarray analysis indicated that compared with the Exos group, 844 miRNAs were upregulated and 367 miRNAs were downregulated in the M2-Exos group (≥ 1.5-fold change, p < 0.05; Fig. 5A). We selected five miRNAs with significant upregulation: miR-672-5p, miR-136-5p, miR-21-5p, miR-218-5p, and miR-338-3p and used qRT-PCR to verify their expression in exosomes. Results indicated that, compared with the Exos group, the expression of miR-672-5p, miR-136-5p, miR-21-5p, and miR-218-5p in the M2-Exos group increased, and the expression of miR-672-5p increased more significantly (p < 0.001; Fig. 5B). Based on the above results, we focused on miR-672-5p and further studied whether M2-Exos regulated neuronal pyroptosis through miR-672-5p.

MiR-672-5p is upregulated in M2-Exos and delivered to neurons through exosomes.
To verify the mechanism of miR-672-5p in the regulation of LPS+ATP induced neuronal cell death, we constructed an miR-672-5p knockdown (miRKD) lentivirus and its corresponding negative control (miR-NCKD). After transfection into M2, qRT-PCR was used to verify the transfection efficiency (Fig. 5C). Exosomes were extracted from M2 transfected with miRKD and miR-NCKD, named miRKD-M2-Exos and miR-NCKD-M2-Exos, respectively. The qRT-PCR results indicated that the expression of miR-672-5p in miRKD-M2-Exos was lower than that in miR-NCKD-M2-Exos (p < 0.001; Fig. 5D). miRKD-M2-Exos and miR-NCKD-M2-Exos were next added to neurons, respectively. The qRT-PCR results indicated that the expression level of miR-672-5p in neurons with miRKD-M2-Exos addition was lower than that in neurons with miR-NCKD-M2-Exos addition (p < 0.001; Fig. 5E). Based on the above results, miR-672-5p derived from M2 can be transferred to target neurons.
M2-Exos inhibits neuronal pyroptosis by transferring miR-672-5p both in vivo and in vitro
To study the effects of miR-672-5p on M2-Exos mediated functional behavior recovery and neuronal pyroptosis after TSCI in vivo, we further conducted a series of experiments. After treatment with miRKD-M2-Exos and miR-NCKD-M2-Exos, we examined the BMS and LSS swimming scores and analyzed the rotary and footprint test results. Results indicated that miR-NCKD-M2-Exos could promote the recovery of hindlimb function in mice after TSCI, while miRKD-M2-Exos counteracted these therapeutic effects (Supplementary Fig. S2A, 2B; Fig. 6A-D). Similarly, at 28 days post-TSCI, anatomical changes in the mouse spinal cord were analyzed by immunofluorescence (Supplementary Fig. S2C). The results indicated that the number of NF200+ cells in the injured area of mice in the miRKD-M2-Exos group was lower than that of mice in miR-NCKD-M2-Exos group (p < 0.01; Fig. 6E), showing that miRKD-M2-Exos could eliminate the promoting effect of M2-Exos on axon regeneration.

M2-Exos inhibits neuronal pyroptosis by transferring miR-672-5p in vivo and in vitro. CHECK
Then, we studied the function of miR-672-5p in regulating neuronal pyroptosis after TSCI. ELISA results indicated that the pro-inflammatory cytokines IL-1β, IL-18, and TNF-α were upregulated in the spinal cords of the miRKD-M2-Exos group compared with miR-NCKD-M2-Exos group (Fig. 6G). The results of qRT-PCR and Western blot showed that the expression of caspase-1, caspase-1 p20, IL-1β, and IL-18 in spinal cord tissue of the miRKD-M2-Exos group was upregulated compared with that in the miR-NCKD-M2-Exos (p < 0.001; Fig. 6F, 6H, 6I). Immunofluorescence was used to detect the expression of caspase-1 p20 at 7 days post-TSCI (Supplementary Fig. S3). Results indicated that the number of caspase-1 p20+/NeuN+ neurons, as a percentage of total NeuN+ neurons, in the miRKD-M2-Exos group was lower than that in the miR-NCKD-M2-Exos group (p < 0.001; Fig. 6J). The above in vivo experimental results indicated that M2-Exos inhibited neuronal pyroptosis via an miR-672-5p-mediated mechanism.
On the basis of the above in vivo experiments, we further studied the in vitro the ability of M2-Exos derived miR-672-5p to regulate neuronal pyroptosis. Compared with the miR-NCKD-M2 group, ELISA showed that the expression of the pro-inflammatory cytokines IL-1β, IL-18, and TNF-α in the miRKD-M2-Exos group upregulated (p < 0.01; Fig. 6L), while qRT-PCR and Western blot showed that the expression of caspase-1, caspase-1 p20, IL-1β, and IL-18 in neurons of the miRKD-M2-Exos group upregulated (p < 0.001; Fig. 6K, 6M). Similar experimental results were obtained for the cell fluorescence intensities of caspase-1 p20, IL-1β, and IL-18 (Supplementary Fig. S4A–C; Fig. 6N). Combining the in vivo and in vitro experiments results, it can be confirmed that M2-Exos inhibited neuronal pyroptosis via an miR-672-5p-mediated mechanism.
Exosomal miR-672-5p regulates AIM2 by directly targeting the 3′-UTR region
To consider the potential mechanism of exosomal derived miR-672-5p regulation of neuronal pyroptosis, we focused on the target gene of miR-672-5p in neurons. According to the miRNA target online database, we found that AIM2 was the likely target gene of interest. We established WT and MUT 3'-UTR sequences of AIM2 due to the potential binding sites, in order to verify whether the 3'-UTR of AIM2 is a direct target of miR-672-5p (Fig. 7A). The transfected neurons were then analyzed by luciferase reporter gene. When miR-672-5p was knocked down and co-transfected with the AIM2 WT luciferase construct instead of the MUT construct, the relative activity of luciferase increased (p < 0.01; Fig. 7B). The qRT-PCR and Western blot showed that knockdown of miR-672-5p increased the expression level of AIM2 mRNA and protein (p < 0.001; Fig. 7C, 7D), which further confirmed that AIM2 is the downstream target gene of miR-672-5p.

Exosomal miR-672-5p regulates AIM2 by directly targeting the 3′-UTR.
Exosomal miR-672-5p regulates neuronal pyroptosis through an AIM2/ASC/caspase-1 signaling cascade
We further conducted several in vitro gain- and loss-of-function experiments in order to explore the relationship between exosomal miR-672-5p and AIM2. To verify that exosomal miR-672-5p regulates neuronal pyroptosis through regulation of AIM2, we silenced AIM2 expression in neurons by using shRNA technology. qRT-PCR results indicated that the expression of caspase-1, IL-1β, and IL-18 in the miRKD-M2-Exos-shAIM2 group was lower than that in the miRKD-M2-Exos-shNC group (p < 0.001; Fig. 8A). Similarly, ELISA results indicated that the expression of the pro-inflammatory cytokines IL-1β, IL-18, and TNF-α in the miRKD-M2-Exos-shAIM2 group was lower than that in the miRKD-M2-Exos-shNC group (p < 0.05; Fig. 8B). These results indicated that miRKD-M2-Exos could eliminate the anti-neuronal pyroptosis effect of M2-Exos, and knockdown of AIM2 reversed this function.

Exosomal miR-672-5p regulates neuronal pyroptosis through an AIM2/ASC/caspase-1 signaling cascade.
The AIM2-ASC-caspase-1 inflammasome is an important mechanism mediating neuronal pyroptosis. We further detected the expression of the AIM2-ASC-caspase-1 inflammasome by Western blot. Results indicated that, compared with the miRKD-M2-Exos-shNC group, the expression of AIM2, ASC, caspase-1 p20, IL-1β, and IL-18 in the miRKD-M2-Exos-shAIM2 group decreased (Fig. 8C). Similar experimental results were obtained for the cell fluorescence intensities of caspase-1 p20, IL-1β, and IL-18 (Supplementary b SF4 Fig. S5A–C; Fig. 8D). Taken together, the results suggest that exosomal miR-672-5p participates in the M2 Exos-mediated anti-neuronal pyroptosis effect by targeting the AIM2/ASC/caspase-1 signaling cascade.
Discussion
TSCI destroys the normal structure and function of the patient's spinal cord and as a result, has a high disability and mortality rate. At present, the treatment of TSCI mainly focuses on the secondary spinal cord injury as there is no effective treatment for the primary spinal cord injury. Spinal cord microenvironment regulation is an important research direction of SSCI treatment strategy in recent years. Spinal cord microenvironment regulation includes inflammation, PCD, immune and nerve regeneration regulation. Here, we demonstrated that M2-Exos inhibited neuronal pyroptosis, improved the spinal cord microenvironment and promoted axon regeneration, resulting in the recovery of functional behavior in mice after TSCI. At the same time, M2-Exos downregulated AIM2 through miR-672-5p, inhibited the AIM2/ASC/caspase-1 signaling pathway and reduced neuronal pyroptosis, which, likewise, resulted in functional and behavioral recovery after TSCI.
Notably, our study showed that M2-Exos could regulate pyroptosis and promote the recovery of functional behavior after TSCI, which provides a feasible research direction for the treatment of TSCI. This research aims to study the effect and mechanism of exosomes secreted by M2 microglia in the treatment of TSCI. Therefore, in terms of grouping, it was mainly divided into M2-Exos treatment group, Exos treatment group, and PBS control group. In the part of mechanism research, it was mainly divided into shRNA intervention group and NC control group. In the part of phenotypic study in vivo, we mainly discussed the role of M2-Exos and Exos in the treatment of TSCI, without discussing the difference in spinal cord function between them and sham operation group. Although the manner of this grouping had no significant statistical impact on our comparison of the therapeutic effects of M2-Exos and Exos on TSCI. Surely, this grouping method does limit our research, which affects us to judge whether the M2-Exos therapeutic effect could realize the significant reversal and thorough treatment of TSCI.
Microglia are innate immune effector cells of the CNS and participate in the cellular immune process of nervous system. After TSCI, resting microglia are activated and polarized into M1 and M2, with M1 and M2 in dynamic balance. 13 -15 However, due to the lack of strictly defined scoring phenotypic criteria, the use of the terms M1 and M2 remains controversial. 41 Microglia polarization is dynamic in time and related to tissue microenvironment. This dynamic process is difficult to evaluate. 42 M2 can be further divided into four subclasses: M2a, M2b, M2c, and M2d. M2a microglia are the most classic M2 phenotype and which are induced by IL-4 stimulation. 43 -45 Increasing studies have indicated that M2 play a pivotal role in ameliorating the spinal cord microenvironment after TSCI and promoting functional recovery. Lima and colleagues 46 found that systemic administration of IL-4 decreased the number of M1 and increased the number of M2 in mice after TSCI. In addition, neuronal markers and oligodendrocyte markers were also increased. Kisucká and colleagues 47 demonstrated that the neural prognosis of TSCI patients was positively correlated with the expression of M2 markers and that regulating the microglial phenotype during the first week after TSCI was crucial for neuroprotection. Kobashi and colleagues 48 demonstrated that transplantation of M2 could effectively ameliorate the motor function of TSCI mice and repair the retrograde axonal transport from the neuromuscular junction to the upstream.
Based these previous research findings, we focused on M2 in the treatment of TSCI. Current studies consider that M1 and M2 have mutual transformation. Although transplantation of M2 for the treatment of TSCI has been reported, the spinal cord microenvironment after TSCI is complex, it is difficult to avoid the transformation of exogenous M2 into M1. In addition, exogenous M2 may also be harmful if the time, dose or location of transplantation are not optimally controlled. 49 Therefore, it is more feasible to regulate the expression of endogenous M2 or utilize the secretory products of M2 for the treatment of TSCI. Kobashi and colleagues found that more than 90% of IL-4 induced M2 to express CD206. Therefore, we used CD206 to label IL-4 mediated M2. 48
Exosomes are nanoscale membrane vesicles secreted by cells, which were first discovered and reported by Pan and Johnstone in 1983. 50 Exosomes are one of the important ways of intercellular signal transduction and paracellular secretion. 51 -53 Studies on exosomes in the treatment of CNS diseases have been gradually reported. Riazifar and colleagues 54 demonstrated that the exosomes secreted by IFN-γ stimulated MSCs reduced demyelination, upregulated the expression of Treg cells and induced anti-inflammatory and neuroprotective responses, indicating that that they can be used as s cell-free therapy to treat multiple sclerosis. Liu and colleagues 30 demonstrated that MSC-derived exosomes played a significant role in promoting angiogenesis, reducing neuronal apoptosis, inhibiting the inflammatory response, promoting axon regeneration and inhibiting glial scar and A1 reactive astrocyte activation. In addition, exosomes have unique advantages such as noncytotoxic effects, high drug loading capacity and low immunogenicity. 55 Therefore, the number of reports on the construction of drug delivery systems using exosomes to treat CNS diseases has increased in recent years.
Haney and colleagues 56 developed a new secretion-based catalase delivery system for the treatment of Parkinson's disease (PD). The delivery system provided significant neuroprotective effects in PD models both in vitro and in vivo. Alvarez-Erviti and colleagues 57 delivered small interfering RNA to the mouse brain by intravenous injection of rabies viral glycoprotein (RVG)-targeted exosomes, so as to knock out BACE1, a therapeutic target for Alzheimer's disease. In addition to therapeutic effects, exosomes can also have diagnostic effects. Fiandaca and colleagues 58 found that the development of Alzheimer's disease 10 years before the clinical onset can be predicted by detecting the expression level of P-S396-tau, P-T181-tau, and Aβ1-42 in neurogenic blood exosomes. In view of these results, we focused on M2-Exos to determine whether they regulate the spinal cord microenvironment after TSCI.
Pyroptosis is a type of PCD with pro-inflammatory and cytolytic properties, and which is involved in the immune response of CNS diseases. 59 Current studies have shown that pyroptosis plays a key role in a variety of neurological diseases and the pathogenesis of TSCI. 20,60 -62 Zheng and colleagues 63 found that CORM-3 inhibited the occurrence of inflammasome signals and pyroptosis, so as to reduce neuronal death and improve the recovery of motor function after TSCI. De Rivero and colleagues 64,65 demonstrated that pyroptosis regulated by NALP1 inflammatory bodies plays a regulatory role in the prognosis of cervical spinal cord injury in rats, and can also improve the histopathology after traumatic brain injury. Li and colleagues 27 demonstrated that AIM2 activates AIM2 inflammatory bodies and regulates neuronal pyroptosis through ectopic double stranded DNA in a mouse spinal cord ischemia-reperfusion model. Neurons mainly express AIM2 and NLRP1 inflammatory bodies, but whether they express NLRP3 is still controversial. 25,66 Current studies have shown that microglia play a pivotal role in the spinal cord microenvironment inflammatory response after TSCI, and that the level of neuronal pyrosis is closely related to the spinal cord microenvironment inflammatory response. Therefore, based on the study of the effect of M2-Exos on TSCI, we focused on whether M2-Exos can regulate neuronal pyroptosis and further explored their regulatory mechanism.
MiRNAs exist widely in organisms and can specifically bind to target mRNA, leading to its degradation or inhibiting its translation, which in turn regulate genes after transcription. 67,68 At present, there are a great number of studies on miRNAs regulating the expression of key pathway proteins after TSCI. Liu and colleagues 32 found that miR-216a-5p can pass through TLR4/NF-κB/PI3K/Akt signaling pathway to regulate microglial polarization and affect the recovery of functional behavior in mice after TSCI. Zhang and colleagues 69 constructed a three-dimensional fibrous hydrogel scaffold containing miR-132/miR-222/miR-431. It was found that the scaffold could promote axon regeneration and myelin regeneration through encapsulated miRNAs, which resulted in neural functional recovery in rats after TSCI. However, there are relatively few studies on pyroptosis after TSCI regulated by miRNAs, especially neuronal pyroptosis.
Sheng and colleagues 70 demonstrated that miR-22 could inhibit microglial pyroptosis by targeting gasdermin D, which, in turn, inhibited the inflammatory level and neurological damage after TSCI in rats. Xu and colleagues 71 found that TLR4 was activated after TSCI and promoted the expression of long noncoding RNA (lncRNA)-F630028O10Rik. This lncRNA is a competing endogenous RNA of the miR-1231-5p/Col1a1 axis, which can enhance microglial pyroptosis after TSCI by activating the PI3K/Akt pathway.
In this study, we used an exosomal miRNA microarray assay to determine miRNAs that were statistically significant expressed in M2-Exos compared with Exos, with miR-672-5p being the most statistically significant. Verify that miR-672-5p can be transferred from M2-Exos to target neurons. We next showed that knockout of miR-672-5p could antagonize the inhibitory effects on neuronal pyroptosis and functional recovery in TSCI mice by M2-Exos. Next, through experiments such as miRNA target online database and luciferase reporter gene analysis, we found that miR-672-5p regulated neuronal pyroptosis through the AIM2/ASC/caspase-1 signal pathway.
In conclusion, we discovered that M2-Exos promoted functional behavior recovery in TSCI mice and inhibited the inflammatory response of the spinal cord microenvironment through miR-672-5p. In addition, we found that the target protein of miR-672-5p is AIM2, and that miR-672-5p inhibited neuronal pyroptosis through the AIM2/ASC/caspase-1 signaling pathway by inhibiting the expression of AIM2. Our findings broaden the understanding of exosomal regulation of pyroptosis in the spinal cord microenvironment, and highlight the prospect of M2-Exos in the treatment of TSCI. We believe that M2-Exos may provide a new basis for direct interactions between microglia and neurons in the spinal cord microenvironment. At the same time, our study also complements the theoretical relationship between miRNAs and neuronal pyroptosis after TSCI. Therefore, the combination of miRNAs and M2-Exos may be a minimally invasive treatment strategy against TSCI.
Footnotes
Acknowledgments
We thank International Science Editing for editing this manuscript. We would like to thank the Core Facility of the First Affiliated Hospital of Nanjing Medical University for its help in the experiment.
Funding Information
This work was sponsored by the National Natural Science Foundation of China, Grant/Award Number: 82072437, 81772352; Postgraduate Research Innovation Program of Jiangsu Province, Grant/Award Numbers: KYCX19_1161, KYCX20_1434, KYCX21_1613.
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Table S1
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.