
Abstract
The auditory organs, including the tympanic membrane, cochlea, and central auditory pathway, are the most fragile components of the human body when exposed to blast overpressure. Tympanic membrane perforation (TMP) is the most frequent symptom in blast-exposed patients. The impact of TMP on the inner ear and central auditory system, however, is not fully understood. We aimed to analyze the effect of blast-induced TMP on the auditory pathophysiological changes in mice after blast exposure. Mice aged seven weeks were exposed to blast overpressure to induce TMP and allowed to survive for two months. All TMP cases had spontaneously healed by week three after the blast exposure. Compared with controls, blast-exposed mice exhibited a significant elevation in hearing thresholds and an apparent disruption of stereocilia in the outer hair cells, regardless of the occurrence or absence of TMP. The reduction in synapses in the inner hair cells, which is known as the most frequent pathology in blast-exposed cochleae, was significantly more severe in mice without TMP. A decrease in the number of excitatory central synapses labeled by VGLUT-1 in the cochlear nucleus was observed, however, regardless of the absence or presence of TMP. Our findings suggest that blast-induced TMP mitigates peripheral cochlear synaptic disruption but leaves the central auditory synapses unaffected, indicating that central synaptic disruption is independent of TMP and peripheral cochlear synaptic disruption. Synaptic deterioration in the peripheral and central auditory systems can contribute to the promotion of blast-induced hearing impairment, including abnormal auditory perception.
Introduction
Auditory dysfunction is a common complication after blast injury. 1,2 Among the various pathologies leading to blast-induced auditory dysfunction (BAD), sensorineural auditory damage is untreatable and is associated with a decline in the quality of life. 3 Clinically, tympanic membrane perforation (TMP) is the most frequently observed complication in patients exposed to explosions. 4 A TMP is considered a potential indicator of the severity of blast-induced neurological injury, because ears with TMP experienced a higher peak pressure of the blast wave. 5 One study, however, observed less BAD on the ruptured versus the intact side in cases of unilateral TMP, 6 despite the likelihood that the ruptured side experienced a higher peak sound pressure level (SPL). 7
Recent research in a chinchilla model has also revealed that, in the short term, TMP caused by multiple blast exposures might have a protective role against peripheral cochlear synaptopathy, indicating that TMP could buffer the harmful blast overpressure transmitted to the cochlea. 7 There are limited longer-term quantitative assessments and central pathophysiological observations regarding blast-induced auditory deterioration, however. 8,9 Further, a single blast exposure is ideally suited for investigating the sole effect of TMP on BAD, because a series of blasts would lead to blast exposure after TMP occurrence.
Therefore, a quantitative analysis of possible correlation between the occurrence of TMP and severity of BAD, in terms of both peripheral and central auditory damages, in TMP and non-TMP ears exposed to the same peak pressure of a single blast shock wave has never been performed.
In the current study, we investigated the central and peripheral auditory pathophysiology developing over the long term in TMP and non-TMP ears after a single blast exposure of the same intensity. The pathological evaluation assessed the integrity of the cochlear synapses of the inner hair cells (IHCs), which is closely related to the pathogenesis of tinnitus and hyperacusis 10,11 ; the morphology of the outer hair cells (OHCs), which directly affects the hearing threshold 4 ; the properties of auditory neurons (ANs); and the central synaptic connectivity of the ANs at the cochlear nucleus (CN) because these elements are frequently damaged after blast exposure. 8,12,13
Methods
Animals
Twenty-nine CBA/J mice (male, six-weeks old), whose hearing function was already matured, 14 were divided into three groups: (1) control ears group: ears without blast exposure (n = 16 ears); (2) TMP(+) ears group: ears of mice exposed to a blast wave at seven weeks of age that exhibited TMP (n = 26 ears); and (3) TMP(−) ears group: ears that displayed no TMP after blast exposure (n = 16 ears). All ears were removed for assessment at 15 weeks of age.
Blast exposure
To generate and expose the mice to the blast wave, we used our established blast-tube system (see Supplementary Experimental Procedures and Supplementary Fig. S1A). 13 In our previous study, mice were exposed to blast shock waves in a diagonally upward direction to prevent the occurrence of TMP. Consequently, mild and pure cochlear damage without TMP were observed. 13
In this study, to create the TMP model, we changed the direction of the blast shock waves such that the mice now received them from the front of their heads. The characteristics of the blast shock waves used were the same as those of the shock waves used in the aforementioned previous study (Supplementary Fig. 1A). Further, a face mask was used to avoid traumatic eye, lung, and airway injuries through the mouth and nose. The pressure waveform of the blast wave generated by our blast-tube system showed a sharp rise to the corresponding to the shock wave, followed by the positive peak pressure of 280 kPa and negative pressure (Supplementary Fig. S1B). The frequency of the median root mean squared amplitude of the blast wave generated by our blast tube was 195 Hz (Supplementary Fig. S1C).

Assessment of the spontaneous healing ability of the tympanic membrane ™. (
Cochlear function test
As cochlear function tests, measurements of the auditory brainstem response (ABR) and distortion product otoacoustic emission (DPOAE) were performed, as described in the Supplementary Experimental Procedures. The ABR is a type of an electroencephalogram signal that is generated in response to an auditory stimulus; it has been used widely to evaluate the auditory function within the cochlea and the auditory center. 15 The ABR wave I represents the summated response from the auditory nerve in the cochlea, while waves II–V represent the responses from the ascending auditory pathway. 16 In clinical settings and in research, DPOAE is commonly used to test the biomechanical gain functions of the OHCs in the cochlea; OHCs represent a biological motor that amplifies the motion of the sensory epithelium. 17
Histological assessment
Cochlear tissues were immunostained with primary anti-C-terminal binding protein 2 (CtBP2, BD Bio-sciences, San Jose, CA), anti-glutamate receptor 2 (GluR2, Millipore, Bedford, MA), anti-Caspr (Millipore, Bedford, MA), anti-NrCAM (Abcam, Waltham, MA), and anti-myosin7a (Proteus Biosciences Inc., Ramona, CA) antibodies, and then incubated with appropriate secondary antibodies labeled with Alexa Fluor (Molecular Probes, Eugene, OR), as described previously. 18 For the CN study, the brainstems were removed, and frozen transverse sections were prepared. The assessment of cochlear hair cells (HCs), synapses, ANs, and CN were conducted as shown in the Supplementary Experimental Procedures.
Statistical analyses
One-way analysis of variance (ANOVA) with correction for multiple comparisons using the Tukey post hoc test was conducted for the between-group comparisons when the data were normally distributed. Non-parametric Kruskal–Wallis test was performed for the non-normally distributed data. Statistical differences in DPOAE and ABR measurements, synaptic counts, and stereociliary disruption were analyzed using two-way ANOVA followed by the Tukey multiple comparison test. The effect size was calculated using the Cohen d value. All data are presented as means ± standard errors. The statistical significance level was set at a p value <0.05.
Results
Rupture and healing process of the TM
We assessed the spontaneous healing ability of the TM after blast wave exposure using a small digital endoscope (AE-C1 endoscopic system, AVS Co., LTD, Tokyo, Japan; Fig. 1A). A TMP was observed in 61.9% of the ears (26/42; unilateral TMP, 10 ears from 10 subjects, bilateral TMP: 16 ears from eight subjects) 1 day after blast exposure. Further, TMP was observed in 12 left ears and 14 right ears, indicating no significant differences in the susceptibility to TMP laterality among individual subjects (p = 0.75, Wilcoxon pair-wise test).
The average size of TMP was 18.0% of the total TM surface one day after exposure and gradually decreased after one to three weeks, indicating a substantial healing ability of the TM (Fig. 1B). The average duration for TMP closure was 2.2 weeks, and all TMP ears eventually exhibited complete closure. We observed a thickened TM at two months after blast exposure (Fig. 1C), however, and there were significant between-group differences in the thickness of the TM (F (2,12) = 86.22, p < 0.0001, and Cohen d = 3.90; one-way ANOVA; Fig. 1D).
The TMP(+) ears showed a significantly greater TM thickness than the control and TMP(−) ears (TMP(+) vs. control: p < 0.01 and Cohen d = 3.36; TMP(+) vs. TMP(−): p < 0.01 and Cohen d = 3.39; one-way ANOVA followed by the Tukey post hoc test). Further, we observed no evidence of fracture or dislocation of the ossicular chain in the middle ear after blast exposure (Supplementary Fig. S2).
Assessment of cochlear function
The TMP(+) ears were composed of unilateral TMP(+) and bilateral TMP(+) ears, while TMP(−) ears were composed of unilateral TMP(−) and bilateral TMP(−) ears. To elucidate the physiological differences between unilateral TMP(+) and bilateral TMP(+) ears and between unilateral TMP(−) and bilateral TMP(−) ears, we performed a three-factor ANOVA (unilateral or bilateral × TMP × frequency) for all ears.
No significant differences were observed in the physiological performances between the unilateral TMP(+) and bilateral TMP(+) ears and between the unilateral TMP(−) and bilateral TMP(−) ears (Supplementary Table S1). Thus, for all pathophysiological comparisons hereafter, the unilateral TMP(+) and bilateral TMP(+) ears have been grouped together as the “TMP(+) ears,” and the unilateral TMP(−) and bilateral TMP(−) ears have been grouped together as the “TMP(−) ears.”
To examine the effects of TMP caused by blast exposure on cochlear function, we measured the ABR and DPOAE thresholds in all ears. In ABR measurements, one day after blast exposure, the TMP(+) and TMP(−) ears revealed a significant elevation in ABR thresholds compared with those of controls (TMP(+) vs. control: p < 0.01 at all tested frequencies, Cohen d = 1.51; TMP(−) vs. control: p < 0.01 at all tested frequencies, Cohen d = 0.89; two-way ANOVA followed by the Tukey post hoc test; Fig. 2A). Further, the ABR thresholds in the TMP(+) ears were significantly higher than those in the TMP(−) ears (TMP(+) vs. TMP(−): p = 0.04 at 8.0 kHz, p < 0.01 at 11.3 kHz, and Cohen d = 0.57; two-way ANOVA followed by the Tukey post hoc test).
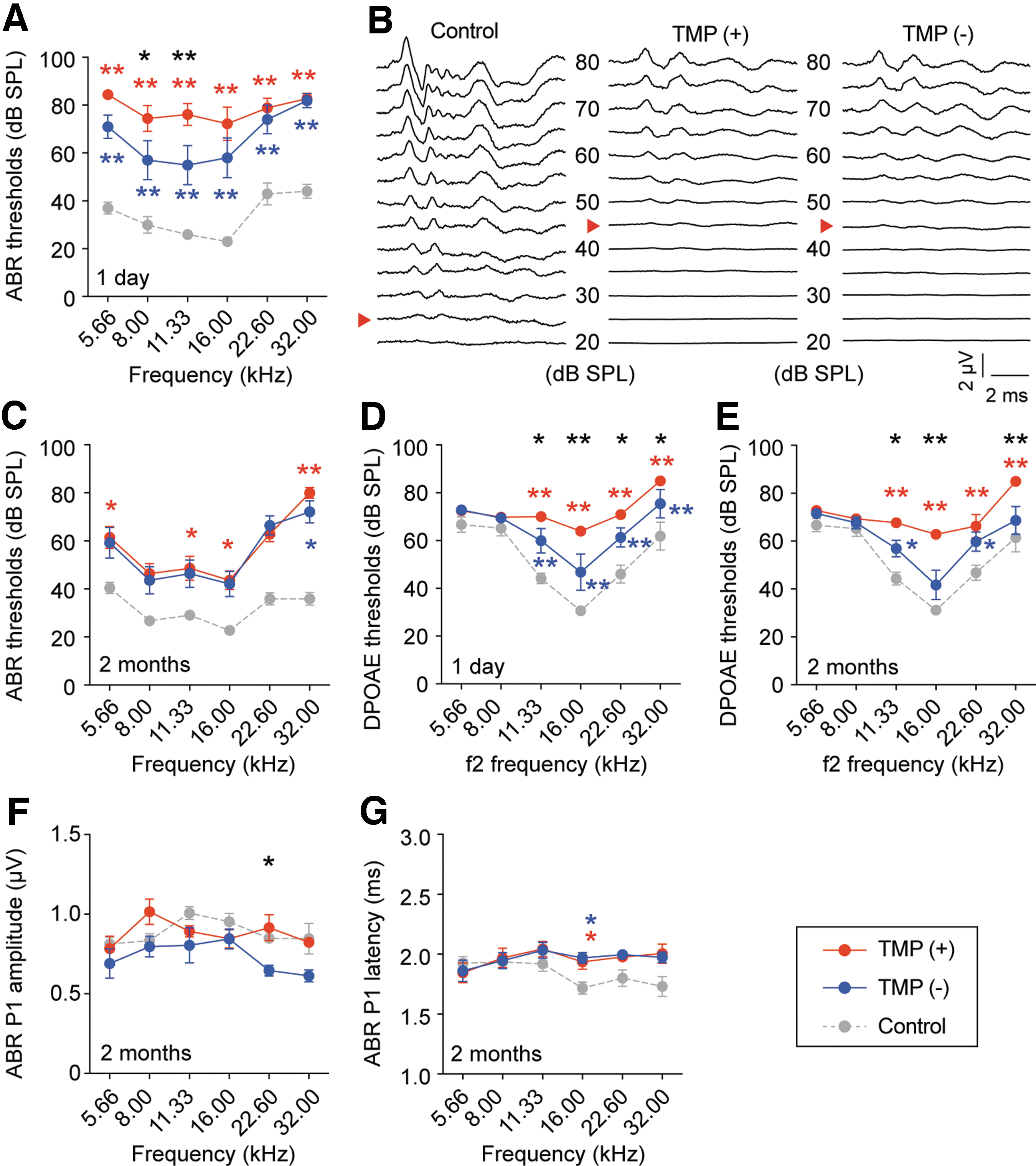
Cochlear functions in tympanic membrane perforation (TMP)(+), TMP(−), and control ears after blast exposure. (
Two months after blast exposure, the TMP(+) ears showed recovery of their ABR thresholds at a level similar to those of the TMP(−) ears, and there was no significant difference between the TMP(+) and TMP(−) ears at all frequencies (p = 0.2 and Cohen's d = 0.42; two-way ANOVA; Fig. 2B, C).
The ABR thresholds in both the TMP(+) and TMP(−) ears, however, were significantly higher than those in the controls (TMP(+) vs. control: p = 0.01 at 5.66 kHz and 11.3 kHz, p = 0.02 at 16.0 kHz, p < 0.01 at 32.0 kHz, and Cohen d = 0.66; TMP(−) vs. control: p = 0.01 at 32.0 kHz and Cohen d = 1.07; two-way ANOVA followed by the Tukey post hoc test).
Next, we measured the DPOAE thresholds in all ears. One day after blast exposure, the TMP(+) and TMP(−) ears exhibited a significant elevation in the DPOAE thresholds compared with the controls (TMP(+) vs. control: p < 0.01 at 11.3 kHz, 16.0 kHz, 22.6 kHz, and 32.0 kHz and Cohen d = 1.27; TMP(−) vs. control: p < 0.01 at 11.3 kHz, 16.0 kHz, 22.6 kHz, and 32.0 kHz and Cohen d = 1.10; two-way ANOVA followed by the Tukey post hoc test; Fig. 2D). Further, there was a significant difference in the DPOAE thresholds between the TMP(+) and TMP(−) ears (p = 0.03 at 11.3 kHz, p < 0.01 at 16.0 kHz, and p = 0.04 at 22.6 kHz and 32.0 kHz and Cohen d = 0.02; two-way ANOVA followed by the Tukey post hoc test).
Two months after blast exposure, although the TMP was completely closed in the TMP(+) ears, the DPOAE thresholds of the TMP(+) and TMP(−) ears remained significantly higher than those of the control ears (TMP(+) vs. control: p < 0.01 at 11.3 kHz, 16.0 kHz, 22.6 kHz, and 32.0 kHz and Cohen d = 0.50; TMP(−) vs. control: p = 0.04 at 11.3 kHz and 22.6 kHz and Cohen d = 0.35; two-way ANOVA followed by the Tukey post hoc test; Fig. 2E). Further, a significant difference between the TMP(+) and TMP(−) ears was also observed (p = 0.04 at 11.3 kHz, p < 0.01 at 16.0 kHz and 32.0 kHz, and Cohen d = 0.13; two-way ANOVA followed by the Tukey post hoc test).
These results indicated that the TMP(+) ears had a significant elevation of the DPOAE thresholds without significant elevation of the ABR thresholds compared with the TMP(−) ears, suggesting that the TMP(+) ears might have OHC dysfunctions as well as conductive pathology. The reason is that sound energy passes through the middle ear once to generate ABR and twice to detect DPOAE responses. Consequently, conductive pathology showed dynamic changes in DPOAE thresholds compared with ABR thresholds. 19,20
Next, to further investigate the cochlear function after blast exposure, we measured ABR P1 amplitude and latency, which are affected by AN function such as synapse integrity, myelination, and axon caliber. 21 To avoid the influence of threshold changes, the ABR P1 amplitudes and latency at 10 dB SPL above the thresholds were measured two months after blast exposure. 22 A significant decrease in the ABR P1 amplitude was observed in the TMP(−) ears compared with the TMP(+) ears (p = 0.04 at 22.6 kHz and Cohen d = 0.36; two-way ANOVA followed by the Tukey post hoc test) (Fig. 2F).
There was no significant difference, however, in the ABR P1 amplitude between the TMP(+) and control ears at all frequencies (p > 0.99, Cohen d = 0.02; two-way ANOVA followed by the Tukey post hoc test). In addition, the ABR P1 latency was significantly prolonged in the TMP(+) and TMP(−) ears compared with that in the control ears (TMP(+) vs. control: p = 0.04 at 16.0 kHz and Cohen d = 0.50; TMP(−) vs. control: p = 0.02 at 16.0 kHz and Cohen d = 0.50; two-way ANOVA followed by the Tukey post hoc test; Fig. 2G). , No significant difference, however, between the TMP(+) and TMP(−) ears was observed in the ABR P1 latency (p > 0.99, Cohen d < 0.01; two-way ANOVA).
Survival of cochlear HCs and ANs
Based on our ABR findings demonstrating blast-induced cochlear dysfunction, cochlear morphology was assessed (Supplementary Fig. S3A). There was no significant loss of IHCs and OHCs in any group (IHC, p = 0.07 and Cohen d = 0.27; OHC, p = 0.09 and Cohen d = 0.13; two-way ANOVA; Supplementary Fig. S3B). About 95% of ANs have synapses with IHCs via their peripheral processes. 23 Although IHC loss was not observed in our blast-exposed mice, to assess AN degeneration, the number of spiral ganglion neurons (SGNs) was measured (Supplementary Fig. S3C). The density of SGNs was not significantly different among the groups (F(2,18) = 1.74, p = 0.20, and Cohen d = 0.45; one-way ANOVA; Supplementary Fig. S3D).
Peripheral cochlear synapses
To investigate further the influence of TMP caused by blast exposure on the cochlear synapses between IHCs and SGNs, cochleae were immunostained with both CtBP2 and GluR2, which are the markers of cochlear pre-synaptic ribbons and post-synaptic membranes, respectively (Fig. 3A). Although numerous synaptic puncta were observed in the control ears, we found fewer puncta in the high-frequency regions of the ears in blast-exposed mice. The number of synapses was significantly lower in the TMP(−) ears than in the control and TMP(+) ears (TMP(−) vs. control: p < 0.01 at 16.0 kHz, 22.4 kHz, and 32.0 kHz and Cohen d = 0.42; TMP(−) vs. TMP(+): p = 0.01 at 22.6 kHz and Cohen d = 0.23; two-way ANOVA followed by the Tukey post hoc test; Fig. 3B).
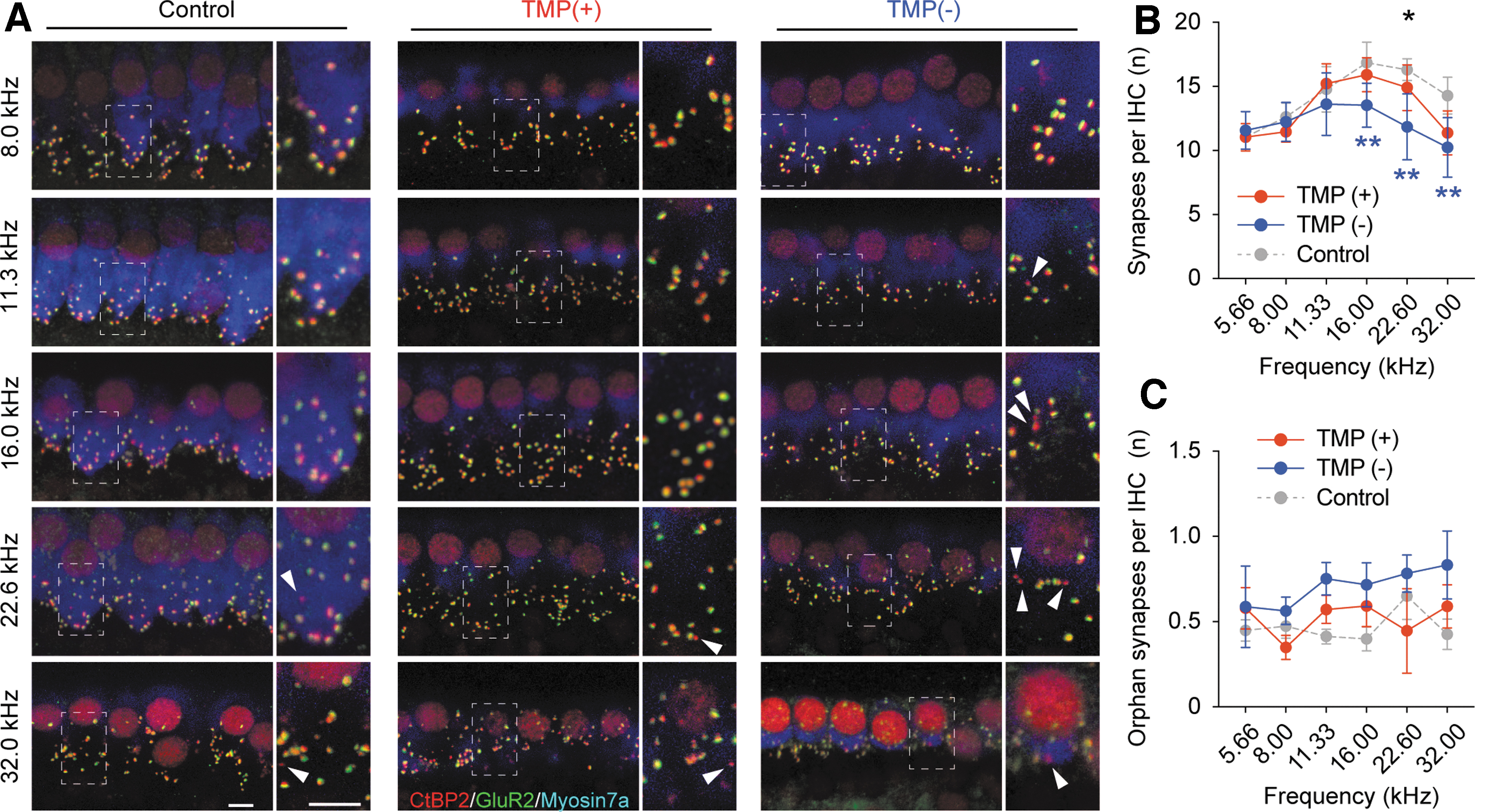
Cochlear synapses after blast exposure. (
These trends of significant differences between the TMP(−) and TMP(+) ears at 22.6 kHz were similar to the trends observed for the ABR wave I amplitude (Fig. 2F). Interestingly, however, the TMP(+) ears displayed no significant reduction in the number of synapses compared with that of the controls at all tested frequencies (TMP(+) vs. control: p = 0.054 and Cohen d = 0.19; two-way ANOVA), consistent with the results regarding ABR P1 amplitude. Similarly, the number of orphan synapses, which are pre-synaptic ribbons lacking apposed glutamate receptor patches, tended to be higher in the TMP(−) ears than in the TMP(+) ears; however, no statistically significant differences were observed among the groups at all tested frequencies (Cohen d = 0.43; two-way ANOVA followed by the Tukey post hoc test; Fig. 3C).
Stereocilia of HCs
Next, we observed the stereocilia bundles of HCs, 13 because blast exposure leads to the stereociliary disruption of HCs and it could be one of the causes for the elevation of hearing thresholds. 13 Deformed stereocilia in OHCs, such as aggregated, bent, or tangled stereocilia, were observed in blast-exposed cochleae (Fig. 4A,B). In contrast, no obvious stereociliary disruption was observed in IHCs after blast exposure.

Surface structures in the hair cells (HCs) after blast exposure. (
We further performed a quantitative assessment of the stereociliary bundle disruption in the OHCs for each group (Fig. 4C). The stereociliary disruption ratio was significantly increased in the blast-exposed TMP(−) ears compared with that in the controls (TMP(−) vs. control: p = 0.04 at 22.6 kHz and Cohen d = 0.30; two-way ANOVA followed by the Tukey post hoc test). There was no significant difference, however, between the TMP(+) and TMP(−) ears at all tested frequencies (p = 0.73 and Cohen d = 0.09; two-way ANOVA followed by the Tukey post hoc test). These results indicate that stereociliary disruption in OHCs could be the major cause for the significant elevation of ABR and DPOAE thresholds observed after blast exposure.
Properties of ANs
The observation that ABR P1 latency was longer after blast exposure suggested that the property of the ANs might be disrupted. Consequently, we evaluated the axon caliber and myelination of the ANs (Supplementary Fig. S4A). No significant difference was observed in the axon caliber and g-ratio among groups (axon caliber: p = 0.58 and Cohen d = 0.14 [Kruskal–Wallis test]; g-ratio: F(2,294) = 1.51 and p = 0.22; Supplementary Fig. S4B).
A previous study reported that the ANs showed a reduction in the density and nodal length of the nodes of Ranvier in noise-induced cochlear damage. 24 Therefore, to investigate the density and the length of the nodes of Ranvier, cochlear pieces were immunostained with the nodal protein NrCAM, which stained the nodes and unmyelinated axons, in combination with the paranodal protein Caspr (Supplementary Fig. S4C). Although the immunostaining for NrCAM and Caspr indicated the presence of dysmyelinated AN fibers after blast exposure, the quantification of the nodal density and length revealed no significant differences after blast exposure (nodal density: F(2,15) = 0.04, p = 0.96, and Cohen d = 0.14 [one-way ANOVA]; nodal length: p = 0.76 and Cohen d = 0.03 [Kruskal–Wallis test]; Supplementary Fig. S4D).
Synaptic assessment of cochlear nucleus
The auditory system compensates for peripheral auditory loss through increased central neural activity, including in the CN where all AN axons project. This alternative gain in the central auditory activity in response to the reduction in auditory inputs is referred to as “central compensation” and is associated with abnormal auditory perception. 25,26
To investigate the CN changes in response to blast-induced cochlear damage, we measured the amplitude of ABR wave 2, generated by the CN activity. The relative ABR amplitude from peak 2 to peak 1 (P2/1) tended to be lower in the TMP(+) and TMP(−) ears; a statistically significant difference was observed among the groups (p = 0.006, Cohen d = 0.49; two-way ANOVA followed by the Tukey post hoc test; Fig. 5A). No significant difference was observed between TMP(+) and TMP(−) ears (p = 0.86, Cohen d = 0.08; two-way ANOVA followed by the Tukey post hoc test).

Vesicular glutamate transporter-1 (VGLUT-1) and glutamic acid decarboxylase (GAD65) expression in the ventral cochlear nucleus and quantitative analyses of puncta density. (
Further, the ABR latency from peak 1 to peak 2 (P1-2) was significantly longer in the TMP(+) and TMP(−) ears than in the control ears (TMP(+) vs. control: p < 0.01 at 16.0 kHz and Cohen d = 0.54; TMP(−) vs. control: p = 0.02 at 11.3 kHz and 16.0 kHz and Cohen d = 0.66; two-way ANOVA followed by the Tukey post hoc test). No significant differences were exhibited in the ABR latency P1-2 between TMP(+) and TMP(−) ears (p = 0.99, Cohen d = 0.09; two-way ANOVA followed by the Tukey post hoc test).
Next, we evaluated the expression of vesicular glutamate transporter-1 (VGLUT1) and glutamic acid decarboxylase (GAD65) in the ventral CN (VCN) to identify the effects of blast exposure on excitatory and inhibitory synapses, respectively (Fig. 5B). The auditory nerve fibers with synapses onto the CN are labeled using VGLUT-1. 27,28 Especially, the characteristic rings of VGLUT1 puncta in VCN represent auditory-nerve endbulbs of Held around the spherical and globular bushy cells. 29
The VGLUT-1 puncta density was significantly lower in the TMP(+) and TMP(−) ears than in the control ears (TMP(+) vs. control: p < 0.01 and Cohen d = 1.33; TMP(−) vs. control: p < 0.01 and Cohen d = 0.91; one-way ANOVA followed by the Tukey post hoc test). No significant difference in VGLUT-1 expression was observed, however, between the TMP(+) and TMP(−) ears (p = 0.83, Cohen d = 0.42; one-way ANOVA followed by the Tukey post hoc test).
Further, quantitative analysis of GAD65 puncta and neural density in the VCN revealed no significant differences among the groups (GAD65: F(2,12) = 0.95, p = 0.42, and Cohen d = 0.59; neural density: F(2,12) = 0.34, p = 0.72, and Cohen d = 0.23; one-way ANOVA), suggesting that damage to the central auditory synapses was not correlated with the occurrence of TMP.
Discussion
Taken together, our results show that although blast-exposed mice had both peripheral and central auditory damage, no significant pathophysiological differences were observed according to the occurrence or absence of TMP, except for TM thickness, DPOAE thresholds, ABR P1 amplitude, and the number of cochlear synapses. Repaired TM remained significantly thickened at least two months after blast exposure. This conductive pathology could be a major cause of the elevation of the DPOAE thresholds in the TMP(+) ears, although the elevation of the ABR thresholds and the disruption of OHC stereocilia were comparable in the TMP(+) and TMP(−) ears. Further, a significant reduction in the ABR P1 amplitude and the number of cochlear synapses without HC and SGN loss were observed in the TMP(−) ears.
These results indicated the occurrence of blast-induced TMP was correlated with peripheral cochlear synaptopathy, which was paired with a lesser functional decline than that found in the absence of TMP. Other end-points, such as hearing thresholds and mechanical damages in OHC stereociliary disruption, were largely not associated with the occurrence of TMP.
The observed stereociliary disruption is a unique characteristic of BAD pathology that is consistent with a previous report showing similar blast-induced cochlear degeneration without TMP. 12 Importantly, our data revealed that significant cochlear synaptopathy was not observed in the TMP(+) ears but was seen in the TMP(−) ears (Fig. 3). Consistent with our results, another study using chinchillas exposed to multiple blasts showed that ears with TMP displayed less cochlear damage than ears with intact TM. 7 Overall, blast-induced TMP might attenuate IHC cochlear synaptopathy by reducing the power of the blast shock wave passing through the TM, but may not mitigate the OHC stereociliary disruption.
One possible explanation for our results is that the blast shock wave transmitted mainly through bone conduction, which is not affected by the presence of TMP, caused OHC stereociliary disruption. Consistent with this hypothesis, we previously reported that shock waves radiated to the temporal bone can lead to OHC stereociliary disruption through bone conduction. 12 Clinical studies in humans reported no significant difference in audiometric thresholds in blast-exposed ears with and without TMP, 30 consistent with our ABR results; the degeneration of cochlear synapses precedes both HC loss and hearing threshold elevation. 31,32
Further, in terms of the cochlear pathology, the number of cochlear synapses at 22.6 kHz was significantly reduced in the TMP(−) ears compared with the number in the TMP(+) ears; this finding was consistent with the findings of the ABR wave I amplitudes. Overall, cochlear synaptopathy tended to be severe and ABR latency tended to be prolonged in the high-frequency region.
We believe that the pathophysiological differences in the auditory neurons between the TMP(+) and TMP(−) ears were mainly observed in the high frequency region because the base of the cochlea (where a high frequency sound transduces) was more vulnerable compared with the apex of the cochlea (where a low frequency sound transduces). The reason is that the shock wave traveled from the base to the apex of the cochlea, thereby exposing the cochlear base to the strong impact of the blast shock wave through the TM. The TMP(+) ears, however, showed a significant decrease in the DPOAE thresholds without significant differences in the ABR thresholds, indicating that the TMP(+) ears have a conductive pathology, which has a greater impact on the DPOAE than the ABR thresholds. 19,20
Although we observed no obvious dislocation or fracture in the ossicular chain (Supplementary Fig. 1), the thickened TM might be one of the causes for the observed elevation in the DPOAE thresholds, as similarly reported in human patients. 33
Despite a reduced cochlear output after ototoxic insults, central neural activity in the CN is often paradoxically enhanced, which is termed “central compensation,” 25,26,34 and is hypothesized to be a potential mechanism involved in abnormal auditory perceptions, such as tinnitus. 26 Currently, little is known about central compensation after blast exposure and whether central auditory deficits are affected by TMP or blast-induced cochlear synaptopathy.
In our study, we demonstrated that relative CN activity measured by the ABR amplitude P2/1 was significantly decreased two months after blast exposure (Fig. 5A) regardless of the occurrence of TMP. We further observed the imbalance between the excitatory and inhibitory synaptic density in the CN of blast-exposed mice, consistent with previous studies after cochlear damage. 22,27,35 Our results suggest that central neural activities in the CN are not compensatively enhanced after blast exposure, even though the blast shock waves cause cochlear synaptic disruption and reduction of cochlear output. The possible reasons why central compensation was not observed are that minimal peripheral pathology such as cochlear synaptopathy without apparent loss of HCs and SGNs might be inadequate to induce central compensation, as observed in acoustic trauma. 34
Further, the auditory system is more vulnerable to blasts than to only noise, which predominantly leads to cochlear damage only. 36 Otherwise, central auditory damage might be independent of peripheral auditory damage because of the effects of the direct transmission of blast shock wave in the central auditory systems. 37,38 Based on this hypothesis, it is reasonable that the blast can cause auditory impairment even if hearing and cochlear function is normal.
The clinical data from blast-exposed humans suggests that blast exposure leads to defects in hearing in complex auditory environments, known as “hidden hearing loss.” 39 Moreover, the imbalance between the excitatory and inhibitory synapses may lead to impaired neurotransmission, which could ultimately contribute to the generation of abnormal auditory perception, such as tinnitus or hyperacusis. 40
There are several limitations to our study. First, there are potential confounding factors, because we speculated that TMP could buffer the harmful blast overpressure transmitted to the cochlea based on the pathophysiological differences between the TMP(+) and TMP(−) ears, but we did not directly measure the power of the blast shock wave delivered to the cochleae through the TM. Second, we were unable to assess whether the central synaptic imbalances observed in this study correlated with BAD in humans. Because behavioral measurements for tinnitus and hyperacusis were not performed in the blast-exposed animals, the detailed mechanisms of blast-induced BAD also remain unclear.
Finally, although peripheral cochlear damage was reported to lead usually to secondary central degeneration in the auditory pathway, 4 our findings revealed that central auditory damage might be independent of peripheral auditory damage. This indicates that direct transmission of the blast shock wave could affect the central auditory systems.
Further studies will be necessary to elucidate, independently, the mechanisms of central degeneration arising from the following: (1) the primary influence of the shock wave to the brainstem via bone conduction and (2) the secondary influence of cochlear deterioration. In addition, further studies are warranted to investigate the effect of blast exposure on central neural circuits, including the entire auditory pathway, and to elucidate the pathology of BAD. 42,43
Taken together, our findings have significant clinical implications for the understanding and management of human BAD. Although blast-induced cochlear degeneration may be influenced by the distance from the blast, physical orientation, and cochlear vulnerability, our findings of cochlear synaptopathy may shed light on the mechanisms of BAD, including abnormal auditory perception in the military population. Based on our results, TMP seems to have a protective effect, and TM resilience may be a key determinant of BAD severity. Further clinical studies will be necessary to elucidate the detailed effects of TMP on BAD in humans.
Conclusions
Blast-exposed mice exhibited both peripheral and central BAD, regardless of the presence or absence of TMP. Although central changes were not affected by the presence of TMP, the TMP(−) ears exhibited significantly more cochlear synaptopathy than the TMP(+) ears. Our study has important clinical implications, suggesting that TMP could buffer against peripheral cochlear synaptopathy without having a similar effect on the central auditory synapses.
Experimental Protocol
Experimental protocols in this study were approved by the Institutional Animal Care and Use Committee (#18050) and conducted in accordance with the relevant regulations and guidelines.
Footnotes
Acknowledgments
We wish to thank Yayoi Ichiki for providing research assistance.
Authors' Contributions
T.K. and K.M. designed the experiments. T.K., K.M., Y.S., and A.S. performed the experiments and analyzed the data. T.K. and K.M. wrote the manuscript. All authors reviewed the manuscript.
Funding Information
This work was supported by a grant for Advanced Defense Medical Research provided by the Japanese Ministry of Defense (Grant Number A-4, A.S.) and two grants from JSPS KAKENHI (Grant Numbers 21K09573 [K.M.] and 20K18263 [T.K.]).
Author Disclosure Statement
The authors declare that they have no conflict of interest.
Supplementary Material
Supplementary Experimental Procedures
Supplementary Table S1
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.