
Abstract
Immediately following spinal cord injury (SCI) patients experience pain associated with injury to the spinal cord and nerves as well as with accompanying peripheral injuries. This pain is usually treated with opioids, and most commonly with morphine. However, in a rodent model we have shown that, irrespective of the route of administration, morphine administered in the acute phase of SCI undermines long-term locomotor recovery. Our previous data suggest that activation of kappa opioid receptors (KORs) mediates these negative effects. Blocking KORs with norbinaltorphimine (norBNI), prior to a single dose of epidural morphine, prevented the morphine-induced attenuation of locomotor recovery. Because numerous cellular changes occur with chronic opioid administration compared with a single dose, the current study tested whether norBNI was also effective in a more clinically relevant paradigm of repeated, intravenous morphine administration after SCI. We hypothesized that blocking KOR activation during repeated, intravenous morphine administration would also protect recovery. Supporting this hypothesis, we found that blocking KOR activation in young, male rats prevented the negative effects of morphine on locomotor recovery, although neither norBNI nor morphine had an effect on long-term pain at the doses used. We also found that norBNI treatment blocked the adverse effects of morphine on lesion size. These data suggest that a KOR antagonist given in conjunction with morphine may provide a clinical strategy for effective analgesia without compromising locomotor recovery after SCI.
Introduction
Immediately after spinal cord injury (SCI), people often experience severe pain, resulting from damage to the spinal nerves, tissue, vertebrae, and accompanying peripheral injuries. Clinicians commonly prescribe opioids to manage this pain. In fact, in a retrospective analyses of the effects of opioids on pain and recovery in the clinical population we found that every SCI patient included in the study, received at least one dose of opioids in the first 24 h after injury, with morphine administered 80% of the time. 1
Unfortunately, however, in both our clinical and pre-clinical studies with rats we have found that early opioid administration undermines recovery after SCI. In the rat SCI model, we have repeatedly shown that morphine treatment in the acute phase of injury increases cell death at the lesion site, increases the development of chronic pain, reduces locomotor recovery, and decreases weight gain. 2 –5 Similarly, in the clinical population, we found a positive correlation between the amount of opioids prescribed in the acute phase of SCI and pain ratings 1 year post-injury. 1 Given these adverse secondary consequences, caution is warranted when using opioids to treat pain after SCI. However, as patients vary significantly in their responses to pain medications, clinicians need an array of analgesic options. Indeed, untreated pain following SCI has also been linked to reduced locomotor recovery, increased chronic pain, and cognitive deficits. 6 –11 Rather than removing opioids from SCI management strategies, it is critical that we identify the mechanism underlying the negative effects of morphine on recovery after SCI.
Our previous data suggest that activation of classic opioid receptors may play a role in the morphine-induced attenuation of recovery. Although morphine binds primarily to mu opioid receptors (MORs), it also binds to other classic opioid receptors, including kappa and delta opioid receptors (KORs and DORs). 12,13 In a previous study, we found that selective activation of KORs, but not MORs or DORs, reduced locomotor recovery and increased the incidence of pain in a rodent model of SCI, outcomes that are also seen with epidural morphine administration. 14 Building on this study, Aceves and coworkers tested whether a KOR antagonist, norbinaltorphimine (norBNI), could prevent the negative outcomes of morphine on recovery after injury. 15 They found that norBNI blocked the adverse effects of a single dose of epidural morphine.
Although single dose, epidural studies provide insight into the spinal mechanisms of how morphine effects recovery, intravenous infusion is the canonical route of administration of morphine in the clinic. Moreover, in a clinical setting, opioids are administered repeatedly. Prior literature indicates that numerous changes occur with repeated opioid administration, which can lead to both opioid tolerance and opioid-induced hyperalgesia (OIH). 16,17 Tolerance occurs when there is a decrease in the pharmacological response following repeated drug administration, whereas OIH is the paradoxical increased sensitivity to nociceptive stimuli that occurs with repeated opioid exposure. Although the exact mechanism behind these outcomes remains to elucidated, the development of tolerance and OIH highlight the potential differences between the effects of single and the effects of repeated opioid administration, and underscore the importance of investigating potential therapeutics in a repeated, intravenous administration paradigm.
To address this, the current experiment investigated whether the KOR antagonist, norBNI, was sufficient to block the adverse effects of repeated, intravenous morphine in a rodent model of SCI. We hypothesized that antagonizing KORs with norBNI would be sufficient to protect against the negative effects of repeated, intravenous morphine. Assessing pain, locomotor recovery, tissue sparing, and general health after SCI, we found that blocking KOR receptors during repeated morphine administration did effectively protect locomotor recovery.
Methods
Subjects
Thirty-eight male Sprague–Dawley rats (Envigo, Houston, TX, USA) ∼90–110 days old (300–350 g) served as subjects. Seven rats were subsequently removed from the study based on a priori exclusion criteria. Subjects were excluded from the study if their (1) average Day 1 Basso, Beattie and Bresnahan (BBB) score was >6.0, and (2) the jugular catheter was not patent for all 7 days of intravenous administration. Data for one rat treated with norBNI only was not completely recorded, and therefore was removed during analysis, and five rats died prior to the end of the experiment. Mortality was distributed across groups; one rat from the vehicle group, three treated with only morphine, and one treated with only norBNI. In total, therefore, 31 subjects completed the study (n = 8 vehicle controls, 8 morphine only, 6 norBNI only, and 9 norBNI + morphine).
The rats were individually housed in Plexiglas bins (45.7 [length] × 23.5 [width] × 20.3 [height] cm), with a fixed 12-h light/dark cycle, and access to both food and water ad libitum. All testing and surgeries occurred during the light cycle. Following the contusion injury, subjects' bladders were expressed manually every morning (between 7:00 and 9:00 a.m.) and evening (between 4:30 and 6:30 p.m). This schedule was maintained until subjects regained bladder function (operationally defined as voiding on their own for 3 consecutive days). All of the experiments reported here were reviewed and approved by the Institutional Animal Care Committee at Texas A&M University, and were consistent with the National Institutes of Health (NIH) guidelines for animal care and use. The experimental design is outlined graphically in Figure 1 and described in further detail in the following sections.

Graphical representation of experimental timeline.
Intravenous jugular catheter implantation
Five days prior to a contusion injury, rats were implanted with a jugular catheter. They were anesthetized with isoflurane anesthesia (2–3% gas), and their back and chest were shaved and disinfected with alcohol and iodine. Next, a 3 cm horizontal incision was made below the shoulders, and a 1 cm superficial incision was made above the jugular vein. The jugular vein was isolated from the surrounding connective tissue. Placing the rat on its right side, a subcutaneous tunnel beginning at the incision on the back, traveling over the shoulder and ending at the neck, was made by carefully separating the skin from the muscle. A catheter, consisting of Silastic tubing (0.025-mm ID), was passed through this tunnel. The catheter was attached to a syringe and flushed with sterile saline. Then using an 18-G needle as a guide, the Silastic tubing was inserted into the vein and secured with sutures and Vetbond. Successful insertion of the catheter was tested by drawing blood back into the tubing. Once placement was confirmed, a back mount pedestal (model 313-00BM-10-SPC; Plastics Inc., Roanoke, VA, USA) was connected to the catheter and implanted subcutaneously. All incisions were then closed with Michel clips. Finally, the catheter was flushed with 150 uL of heparin-saline (1000 units/mL), to prevent blood from clotting inside the tubing, and capped. Subjects were also given 3 mL of saline subcutaneously to compensate for fluid loss. The rats were kept in a recovery room maintained at 26.6°C for the first 12–16 h following catheter implantation.
Spinal contusion injury
Five days after the implantation of the jugular catheter, the rats underwent a moderate contusion injury using the Infinite Horizon (IH) spinal cord impactor (PSI, Fairfax Station, VA, USA) as previously described. 18 Briefly, subjects were anesthetized using isoflurane (2–3% gas). Once a stable level of anesthesia was reached, the dorsal spinous process at T12 was removed and the spinal cord was exposed. The vertebral column was fixed within the IH device using two Adson forceps, and a moderate, midline contusion injury was produced by applying an impact force (150 kdyn with a 1 sec dwell time) onto the exposed spinal cord. Following injury, the rats were removed from the impactor device and the incision was closed with Michel clips. To compensate for fluid loss, subjects were given 3 mL of saline subcutaneously after surgery. The rats were kept in a recovery room maintained at 26.6°C for the first 24 h post-injury.
Epidural catheter implantation
All rats were implanted with an epidural catheter immediately following the contusion injury and prior to closing the incision site. This 15 cm catheter consisted of polyethylene tubing (model 51150, Stoelting, Co., Wood Dale, IL, USA) and a silicone ball positioned 4 cm from the end of the tubing. The catheter was inserted into the epidural space, with the end positioned 2 cm caudal to the spinal injury. To prevent cannula movement, Vetbond was used to adhere the silicone ball to the subcutaneous connective tissue above the exposed spinal cord. The skin incision above the laminectomy was then closed with Michel clips. The externally exposed end of the catheter was further secured within the wound clips. The end of the exposed catheter was also sealed with heat to prevent any contaminates from being introduced directly to the spinal cord. The catheter was removed on Day 7 post-injury, following the end of post-administration pain testing, to prevent any confounding effects of inflammation with chronic implantation. 19 With minimal pressure, the catheter slowly slid out of the epidural space.
Drug administration
Twenty-four hours following the contusion injury, half of the rats received intravenous morphine (10 mg/mL) on an escalating dose schedule, and the other half received an equivalent volume of intravenous saline (0.9%). On Days 1–2, the subjects were treated with 10 mg of morphine, on Days 3–4 they were treated with 20 mg, and on days 5–7 they were treated with 30 mg. Morphine was administered in 5 mg doses and doses were separated by 1 h up to the daily dose, to prevent overdose. This dosing schedule was derived from previous experiments assessing analgesia, self-administration, and cell expression following SCI. 5,20 Heparin-saline (1000 U/mL) was used to flush catheters following drug administration.
Prior to the first administration of morphine on Day 1, subjects were also pre-treated with 0.32 μmol (dissolved for a final volume of 8 μL in ultra-purified water) of norBNI via the epidural catheter, or the equivalent volume of 0.9% saline. NorBNI was administered 2 h prior to morphine administration, to account for the slow onset of its antagonistic action at the KOR. 21 –23 Following administration of norBNI, catheters were flushed with 10 μL saline. Subjects were re-treated with norBNI (0.32 μmol) or saline on Day 3 post-injury, prior to the initiation of the daily morphine administrations, to prevent morphine-induced KOR activation during the remaining days of administration. 24 –26
Assessment of sensory reactivity
Thermal reactivity
Thermal reactivity was assessed with the tail-flick test immediately before and 30 min after final drug treatment on Days 1, 3, and 7 post-injury. Rats were placed in restraining tubes (6 cm in diameter, 23 cm in length) and acclimated to the tail flick apparatus for 15 min (IITC Life Science Inc., Woodland Hills, CA, USA). Prior to testing, the intensity of the light was set to elicit a baseline tail-flick response in ∼3–4 sec in an intact rat. For test trials, a hot light of the same temperature was focused onto the rat's tail and the latency to flick the tail away from the stimulus was recorded. If a subject failed to respond, the test trial was automatically terminated after 8 sec of heat exposure. Two tests were conducted at 2 min intervals, and the second tail-flick latency was recorded. This test was used to assess both OIH (immediately before) and tolerance (30 min following) after repeated drug administration, on Days 1, 3, and 7 post-injury. The tail-flick test was also used to assess long-term sensory reactivity thresholds on days 14, 28, and 42 post-injury.
Tactile reactivity
Mechanical reactivity was measured using von Frey filaments (Semmes-Weinstein Aesthesiometer, Stoelting Co., Chicago, IL, USA) prior to and 30 min after final drug administration on Days 1, 3, and 7 post-injury. Mechanical reactivity was also assessed on Days 14, 28, and 42 to test for the development of long-term pain. Filaments of increasing diameter (correlating with the target force delivered) were applied to the L4 dermatome on the plantar surface of the hind paw for 2 sec. The force applied was increased until the rats exhibited both a motor (hind paw withdrawal) and vocal response. 27 The force (g) of the stimulus that produced a response was calculated using the formula provided by Semmes-Weinstein: Intensity = log10 (10,000g force). If a motor and/or vocal response was not observed, testing was terminated at a force of 300g.
Von Frey filaments were also used to assess at-level pain with the girdle test on Days 14, 28, and 42 post-injury. In this test, the filament corresponding to 26g of force was applied to 44 points across the subjects' back in a grid-like pattern. As rats do not normally vocalize in response to this stimulus, vocalizations indicated that rats experienced a noxious event. The number of vocalizations was recorded and normalized by the following formula: (Nv × 100)/total number of applications.
Assessment of locomotor recovery
Locomotor recovery was assessed using the BBB scale. 28,29 The BBB scale is locomotor rating scale used to characterize the hindlimb function of rats following SCI. This scale rates movement of the three major hindlimb joints (hip, knee, and ankle), plantar stepping, and coordination between paws. BBB scores were collected daily for the first 7 days post-injury, then every other day from Day 9 to Day15, every third day from Days 18 to Day 30, and every fourth day from Day 34 to Day 42. For this assessment, rats were placed in an open enclosure (99 cm in diameter, 23 cm deep) and observed for 4 min. Investigators were blind to a subject's experimental treatment, and care was taken to ensure that all investigators had high intra- and inter- observer reliability (all rs > 0.89).
Motor function was also assessed on Day 42 post-injury using the beam and ladder walk tests. 29 Prior to testing, subjects were habituated to both the beam and ladder apparatus for 3 consecutive days (8 min per habituation). The tapered beam walk task provides an index of postural stability, and a gross measure of paw placement. The tapered beam is 17.14 cm wide at the beginning and 0.95 cm wide at the end. On this test, the width at which each foot failed to plantar place on the beam was recorded, and the scores for the left and right legs were averaged to provide a score for beam walk performance. The ladder task requires subjects to cross a horizontal ladder (20 cm wide; 37 rungs at 2.5 cm intervals), providing a measure of the fine motor abilities of the hind paws. Using post hoc frame-by-frame video analysis, the number of times that subjects did not successfully place their hind paws on a rung (their paw slipped between rungs) was recorded.
Lesion Analysis
On Day 43 post-injury, subjects were deeply anesthetized (100 mg/kg of Beuthanasia, i.p.) and perfused intracardially with phosphate-buffered saline followed by 4% paraformaldehyde. Following perfusion, a 1.5 cm spinal segment around the lesion site was extracted and prepared for cryostat sectioning. Tissue was embedded in optimal cutting temperature compound. Then, during sectioning, every 10th transverse slice (20 μm width) was collected and mounted directly onto Superfrost Plus slides. The mounted tissue was stained with cresyl violet for Nissl substance and Luxol fast blue for myelin. 28 Sections ±0, 600, 1200, 1800, and 2400 μm rostral and caudal from the lesion center were analyzed using Stereo Investigator software (MFB Bioscience, Williston, VT, USA). Three indices of lesion magnitude were derived for each section: lesion, residual gray matter, and residual white matter. The lesion was defined by identifying and tracing around the areas of cystic formations. Residual gray matter was identified by Nissl-stained areas containing neurons and glia. Areas lacking gliosis denoted spared white matter. 28 The total area of each cross section was determined by summing the lesion, residual gray matter, and residual white matter.
A correction factor was used to control for variability in the section area across subjects. This correction factor was derived from age-matched, undamaged cord sections. 30 Standardizing the area across sections allowed us to estimate the degree to which tissue was “missing” (i.e., atrophy, necrosis, or apoptosis). An accurate assessment of the degree to which a treatment has impacted the spinal cord includes both the remaining “damaged” tissue, and resolved lesioned areas. Summing the amount of “missing” tissue and the measured “damaged” area allowed us to derive an index of the relative lesion percent relative lesion) in each section that was comparable across sections. We also computed the relative percent of gray and white matter remaining in each section, relative to intact controls. Previously, using this method, we have found a high correlation between relative lesion and measures of behavioral recovery including BBB locomotor scores and recovery of bladder function. 30,31
Statistical analysis
Three-way repeated measures analysis of variance (ANOVA), with day post-injury, morphine, and norBNI treatment as independent variables, were used to analyze the sensory reactivity data. When analyses revealed significant main effects or interactions (p < 0.05), Tukey's multiple comparison tests were used for post-hoc analysis.
The effects of morphine and norBNI on locomotor recovery across days were assessed with an analysis of covariance (ANCOVA). Day 1 BBB scores collected prior to drug treatment were used as the covariates. Use of the Day 1 covariate considerably reduces unexplained variance between groups and, as a result, increases statistical power. Additional tests of motor function, including data collected for the beam and ladder walk tasks, were analyzed with two-way (morphine and norBNI) ANOVAs. Finally, two- and three-way ANOVAs with independent variables of distance from the center of the lesion, morphine, and norBNI treatment were used to analyze data for lesion size and tissue sparing. Tukey's multiple comparison tests were used for post-hoc analyses.
Results
An escalating dose of intravenous morphine prevented the development of opioid-induced tolerance
Intravenous morphine produced robust analgesia following SCI. A three-way repeated measures ANOVA revealed a main effect of morphine on tail-flick latency (F [1, 27] = 213.1, p < 0.0001, Fig. 2A) regardless of norBNI administration. There was no main effect of norBNI (F [1, 27] = 1.26, p > 0.05), day (F [2, 54] < 1.0, p > 0.05) across testing days and no interactions between morphine or norBNI and day of administration (F [1, 27] = 1.28, p > 0.05). The analgesic effects of morphine persisted across all 7 days and norBNI had no effects on analgesic efficacy.
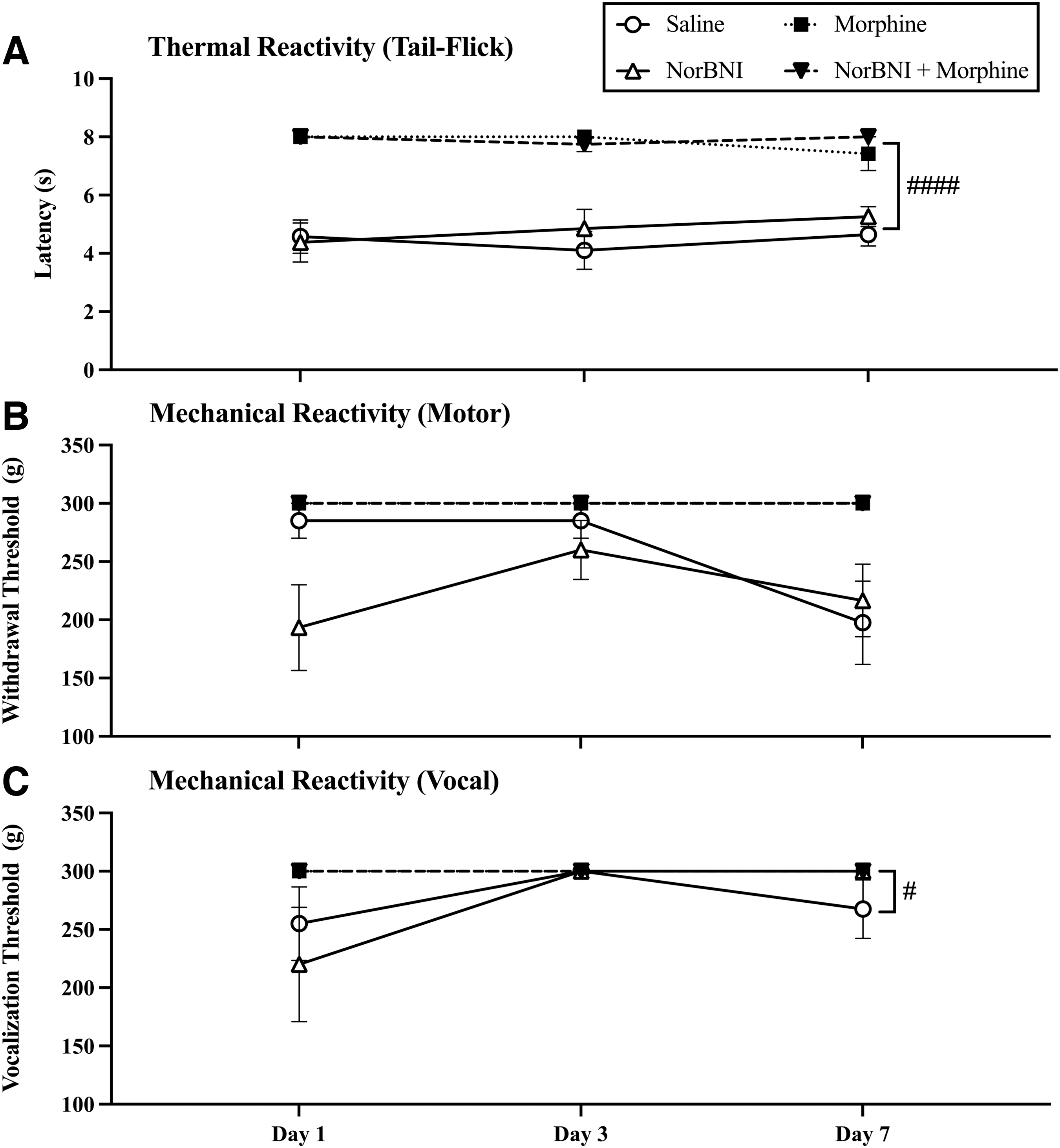
Acute pain testing, after the final dose of morphine on Days 1, 3, and 7 post-injury, revealed that the escalating dose of morphine prevented the development of opioid-induced tolerance. Morphine treatment resulted in maximum pain reactivity thresholds (i.e., no reactivity to stimulation) in (
Similarly, there was a main effect of morphine on mechanical reactivity for both motor (F [1, 27] = 34.06, p < 0.0001) and vocal responses (F [1, 28] = 5.08, p = 0.03) across days. Overall, morphine reduced motor and vocal responses to mechanical stimulation. There was also an interaction between day of administration and morphine administration for both motor (F [1, 27] = 3.65, p = 0.03) and vocal (F [1, 27] = 6.12, p = 0.004) reactivity. Morphine produced robust analgesia across the first 7 days of administration, whereas reactivity thresholds decreased in the saline-treated groups across days. NorBNI did not affect either motor (F [1, 27] = 2.46, p > 0.05) or vocal (F [1, 27] < 1.0, p > 0.05) reactivity with mechanical stimulation (Fig. 2B and C). Overall, these data suggest that tolerance did not develop with the escalating dose of morphine across days and that norBNI does not affect the analgesic potency of morphine.
Repeated morphine administration produced OIH
Thermal and tactile reactivity were also assessed prior to daily drug administration to monitor the development of OIH. There were no significant baseline differences in thermal or mechanical reactivity across groups prior to drug administration on Day 1 (F [1, 27] ≤ 1.0 for all tests, p > 0.05). However, by the 7th day of morphine administration there was evidence of OIH in motor responses to von Frey stimulation. A three-way ANOVA revealed a main effect of morphine (F [1, 27] = 34.06, p < 0.0001) across days, a main effect of day of administration (F [2, 54] = 17.25, p < 0.0001), and an interaction between day of administration and morphine (F [1, 27] = 3.65, p = 0.03, Fig. 3B). Morphine-induced hypersensitivity developed in spite of norBNI treatment, as post-hoc analyses reveal that morphine and norBNI-morphine treated subjects did not differ on day 7 of administration (p = 0.99). There was no effect of testing day, morphine, or norBNI treatment on the tail-flick assay or on the vocal response to von Frey stimulation (all Fs < 3.0, p > 0.05) (Fig. 3A and C).
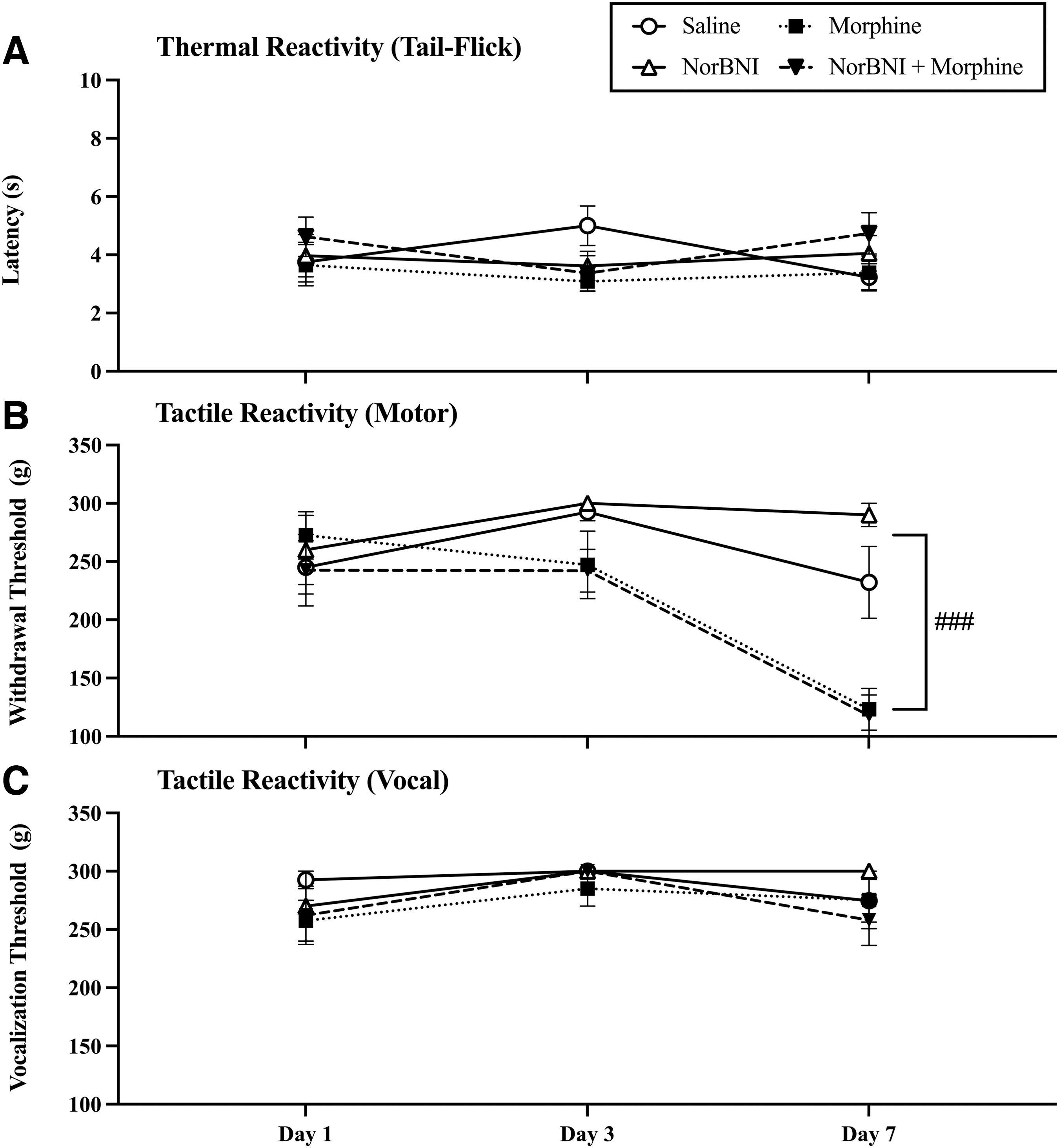
Repeated morphine administration produced opioid-induced hyperalgesia (OIH). Analysis of pain thresholds prior to drug administration on Days 1, 3, and 7 post-injury revealed that irrespective of norbinaltorphimine (norBNI) treatment, morphine significantly reduced (
Morphine increased weight loss after injury
Body weights were recorded as an indicator of general health. To control for differences in starting weights, weight loss was calculated by subtracting body weight prior to injury from the daily weight recorded post-injury. Regardless of treatment, all subjects lost weight in the first week following SCI. By 4 weeks post-injury, all subjects had regained their weight. However, there was a significant interaction between day post-injury and morphine treatment (F [18, 486] = 10.52, p < 0.0001) and a three-way interaction among morphine, norBNI, and day post-injury (F [18, 48] = 2.03, p = 0.007, Fig. 4). Subjects treated with morphine lost more weight following injury than their saline-treated counterparts from Day 7 to Day 14 post-injury. Although norBNI appeared to reduce weight loss from both SCI and morphine administration, post-hoc analysis found that the norBNI-only and norBNI-morphine groups did not differ from the saline-only or morphine-only groups, respectively.

Morphine increased weight loss following injury. Although all subjects lost weight following injury, morphine treatment exacerbated these effects and prolonged the time taken for subjects to return to their pre-injury weight. However, norbinaltorphimine (norBNI) improved weight gain in morphine treated subjects Results shown as mean ± standard error of the mean (SEM).
NorBNI pre-treatment prevented the morphine-induced attenuation of recovery
BBB scores did not differ across groups on Day 1 post-injury (F [1, 27] < 1.0, p > 0.05). Scores ranged from 1.75 ± 0.574 (mean ± standard error of the mean [SEM]) for rats treated with norBNI only to 3.06 ± 0.76 for rats treated with saline. Replicating previous studies, a mixed-model ANCOVA revealed that 7 days of morphine administration significantly undermined locomotor recovery (F [1, 26] = 7.11, p = 0.013) across the all testing days. There was no main effect of norBNI (F [1, 26] = 1.27, p > 0.05) or any interaction between day and drug treatment group (F [1, 27] < 1.0, p > 0.05). Repeated measures ANOVAs, comparing differences between select groups, revealed that subjects treated with morphine recovered significantly less than saline-treated subjects (F [1, 13] = 6.191, p = 0.027, Fig. 5A). Subjects that were treated with both norBNI and morphine, however, did not differ from vehicle-treated controls. No other groups differed significantly from one another (Fs < 3.0, p > 0.05).
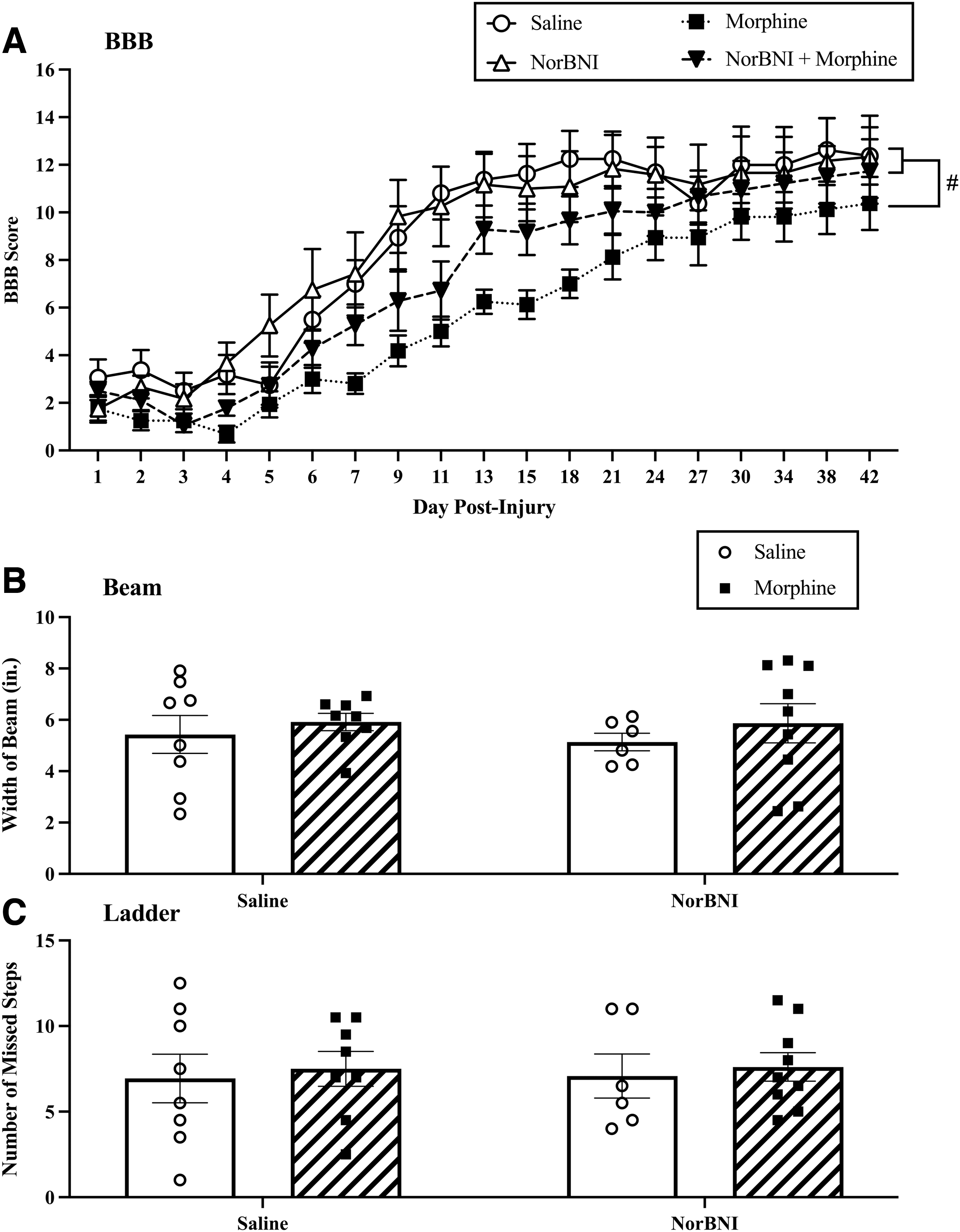
Norbinaltorphimine (norBNI) prevented the negative effects of morphine on locomotor recovery. (
Motor recovery was further evaluated on Day 42 post-injury using the beam and ladder tests. There was no significant effect of morphine on beam (F [1, 27] < 1.0, p > 0.05) or ladder performance (F [1, 2] < 1.0, p > 0.05). Similarly, norBNI did not affect beam (F [1, 27] < 1.0, p > 0.05) or ladder (F (1, 27) < 1.0, p > 0.05) scores, and there was no interaction between morphine and norBNI on either task (all Fs [1, 2] < 1.0, p > 0.05, Fig. 5B and C).
Morphine administration transiently reduced pain thresholds
To examine the effects of treatment on chronic pain, thermal and mechanical reactivity were re-assessed on Days 14, 28, and 42 post-injury. A three-way repeated measures ANOVA revealed no main effect of morphine on thermal reactivity (F [1, 2] < 1.0, p > 0.05) across days. However, there was a significant interaction between day post-injury and morphine (F [1, 27] = 3.97, p = 0.02, Fig. 6A) in the tail-flick assay. Using a two-way ANOVA to more closely investigate the effects of morphine on pain thresholds, we found a main effect of morphine on tail-flick latencies on Day 14 (F [1, 27] = 4.45, p = 0.04). However, norBNI did not rescue this effect, as post-hoc analysis found that morphine-only and norBNI-morphine treated rats did not significantly differ from one another (p = 0.44). The morphine-induced reduction in pain thresholds on Day 14 was also observed in motor (F [1, 27] = 21.39, p = 0.009) and vocal (F [1, 27] = 24.23, p = 0.006) responses to von Frey testing (Fig. 6B and C), but not in girdle testing (F [1, 27] < 1.0, p > 0.05). Similarly to tail-flick responses, morphine reduced pain thresholds irrespective of norBNI treatment, as morphine only and norBNI-morphine groups did not differ for either motor (p = 0.98) or vocal (p = 0.54) response to von Frey. There were no main effects or interactions for the girdle assay across days (Fs < 3.00, ps > 0.05, Fig. 6D). In this study, therefore, morphine with or without norBNI administration transiently reduced pain thresholds until subjects recovered to a level commensurate with their saline-treated counterparts.

Morphine transiently increases pain-like behaviors. There is a temporary reduction in pain thresholds at Day 14 in morphine-treated subjects; however, no other differences between groups were observed on subsequent testing days for (
NorBNI treatment reduced morphine-induced tissue loss
Three-way ANOVAs were used to investigate the effects of morphine and norBNI on lesion size by evaluating percent damage and relative lesion. These analyses revealed a main effect of norBNI on percent damage (F [1, 26] = 6.94, p = 0.01, Fig. 7A). Rostral to lesion epicenter, epidural norBNI treatment reduced percent damage from 10.77% to 1.74% in morphine-treated rats. Similarly, epidural norBNI reduced percent damage, albeit to a lesser extent, from 4.08% to 1.93% in saline-treated subjects. Therefore, damage was decreased in subjects treated with norBNI, regardless of whether they were treated with morphine. Two-way ANOVAs were then used to investigate which area of the cord was impacted by treatment. These analyses revealed that norBNI treatment reduced tissue loss caudal to the injury site (F [1, 26] = 11.82, p = 0.002), and although not statistically significant, there was a trend for a main effect of norBNI rostral (F [1, 26] = 4.84, p = 0.053) to the lesion center. Planned comparisons revealed that subjects treated with norBNI and morphine had significantly less percent damage than those treated with morphine alone (p = 0.02). NorBNI did not affect percent damage at the injury site (F [1, 26] = 2.22, p > 0.05).
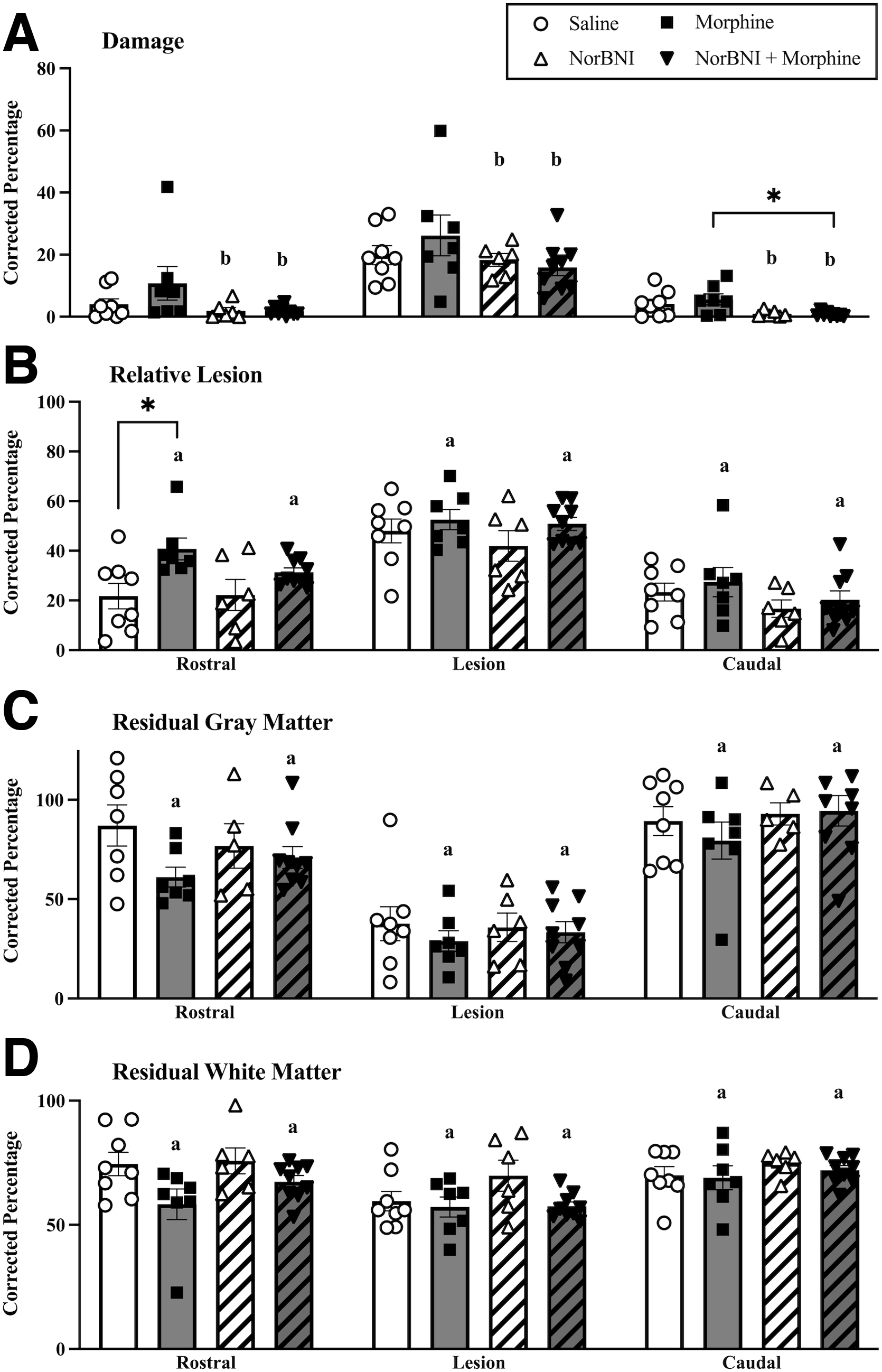
Norbinaltorphimine (norBNI) increases tissue sparing, while morphine increases tissue loss. NorBNI, regardless of morphine treatment, significantly reduced % damage (
Conversely, morphine increased relative lesion and reduced residual tissue. A main effect of morphine was observed on percent relative lesion (damage and missing tissue, F [1, 26] = 8.20, p = 0.008) (Fig. 6B), residual gray (F [1, 26] = 4.75, p = 0.038) and white matter (F [1, 2] = 7.55, p = 0.01) (Fig 6C and D). All of the effects of morphine were observed rostral to the lesion; relative lesion (F [1, 26] = 10.04, p = 0.003), residual gray matter (F [1, 26] = 4.53, p = 0.042) and white matter (F [1, 26] = 4.71, p = 0.04). Rostral to the lesion epicenter, morphine increased percent relative lesion (percent damage + percent missing) from 21.75% to 40.76%. Whereas norBNI reduced percent relative lesion for morphine-treated subjects (31.35%) but not for saline-treated subjects (22.19%) respective to their controls. Morphine also reduced gray matter from 87.08% to 61.07%, and reduced white matter from 74.51% to 58.27% relative to saline controls, rostral to the lesion center.
Discussion
Epidural norBNI treatment prevented the adverse effects of repeated, intravenous morphine administration on locomotor recovery in the rat SCI model. Rats treated with morphine alone had significantly less recovery of locomotor function than their saline-treated counterparts even 35 days after the final administration of morphine. NorBNI pre-treatment blocked these effects. NorBNI also prevented the morphine-induced increase in lesion size. There was a main effect of morphine on lesion size, but this was driven by the subjects treated with morphine alone. In fact, irrespective of morphine treatment, norBNI reduced percent damage rostral and caudal to the lesion site. These data suggest that an adjuvant that inhibits KORs during morphine administration may allow for effective pain management, without a potential reduction in the prognosis for locomotor recovery.
Prior literature has repeatedly linked KOR activation to reduced recovery following SCI. Faden and colleagues found that intrathecal administration of dynorphin peptides, but not other neuropeptides, produced motor dysfunction in the hindlimbs of intact rats. 32 –35 Interestingly, this paraparesis was attenuated by norBNI administration, suggesting that dynorphin may negatively impact SCI through KORs. 36 Further investigation revealed that the immunoreactivity of dynorphin within the lesion is correlated with the degree of injury and subsequent motor recovery following SCI. 33 The data reported here support these findings, demonstrating that KOR activation, with morphine administration, plays an important role in recovery of motor function.
By extension, the data suggest that opioids that do not engage the KOR system might produce robust analgesia in the acute phase of SCI without compromising locomotor recovery. It is well established that opioids vary in their capacity to cross the blood–brain barrier, their activity at classic and non-classic opioid receptors, and their immunosuppressive properties, each of which could lead to different molecular outcomes and potential effects on SCI recovery. 37 –39 Although most opioids used for pain management are at least partial MOR agonists, their affinity for other classic opioid receptors vary. For example, both morphine and oxycodone are known to interact directly with KORs, whereas fentanyl does not. Based on receptor binding affinities, fentanyl may be a better pain management option than morphine in the acute phase of SCI. 40 –42
Even when opioids do not directly bind to KORs, however, they can still act on these receptors by increasing dynorphin production via the extracellular signal-regulated protein kinase (ERK)1/2 pathway. ERK1/2 is the upstream kinase responsible for cyclic adenosine monophosphate (cAMP)-response element binding protein (CREB) regulation of dynorphin gene production. 43 Indeed, our laboratory has found that intravenous morphine administration following SCI increased the levels of ERK1/2 and dynorphin in microglia and macrophages isolated from the injured spinal cord (Terminel and coworkers, submitted for publication). Similarly, other publications suggest that other common clinically used opioids, including fentanyl and oxycodone, increase pro-dynorphin and dynorphin in nervous tissues. Rivat and coworkers found that subcutaneous fentanyl increased spinal dynorphin expression in rats, and that this increase was exacerbated in subjects following a plantar incision. 44 Other researchers have found that intraparietal oxycodone can increase pro-dynorphin and dynorphin A levels in numerous brain regions, including the striatum and hippocampus. 45 Based on our retrospective clinical study, we found that morphine, fentanyl, hydromorphone, and oxycodone were the most common opioids to be administered in the first 24 h following SCI. 1 With reported links between these clinically-relevant opioids and increased dynorphin levels, it is likely that these opioids would produce similar outcomes to acute morphine treatment in the context of SCI. To address this opioid-induced increase in dynorphin, buprenorphine, an unique opioid that is a partial MOR agonist and a KOR antagonist should be investigated. 46
In animal studies, buprenorphine is shown to be a safe and effective analgesic in the acute phase of SCI. In fact, buprenorphine is commonly recommended by Institutional Animal Care and Use Committees (IACUC) as a post-operative analgesic. 47 Moreover, in a SCI model, Santiago and coworkers demonstrated that 3 days of subcutaneous buprenorphine did not affect locomotor recovery. 48 Kouya and coworkers also found that buprenorphine alleviated neuropathic pain-related behaviors after SCI. 49 This is an important finding, as previous studies suggest that canonical MOR agonists fail to treat neuropathic pain in both the clinic and experimental models. 50,51 The differences in buprenorphine pharmacology, relative to more commonly used opioids such as fentanyl and morphine, may underlie the differential efficacy in neuropathic pain treatment. These data suggest that buprenorphine following SCI may not only protect, but also improve recovery; based on our data, we hypothesize that these positive outcomes are mediated by inhibiting KOR activation.
The literature also suggests that in addition to preventing the negative effects of dynorphin on locomotor recovery, KOR antagonists could protect against opioid-induced pain. Previous studies have found that supraphysiological levels of dynorphin can be pronoiceptive. 36,52 –55 Dynorphin alone can increase pain by acting upon glutamate receptors leading to excitotoxic effects in neurons and oligodendrocytes, and potentially destabilizing astrocytes. 52 However, in the present study, neither norBNI nor morphine had an effect on the development of long-term pain. At first glance, this is in contrast to our previous studies with morphine and to the literature, but we hypothesize that the effects of morphine and dynorphin on pain and locomotor function are likely dose dependent. 20,52 –57 Previously, we showed that the dose of morphine used in the current study produces robust cell death and a subsequent loss of motor function. 5 Notably, with increased cell death, sensory function would also be significantly decreased, resulting in an absence of pain-like behavior. We presume that lower doses of morphine may cause hyperexcitability of cells without significant cell death, unmasking allodynia without substantial effects on locomotor recovery. Indeed, high concentrations of dynorphins can act on N-methyl-d-aspartate (NMDA) receptors to induce excitotoxicity leading to increased motor neuron death. 58 –60 In other studies, however, NMDA activation has also been linked to neuronal sensitization and the development of pain. 15 This may suggest that the effects of NMDA activation on pain and cell death are also on a continuum, in that lower amounts of activation lead to neuronal sensitization and higher amounts lead to excitotoxicity. Supporting this hypothesis, Martin and coworkers found that higher amounts of morphine exposure reduced magnesium inhibition of NMDA receptors compared with untreated and acutely exposed brain slices. 61 Although the escalation of the dose of morphine used in present study prevented the development of tolerance across repeated administrations, this dosing schedule may also be a limitation, as increased cell death might mask effects of morphine on the development of pain.
In the present study, however, we observed OIH that was not blocked by norBNI. We hypothesize that this persistent OIH is a result of residual NMDA activation. Although norBNI has been previously shown to reduce NMDA activation, possibly contributing to the reduction in excitotoxicity and increasing subsequent tissue sparing in the current study, there are likely multiple sources of NMDA modulation. 62 For example, morphine facilitates the removal of the magnesium block from NMDA receptors via upregulation of protein kinase Cγ, leading to NMDA channel activation. 63 –66 Moreover, MOR and NMDA receptors have been previously reported to be directly associated. 67 –69 Interestingly, the effects of NMDA activation on pain may also be dose dependent, as Vuckovic and coworkers found a dose response of the NMDA antagonist, MK-801, on the expression of pain behavior in rats. 70 Therefore, norBNI may be preventing some sources of NMDA activation; however, it is likely that morphine is still able to modulate NMDA receptors via alternate mechanisms, leading to the OIH observed in the present study.
While the current study identifies potential strategies for the safe use of morphine following SCI, further investigation into the dose-response effects of morphine on recovery is necessary. Several other limitations in the present study are also noted. First, to increase clinical utility it will be critical to investigate whether systemic norBNI could also prevent the adverse effects of intravenous morphine. Repeated epidural administrations of norBNI in clinical pain management would not be feasible. The bioavailability of norBNI was also not investigated in the present study. However, norBNI has been repeatedly cited to cross the blood–brain barrier and produce long-lasting (∼ 28 days) KOR inactivation. 71 –73 Nonetheless, epidural administration may not selectively target the spinal cord and may also result in KOR inactivation in the dorsal root ganglia. Focusing on the loci of norBNIs action, it is interesting to note that the significant protective effects of norBNI were observed caudal to the spinal injury, with non-significant trends for protection at the center and rostral to the lesion center. In contrast, the effects of morphine were localized rostral to the lesion site. It is possible that administering norBNI rostral, rather than caudal, to the lesion site, would increase the protective effects of norBNI administration. Future studies could also investigate which cell types increase dynorphin production during morphine administration to develop targeted cell-specific therapeutics. Numerous cell types in the spinal cord can release dynorphin which may lead to the negative effects on locomotor recovery reported here. However, it is unknown which cell type produces these effects. Prior literature has suggested that glial-mediated, but not neuronal-mediated, dynorphin contributes to allodynia. 74,75 Selectively targeting glial-derived dynorphin may be optimal. Finally, the effects of sex were not examined in the present study. Despite the incidence of SCI being far greater in males, there is still a need to study these phenomena in females. As previous studies have found sex differences in both pain perception following SCI and the efficacy of morphine, future studies should address the potential for sex differences in the morphine-induced attenuation of SCI recovery. 76 –79
Conclusions
Given the frequency of opioid prescription in the acute phase of SCI and the negative effects found both in the clinic and in experimental models, it is critical that we identify the mechanisms underlying the adverse consequences of opioids and improve the safety and utility of these indispensable analgesics. The current studies highlight the role of KOR activation in the morphine-induced attenuation of locomotor recovery after SCI. Coupled with our previous study showing that norBNI is also effective with epidural morphine, norBNI appears to be a potential adjuvant for clinical intervention, preventing the negative impact of morphine on recovery of locomotor function after SCI.
Footnotes
Authors' Contributions
Josephina Rau was responsible for the methodology, validation, formal analysis, investigation, resources, writing – original draft, visualization, supervision, and project administration. Annebel Hemphill, Kendall Araguz, Rachel Cunningham, Alexander Stefanov and Lara Weise were responsible for investigation. Michelle A. Hook was responsible for conceptualization, methodology, writing – review and editing, supervision, project administration, and funding acquisition. All authors read and approved the final manuscript.
Funding Information
This study was supported by the National Institute on Drug Abuse (NIDA) Drug Supply Program and by the Office of the Assistant Secretary of Defense for Health Affairs, through the Spinal Cord Injury Program under Award No. W81XWH 17-1-0629.
Author Disclosure Statement
No competing financial interests exist.