
Abstract
Chronic, often intractable, pain is caused by neuropathic conditions such as traumatic peripheral nerve injury (PNI) and spinal cord injury (SCI). These conditions are associated with alterations in gene and protein expression correlated with functional changes in somatosensory neurons having cell bodies in dorsal root ganglia (DRGs). Most studies of DRG transcriptional alterations have utilized PNI models where axotomy-induced changes important for neural regeneration may overshadow changes that drive neuropathic pain. Both PNI and SCI produce DRG neuron hyperexcitability linked to pain, but contusive SCI produces little peripheral axotomy or peripheral nerve inflammation. Thus, comparison of transcriptional signatures of DRGs across PNI and SCI models may highlight pain-associated transcriptional alterations in sensory ganglia that do not depend on peripheral axotomy or associated effects such as peripheral Wallerian degeneration. Data from our rat thoracic SCI experiments were combined with meta-analysis of published whole-DRG RNA-seq datasets from prominent rat PNI models. Striking differences were found between transcriptional responses to PNI and SCI, especially in regeneration-associated genes (RAGs) and long noncoding RNAs (lncRNAs). Many transcriptomic changes after SCI also were found after corresponding sham surgery, indicating they were caused by injury to surrounding tissue, including bone and muscle, rather than to the spinal cord itself. Another unexpected finding was of few transcriptomic similarities between rat neuropathic pain models and the only reported transcriptional analysis of human DRGs linked to neuropathic pain. These findings show that DRGs exhibit complex transcriptional responses to central and peripheral neural injury and associated tissue damage. Although only a few genes in DRG cells exhibited similar changes in expression across all the painful conditions examined here, these genes may represent a core set whose transcription in various DRG cell types is sensitive to significant bodily injury, and which may play a fundamental role in promoting neuropathic pain.
Introduction
Many neuropathic pain conditions involve plasticity of the peripheral somatosensory system. 1 After peripheral nerve injury (PNI), primary sensory neurons with cell bodies in dorsal root ganglia (DRGs) undergo functional alterations leading to amplification of noxious and innocuous stimuli (hyperalgesia and allodynia) and continuing activation of central pain pathways, 1,2 in part via increased electrical activity in primary somatosensory neurons. 1,3 -5 Alterations in gene and protein expression in DRG neurons have been linked to neuropathic pain. 6 –9 PNI can produce axonal injury, degeneration, regeneration, and sprouting, plus synaptic remodeling, inflammatory responses, death of DRG cells, and altered electrical activity of sensory neurons. 10 -12 Unbiased transcriptomic profiling of single primary somatosensory neurons shows that PNI induces a transcriptional response in axotomized sensory neurons in DRGs 13 and trigeminal ganglia. 14 This response suppresses cell identity and promotes axonal regeneration by inducing neuronal transcription factors such as ATF3, which induce the expression of regeneration-associated genes (RAGs). Although links between RAG expression and neuronal regeneration are evident, contributions of RAG expression to neuropathic pain remain unclear, 14 albeit plausible. 15
PNI also induces transcriptional alterations in non-neuronal DRG cells, including satellite glial cells and macrophages. 16 Insights into potential contributions of axotomy-independent gene expression in diverse cells to neuropathic pain might come from examination of DRGs in a neuropathic pain condition, contusive spinal cord injury (SCI), that involves little peripheral regeneration. Spinal contusion directly injures few peripheral axons but, like PNI, produces chronic pain linked to persistent hyperexcitability of DRG neurons. 17,18 In contrast, SCI models such as spinal transection, hemisection, or focal dorsal column transection axotomize large numbers of the central axons of sensory neurons, which fail to regenerate. These sectioning models are less clinically relevant than contusive SCI models. 16,19 -21
A major question is how transcriptional alterations induced in DRGs by contusive SCI compare with those induced by PNI. We have analyzed transcriptomic data from a rat thoracic spinal contusion model and combined it with a meta-analysis of RNA-seq datasets from common rat PNI models. We find major differences in the expression of many genes, including RAGs and long noncoding RNAs (lncRNAs). Unexpected evidence indicates that many SCI-associated alterations in DRG gene expression are caused by surgical injury rather than cord injury. Few transcriptomic similarities were found between rat neuropathic pain models and human DRGs linked to neuropathic pain, but the shared transcriptional alterations across species and models may reveal core genes important for driving persistent pain.
Methods
Spinal cord injury and sham surgery
All procedures complied with guidelines of the International Association for the Study of Pain and were approved by the institutional animal care and use committee. Male rats (200-300 g) were maintained under a 12:12 h reversed light/dark cycle, and all experiments were performed during the dark phase. 17 Spinal contusion injury was produced as described previously. 17,18 Briefly, rats receiving SCI were anesthetized with ketamine (80 mg/kg), xylazine (20 mg/kg), and acepromazine (0.75 mg/kg) before laminectomy of T10 vertebrae followed by an impact to the center of the cord at spinal T11 using an Infinite Horizon impactor (150 kdyne, 1-sec dwell time). The muscle layer was then sutured with 4.0 Vicryl and the skin closed with stainless steel wound clips. This injury produces equivalent effects on DRGs on both sides of the cord. 17
All SCI animals exhibited Basso, Beattie, and Bresnahan (BBB) hindlimb motor scores of 0–122 1 day after injury. Hindlimb motor function showed partial recovery when examined 1 month after SCI, as reported previously. 17,18 We define SCI as injury to the spinal cord itself, produced in our studies by controlled impact to the cord as part of the SCI procedure (SCIp). Data are reported for the SCIp group, which necessarily included surgical injury to skin, muscle, and bone as well as cord injury from the spinal impact. The SCI component of SCIp is inferred from differences between the SCIp group (n = 4) and the sham surgery control group (ShamSCI; n = 4). Post-operative effects of ShamSCI are inferred from differences between the ShamSCI and NaïveSCI (n = 4). groups run at the same time.
The methods used for our SCIp group differed from the SCIp methods used by another group for transcriptomic profiling of DRGs after SCI, 23 the results of which we use for comparisons. Their SCIp rats were injured at spinal level T2 by either contusion with an NYU impactor or complete transection with a scalpel. Other differences are described in the “Results” section.
DRG excision, RNA isolation, library preparation, and sequencing
Rats (four per group) were anesthetized 1 month after injury with an intraperitoneal injection of pentobarbital/phenytoin (390 mg/kg) in the mid-morning, prior to cardiac perfusion with ice-cold phosphate-buffered saline. Thoracic and lumbar DRGs were excised bilaterally below spinal level T11, yielding 16 DRGs per rat. DRGs from each rat were pooled and put into RNAlater reagent (Sigma Aldrich) for RNase-free homogenization on ice using a tissue homogenizer (OMNI International TH). Total RNA was extracted using RNeasy Mini Kit (QIAGEN). RNA was eluted with RNase-free water and its quality validated using the Agilent 2100 Bioanalyzer RNA 6000 Nano kit.
Rat neuropathic pain models and RNA-seq datasets
We combined our rat SCI RNA-seq data with datasets from three unilateral PNI models: spinal nerve transection (SpNT), 24,25 chronic constriction injury (CCI) of the sciatic nerve, 26 and partial sciatic nerve ligation (pScNL). 27 Rat strain, age, and sex, site of DRG sample isolation, number of days post-injury, and the type of control used (sham surgery and/or uninjured naïve) differed across studies (Table 1). We also compared our SCIp RNA-seq data to data obtained with two similar SCI models. 23 Read mapping and gene expression quantification were performed using a previous pipeline. 28 Datasets averaged 87.5% uniquely mapped reads (Supplementary Table S1). Supplementary Table S2 contains the raw counts matrix.
Peripheral Nerve Injury (PNI) and Spinal Cord Injury (SCI) Datasets Used for Transcriptomic Analyses
SCIp involves both injury to the spinal cord (SCI) and substantial surgical injury produced during exposure of the cord. The surgical injury component is estimated with sham surgery controls run at the same time (ShamSCI group).
DPI, days post-injury AUTH: DRG, dorsal root ganglia; SpNT, spinal nerve transection; CCI, chronic constriction injury of sciatic nerve; pScNL, partial sciatic nerve ligation; SCIp, spinal cord injury procedure.
Batch effect adjustment
Given that rat neuropathic pain datasets belong to independent studies and thus have inherent variability, we used ComBat, included in the Surrogate Variable Analysis (SVA) package, for batch effect adjustment. 29 Normalized and batch effect adjusted gene expression matrixes are found in Supplementary Table S2.
Classification of datasets
Datasets for each treatment group were assigned two scores, a DRG inflammation score and a DRG peripheral axotomy score, based on reported consequences of each injury treatment (see Table 2 and the “Results” section). Plots of principal components 1 and 2 of the normalized batch effect-adjusted count matrixes revealed clustered samples and outliers. Pearson correlation coefficients from the gene expression matrixes were used for hierarchical clustering utilizing complete linkage. Clustering of the Pearson correlation coefficient matrixes validated the classification method and identified outlier samples, which were excluded from further analysis.
Scores on Two 5-Point Scales Assigned to Datasets according to the Fraction of Sensory Neurons Likely to Have Had Their Peripheral Axons Transected (DRG Peripheral Axotomy Score) and Reports of Inflammatory Effects Likely to Influence DRG Cells (DRG Inflammation Score) by Each Treatment
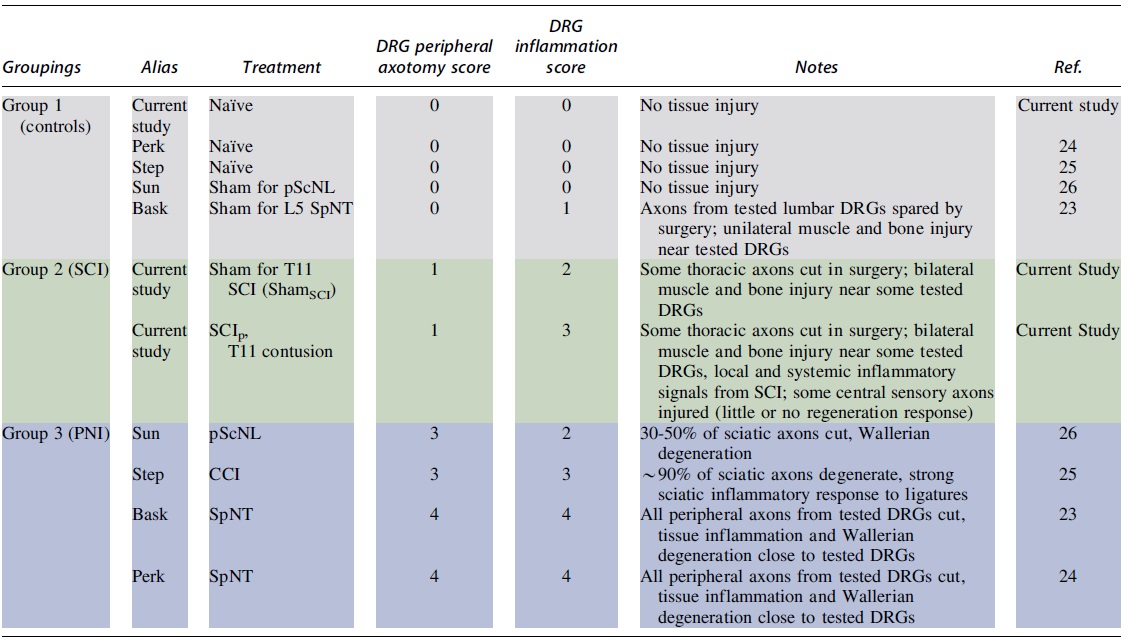
Colors show the groupings of treatment conditions based upon overall similarities of DRG peripheral axotomy scores and DRG inflammation scores.
DRG, dorsal root ganglia; pScNL, partial sciatic nerve ligation; SpNT, spinal nerve transection; SCI, spinal cord injury; ShamSCI, sham surgery control group; SCIp, spinal cord injury procedure; CCI, chronic constriction injury of sciatic nerve; PNI, peripheral nerve injury.
Analysis of differential gene expression
Batch effect-adjusted gene expression matrixes were used for finding differentially expressed genes (DEGs) using the limma R package. 30 Groups clustered by injury type were compared with groups with little or no injury. DEGs were defined by absolute value of log2 fold-change >0.5, false discovery rate (FDR) <0.05, and normalized gene expression >1.0 in at least one sample. DEGs are listed in Supplementary Table S4.
Gene set enrichment analysis
Lists of DEGs corresponding to samples in each principal component analysis (PCA) cluster were used to define significantly over-represented gene sets. The enriched gene sets are listed in Supplementary Table S5.
Pain protein–protein interaction networks
To understand the molecular mechanisms associated with neuropathic pain we built a protein–protein interaction (PPI) network based on a curated list of 1002 protein-protein interactions of 611 proteins associated with pain. 31
Gene set variation analysis
To compare rat pain datasets with single-cell RNA-seq gene signatures representing the cellular diversity underlying the DRG with and without injury, we adopted the gene set variation analysis approach. 32 Gene signatures were obtained from a recent study in which authors performed single-nucleus RNA-seq over multiple time-points on lumbar DRG cells from adult mice in naïve conditions, and after three PNI conditions: sciatic nerve transection + ligation (ScNT), spinal nerve transection (SpNT), and sciatic nerve crush. 13 Enrichment scores with FDR <0.05 were considered significant and were grouped through hierarchical clustering (see heatmaps in figures).
Guilt-by-association analysis
To infer potential functions of differentially expressed lncRNAs, we adopted the guilt-by-association approach. 33 This widely used approach to inferring gene function assumes that genes that are highly correlated will share some biological function. Thus, potential lncRNA functions were predicted to overlap with those of protein-coding genes having highly similar expression patterns and published functional annotation. For each differentially expressed lncRNA, a ranked list of protein-coding genes was obtained by ordering the Pearson correlation coefficient between the lncRNA and protein-coding genes using their gene expression in all datasets. Gene Set Enrichment Analysis (GSEA) 34 was performed to identify significantly enriched gene sets for each lncRNA. Additional information is provided in the Supplementary Methods.
Results
Transcriptional profiles of DRGs in SCI and PNI models differ
We performed a meta-analysis of data from publicly available DRG RNA-seq datasets collected in several rat unilateral PNI models and combined it with our SCI data. Rat PNI models included spinal nerve transection (SpNT), 24,25 chronic constriction injury (CCI) of the sciatic nerve, 26 and partial ligation of the sciatic nerve (pScNL; Table 1). 27 A separate comparison was made to DRG transcriptional profiles reported for two different thoracic SCI models in another study. 23 All data came from DRG samples collected 7 days or later after injury.
Raw sequencing files were processed with a previously developed RNA-seq pipeline using a comprehensive gene annotation including lncRNAs. Raw counts and FPKM values obtained from processed datasets are listed in Supplementary Table S2. To account for technical variations and batch effects inherent to datasets from independent studies we implemented ComBat 29 using log2 transformed normalized counts. PCA plots before batch effect adjustment showed a separation of samples by study (Supplementary Fig. S1A). The location of samples in the PCA plots became more homogeneous after batch effect adjustment (Fig. 1A and Supplementary Fig. S1B). The variability explained by PC1 increased after batch effect adjustment from 62.4% to 81.5%. A linear combination of PC1 and PC2 separated most of the injury and control samples. Supplementary Table S2 shows the normalized gene expression matrix after batch effect adjustment.
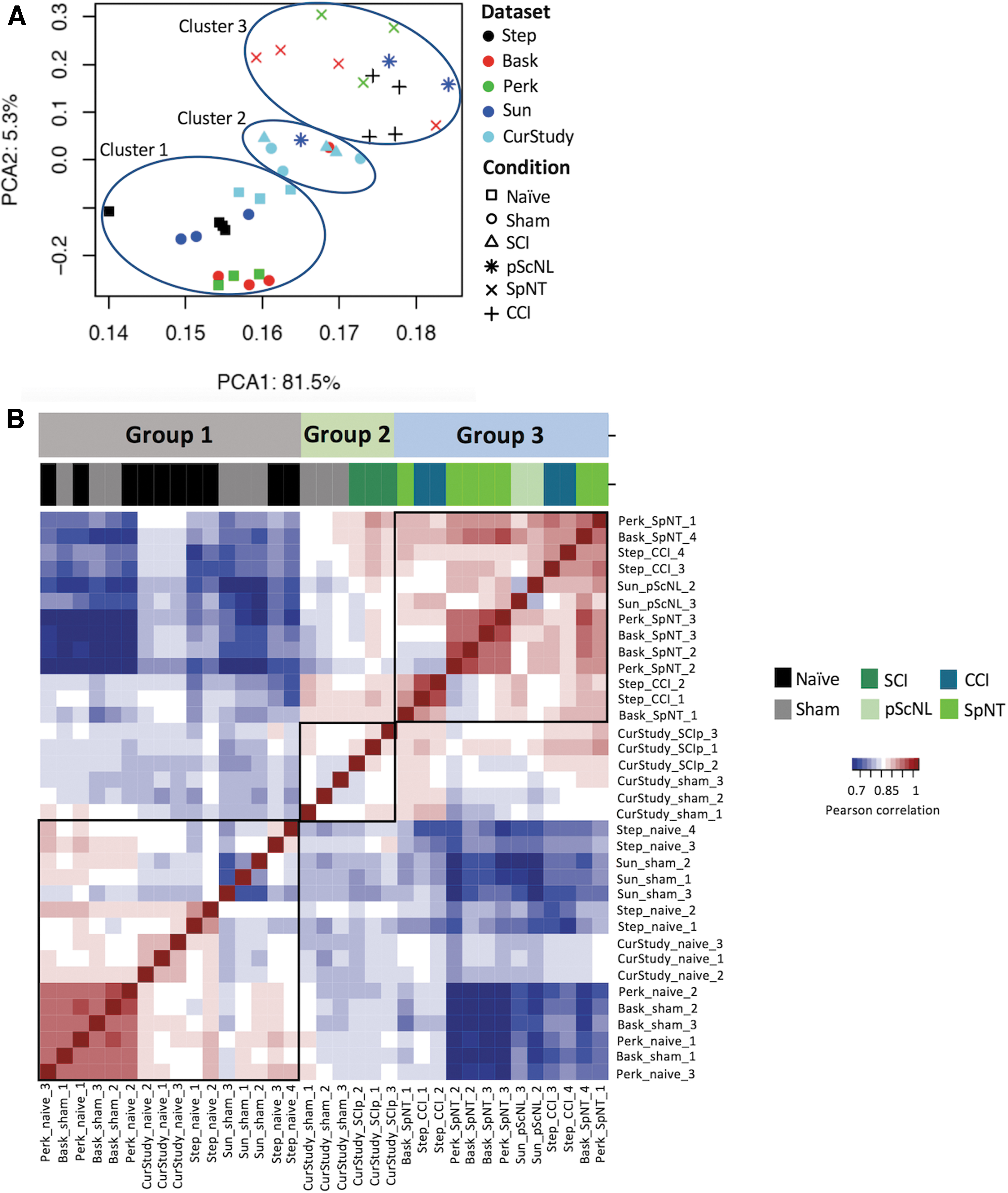
Clustering of central and peripheral pain datasets analyzed.
PCA plots showed a separation of DRG datasets into three clusters corresponding to treatment types (Fig. 1A). PCA Cluster 1 included control treatments producing little or no tissue injury: naive groups and sham surgery groups for SpNT (ShamSpNT) and for pScNL (ShampScNL). Cluster 2 included two treatments, our SCI procedure (SCIp) and the corresponding sham surgery procedure (ShamSCI). Cluster 3 included the rat PNI treatments (SpNT, CCI, and pScNL). To further characterize relationships between DRG gene expression and injury, we assigned crude scores for two major consequences of tissue injury for DRGs in each study (Table 2). The 5-point DRG peripheral axotomy score represents the fraction of sensory neurons in sampled DRGs likely to have had their peripheral axons transected. Because injury to central sensory axons in dorsal roots and dorsal columns induces almost no regeneration and possibly less pain than induced by axotomizing injuries of peripheral nerves, 20,35 -37 we did not include injury to the central axons of DRG neurons in the peripheral axotomy score. A maximal score (4) was given if >90% of axons in sampled DRGs were likely to be severely injured (cut in SpNT models), 38 while a score of 3 was given if ∼30-90% of the axons were likely to be severely injured (as indicated in pScNL and CCI models by extensive axonal degeneration). 39 –42
Because very large changes in gene expression have been reported for single DRG neurons after peripheral axotomy, 13 we assumed that axotomy of as few as 30% of the DRG neurons would result in easily detectable transcriptomic effects in the corresponding DRGs. While the SCI group was likely to receive injury to dorsal column axons (central A-fibers) bilaterally, and to some of the small fraction of central C-fibers innervating the thoracic injury site, 17 no peripheral C-fibers should have been injured by the spinal impact. However, both the SCIp and ShamSCI procedure, each of which can induce persistent pain, 43 would axotomize some sensory neurons in the minority of the sampled DRGs (primarily T12) that innervate skin, muscle, and bone injured during exposure of the spinal impact site (which represents a small fraction of the 16 sampled DRGs' receptive fields). Therefore, both the SCIp and ShamSCI groups received a total peripheral axotomy score of 1.
The 5-point DRG inflammation score represents exposure to inflammatory factors likely to stimulate DRG cells. Major contributors to the DRG inflammation score include proximity of sampled DRGs or sensory axons to a sufficiently recent surgical wound (SpNT and ShamSpNT unilaterally, and some SCIp and ShamSCI DRGs bilaterally, associated with the local release of cytokines and damage-associated molecular patterns [DAMPs]), and published reports that the injury model is associated with increased numbers of inflammatory cells in DRGs (PNI and SCIp) or with widespread inflammation (e.g., elevated cytokine and DAMP levels in blood and/or CSF) (SCIp). 44 –48 In addition, it was assumed that peripheral inflammation associated with nerve injury might alter the transcriptional state of the DRG by release of mediators from inflammatory cells responding to Wallerian degeneration close to the ganglion (especially in the SpNT groups) and possibly in neurons by retrograde axonal transport of molecules such as NGF in signaling complexes 49 from distant sites of inflammation. Both the SCIp and ShamSCI DRGs near T11 would be exposed to local inflammation resulting from the vertebral surgery, but all sampled DRGs in the SCIp group would also be exposed to inflammatory signals in the CSF resulting from SCI, 44,50 so the DRG inflammation scores assigned for ShamSCI and SCIp DRGs were 2 and 3, respectively.
Cluster 1, containing Naïve, ShamSpNT, and ShampScNL groups, had peripheral axotomy scores of 0 and inflammation scores of 0 to 2 (from tissue damaged during sham surgery). Cluster 2, containing SCIp and ShamSCI groups, had relatively low peripheral axotomy scores (1) and moderate or moderately high inflammation scores (2 or 3). Cluster 3 (Fig 1A), containing the PNI groups (SpNT, CCI, and pScNL), was characterized both by high DRG peripheral axotomy scores (3 or 4) and by moderate to high DRG inflammation scores (2 to 4). The location of nearly all the PNI Sham groups in Cluster 1 suggests that any contralateral effects of PNI (e.g., from systemic inflammatory effects) 48 on gene expression in uninjured DRGs are relatively weak.
Using PCA and correlation coefficients, we identified one Sun and colleagues pScNL sample 27 and one Baskozos and colleagues ShamSpNT sample 24 as outliers and removed them (Supplementary Fig. S2). Potential reasons for these outliers include insufficient axotomy during PNI in the pScNL rat, and inadvertent damage to the spinal nerve in the ShamSpNT rat. The heatmap of the remaining samples (Fig. 1B) demonstrates the alignment of treatment groups 1 to 3 (Table 2) with corresponding PCA clusters 1 to 3 (Fig. 1A).
SCIp and ShamSCI induce lower expression of RAGs in DRGs than do PNI models
The largest difference in injury-related factors between PCA Cluster 3 and both Clusters 1 and 2 was in the degree of peripheral axotomy of the tested DRGs, although the inflammation scores were also highest in Cluster 3. Confirming the importance of peripheral axotomy, we observed much higher pooled expression of canonical DRG RAGs (Atf3, Sox11, Sprr1a, and Flrt3) 13 in Cluster 3 (SpNT, CCI, pScNL) than in Clusters 1 and 2 (Fig. 2A), which had lower peripheral axotomy scores. Compared with controls, significantly higher fold-changes in expression of other RAGs and regeneration-associated transcription factors also were found in Cluster 3 (Supplementary Fig. S1C, S1D ; Supplementary Table S3). Importantly, while a small but significant increase in expression of these RAGs was found in the SCIp group compared with the corresponding NaiveSCI group, no significant difference was found between the SCI and ShamSCI groups (Supplementary Table S3; Fig. 2A). The lack of a difference in RAG expression between the SCIp and ShamSCI groups mirrors the overlap of these groups in the PCA clusters (Fig. 1A) and is consistent with most of the axotomy in the SCIp group being produced by transection of a minority of peripheral axons during the SCI surgery rather than by the spinal impact itself. This possibility is also supported by the reported lack of any effect of spinal transection at T12 on RAG expression in L1 DRGs, which probably did not innervate the incised region (∼2 cm distant). 51

Expression of canonical regeneration-associated genes (RAGs) and inclusion of an additional SCI dataset.
A second study yields SCIp profiles in Cluster 2 and it suggests that DRG transcriptional profiles are sensitive to diverse stresses
One other study 23 has reported DRG transcriptomes following contusive SCI in rats. For direct comparisons, we processed their datasets with the same pipeline and added them to our PCA analysis (Supplementary Fig. 2B). In contrast to our study, this study used female instead of male rats, their SCI was at the spinal T2 rather than T11 level, they sampled DRGs from T11 to L4 rather than T12 to L6 (i.e., they sampled neither the largest L5 DRG nor DRGs close to the contusion site), and they reported transcriptional effects of both spinal contusion and complete spinal transection. Despite these differences, all their SCIp samples (taken ∼3 months after spinal contusion or transection) were found in PCA Cluster 2, close to our T10 contusion samples (taken 1 month after spinal contusion). The clustering of SCIp samples from both studies outside of Cluster 3 confirms that SCIp induces changes in DRG gene expression that differ markedly from the transcriptional responses to PNI seen in Cluster 3.
Chariker and colleagues (2019) 23 used an uninjured NaiveSCI group as their sole control group for SCI. Surprisingly, all their NaiveSCI samples were found close to their SCI samples in Cluster 2, and were closer to all the SCIp and ShamSCI samples in both studies than to our NaiveSCI samples in Cluster 1 (Fig. 2B). The difference between the two NaiveSCI subclusters from each SCI study might in part be a sex difference, as 860 DRG genes have been reported to be differentially expressed between Naive male and female rats. 26 Moreover, several transcriptional regulators linked to injury and pain (see below), including Fos, Junb, and Socs3, showed much greater upregulation in female than male DRGs after CCI. 26 However, many more DRG genes (1904) were differentially expressed between our NaiveSCI group and the Chariker and colleagues. NaiveSCI group, suggesting the contribution of an additional differential factor affecting DRG gene expression in their study. An unusual feature of the Chariker and colleagues study 23 (see also 52 ) was that the housing condition for all groups was designed to restrict movement and thereby mimic SCIp -induced inactivity beginning 2-3 months before DRG processing. It is plausible that this prolonged inactivity produced a systemic stress, perhaps manifested as persistent low-grade inflammation. 53,54 Finding the Chariker and colleagues' Naive samples in PCA Cluster 2 instead of Cluster 1 with the other Naïve groups suggests that stress from forced inactivity may have significantly altered DRG gene expression. Because such stress differs from the injuries that are our focus here, the remaining SCIp analyses do not include data from the Chariker and colleagues study. 23
Fewer DEGs are found in DRGs after SCIp than PNI but some of the most highly enriched DEGs are shared
To identify genes that change expression after SCIp and PNI, we used limma 30 to compare the normalized and batch effect-adjusted gene expression matrixes of samples in Clusters 2 and 3 (representing procedures producing neuropathic pain) to those in Cluster 1 (representing control groups lacking neuropathic pain). Differentially expressed genes (DEGs) were defined as those showing an absolute value of log2-transformed fold-change >0.5, and FDR <0.05. DEGs with a normalized expression <1 in all samples were filtered out.
Whereas 2371 DEGs were found in Cluster 3 compared with Cluster 1 (Fig. 3B), only 636 DEGs were found in Cluster 2 compared with Cluster 1 (Fig. 3A). Only 582 DEGs were found in Cluster 3 compared with Cluster 2. DEGs in all comparisons included protein-coding and lncRNAs. Heatmaps of DEGs found comparing Clusters 2 and 3 to Cluster 1 are presented in Figure 3C and 3D. All DEGs are listed in Supplementary Table S4. These results show that the treatments having the highest peripheral axotomy scores and highest DRG inflammation scores (PNI models) change the expression of many more DRG genes than do the SCI-associated treatments (SCIp and ShamSCI).
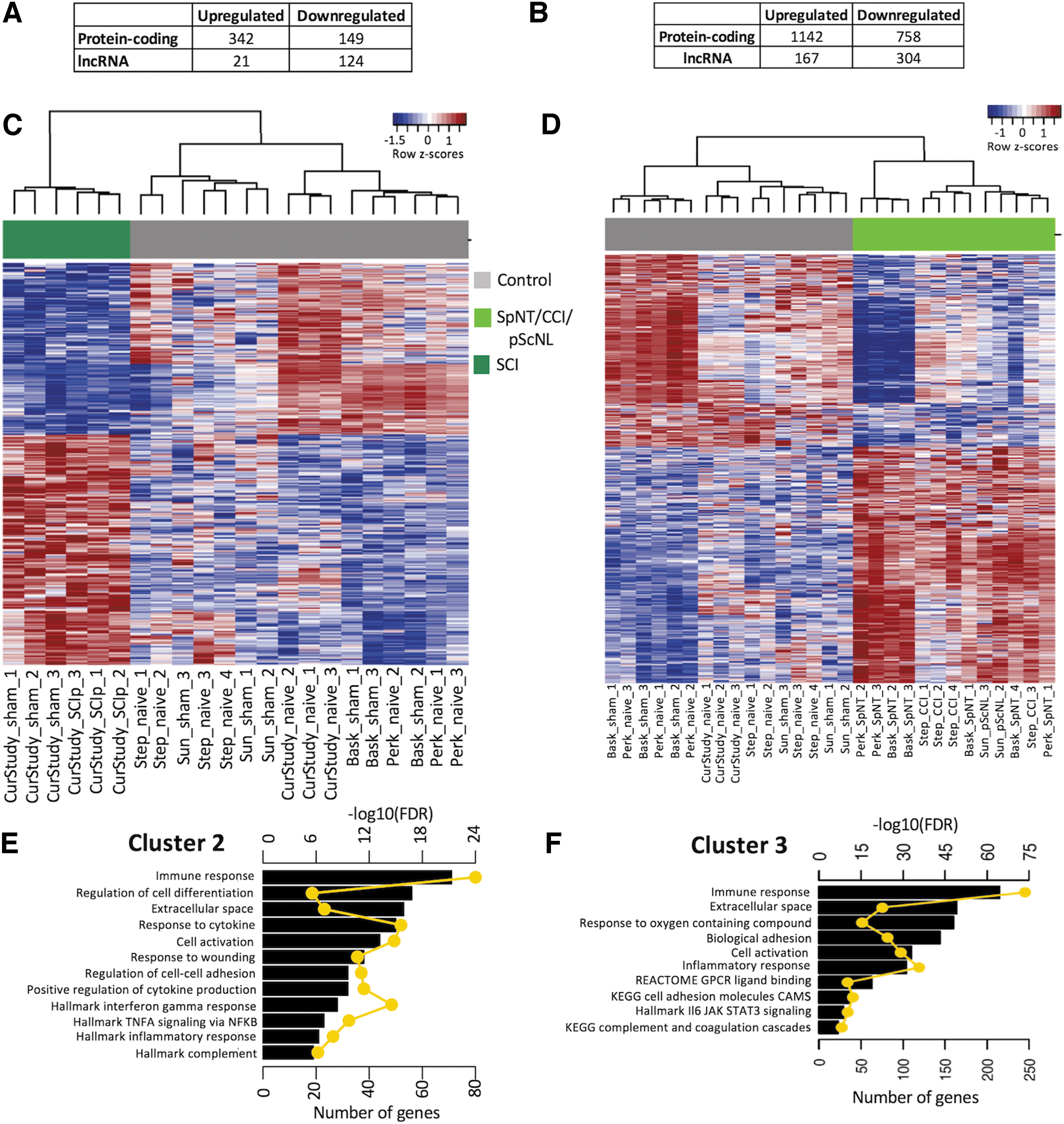
Analysis of differential gene expression.
To explore potential biological functions of DEGs in Clusters 2 and 3 we performed a gene set enrichment analysis. In Cluster 2, 261 and 14 significantly enriched gene sets (FDR <0.05 and number of overlapping genes ≥5) were found with upregulated or downregulated DEGs, respectively. The most enriched upregulated gene sets have been linked to inflammatory, immune, and wounding responses, including complement, cell activation, extracellular matrix, TNFα signaling, and interferon gamma responses (Fig. 3E). Enriched downregulated functions were mainly associated with plasma membrane functions (Supplementary Fig. S3).
Enrichment analysis for the up- and downregulated DEGs in Cluster 3 yielded 544 and 72 enriched genes, respectively. Interestingly, despite the substantially larger number of enriched genes in Cluster 3 than Cluster 2, the most enriched gene sets were often the same in the two clusters (Fig. 3F). For upregulated gene sets, these include associations with immune, inflammatory, extracellular space, adhesion molecule, and complement functions. For downregulated gene sets, ion transport and ion channel functions were highly enriched in both clusters, while gene sets linked to electrical and synaptic signaling in neurons were also enriched in Cluster 3 (Supplementary Table S5).
Many DEGs are shared by the SCIp and ShamSCI groups
The proximity of the SCIp to ShamSCI samples on the PCA graph (Fig. 1A) indicated many genes induced by these treatments were shared. Indeed, a comparison of DEGs relative to the NaïveSCI group showed that 59% of the 3617 SCIp DEGs were the same as in the 3026 ShamSCI DEGs (Fig. 4A; Supplementary Table S4). When ranked by the largest expression differences, the top 100 DEGs included 37 shared genes. This overlap suggests that many changes in DRG gene expression in the SCIp group are caused by the tissue injury and consequent inflammation produced by the surgery to expose the spinal cord. Nevertheless, comparison of the SCIp and ShamSCI groups showed that expression of 207 DRG genes was preferentially altered by SCI itself. To broaden the list of DEGs potentially important for SCI-induced pain, we used a p value <0.05 and the previously described filtering thresholds (absolute value of log2 fold-change >0.5 and gene expression >1). Some of the 252 genes that were upregulated in SCIp samples versus ShamSCI samples (SCI-induced genes, Supplementary Table S4) have been implicated previously in regulating pain. These include vasoactive intestinal peptide (Vip), 55 c-Fos (Fos), 56 and urocortin (Ucn), a protein that may inhibit pain. 57 Some of the 219 genes that were significantly downregulated in SCIp versus ShamSCI samples (SCI-repressed genes; Supplementary Table S4) have also been implicated in pain, for example natriuretic peptide C (Nppc), which may induce pain. 58
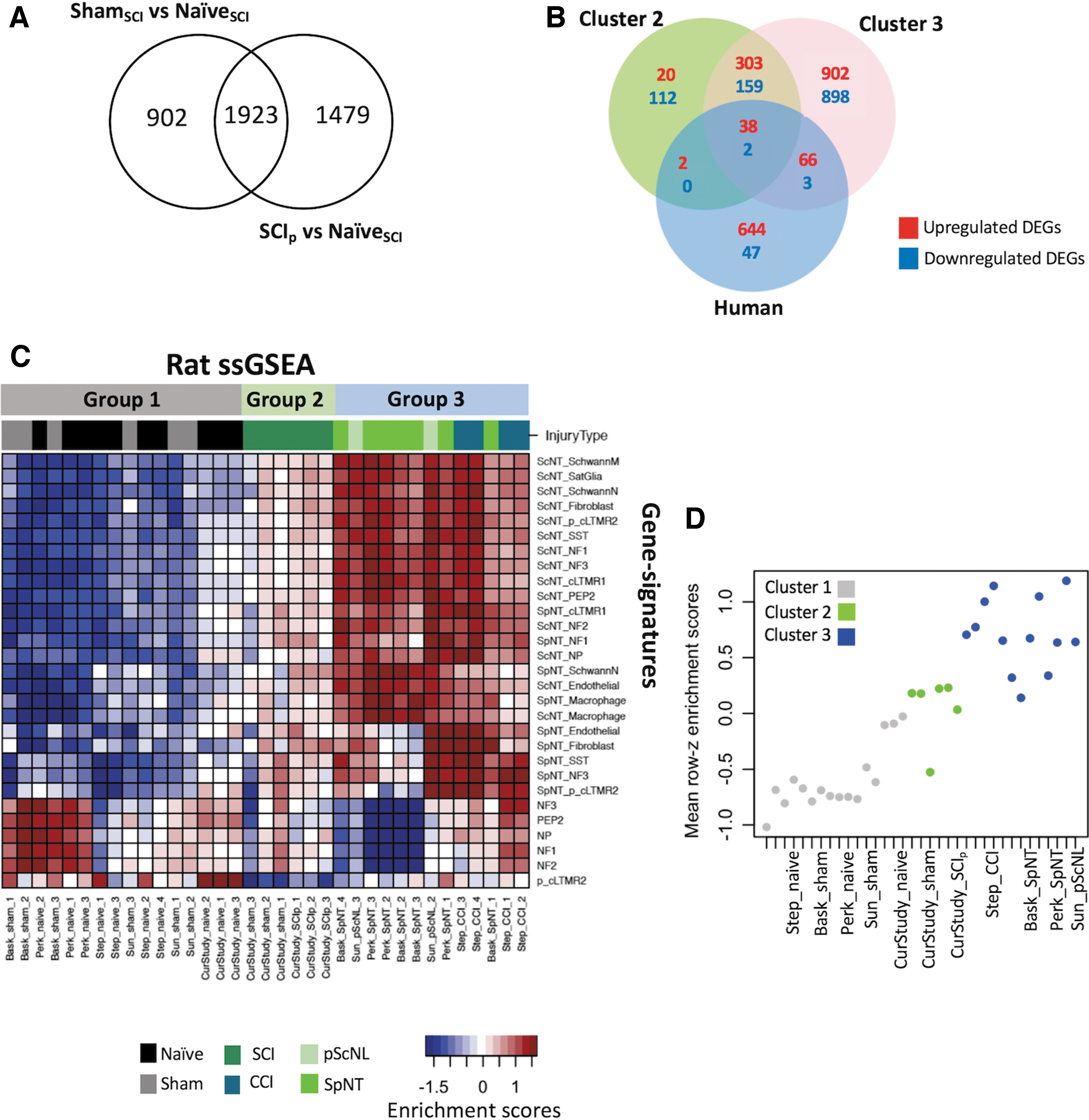
Differentially expressed genes (DEGs) shared between spinal cord injury procedure (SCIp) and sham surgery control (ShamSCI) groups, between rat neural injury models and a human neuropathic pain study, and single-sample Gene Set Enrichment Analysis (GSEA).
Few DEGs are shared between DRGs in rat neural injury models and human pain-associated DRGs
A pioneering study analyzed transcriptomes from DRGs excised from patients in which tumors compressed nerve roots. 59 DEGs were found in DRGs corresponding to painful dermatomes compared with DRGs corresponding to dermatomes without reported pain. Like DRGs in SCI conditions, these human DRGs were subject to nearby inflammation and possibly some axotomy of peripheral fibers. We compared human genes that were differentially expressed in pain-associated DRGs (pain DEGs) versus DRGs without a pain association to rat genes that were differentially expressed in Clusters 2 and 3. We found an overlap of only 40 (5%) and 104 (14%) upregulated DEGs out of 750 upregulated human pain DEGs with DEGs found in Cluster 2 (SCIp and ShamSCI groups) and Cluster 3 (PNI groups), respectively (Fig. 4B). Only two and five downregulated DEGs overlapped in human pain DEGs and Cluster 2 and Cluster 3 DEGs, respectively. The 38 upregulated DEGs overlapping all subsets included genes coding for complement proteins (C1QA, C1QB, and C1QC), leucine zipper proteins (FOS), suppressor of cytokine signaling (SOCS3), and transcription factors (ATF3, MYC, and JUN; Supplementary Table S6).
Rat DRG expression profiles correlate with single-cell gene signatures of diverse mouse DRG cell types
Given that all datasets in this study were obtained from bulk DRG samples, we considered whether the observed transcriptional differences resulted from dissimilar cellular composition. Such changes might occur because of preferential death of specific cellular populations, dedifferentiation, 13 other phenotypic changes, and proliferation or infiltration of non-neuronal cell types. 16,47,60 To gain insight into possible changes in phenotypic composition of the rat DRG datasets, we compared the transcriptional profiles from this meta-analysis to profiles of mouse DRG cells at single-nucleus resolution with and without sciatic nerve transection (ScNT) or spinal nerve transection (SpNT). 13
Single sample gene set enrichment analysis (ssGSEA) 61 yielded enrichment scores for each pairing of DRG rat expression profiles and single cell-derived DRG gene signatures (Supplementary Table S7). Enrichment scores indicate the degree to which genes representing cell types are coordinately up- or downregulated within DRG expression profiles after injury. High positive enrichment scores indicate an expression profile resembling the expression pattern of the cell type gene profile evaluated. Significant differences in enrichment scores (FDR <0.05) between control and injury samples were determined with Limma. 30
Using heatmaps of enriched scores and hierarchical clustering, we found three clusters that correspond to previously defined neuronal groups (Fig. 4C). Rat Naïve and Sham samples showed transcriptional profiles similar to mouse single-cell Naïve gene signatures, with significant enrichments of NF1, NF2, NF3, NP, PEP2, and p_cLTMR2 neuron subpopulations. Datasets of Cluster 2 (SCIp and ShamSCI) showed no high enrichment with any neuronal subpopulation. Interestingly, Cluster 2 enrichment scores were more similar to those of Cluster 3 than Cluster 1. This suggests that, after SCIp and ShamSCI, some DRG neurons acquire phenotypes similar to PNI. 13 The mean enrichment scores for all cell types in each dataset showed a continuum in which Naïve and Sham groups had the highest negative enrichment, whereas datasets of Cluster 3 had positive enrichment (Fig. 4D), while Cluster 2 datasets (SCIp and ShamSCI) occupied an intermediate position. The high enrichment scores of the rat SpNT, pScNL, and CCI DRG groups associated with both neuronal and non-neuronal cell types in the single-cell mouse study (sensory neurons, macrophages, satellite glial cells, Schwann cells, fibroblasts, endothelial cells) confirm that PNI in rodents causes large phenotypic changes in diverse cell types, presumably reflecting alterations related to axonal regeneration and inflammation.
Downregulated lncRNAs are prominent in SCIp groups
We ranked DEGs in the Cluster 2 groups (SCIp and ShamSCI) by fold-change relative to Cluster 1 and observed that the top 12 downregulated genes were lncRNAs. In contrast, the top downregulated DEGs in the PNI groups were mainly protein-coding genes. The majority of upregulated DEGs in SCI and PNI models were protein-coding genes. Intrigued by why the most downregulated SCI genes were lncRNAs, we performed a guilt-by-association analysis to infer potential functions for each differentially expressed lncRNA. Protein-coding genes were ranked by the correlation of their gene expression with lncRNAs across all samples, and a gene set enrichment analysis (GSEA) was performed using previously defined gene sets. Normalized enrichment scores (NES) for every gene set and lncRNA were assessed for the degree to which each was overrepresented at the top or bottom of the ranked list of protein-coding genes (Supplementary Table S8). Ten examples of the gene sets that were highly enriched using the top 12 rat SCIp downregulated lncRNAs are shown in Figure 5A.
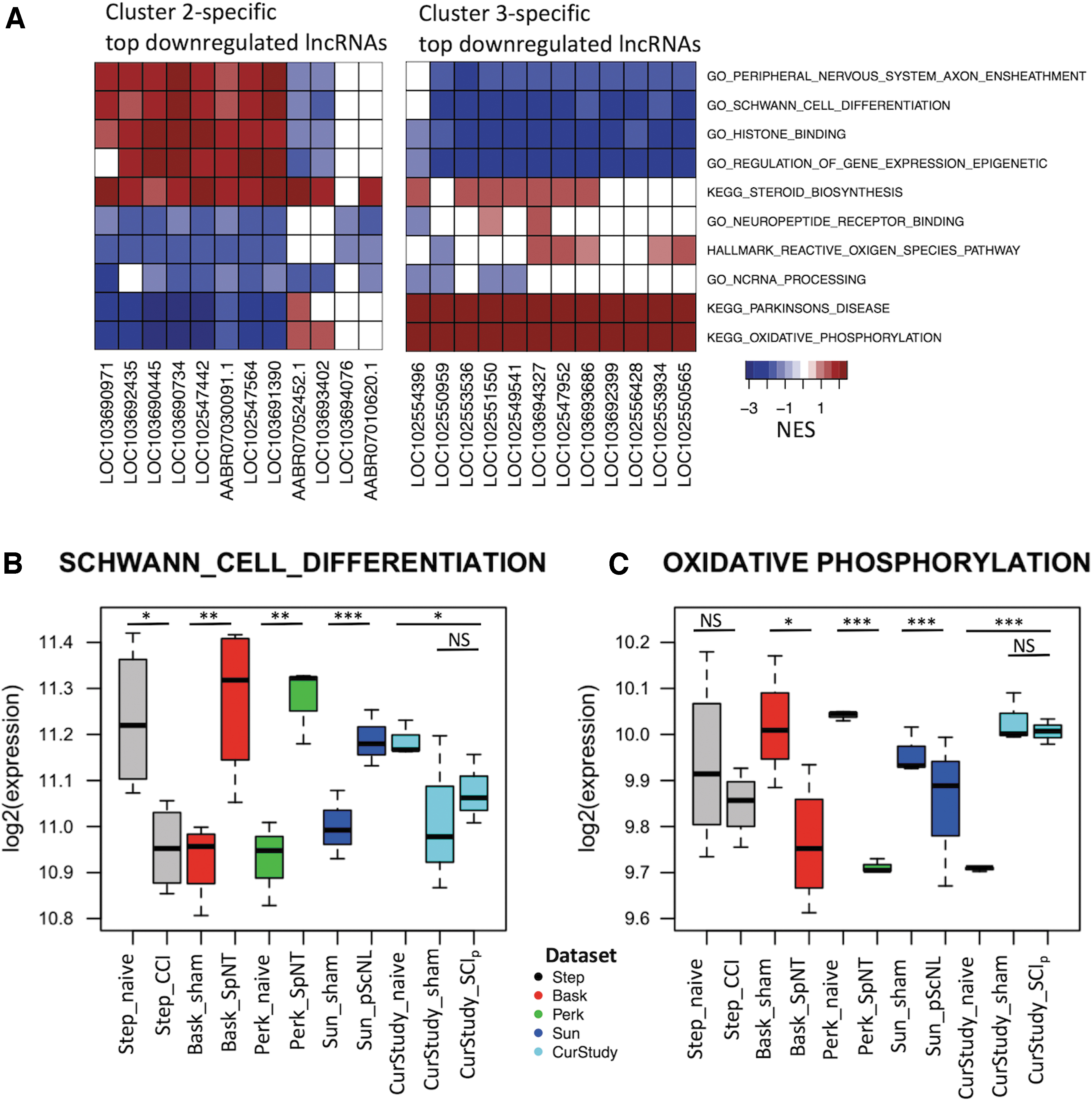
Guilt-by-association analysis.
Because the lncRNAs were downregulated, a negative NES represents upregulation of the genes in the associated protein-coding gene set. A positive correlation yields a positive NES, meaning genes in that set were downregulated (Supplementary Table S9). With the caveat that a given gene or protein may have multiple or complex functions, our results suggest that downregulated lncRNAs in rat SCIp and ShamSCI groups are involved in decreasing peripheral axon ensheathment (myelination), and two other functions that could reduce myelination as well as tissue repair (Schwann cell differentiation and steroid biosynthesis; Fig. 5A). 62 In addition, functions involved in transcriptional control (histone binding and regulation of gene expression) were associated with downregulated lncRNAs in rat SCIp, suggesting decreased binding of chemical modifiers to histone proteins and decreased gene regulation. Decreased histone modification activity would impair modifications of a cell's transcriptional regulatory mechanisms. 63 Decreased synthesis of steroids may also reduce neurotrophic and neuroprotective effects, 64 reducing recovery after SCI. Downregulated lncRNAs in rat SCIp and ShamSCI groups were also associated with increased expression of genes involved in neuropeptide receptor binding, generation of reactive oxygen species, noncoding RNA processing, Parkinson's disease, and oxidative phosphorylation. Surprisingly, the most downregulated PNI-associated lncRNAs in the majority of gene sets showed opposite enrichment directions in Clusters 2 and 3 (Fig. 5A). For example, the gene sets associated with Parkinson's disease and oxidative phosphorylation have a positive NES after PNI (Cluster 3), indicating these functions are decreased after axotomizing nerve injury, in contrast to SCIp (Cluster 2). Oxidative phosphorylation in the mitochondria includes genes associated with Parkinson's disease 65 ; their reduced expression may decrease energy production and increase neuronal death. Similarly, after axotomizing nerve injury there was a moderate decrease in expression of genes involved in binding of neuropeptides to their receptors (potentially due to neuronal death). Interestingly, downregulated lncRNAs in both SCIp and PNI injuries were associated with genes that increase the processing of noncoding RNAs, suggesting a higher conversion of primary noncoding RNAs to mature noncoding molecules. The latter finding reinforces emerging roles of lncRNAs in nerve injury and neuropathic pain. 66 Genes significantly correlated (p < 0.05) with DE lncRNAs and enriched in the aforementioned gene sets are listed in Supplementary Table S10.
To further investigate changes in expression of the gene sets found, we used box plots for each dataset to show the mean log2-transformed gene expression of all genes belonging to the indicated gene category. Figure 5B shows the direction of regulation of genes in the Schwann cell differentiation category across conditions. Two PNI conditions found to be in Cluster 3 (SpNT, pScNL) exhibited a clear upregulation of this gene set, while SCIp and ShamSCI groups in Cluster 2 appear to have this gene set downregulated, consistent with positive NES scores. Even though the CCI group is in Cluster 3, the expression of Schwann cell differentiation genes in this group was downregulated, possibly related to more extensive axonal degeneration in this model. Similar results were found for the oxidative phosphorylation gene set (Fig. 5C), albeit in the opposite direction compared with the Schwann cell differentiation set. The directions of regulation depicted in the box plots confirm what was found through the guilt-by-association analysis of downregulated lncRNAs in Clusters 2 and 3, except for the CCI group. Supplementary Table S11 shows the metrics used to build Figure 5B and 5C.
Common signaling genes are implicated in DEGs shared by the injury models
To examine similarities across injury groups, we compared Cluster 2 and Cluster 3 DEGs in different neuropathic pain models (Fig. 6A). 502 DEGs were shared between Clusters 2 and 3, with a high proportion (79%) of Cluster 2 DEGs included in Cluster 3 DEGs. Upregulated genes common to both clusters were mainly protein-coding genes (94%) with few lncRNAs, whereas common downregulated genes were similarly distributed between protein-coding RNAs (55%) and lncRNAs (45%). To further compare gene expression across clusters, we examined fold-changes of the common DEGs (Fig. 6B). We found a significantly higher change (p = 0.00022) in Cluster 3 than Cluster 2, consistent with the higher peripheral axotomy scores and DRG inflammation scores of Cluster 3. Supplementary Table S12 lists the DEGs common and specific to different injury groups.
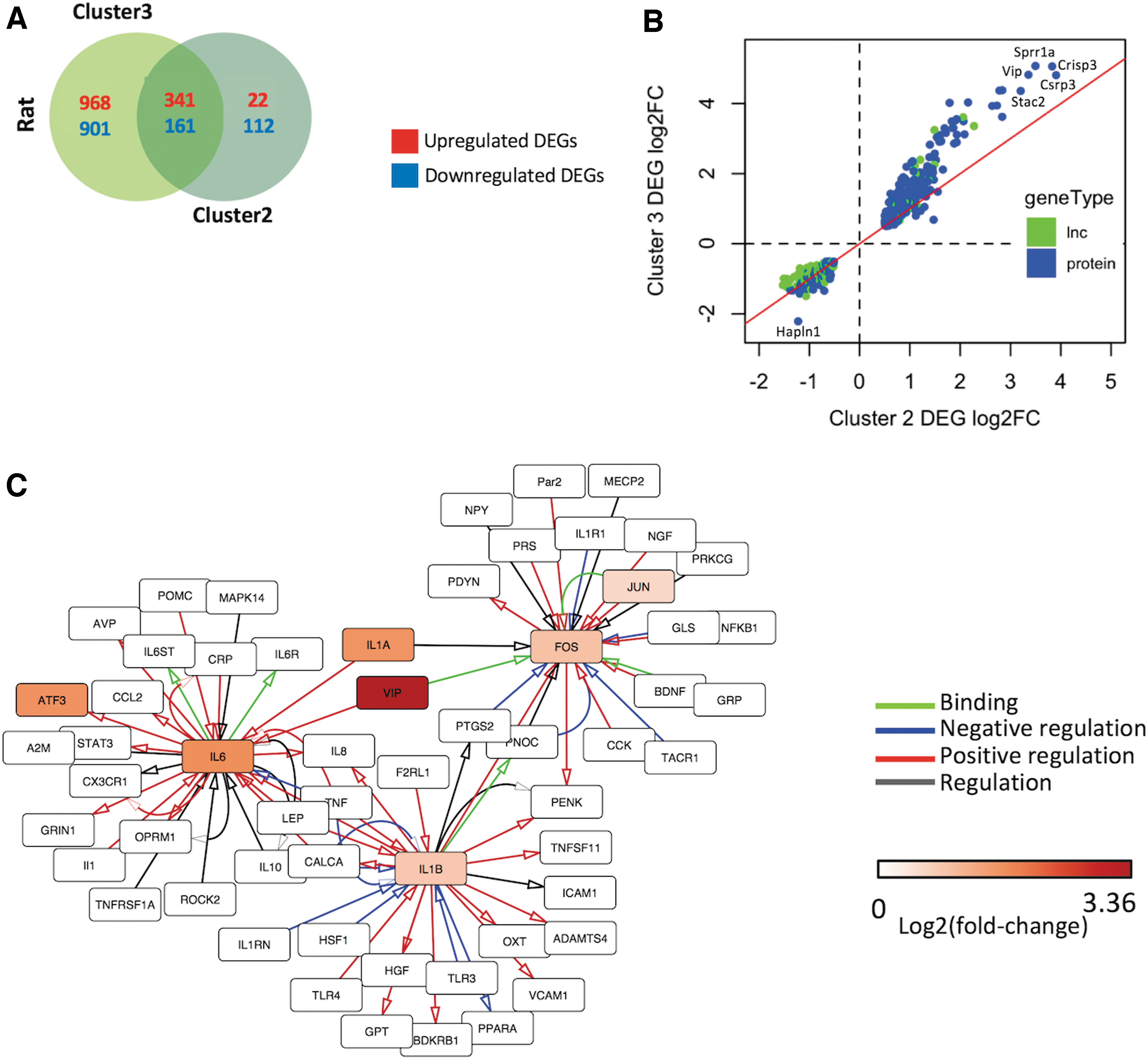
Analysis of differentially expressed genes (DEGs) common to neuropathic pain models.
To help elucidate signaling components in the pathways that change expression in SCI and PNI models, we used a curated human PPI network consisting of 1002 interactions of 611 proteins previously implicated in pain 31. Of the 502 rat DEGs common to the injury models examined, 34 had 161 known interactions. DEGs with the highest number of interactions included those coding for inflammatory cytokines IL-6, IL-1α, and IL-1β, and transcription factors FOS and JUN. Figure 6C shows the subnetwork containing the three most highly connected hub genes.
Discussion
Much is known about alterations in DRG cells that accompany neuropathic pain but how pain-linked alterations of these cells can persist chronically remains a mystery. 5 One possibility is that transcriptional reprogramming of DRG neurons and/or other DRG cells underlies persistent alterations that promote pain, as has been shown for alterations in several DRG cell types that promote axonal regeneration after PNI. 13,16 Apparently similar neuropathic pain can also be induced by damage to the spinal cord and other parts of the central nervous system 1,17 -19,67 -69 and such injury fails to induce significant axonal regeneration. 20,37 Because both inflammatory signals and peripheral axotomy have been suggested to induce persistent pain, 15,70 it is useful to compare pain-inducing injury models that produce quite different degrees of DRG inflammation and peripheral axotomy. Focusing on rats, we have compared DRG transcriptional profiles in a contusive SCI model that damages few peripheral sensory axons while producing both neuropathic pain 17,18 and central neuroinflammation 44,50 to published DRG transcriptomes in PNI models known to produce axotomy, inflammatory reactions, and persistent pain. 71
To measure transcriptional alterations, we downloaded and analyzed raw RNA sequencing datasets from rat SpNT, pScNL, and CCI models in combination with a dataset from our rat SCI model. PCA on these datasets and their corresponding controls indicated three clusters distinguished by type of treatment. To examine correlations of the treatments with two major factors widely considered important for neuropathic pain, we characterized each model and its control treatment along two crude 5-point scales based on published descriptions of the models. The DRG peripheral axotomy score represents the rough fraction of sensory neurons in sampled DRGs that had their peripheral axons transected. Injury to the central axons of DRG neurons was ignored because such injury induces almost no regeneration and possibly less pain than is induced by axotomizing injuries of peripheral nerves. 20,35 -37 The DRG inflammation score represents reported inflammatory factors that can stimulate DRG cells, including factors released at a nearby surgical site, factors transported within intact fibers in a nerve containing degenerating fibers, and factors involved in systemic and DRG inflammation. 44 –47 Compared with Cluster 1 controls, Cluster 3 (PNI groups) comprises DRGs having substantial peripheral axotomy plus exposure to intense inflammation, while Cluster 2 (the SCIp and ShamSCI groups) comprises DRGs having intermediate degrees of peripheral axotomy and exposure to inflammatory factors.
Compared with Cluster 1, there were nearly four times as many DEGs in Cluster 3 as in Cluster 2, indicating a much larger effect on DRG gene transcription caused by PNI than by SCIp, in agreement with the higher peripheral nerve axotomy and DRG inflammation scores. Differential expression of RAGs, including Atf3, Sox11, Sprr1a, and Flrt3, was also much higher in PNI models than in the SCIp model. However, RAGs were not the most enriched DRG gene sets in Cluster 3. This is consistent with their predominant expression in neurons (the only DRG cells that regenerate peripheral processes), which constitute only ∼10% of the total number of cells in the DRG 8. Interestingly, the most enriched DEGs in Clusters 2 and 3 were often the same. These included upregulated gene sets associated with immune, inflammatory, extracellular space, adhesion, and complement functions, and downregulated gene sets linked to ion channel and ion transport functions.
The lower expression of inflammation-related gene sets in Cluster 2 than Cluster 3 is consistent with the degree of DRG inflammation being lower in the SCI groups at the 1-month post-injury time of sampling than in the PNI models sampled 2-3 weeks after injury. A limitation of our comparisons is that SCI-related and PNI-related transcriptomes are not available for identical times following injury, and temporal differences may contribute to the differences in gene expression among the selected studies. Importantly, the rat bulk DRG gene sets enriched in Cluster 3 largely agree with bulk and single-cell DRG RNA-seq findings from mouse PNI models. 13,16,72 -74 The correlation of rat injury-associated DRG gene signatures with single nucleus-derived subpopulation signatures reported for mice, 13 suggests that PNI and, to a lesser extent, contusive SCI induce phenotypic changes in diverse DRG cell types, including neurons, inflammatory cells, and satellite glial cells. 16
Relatively few DEGs in human DRGs reported to be associated with neuropathic pain 59 were shared with rat genes expressed differentially in both Cluster 2 and Cluster 3 compared with Cluster 1. The small set of shared upregulated genes included several that have been linked to DRG axonal regeneration, including Fos, Jun, Atf3, Myc, 75 and genes for complement proteins. In the case of Fos, Jun, and Atf3, these represent important nodes in a human PPI network implicated in pain. 31,76 Other prominent hub genes in this network we found to be differentially expressed (notably interleukin [Il]-6 and Il-1b) are consistent with major roles played by inflammatory responses involving DRGs in these neuropathic pain models. 44,77 Il-6 and Il-1B inflammatory cytokines were also DEGs in the human neuropathic pain study. Surprisingly, the most upregulated RAG we found in rat neuropathic pain models, Sprr1a, is not differentially expressed in human DRGs. This transcriptional difference adds to recent findings of some pain-related cellular divergences between human and rodent DRGs. 78 –81
Another unexpected finding was the extensive overlap between DEGs induced by SCIp and those induced by the ShamSCI procedure. SCIp transcriptional profiles of DRGs from another contusive SCI study in rats 23 also localized to PCA Cluster 2, supporting the SCIp results we found. Interestingly, in our SCI experiments, the ShamSCI group localized to Cluster 2 rather than to the control Cluster 1, and a majority of the SCI DEGs (compared with the NaïveSCI group) were shared with the ShamSCI group. This overlap of DEGs 1 month after injury is consistent with earlier observations that ShamSCI surgery induced both persistent hyperalgesia 43 and nociceptor hyperexcitability, 82 albeit more weakly than induced by SCI. The overlap of DEGs indicates that many SCIp DEGs represent chronic DRG responses to the substantial tissue injury (to bone, muscle, and skin) needed to expose the cord rather than responses to spinal injury alone. Some of these shared DEGs might contribute both to persistent postsurgical pain and to SCIp pain. Nonetheless, a few genes were differentially expressed between the SCIp and ShamSCI groups, and it will be interesting to determine whether some of these (e.g., Fos, Vip, and Urc) might be especially important for in DRGs central neuropathic pain. For all neuropathic pain models, including SCI, mRNA translational changes, independent of transcriptional alterations, are also likely to play key roles in the development and maintenance of persistent pain, as observed for PNI. 9,83
Transcriptional profiles of DRG neurons after SCI are also available from a mouse study that examined transcriptomes of small neurons innervating hairy skin of the hindpaw 4 days after thoracic clip compression. 84 However, while Fos in the mouse study was also an upregulated DEG in this study, many of the other neuronal DEGs differed from the SCIp DEGs in whole DRGs found in the present study. A major experimental difference was their use of dissociated, live-sorted neurons. Their finding of substantial Atf3 expression in the NaiveSCI group suggests that transcriptional alterations caused by acute stresses from recent dissociation contributed to the different SCIp transcriptional profiles. In addition, SCI may have transcriptional effects during the first few days after injury that differ from the chronic effects we analyzed here. Notably, SCI triggers an enhanced intrinsic growth state that is expressed in dissociated nociceptors 3 days but not 1 month after injury. 85 This raises the question of whether subsets of the genes promoting peripheral axonal regeneration after PNI also promote the central sprouting of nociceptors observed acutely after SCI. 68,86,87
We also examined the expression of lncRNAs, which can regulate gene expression 88,89 and might be involved in DRG alterations related to axonal regeneration and neuropathic pain. 90,91 Analysis of DRG datasets disclosed partial overlap of DEGs and especially of highly enriched genes between Cluster 2 (SCIp and ShamSCI groups) and Cluster 3 (PNI groups). Remarkably, the top 12 downregulated DEGs in Cluster 2 were lncRNAs, in stark contrast to Cluster 3 where the top 13 downregulated DEGs were protein-coding genes. The most downregulated lncRNAs in Cluster 2 datasets were associated with gene sets (e.g., Schwann cell differentiation and oxidative phosphorylation) that were not enriched in GSEA of protein-coding genes alone. Differences we observed in the regulation of these gene sets between Cluster 2 and much of the Cluster 3 data set may reflect the profound differences in axonal regeneration capacities of the CNS compared with peripheral nerves. While a functional characterization of these novel lncRNAs is still needed, our results suggest that downregulation of some of these lncRNAs may contribute to neuropathic pain after SCI, and may offer potential therapeutic targets.
In sum, our combined analysis has revealed striking differences between transcriptional responses in PNI models and in a common SCI model, which is attributed to much less injury to peripheral axons and less inflammation of the DRG in SCI than in PNI. While some prominent injury-associated DEGs are shared across PNI and SCI models, much of the altered expression after SCIp can be explained by persistent transcriptional responses to the tissue injury required to access the cord for experimental SCI. Large transcriptional differences also were found between all the rat injury models examined and the only reported transcriptional analysis of human DRGs linked to neuropathic pain. The differences across injury models (including lncRNA responses) and species highlight the transcriptional sensitivity of DRGs to various injury-related stresses, the diversity of transcriptional responses in DRG cells, and perhaps to differences in associated pain. Conversely, the small number of genes that consistently changed expression across diverse, painful injury models may offer insight into core genes and fundamental mechanisms important for driving many forms of persistent pain.
Footnotes
Acknowledgments
We thank Emily Spence for performing SCIp and ShamSCI surgeries, Yan Wang for assistance in isolating DRGs, and Haichao Wei for his help with data analysis. The authors acknowledge the Texas Advanced Computing Center (TACC) at The University of Texas at Austin for providing HPC resources that have contributed to the research results reported within this paper.
Availability of Data
The datasets generated in the current study are available from NCBI/GEO under accession number GSE202111. Publicly available datasets used in the meta-analysis can be found under accession numbers GSE100122, GSE107180, GSE53762, and GSE117526.
Authors' Contributions
CWD, ETW, and JQW conceived the study and acquired funding. CWD, RCDD, ETW, and JQW further developed the conceptualization and experimental design. YL collected and processed the DRGs. YY performed the RNA sequencing. RCDD conducted the data analysis. All authors interpreted the results. RCDD and ETW wrote the manuscript with input from all authors, and all authors edited and reviewed the manuscript.
Funding Information
This work was supported by grants from the NIH (R01 NS088353, R21 NS113068-01, R21EY028647-01) and The Knight fund- Senator Lloyd & B.A. Bentsen Center for Stroke Research to JQW, and by NIH Grant NS091759 to CWD and ETW.
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Methods
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
Supplementary Table S6
Supplementary Table S7
Supplementary Table S8
Supplementary Table S9
Supplementary Table S10
Supplementary Table S11
Supplementary Table S12
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.