
Abstract
Traumatic brain injury (TBI) has been associated with several lasting impairments that affect quality of life. Pre-clinical models of TBI have been studied to further our understanding of the underlying short-term and long-term symptomatology. Neuromodulation techniques have become of great interest in recent years as potential rehabilitative therapies after injury because of their capacity to alter neuronal activity and neural circuits in targeted brain regions. This systematic review aims to provide an overlook of the behavioral and neurochemical effects of transcranial direct current stimulation (tDCS), transcranial magnetic stimulation (TMS), deep brain stimulation (DBS), and vagus nerve stimulation (VNS) in pre-clinical TBI models. After screening 629 abstracts, 30 articles were pooled for review. These studies showed that tDCS, TMS, DBS, or VNS delivered to rodents restored TBI-induced deficits in coordination, balance, locomotor activity and improved cognitive impairments in memory, learning, and impulsivity. Potential mechanisms for these effects included neuroprotection, a decrease in apoptosis, neuroplasticity, and the restoration of neural circuit abnormalities. The translational value, potential applicability, and the interpretation of these findings in light of outcome data from clinical trials in patients with TBI are discussed.
Introduction
Traumatic brain injury (TBI) is a life-altering incident classified as an injury to the head resulting from external physical forces. 1,2 TBI currently remains a critical public health concern and is the leading cause of mental health illnesses, death, and disabilities associated with trauma. 3,4 In addition to the initial primary neurological deficits, TBI may be followed by a spectrum of progressive long-term physical and psychosocial impairments, including cognitive dysfunction (e.g., attention, memory), neurological symptoms (e.g., headaches, dizziness), neuropsychiatric disorders (e.g., anxiety, post-traumatic stress disorder), and late onset neurodegenerative diseases. 5
Clinical cases of TBI are drastically heterogeneous because of the varying degrees of primary injury and secondary complications that arise at molecular and cellular levels. 6 To acquire a more comprehensive understanding of the complex mechanisms associated with TBI and develop appropriate treatment modalities, the use of pre-clinical models may be a useful strategy.
Neuromodulation is an evolving field of medical technology through which the functioning of the central, peripheral, or autonomic nervous systems may be reversibly altered using electrical, chemical, or mechanical principles. 7 Non-invasive neuromodulation strategies investigated in patients with TBI include transcranial magnetic stimulation (TMS) 8 and transcranial direct current stimulation (tDCS). 9 The TMS utilizes a head coil to generate a magnetic field, and subsequently electrical currents, that modulate neuronal activity in underlying brain structures. 10,11 During tDCS, direct current stimulation is delivered to induce a subthreshold modulation of neuronal excitability. 11
Invasive neuromodulation techniques proposed for the management of TBI include deep brain stimulation (DBS) and vagus nerve stimulation (VNS). The former involves the use of electrical stimulation delivered to the brain parenchyma via implanted electrodes. 12,13 VNS involves the electrical stimulation of the vagus nerve to modulate physiological processes and brain regions receiving associated projections. 14
To date, no article has systematically addressed the efficacy of neuromodulation interventions in pre-clinical models of TBI. This review was conducted to answer whether neuromodulation strategies (tDCS, TMS, DBS, and VNS) improve TBI-induced behavioral deficits in pre-clinical models. In addition, we aim to summarize the mechanisms involved in the therapeutic effects of neuromodulation strategies. For each of the included studies, the animal species, intervention, outcome (i.e., post-treatment compared with pre-treatment or to outcome observed in sham-treated groups) and behavioral results will be provided.
Methods
Search strategy
This systematic review followed PRISMA guidelines (Fig. 1). A literature search was conducted using the PubMed and Cochrane databases until March 31, 2022 for original articles with no limit for publication date. Studies were required to match the search terms “traumatic brain injury AND deep brain stimulation,” “traumatic brain injury AND transcranial magnetic stimulation,” “traumatic brain injury AND transcranial direct current stimulation,” or “traumatic brain injury AND vagus nerve stimulation.”
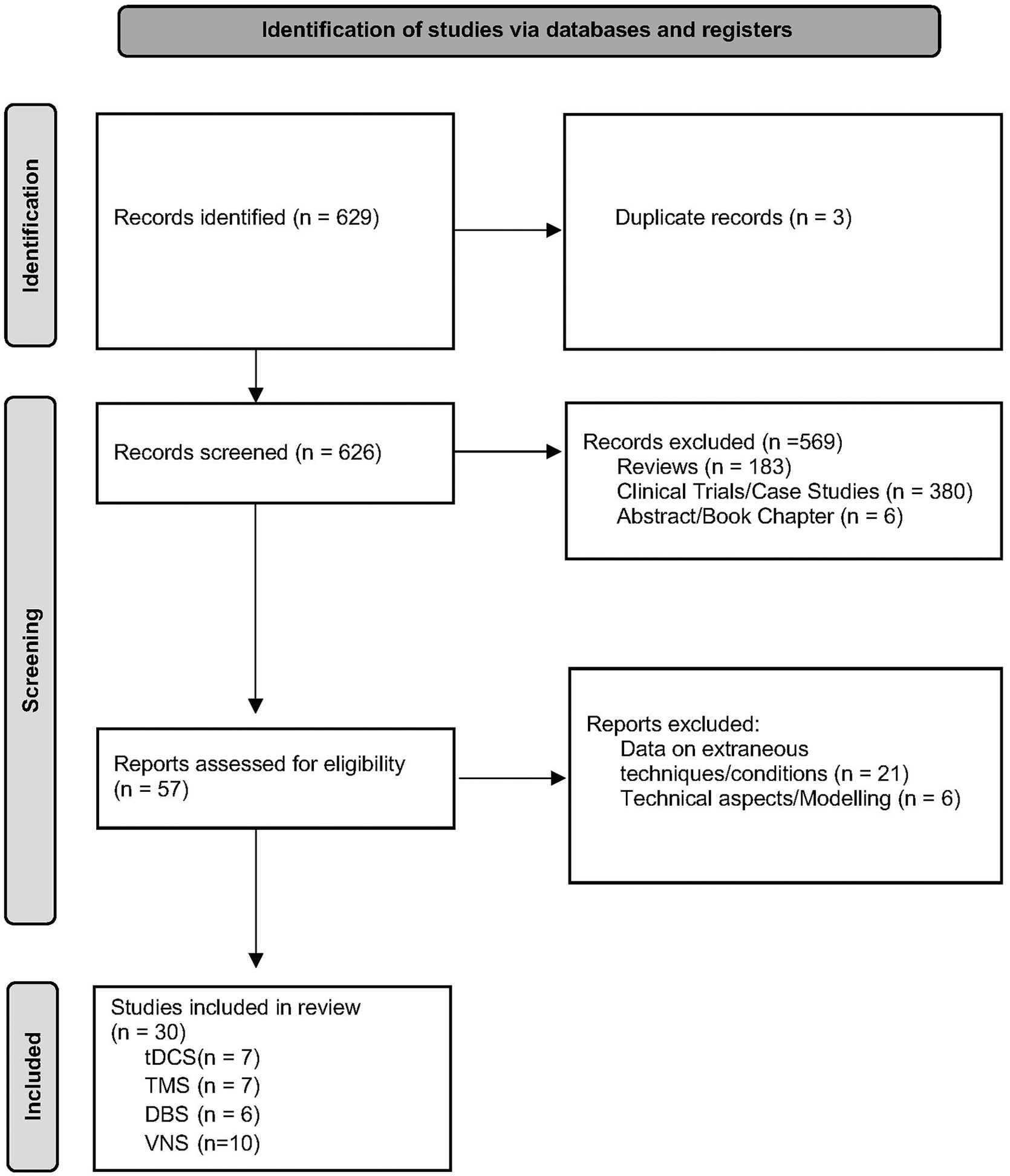
Prisma flowchart of study selection. DBS, deep brain stimulation; tDCS, transcranial direct current stimulation; TMS, transcranial magnetic stimulation; VNS, vagus nerve stimulation.
Abstracts were reviewed by two independent investigators. Relevant articles selected for full-text review had to present pre-clinical data and describe behavioral outcomes or mechanistic data. Excluded papers were reviews, clinical trials, case series, and articles that presented data on other conditions, techniques, technical aspects, modeling, or abstracts. Only studies written in English were included.
Data extraction
Data were extracted to identify common and specific variables. Common variables included species, sex, TBI severity, TBI model, cortical region injured, behavioral output, neurochemical and electrophysiology measures. Variables recorded for each technique included stimulation parameters, time frame, and stimulated target.
Results
The search retrieved 629 abstracts, 30 of which were pooled for analysis. The remainder were excluded for the following reasons: three articles were repeated in more than one search, 183 were reviews, 380 clinical trials or case series, six were abstracts or book chapters, 21 presented data on other conditions or techniques, six were related to technical aspects or modeling. Of the 30 articles retrieved, seven were on tDCS, seven on TMS, six on DBS, and 10 on VNS.
tDCS
We reviewed seven studies investigating tDCS to ameliorate motor deficits, memory performance, and impulsivity in rodent models of TBI, two in mice and five in rats. 15 –21 Four studies were conducted after moderate, 15,16,19,20 two after mild, 17,18 and one after severe injury. 21 The models used were controlled cortical impact (CCI) (n = 3), 15,20,21 weight drop (n = 3), 16 –18 and fluid percussion injury (FPI) (n = 1). 19 TBI was administered either to bilateral cortical regions near the midline (n = 3) 17,18,21 or unilaterally to a single hemisphere (n = 4). 15,16,19,20
Depending on the number of sessions and the time frame after TBI, tDCS was delivered in four major paradigms: short after TBI (minutes to hours) as an acute intervention (minutes to one day) (n = 2), 17,18 a few days after TBI as a chronic intervention (several days to weeks) (n = 1), 16 as an acute intervention long after TBI (several days to weeks) (n = 1), 20 or as a chronic intervention long after TBI (n = 3). 15,19,21
In the most commonly used preparation, the anode was fixed onto the skull over the cortex near the site of injury, while the cathode was attached to the trunk. 15 –20 One study investigated the use of cathodal stimulation with the cathode placed near the site of injury to induce cortical inhibition. 21 As described in Table 1, the current amplitude in studies delivering anodal stimulation ranged from 0.1 mA–0.2 mA. 15 –20 Cathodal tDCS was administered at 0.8 mA. 21 Stimulation was delivered from a few days to four weeks.
Summary of Transcranial Direct Current Stimulation Studies
TBI, traumatic brain injury; M, male; CCI, controlled cortical impact; ML, medial-lateral; AP, anterior-posterior; NS, non-statistical; FPI, fluid percussion injury; BDNF, brain derived neurotrophic factor.
Relative to bregma unless otherwise specified.
When compared with animals with TBI.
tDCS was found to reverse or ameliorate TBI-induced deficits in motor coordination, 6 as measured in tests such as the rotarod performance task (RPT). 15,16,18,19 Anodal stimulation delivered to CCI- or FPI-exposed rodents increased the latency to fall compared with non-stimulated animals. 15,19 In rats exposed to repetitive mild TBI, a single tDCS session was shown to improve RPT 18 and reduce the recovery time of coordination deficits (e.g., the ability of an animal to right itself from a supine to a prone position). 17
Additional impairments after TBI include deficits in working memory, spatial memory, and impulsivity. 5 In a Y maze test, stimulation delivered chronically to CCI- or weight–drop- exposed rodents was found to improve TBI-induced working memory deficits. 15,16 In animals undergoing FPI, tDCS 19 was associated with a significantly shorter latency to locate an escape box in the Barnes maze, suggesting an improvement in spatial memory performance. In a five-choice serial reaction time task, cathodal stimulation was found to significantly reduce impulsive-like responses. 21
In addition to behavioral readouts, a few reports have used electrophysiology and neurochemical tools to investigate potential mechanisms of tDCS in TBI models. In a repetitive mild weight-drop TBI preparation, rats were delivered a single 30-min session of tDCS over the left primary motor cortex. No significant differences were found between stimulated and non-stimulated rats in the amplitude or latency of motor evoked potentials (MEP) before TBI, immediately after, or 1 h after tDCS. tDCS-treated rats, however, but not animals given sham stimulation, were able to recover post-traumatic decreases in MEP amplitude. 18 A similar study found that the peak-to-peak MEP amplitude increased after anodal tDCS compared with pre-treatment baseline. 17
As for neurochemistry, animals receiving tDCS after moderate TBI had an increase in c-fos expression in stimulated cortical regions. 16 In addition, tDCS applied to the perilesional cortex induced a significant increase in the expression of brain derived neurotrophic factor (BDNF). 19 A similar effect was not found in the underlying hippocampus, suggesting that this therapy induces focal neuroplastic responses.
In summary, tDCS delivered during single or multiple sessions mitigates TBI-induced deficits in motor coordination, memory performance, and impulsivity. From a mechanistic standpoint, a series of studies suggested that tDCS may counter the impairment in brain electrical activity induced by repetitive mild TBI, increase neuronal activity, and potentially induce neuroplastic changes in stimulated cortical regions.
TMS
We reviewed seven studies using TMS to manage TBI-induced deficits in motor coordination, balance, locomotor activity, and memory performance, six in rats and one in mice. 22 –28 The models used were weight drop (n = 4), 22,24,27,28 CCI (n = 2), 23,25 and FPI (n = 1). 26 The TBI was moderate in four studies, 23 –26 severe in two, 22,27 and mild in one. 28 Lesions were unilateral in six studies 22 –26,28 and bilateral in one. 27 The three major study designs in the articles pooled for review involved the delivery of TMS as a subacute intervention (for a few days), short after TBI (n = 3), 22,23,28 a few days after TBI as a chronic intervention (n = 2), 24,27 or as a chronic intervention long after TBI (n = 2). 25,26
In the most commonly used configuration, TMS was delivered non-invasively with a probe external to the cranium targeting either the entire cortex or specific cortical regions. Five TMS studies used a stimulation system consisting of a figure-of-eight magnetic coil. 22 –26 One study used an oval-shaped magnetic coil, 29 and one study did not specify the exact configuration of the coil. 28 In most studies, TMS was delivered in bursts or in a repetitive fashion for four days to four weeks. Stimulation parameters are summarized in Table 2.
Summary of Transcranial Magnetic Stimulation Studies
TBI, traumatic brain injury; M, male; CCI, controlled cortical impact; BrdU, bromodeoxyuridine; ML, medial-lateral; AP, anterior-posterior; NS, non-statistical; FPI = fluid percussion injury; GFAP, glial fibrillary acidic protein; NeuN, neuronal nuclear protein; PrPc, cellular prion protein.
Relative to bregma unless otherwise specified.
When compared with animals with TBI.
TMS applied to the cortex of rodents exposed to TBI increased or had a tendency to increase the latency to fall in the RPT, coordination in the beam balance test, and fine motor coordination in the single pellet reaching task. 27,28 Stimulation delivered in the presence of an enriched environment improved TBI-induced impairments in fine motor coordination. 23
TMS administered immediately after repetitive mild TBI improved memory performance in the novel location recognition task, as well as the distance traveled, number of total squares crossed, and a decrease in immobility in the open field. 28 In a pediatric TBI model, TMS delivered to the non-injured primary somatosensory cortex for four weeks from post-neonatal days 16–17 reduced hyperlocomotion activity. 25
In addition to behavioral investigations, a few studies have used electrophysiology and functional imaging techniques to explore mechanisms of TMS in TBI models. In an fMRI study, TMS applied to the primary somatosensory cortex contralateral to the injury side significantly increased cortical responses (189%), synaptic activity (46%), and neuronal firing (200%) in the stimulated hemisphere in comparison to animals that did not receive TMS. 25 Animals receiving TMS in an enriched environment after severe TBI exhibited significantly higher local field potentials, enhanced motor evoked potentials in the ipsilateral forelimb, an increase in cortical excitability, and a higher activation of the primary somatosensory cortex during fMRI compared with animals with TBI that did not receive TMS. 23
Neurochemical analyses in animals treated with TMS have shown a significantly greater number of Bcl-2 cells and a reduced number of BAX cells around the perilesional cortex compared with non-stimulated rodents with TBI. 26 TMS delivered for two weeks has also been associated with augmented cell proliferation and neurogenesis in the subventricular zone, as revealed by an increased number of BrdU-positive and musashi1 positive cells. 24
In addition to neuronal precursors, TMS increased NeuN-positive mature cell density in perilesional cortical zones. 24,28 TMS also reduced the increased caspase-324 and glial fibrillary acidic protein (GFAP) expression 28 observed after TBI, suggesting antiapoptotic and anti-astrogliotic effects. Finally, TMS was shown to modulate cellular prion protein (PrPc) and reduce the expression of proteins involved with the circadian rhythm (CLOCK, CRY2, PER2, GAPDH), 28 while increasing c-fos in the stimulated cortex. 27
In summary, TMS delivered to rodents in regimens that varied from a few days to four weeks countered TBI-induced deficits in coordination, balance, locomotor activity, and memory. Mechanisms involved in the effects of TMS include neuroplasticity, the restoration of abnormal neural circuitry functioning, and possibly changes in regional blood flow.
DBS
We reviewed six studies investigating the effects of DBS in TBI models, particularly focusing on motor coordination, memory, and bladder dysfunction. 29 –34 Five studies were conducted in rats 29 –33 and one in mice. 34 As for other techniques, the degree of TBI ranged from mild (n = 1), 29 to moderate (n = 2), 31,32 and severe (n = 3) 30,33,34 TBI was induced using either FPI (n = 3) 29,31,32 or weight drop (n = 3). 30,33,34 Lesions were unilateral in five studies 29 –32,34 and bilateral in a weight-drop model report. 33
The effects of DBS after TBI were examined in three major paradigms: acute intervention long after TBI (n = 4), 31 –34 chronic intervention long after TBI (n = 1), 29 acute intervention short after TBI (n = 1). 30 The most common configuration was the administration of cathodic stimulation via electrodes stereotactically implanted in different brain targets. The DBS parameters were quite variable, ranging from 7.7–200 Hz, 20–200 μA, and 150 μsec–1 msec (Table 3).
Summary of Deep Brain Stimulation Studies
TBI, traumatic brain injury; M, male; FPI, fluid percussion injury; ML, medial-lateral; AP, anterior-posterior; Iba, Ionized calcium binding adaptor molecule 1; GFAP, glial fibrillary acidic protein; TNFα, tumor necrosis factor alpha; IL, interleukin; CCL, chemokine (C-C motif) ligand 2; CaMKIIα, calcium/calmodulin-dependent protein kinase II; PSD-95, postsynaptic density protein 95; NMDAR, N-methyl-D-aspartate receptor; CDK5, cyclin dependent kinase 5; FJC, fluoro-jade C; TUNEL, terminal deoxynucleotidyl transferase dUTP nick end labeling; F; female; OX1R = orexin receptor type 1; α1-AR, alpha-1 adrenergic receptors; GABABR, gamma-amino butyric acid receptor, type B.
Relative to bregma unless otherwise specified.
When compared with animals with TBI.
Stimulation delivered unilaterally to the lateral cerebellar nucleus (LCN) of rats (12 h/day during four weeks) contralateral to the TBI hemisphere increased dexterity, motor coordination, and was associated with a greater use of the predominant forepaw compared with sham-treated controls. 29 Mice given a single session of central thalamic DBS after severe TBI had an increase in horizontal activity and the total distance travelled in their home cage compared with pre-stimulation values. 34
To manage TBI-induced memory deficits, stimulation was applied to the medial septal nucleus (MSN) for three consecutive days during the Barnes maze test. Stimulated animals, particularly those receiving theta frequency DBS (e.g., 7.7 Hz), presented significantly shorter latencies to locate the escape box and an improved search strategy. 31 Theta frequency MSN DBS was also shown to increase the time exploring novel objects in an exploration task. 32
One of the studies pooled for review specifically investigated the effects of DBS on abnormal bladder functioning. Stimulation applied to the pedunculopontine tegmental nucleus (PPT) four weeks after severe TBI significantly increased the amplitude and duration of vesical contractions, the intercontraction interval, and the void volume compared with non-stimulated controls with TBI. 33
To investigate potential mechanisms of DBS in TBI, both electrophysiology and neurochemistry studies have been conducted. Stimulation delivered to the lateral hypothalamic area (LHA) after TBI induced a significant reduction in δ power, an activity band associated with comatose states. 30 Hippocampal activity recorded within one week of TBI showed that MSN DBS increased theta oscillations. 31,32
LCN DBS was found to reduce Iba1, GFAP, tumor necrosis factor (TNF)α, interleukin (IL)-1β, CCL2 levels, fluoro-jade C (FJC)+, and terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL)+ cells, while increasing the expression of proteins associated with potentiation and excitability in perilesional cortical regions (e.g., calcium/calmodulin-dependent protein kinase II (CaMKII)α, post-synaptic density protein 95 (PSD-95), NMDAR, N-methyl-D-aspartate receptor (NMDAR)1, cyclin dependent kinase 5 (CDK5), and c-fos). 29 LHA DBS was also shown to increase the expression of orexin type 1 (OX1R) and alpha-1 adrenergic receptors (α1-AR), but reduce gamma-aminobutyric acid type B receptor (GABA-B R) through the OX1R pathway. 30
In summary, DBS delivered to the LCN and central thalamus was found to counter TBI-induced deficits in motor coordination, while MSN theta frequency stimulation improved memory performance. PPT DBS improved vesical function. The effects of DBS were likely because of changes in brain activity patterns, synaptic plasticity, or neuroprotective responses, via reductions in apoptosis, neuroinflammation, or neuronal loss.
VNS
We reviewed 10 articles studying the effects of VNS on the behavioral recovery of balance and coordination in TBI models, as well as its neurochemical mech-anisms. 35 –44 Seven studies were completed in rat models, 35 –37,39 –41,43 two in mice, 38,44 and one in rabbits. 42 TBI was delivered either by weight drop (n = 5), 37 –39,43,44 FPI (n = 3),35.36.40 CCI (n = 1), 41 or explosive blast (n = 1). 42 Injury severity varied from moderate (n = 5) 35,36,38,40,41 to severe (n = 3) 37,42,44 and the induction of coma (n = 2). 39,43 Brain injuries were targeted unilaterally in all studies.
VNS therapy was explored in three major treatment paradigms: acute or subacute intervention short after TBI (n = 7), 36 –39,42 –44 chronic intervention short after TBI (n = 2), 35,40 or chronic intervention long after TBI (n = 1). 41 In most studies, VNS was delivered via a cuff electrode implanted in the cervical vagus nerve. Stimulation parameters ranged from 5–30 Hz, 0.5–2 mA, and 0.5–5 msec (Table 4).
Summary of Vagus Nerve Stimulation Studies
TBI, traumatic brain injury; M, male; FPI, fluid percussion injury; ML, medial-lateral; AP, anterior-posterior; GAD, glutamic acid decarboxylase; MDA, melanoma differentiation-associated gene; IL, interleukin; TNFα, tumor necrosis factor alpha; GSH, glutathione; TUNEL, terminal deoxynucleotidyl transferase dUTP nick end labeling; OX1R, orexin receptor type 1; NMDAR, N-methyl-D-aspartate receptor; 5-HT2AR, serotonin receptor 2; H1R, histamine receptor 1; α1-AR = alpha-1 adrenergic receptors; GABABR, gamma-amino butyric acid receptor, type B; F, female; CCI, controlled cortical impact; AQP-4, aquaporin-4; BBB, blood–brain barrier.
Relative to bregma unless otherwise specified.
When compared with animals with TBI.
Both acute 36 and chronic 40 left vagus nerve stimulation were found to significantly improve balance and coordination deficits in rats. 40 Significant improvements were also reported in an isometric pull task, particularly when stimulation was paired with rehabilitative training. 41
Most articles investigating mechanisms of VNS in TBI have focused on neurochemical changes. Some studies showed that acute VNS delivered shortly after injury induced neuroprotective effects. These were characterized by the suppression of TBI-induced increases in proinflammatory cytokines, such as IL-1β, IL-18, IL-6, and TNF-α, and an increase in focal levels of the anti-inflammatory cytokine IL-10, and ghrelin. 37,38,42 VNS has also been shown to inhibit apoptosis, with significant reductions in TUNEL and Bax cells, and increases in Bcl-2 levels being reported. 37
Studies investigating the effects of VNS in comatose animals reported that stimulation increased the expression of oxidation resistance 1 (OX1R), N-methyl-D-aspartate receptor (glutamatergic receptor, NMDAR1), 5-Hydroxytryptamine Receptor 2A (serotoninergic receptor, 5-HT2AR), histamine 1 receptor (H1R), α1-AR receptors, while decreasing GABA-BR. 39,43 The wake-promoting impact of stimulation was diminished in animals administered with orexin antagonists. 43
In addition, VNS was found to mitigate the decrease in glutamic acid decarboxylase (GAD) positive cells 35 and the increased blood–brain barrier (BBB) permeability after TBI. This latter effect was implicated from a decrease in the upregulation of aquaporin 4 (AQP-4), a key protein for the development of BBB-mediated edema. This suggests that VNS may possibly reduce vasogenic elements and cytotoxicity. 44
In summary, VNS was shown to significantly improve deficits in balance, coordination, and motor strength after TBI. This therapy was also found to improve consciousness by modulating putative excitatory and inhibitory neurotransmitter receptors. Finally, VNS delivered shortly after TBI has showcased neuroprotective and anti-apoptotic effects.
Discussion
Studies pooled for review in our manuscript have shown that tDCS, TMS, DBS, or VNS administered in varying schemes after TBI in pre-clinical models mitigate deficits in coordination, balance, and locomotor activity. In addition, these neuromodulatory techniques have also shown therapeutic effects for TBI-related cognitive impairments through improved memory, learning, and impulsivity. Neurochemical analyses provided insight to potential mechanisms related to neuroplasticity, neurogenesis, neuroprotection anti-inflammatory and anti-apoptotic effects, and the reestablishment of normal neural circuitry activity.
Non-invasive neuromodulation
The reviewed studies exploring tDCS as a form of treatment in different rodent models have demonstrated its ability to restore post-injury behavioral and cognitive impairments. Problems with coordination are among the most common motor functioning deficits in rodents and patients after TBI. 6 Studies implementing tDCS acutely or chronically showed a significant recovery in different rodent behavioral tasks. 15,17 –19 In addition, tDCS was found to improve TBI-induced working and spatial memory deficits, 15,16,19 as well as elevated motor impulsivity. 21
In the studies reviewed, potential mechanisms of tDCS in TBI models include a focal increase in cerebral blood flow and oxygenation, changes in cortical excitability, and an increase in neuroplasticity. tDCS may impact cerebrovascular reactivity by retrieving dysfunctional endothelial nitric oxide synthase in the traumatized brain, thus producing more nitric oxide and improving the dilation of cerebral arterioles and cerebrovascular reactivity regulation. 15,20 This would allow an appropriate regulation of blood flow and oxygenation to the injured brain according to its metabolic demands.
Studies also suggest that tDCS is able to alter cerebral excitability, which could promote the recovery of motor and cognitive deficits via an increased synchrony of cortical firing and potentiation. 17,18,21 It is also believed that tDCS is able to promote plasticity in the injured brain by upregulating the expression of neurotrophic factors, such as BDNF. 19,21
Most of the clinical trials investigating tDCS for post-concussive symptoms after mild-moderate TBI have focused primarily on cognitive impairment, with mixed results. 45 Although one non-randomized single blind trial demonstrated sustained improvements in category fluency, two double-blind randomized controlled trials (RCT) revealed no significant differences in cognitive outcomes between active and sham stimulation groups. 45 The use of tDCS for other post-concussive symptoms is limited, with only one retrospective cohort study reporting significant improvements in patients with post-traumatic headache. 45 tDCS has also been explored for cognitive impairment after severe TBI, with similarly mixed results. 46,47
Preliminary evidence suggests that patients in minimally conscious states may respond better than those in vegetative states, with stimulation of prefrontal areas eliciting a better response than parietal areas. 48,49 Response to tDCS in moderate-severe TBI has also been reported and seems to depend on the structure of white matter within a stimulated brain network, highlighting the need for individualized approaches that incorporate neuroimaging and neuronavigation techniques. 50
Studies investigating TMS in TBI models primarily showed improvements in balance and coordination, fine motor control, and decreased hyperactivity. Of the studies reviewed, three of four found significant improvements in balance and coordination after TMS. 23,27,28 One study was not able to detect the same benefits when stimulation was administered one day after TBI, suggesting that the time frame for initiating this therapy may be of importance. 26
As in pre-clinical models, fine motor control dysfunction has also been reported in patients with TBI. 51 Applying TMS in TBI subjects was associated with a certain regain in fine motor coordination, 23,27 suggesting a potential role for this therapy in motor rehabilitation. In a single case, TMS delivered to the cerebellum of a patient with TBI showcased significant motor recovery in balance and coordination. 52 Motor hyperactivity is another behavioral deficit observed in TBI models. 53,54 One study in our review showed that TBI-exposed animals receiving stimulation had significantly reduced hyperactivity compared with non-stimulated controls, 25 suggesting that TMS may be beneficial for the management of this behavioral deficit.
The underlying mechanisms of TMS in TBI are still under investigation. When used during functional recovery, one of these mechanisms may be an increase in cortical excitability. 23,25,27 The enhanced neuronal activity reported after TMS is believed to be associated with gene expression modifications that may help to overcome the hypoactivity observed in injured brain sites, which may be important for the restoration of normal functioning. 25,27 Other studies have suggested that TMS may assist in the reorganization of neural circuits and rhythms that are disrupted after TBI. 22,28 TMS may also help to re-establish more physiological patterns in abnormal neural pathways and circuits, thereby improving motor impairments.
In the clinic, the efficacy of TMS has limited preliminary evidence for post-concussive depression and headache. 55 The only available RCT exploring TMS or post-concussive cognitive impairment reported promising results but was limited by the lack of follow-up data. 56 Seven studies included in a recent review assessed cognition as a secondary outcome in patients with mild TBI. Six of those studies either found no significant improvements or no significant differences between sham and active stimulation. 55 All trials involved stimulation of the dorsolateral prefrontal cortex (dlPFC), either excitatory stimulation on the left (≥ 10 Hz), or inhibitory on the right (≤ 1 Hz). Other specific protocol parameters varied widely across trials. Moreover, a recent double blind RCT found TMS over the left dlPFC to be ineffective for post-concussive cognitive impairment after moderate TBI. 57
Electroconvulsive therapy (ECT) is an effective non-invasive neuromodulation modality for patients with depression and psychotic episodes. 58 Despite a few clinical reports showing efficacy in patients with post-traumatic depression and psychosis, 59 –63 we could not find pre-clinical studies using ECT to manage TBI-induced behavioral deficits searching the terms “traumatic brain injury and electroconvulsive therapy.” As a result, this neuromodulation treatment was not included in our review.
Invasive neuromodulation
Studies applying DBS neuromodulation showcased its ability to recover motor deficits, locomotor activity, coordination, and paw dexterity after TBI. 29,34 DBS was also associated with significant improvements in spatial memory. 31,32 From a translational perspective, these results suggest that DBS could be potentially investigated in the recovery of both motor and cognitive dysfunction in patients with TBI.
The mechanisms of DBS neuromodulation are still not fully understood. In a simplistic fashion, DBS has been proposed to reduce spontaneous neuronal firing while activating fiber pathways. 64 A commonly proposed mechanism for the former involves a progressive depolarization state that culminates with a block of spontaneous action potentials. 64 –66 By driving neuronal appendages, DBS has been proposed to exert some of its distant effects and increase the release of GABA from interneurons in the target region. 64,67 In TBI, some of these effects may be responsible for the reduced neuronal activity in the stimulated area, which may culminate in neuroprotective and antiapoptotic effects. 29,30,68
In addition, an enhanced cortical excitability may reverse the neurodegeneration-induced hypoexcited state associated with traumatic injuries. 29 An alternative mechanism of DBS in TBI involves the restoration of abnormal neural oscillations, including abnormal theta wave patterns, that may disrupt the normal functioning of brain circuits involved in cognitive performance. 31,32 Also noteworthy are the increase in plasticity and potentiation after DBS. 29
A few recently published pre-clinical studies were identified after our literature search was completed. One study showed that LCN DBS was associated with an improvement in spatial and recognition memory tasks in TBI-exposed rodents. 69 Neurochemical analyses showcased significant increases in CaMKIIα, BDNF, and p75NTR in the perilesional cortex. 69 Another study revealed that task-matched acute DBS applied to the nucleus accumbens in mice enhanced cognitive abilities in visuospatial associative learning and long-term memory in the Morris water maze. 70
In pre-clinical TBI models, VNS has been employed mainly in the recovery of coordination and balance deficits. These studies provide behavioral insight into the potential translational use of this therapy in the restoration of motor function. 36,40,41 In addition to behavioral improvements, neurochemical studies have been conducted to ascertain potential mechanisms of VNS. One such mechanism is stimulation-induced neuroprotection. The application of VNS for a short period soon after the brain injury has been shown to suppress proinflammatory cytokines and proapoptotic factors, while increasing the expression of anti-inflammatory cytokines and anti-apoptotic factors. 37,38,42
In TBI-induced coma studies, the wake-promoting effects of VNS seem to occur through the upregulation of excitatory and the reduction in the expression of inhibitory neurotransmitters. 39,43 Specifically, the increased expression of orexins may be involved in the restoration of consciousness after VNS. 39,43
In the clinic, a recent review of DBS for the treatment of those with disorders of consciousness and cognition after severe TBI revealed mixed results. 71 Outcomes focused on either arousal (after stimulation of the ascending reticular activating system or associated thalamic regions) or cognition (DBS delivered to the frontal and prefrontal cortices, fornix, nucleus accumbens, internal capsule, thalamus, or brainstem nuclei).
Across six studies exploring DBS for arousal (n = 73, mostly patients in a vegetative state), approximately 45% of individuals improved. 71 In a six-month double-blind alternating crossover case report, a patient with minimally conscious state had an improvement in behavioral responsiveness after bilateral central thalamus DBS. 72 This suggests that DBS may be effective for disorders of consciousness, particularly when the circuitry involving the midbrain reticular formation, thalamus, and cortex is found to be relatively preserved. 71
Modest improvements in cognitive outcomes (executive function, impulsivity, and initiation of goal-directed behavior) were reported in a small case series (n = 4) of patients who regained consciousness after severe TBI after DBS of the nucleus accumbens and anterior limb of the internal capsule. 73 Notably, functional and cognitive scores improved more substantially when DBS was paired with behavioral therapy compared with DBS alone. 73
Current evidence in humans for implanting VNS devices in TBI is limited to a single case report. After stimulation, the arousal level of a patient who had been lying in a vegetative state for 15 years improved to a minimally conscious state over the course of six months. 74 Coinciding with this change in arousal, VNS had increased activity in occipito-parieto-frontal and basal ganglia regions, as well as metabolic signals in the thalamus. 74 More recently, transcutaneous VNS (a non-invasive method) was applied to five patients with severe TBI in an open-label, prospective, single-arm study. It is unclear whether the modest improvements in measures of consciousness in three of these patients may be attributed entirely to tVNS, but the study did demonstrate the safety, tolerability, and feasibility of this neurotechnology in severe TBI. 75
Translational considerations
Overall notable gaps remain in our knowledge of how neurostimulation actually modulates the brain in TBI. Animal models allow for more detailed probing of these complex mechanisms, along with treatment manipulations across types of injuries and time/stages of TBI (e.g., acute, subacute, and chronic). 76 Invasive neuromodulation strategies for more severe TBI in human populations are still in their infancy. Under these circumstances, the use of pre-clinical models may offer a translational opportunity for new potential treatment targets.
While not included in this review, a recent study exploring DBS in nonhuman primates with severe TBI found improvements in cognitive fatigue and arousal after stimulation of the central thalamus and medial dorsal thalamic tegmental tract (connecting the prefrontal and thalamic structures). 77 In addition, stimulation of the midbrain raphe nuclei in rodent models of severe TBI demonstrated improvements in cognition and motor coordination, implicating dysregulated serotonergic functioning as a potential pathophysiological and neuroanatomical target for DBS. 78 Other targets not yet explored in pre-clinical models would include stimulation of disrupted white matter tracts connecting frontal, thalamic, and midbrain structures. 71
One notable difference between pre-clinical studies and clinical trials is the somewhat better results obtained in the former. In general, the stimulation time frame and settings are relatively similar in studies conducted in both species. One exception, however, is the stimulation current. Considering the differences between electrode dimensions in humans and rodents and the current delivered in pre-clinical and clinical studies, it becomes clear that the charge density related to the stimulation amplitude delivered to rodents is often substantially higher than that administered to humans. It is possible that the administration of higher currents could yield to better clinical results in human trials in general. That said, increasing stimulation settings in humans may lead to adverse effects that may be poorly tolerated.
Another translational shortcoming of pre-clinical research is that many of the target symptoms of human TBI are difficult to model in animals, given their subjective (e.g., depression, headache) and interconnected nature. For example, cognitive impairment after mild TBI is often perpetuated by other common post-concussive symptoms (e.g., headache, depression, insomnia), which makes it difficult to isolate and disentangle in polysymptomatic patients. 79
Human neuromodulation trials in general are moving in the direction of precision and finely localizing targets (e.g., targeted neuroimaging, closed-loop systems). Available coils and stimulation devices in animals may be too large/crude to study similarly targeted stimulation regions (especially in rodent models). Finally, the lack of objective clinical biomarkers for TBI/concussion hampers translational treatment efforts because there are little means to confirm that injuries induced in pre-clinical animals are comparable to human TBI/concussion.
Although these neuromodulation techniques exhibit promising functional and cognitive recovery in pre-clinical models of TBI, there is still uncertainty regarding the ideal stimulation paradigms and parameters. The technique, target, treatment initiation, duration, power, and frequency of stimulation are all important components when considering the therapeutic use of neuromodulation.
Although the studies in this review targeted different brain regions, they ended up producing a similar post-injury recovery in behavioral and cognitive tasks. Moreover, it is still unclear whether neuromodulation is best administered immediately after the injury or as a delayed regimen after the secondary processes of TBI become established. One study suggested that early TMS intervention could be detrimental, because the injured brain may be vulnerable during the early injured state. 26 Other studies, however, suggest that the immediate application of neuromodulation can provide neuroprotective and antiapoptotic effects in the injured brain, which would help to reduce functional loss. 24,28
Conclusion
This systematic review is the first to our knowledge to summarize the effects of tDCS, TMS, DBS, and VNS in pre-clinical models of TBI. It has provided robust and encouraging data for the therapeutic properties of these modalities with a potential for clinical translation. These techniques have demonstrated the ability to recover behavioral impairments in motor and cognitive functioning after TBI. There is also evidence supporting the potential underlying mechanisms for these recovery effects through neuroplasticity, neurogenesis, neuroprotection, and reinstated neural circulatory activity.
Despite limitations, the use of animal models of TBI to study the effects and mechanisms of neuromodulation therapies may be quite valuable. To date, pre-clinical results seem to be more promising than the outcome reported in clinical trials. In future studies, stimulation parameters, timeline, and targets successfully implemented in animals may be translated to the clinic.
Patients with post-concussive syndrome have various neuropsychiatric symptoms. To reflect this scenario, animals exposed to TBI could be tested in a battery of behavioral paradigms (e.g., including tests to measure anxiety, depression-like behavior, memory deficits, impulsivity, and motor coordination). In models of fear- and anxiety-like behavior, response to neuromodulation seems to vary according to the behavioral phenotype of the animals. 80,81 A similar approach may be taken in TBI-exposed rodents to study individual differences in behavior.
Finally, rather than stand-alone therapies, it is more likely that neuromodulation strategies will need to be combined with other therapeutic modalities for an optimal result. Animal models offer the possibility of studying individualized interactions between neuromodulation and medications, as well as rehabilitation regimens.
Footnotes
Authors' Contributions
SS reviewed the literature and contributed with writing. CH conceptualized the study, reviewed the literature, and contributed with writing. AM and MJB contributed with writing. TKR, ACPC, AA and NL reviewed and edited the text.
Funding Information
Research supported by the Canadian Institutes of Health Research and the Harquail Centre for Neuromodulation.
Author Disclosure Statement
No competing financial interests exist.