
Abstract
Spinal cord injury (SCI) above the lumbosacral level often leads to dysfunction of the lower urinary tract (LUT) including detrusor hyper-reflexia, wherein bladder compliance is low, baseline pressures are increased, and filling is accompanied by numerous non-voiding contractions (NVCs) referred to as neurogenic detrusor overactivity. Here, we investigate the expression levels of the serotonin 1A (5-HT1A) receptor in segments both rostral and caudal to the injured site, as well as the effects on micturition of blocking 5-HT1A receptor using pharmacological interventions in spinally intact rats or T8 complete SCI rats. The activities of detrusor and external urethral sphincter (EUS) were assessed with the rats in a conscious condition. Adult female rats were divided into two groups: (1) sham control (T8 laminectomy only) and (2) T8 complete spinal cord transection. The observation period was 2 months after the original SCI. In Western blot analyses, we identified significant upregulation of the 5-HT1A receptor in the T10-L2 and L6/S1 segments after chronic complete SCI. In pharmacological studies, a dose-response study of the 5-HT1A receptor antagonist, WAY100635, indicated alterations in detrusor and EUS activities in spinally intact rats. Interestingly, blocking the 5-HT1A receptor alone resulted in inhibitory effects on NVCs with a reduced number and decreased amplitude, but in an increased interval between NVCs in SCI rats. In addition, the duration of EUS bursting was also significantly increased by WAY100635. These inhibitory effects of WAY100635 on NVCs were diminished by subsequent application of a beta-adrenergic blocker (propranolol). The reduction of NVCs observed by WAY100635 may be the result of blocking the constitutive activities of the 5-HT1A receptor but activating the beta-adrenergic sympathetic pathway, which in turn relaxes bladder activity. Together, the neuroplasticity of the 5-HT1A receptor can be a potential therapeutic target for treatment of bladder dysfunction after SCI.
Introduction
Depending upon the level affected, spinal cord injury (SCI) results in the disruption of motor and autonomic functions with differing consequences for each system, including micturition control. 1,2 It is well established that suprasacral SCI results in neurogenic detrusor overactivity (NDO) caused by a lack of supraspinal inputs regulating voiding reflexes. 2 NDO is characterized by increased frequency of strong involuntary bladder contractions, often resulting in urinary incontinence. NDO development is thought to result from massive neuroplasticity changes occurring in the lumbosacral spinal cord after SCI. One of the proposed mechanisms of NDO is caused by hyperexcitability of C-fiber bladder afferents in sensory nerves. 3 –5 Intravesical therapy with the C-fiber neurotoxin resiniferatoxin can reduce NDO in patients with SCI. 6 Urine storage and release are highly dependent upon central nervous system (CNS) pathways, which are modulated by multiple excitatory and inhibitory neurotransmitters including norepinephrine, gamma-aminobutyric acid, glutamate, serotonin (5-HT), encephalin, and corticotrophin releasing factor. 7 Therefore, specific neurological mechanisms are important considerations in the diagnosis and treatment of voiding disorders.
Along with other parts of the autonomic nervous system; for example, the heart, vasculature, and airways, 8 central 5-HT receptors have been implicated in the control of micturition. 9 There are 14 different structurally-distinct 5-HT receptors, which are divided into seven families (5-HT1–7 receptors), and those receptors that have already been implicated in the control of micturition include 5-HT1, 5-HT2, 5-HT3, and 5-HT7. These receptors seem to modulate all of the pathways involved in the control of micturition – the parasympathetic, the sympathetic, and the somatic 10 – although there are few experimental data on the sympathetic pathway. The 5-HT7 receptor 11 has just been added to this list based on the use of pharmacological interventions with either agonists or antagonists to the 5-HT7 receptor 10 ; but there is very little information regarding its role in regulating micturition control after SCI. In addition, the 5-HT2A/2C receptor agonist (2,5-dimethoxy-4-iodophenyl)-2-aminopropane hydrochloride (DOI) improved voiding efficiency by promoting external urethral sphincter (EUS) burst activity, decreasing outflow resistance, and possibly improving voiding contraction in urethane-anesthetized complete SCI rats. 12 The 5-HT1A receptor is the most plastic and most highly characterized spinal 5-HT receptor, and it modulates a number of different cellular processes in the spinal cord. The 5-HT1A receptor can increase spinal motor neuron excitability pre-synaptically in neonatal rats by modulating the amount of transmitter released, 13 while cell excitability can be decreased post-synaptically by inhibiting adenylate cyclase activity through a Gi protein in fetal mouse spinal cord ganglion explants. 14 In spinally intact rats, 5-HT1A receptor agonists increase and 5-HT1A receptor antagonists decrease urinary frequency through actions at both the supraspinal and spinal levels. 15,16 In addition, the 5-HT1A receptor agonist ([R]-[+]-8-hydroxy-2-di-n-propylamino) tetralin (8-OH-DPAT) can improve voiding efficiency by increasing micturition volume, decreasing residual volume, and increasing the bursting activity of EUS in urethane-anesthetized rats with complete SCI. 17 However, activation of the 5-HT1A receptor by 8-OH-DPAT can significantly suppress reflex bladder activity in both conscious and α-chloralose-anesthetized cats after complete SCI, 18 indicating that there are species-dependent differences in the influence of the 5-HT1A receptor on micturition.
Although the 5-HT receptor family has been investigated in controlling micturition-reflex behavior in different species under normal physiological conditions, 10 there is very little information on 5-HT1A receptor expression levels in the spinal cord (both sympathetic and parasympathetic segments), or on the role of the 5-HT1A receptor in regulating NDO after chronic SCI. Previous studies showed that blocking the 5-HT1A receptor with the 5-HT1A receptor antagonists WAY100635 or NAD-299 inhibited detrusor activity, 15,16,19 but had no effect on EUS activity 17 in spinally intact rats. However, those studies occurred either with the animals awake and did not specifically investigate EUS, 15,16,19 or with the animals under urethane anesthesia, 16,17 which has inhibitory effects on EUS activity. 5,20 In the present study, we determined the spatiotemporal expression patterns and levels of the spinal 5-HT1A receptor at various time points following SCI. In addition, a comprehensive cystometry and EUS electromyogram study was used to determine the effects of pharmacological inhibition of the 5-HT1A receptor on regulating lower urinary tract (LUT) function (both detrusor and EUS activities) in awake spinally intact or SCI animals.
Methods
Animal groups and experimental design
All animals were housed in standard laboratory cages under 12:12-h light–dark cycle conditions with standard rodent chow and water available ad libitum. All experiments were performed during the light cycle. All animal procedures were approved by the Cleveland Clinic Institutional Animal Care and Use Committee. Forty-two adult female Sprague-Dawley rats (220–250 g; Harlan Laboratories, Madison, WI, USA) were randomly divided into two groups: (1) spinally intact group (n = 12) and (2) T8 complete transection group (n = 30). In the spinally intact group total of 12 animals, all 12 were used to pharmacologically test LUT function and then 6 animals were used for Western blot study and the other 6 were used for histological study. In the T8 complete transection group, there were four subgroups at four different time points, including 1 (n = 6), 2 (n = 6), 4 (n = 6), and 8 weeks (n = 18) post-injury. All animals in the 1, 2, and 4 week post-injury groups were used for Western blot study. In the 8 weeks post-injury group, all 18 animals were used to pharmacologically test LUT function by intravenous (n = 12) or intrathecal (n = 6) drug delivery. After assessments of LUT function, 6 of the 12 animals with intravenous delivery were used for Western blot study and the other 6 animals were used for histological study. The spinal cord surgery was performed using ketamine/xylene anesthesia, and LUT functional assessments were prepared using isoflurane surgery and then recovery for 1 h prior to recordings when the animals were awake. All efforts were made to minimize suffering. All procedures were performed in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health.
T8 spinal cord laminectomy and complete transection surgery
Animals had operations performed under general anesthesia using intraperitoneal administration of ketamine (60 mg/kg) and xylazine (10 mg/kg). The body temperature was monitored by a rectal probe and maintained using a heating pad underneath the animal, which automatically adjusted the temperature based on feedback from the rectal probe. A sterile eye lubricant (Paralube) was applied to prevent dehydration. The musculature was cut from T7-T9 following the skin incision and the dorsal surface of the T8 segment was exposed by laminectomy. The spinally intact group did not receive the following injury procedures. For the complete transection group, a complete cut was made by a scissor at T8, resulting a separated gap (2–3 mm) between two stumps. A high-magnification surgical microscope was used to confirm that there was no remaining neural tissue between the two stumps of cord. The overlying musculature was sutured closed and the skin was closed using 4-0 black monofilament suture.
Western blot analyses of spinal cord tissues
The spinally intact animals and animals at four different time points (1, 2, 4, and 8 weeks) post-SCI were subjected to transcardial perfusion with ice-cold phosphate-buffered saline (PBS). The spinal cord rostral to the injury (T5/6), and sympathetic (T10∼L2) and parasympathetic (L6/S1) segments of spinal cord were homogenized in a radioimmunoprecipitation assay (RIPA) buffer supplemented with protease inhibitor and phosphatase inhibitor cocktails (Sigma-Aldrich, St. Louis, MO, USA). All samples were centrifuged, and the supernatant was collected and stored at −80°C until use. Protein concentration was estimated using a BCA protein assay kit (Thermo Scientific, Rockford, IL, USA). Protein lysates were prepared in 4 × Laemmli sample buffer (Bio-Rad, Hercules, CA, USA) and dithiothreitol (DTT) and then incubated at 95°C for 5 min. Equal amounts of protein were loaded into each well (50 μg) of stain-free Novex 4–12% Tris-Glycine Mini Gels, WedgeWell format (Thermo Fisher Scientific, Waltham, MA, USA). After gel electrophoresis, protein samples were transferred to polyvinylidene difluoride (PVDF) membrane (Bio-Rad, Hercules, CA, USA). Blots were blocked in 5% dry milk in tris-buffered saline (TBS) containing 0.1% Tween-20 (TBST) for 1 h at room temperature (RT). Primary antibody (5-HT1A receptor, MAB11041, 1/1000 in TBST containing 5% bovine serum albumen (BSA); Millipore Sigma, Temecula, CA, USA) was incubated overnight at 4°C. After three washes in TBST for 5 min, blots were incubated in secondary antibodies (HRP-conjugated anti-mice IgM, Abcam, Waltham, MA, USA) at RT for 2 h. Chemiluminescence was detected using Clarity Western ECL substrate (Bio-Rad, Hercules, CA, USA), imaged using a Bio-Rad ChemiDoc MP, and analyzed using Imagelab software version 6.1 (Bio-Rad, Hercules, CA, USA) for volumetric analysis of protein expression. 5-HT1A receptor levels were normalized to glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (a loading control, 2118, Cell Signaling Technology, Danvers, MA, USA) as a loading control. All results were normalized by taking the value of the spinally intact group as 100% to obtain a comparative value. The significance was assessed using one-way analysis of variance (ANOVA) followed by the Tukey's post hoc test with Graphpad Prism 8.0 (GraphPad). The 95% confidence level (p < 0.05) was considered statistically significant.
Immunohistochemistry for 5-HT1A receptor and analyses of immunoreactivity intensity
At 8 weeks post-SCI, animals were transcardially perfused with 4% paraformaldehyde (PFA). The spinal cords were removed and stored in 4% PFA overnight, and then transferred to 30% sucrose solution. The L6/S1 segment of spinal cords were sectioned by cryostat in 30 m pieces for an immunohistochemistry process, and the 15 sections (120 m apart) in each animal were selected. The sections were blocked by blocking solution (PBS containing 0.5% Triton X-100 and 3% BSA) and then incubated with antibodies against 5-HT1A receptor (MAB11041, Millipore Sigma) and NeuN (Abcam) overnight at room temperature. All sections were rinsed three times with PBS for 10 min and then incubated at RT for 1 h with secondary antibodies (Abcam) including Alexa Fluor 488-conjugated secondary antibodies and Alexa Fluor 594-conjugated secondary antibodies. The sections were rinsed three times with PBS, and mounting media (Vector Laboratory, Newark, CA, USA) was added before they were cover-slipped. The sections were photographed by a Zeiss confocal microscope for further analysis.
Standardized areas for sampling in sections from each animal in each group were selected by Photoshop and ImageJ. The mean number of pixels containing immunoreactive product in the sampled area was measured and multiplied by the average intensity. This value was subtracted from background immunolabeled intensity, as measured in a separate adjacent section. Graphs were plotted and statistics were assessed using Graphpad Prism 8.0 (GraphPad). The intensities are shown as mean ± standard error of the mean (SEM) in units of the percentage of the mean intensities from spinally intact rats, which are presented as 100%. Significant differences between groups are indicated by a signifier (p < 0.05; p < 0.001) using an unpaired t test. Light intensity and threshold values were maintained at constant levels for all analyses.
Awake urodynamics, EUS electromyography (EMG) recording, and pharmacological interventions
Animals were anesthetized under 2.5% isoflurane with oxygen to perform surgeries and then cystometry and EUS EMG recording were performed when the animals were awake. The urinary bladder was exposed by an incision of the midline abdomen. A catheter (polyethylene 50) was inserted into the bladder through a small incision at the apex of the bladder dome. The bladder end of a pulmonary embolism (PE) catheter was heated to form a collar and was secured with a purse-string suture on the opening of the detrusor. The end of bladder catheter was connected to a pressure transducer (model P23XL-1; Gould Ohmeda, Valley View, OH, USA), an amplifier (Astro-Med, Inc., West Warwick, RI, USA), and an Aladdin single-syringe infusion pump (World Precision Instruments, Sarasota, FL, USA) for signal recording and saline infusion. A normal saline-filled femoral vein catheter was inserted into the femoral vein for drug delivery. The bladder catheter exited from the rostral edge of the abdominal wound, which was closed in layers. The skin above the vascular catheters was also closed. Fine (50μm diameter) Teflon-insulated platinum wire electrodes (A-M Systems, Carlsborg, WA, USA) were inserted percutaneously on both sides of the mid-urethra using a 27-gauge needle for subsequent measurements of EUS EMG activity. Next, rats were placed in a universal restrainer (Braintree Scientific, Inc., MA, USA) to perform physiological tests. Following a 1 h recovery period from anesthesia, continuous cystometrograms (CMGs) were collected using constant infusion (0.1 mL/min) of saline by an infusion pump (World Precision Instruments) through the catheter into the bladder to elicit repetitive voids. The electrodes were connected to an AC amplifier (Astro-Med, Inc.) with high- and low-pass frequency filters at 10 Hz and 1 kHz and a recording system (DASH 8X, Astro-Med, Inc.) at a sample frequency of 10 kHz. Continuous cystometry at a flow rate of 6 mL/h for 1 h prior allowed the LUT to adjust to the flow rate. After 1 h the pump was stopped and the bladder was emptied by aspiration through the bladder catheter via a stopcock port. The pump was restarted and after at least three reproducible filling CMGs were obtained, WAY100635 from Sigma (0.02 mg/kg, 0.2 mg/kg, 1 mg/kg, 2 mg/kg) for spinally intact or WAY100635 (0.2 mg/kg, 1 mg/kg, 2 mg/kg) and propranolol (2 mg/kg) for complete SCI animals were intravenously administered through the right site of femoral vein, and then another filling CMG was performed, from which measurements were taken for three to five filling cycles. Drug solutions were administered in a volume of 0.1 mL followed by a 0.1 mL flush of saline. In addition to the systemic delivery of WAY100635, the intrathecal injection of WAY100635 (0.002 mg/kg, 0.02 mg/kg) with the tip at lumbar segments was used to determine site of action in the SCI animals. For intrathecal catheter preparation, the laminectomy at T10 spine was made to expose the upper lumbosacral segment of spinal cord after midline incision on the back of the skin. A 27 G needle was used to make a small hole in the dura mater and a 32 G intrathecal catheter (CS-32, Braintree Scientific, Inc.) was inserted into the intrathecal space advancing 0.3 cm caudally to approximately reach the L1/L2 spinal segment. The catheter was secured with tissue glue (3M) and the other end of catheter was used for the drug injection during the awake urodynamics and EUS EMG recordings.
Quantification of CMG and EUS EMG recordings
During each of the individual interventions in the CMG and EUS EMG recordings, three to five consecutive and stabilized micturition events were segmented for analysis as described in our previous studies. 21,22 Segmented data included the full duration between the completions of two consecutive voiding contractions. The defined parameters used to analyze both CMG and EUS EMG recoding data are shown in Figure 1. CMG voiding parameters include: the voiding efficacy (%), peak voiding pressure (cmH2O), bladder volume capacity (mL), voiding duration (sec), voiding interval (min), and high-frequency oscillation (HFO) (sec). The period of the voiding cycle from the empty bladder with saline infusion until the first micturition occurred was used to determine voiding efficacy. It was calculated by the percentage of voiding volume in the amount of total saline infused. Non-voiding contractions (NVCs) were determined as contractions with amplitudes >5 cm H2O that were not observed with any successful micturition. The CMG non-voiding parameters include: amplitude (peak pressure – baseline pressure) (cmH2O), single non-voids duration (sec), number of non-voids, and percentage of time with non-void activity during the bladder-filling phase (%). The EMG parameters included (during the bursting phase): the number of EMG bursts, EMG bursting duration (sec), EMG burst interval (sec), and EMG burst amplitude (mV). EMG bursts were qualified as coordinated when they were timed in sync with the CMG HFO. The amplitude of tonic EUS EMG activity during the bladder filling phase also was analyzed. All data collection and data analysis were performed in a blinded fashion. Data are presented as group mean ± SEM, and were analyzed using a one-way ANOVA with a Fisher's post hoc test. Significance level was set to 0.05 for all comparisons.
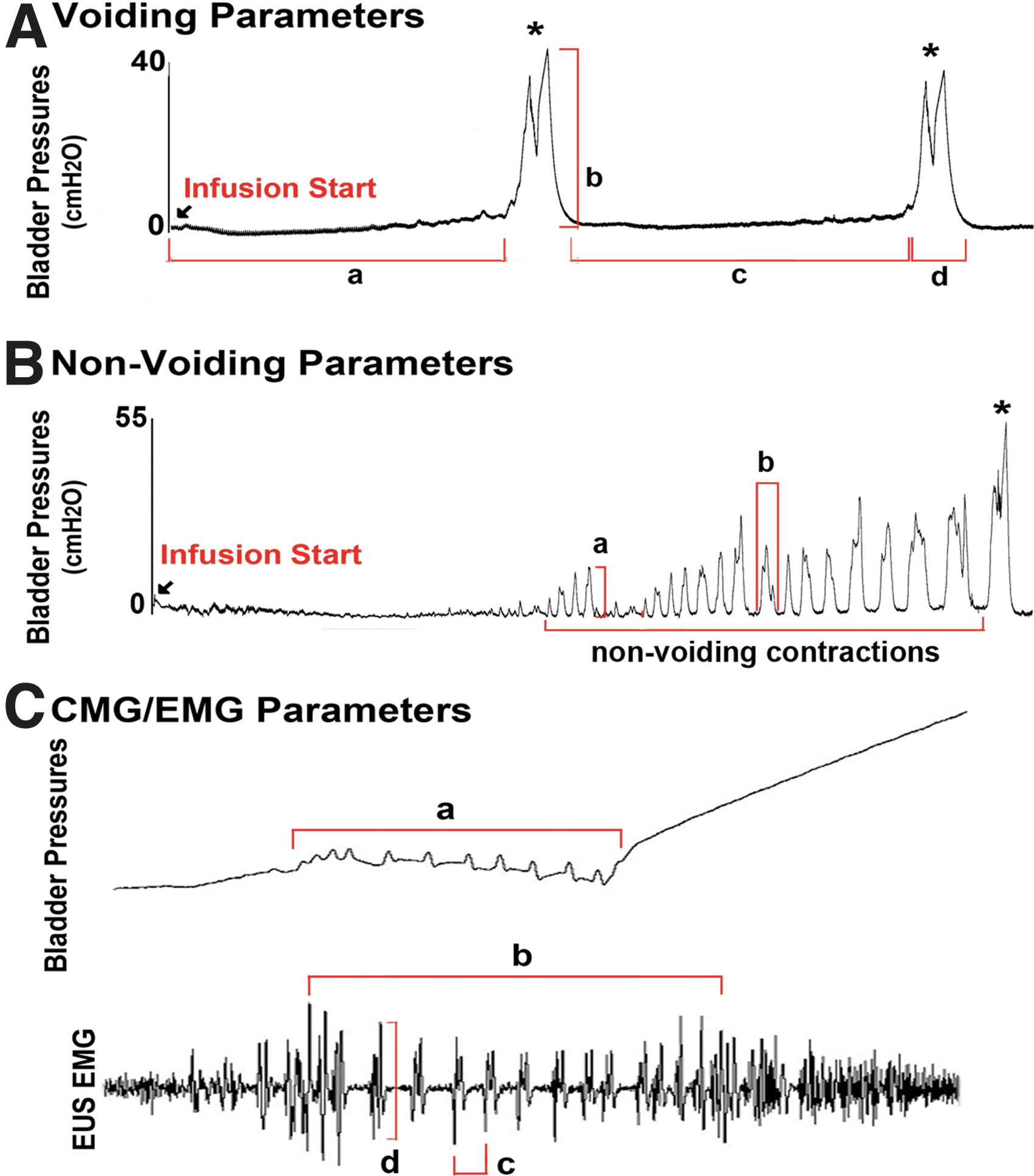
The illustration of parameters of voiding, non-voiding, and electromyography (EMG) in the analyses of cystometrograms (CMG) and external uretheral sphincter (EUS) EMG recordings. (
Results
Upregulation of spinal 5-HT1A receptor after chronic SCI
The expression levels of spinal 5-HT1A receptor at 1, 2, 4, and 8 weeks after T8 SCI in the spinal cord segments both rostral and caudal to the injured site were determined by Western blot analyses. The caudal segments were divided into sympathetic segments (T10-L2) and parasympathetic segments (L6/S1). There were no significant differences in the levels of 5-HT1A receptor rostral to the injured site in SCI animals when compared with the spinally intact animals (Fig. 2A, B). In contrast, there was significant upregulation of spinal 5-HT1A receptor below the injured site over a period of 8 weeks after T8 complete lesion. In the sympathetic spinal cord segment, there was significant upregulation of 5-HT1A receptor starting at 1 week and continuously increasing, even at 4 and 8 weeks post-SCI. These increases were ∼35% (1 week), 42% (2 weeks), 50% (4 weeks), and 58% (8 weeks) following SCI when compared with the spinally intact animals (Fig. 2C, D). Similar to the sympathetic spinal cord segment, the parasympathetic spinal cord segment also showed robust upregulation of 5-HT1A receptor starting at 1 week and continuing through 4 or 8 weeks post-SCI. These increases were ∼30% (1 weeks), 50% (2 weeks), 80% (4 weeks), and 82% (8 weeks) following SCI when compared with the spinally intact animals (Fig. 2E, F).
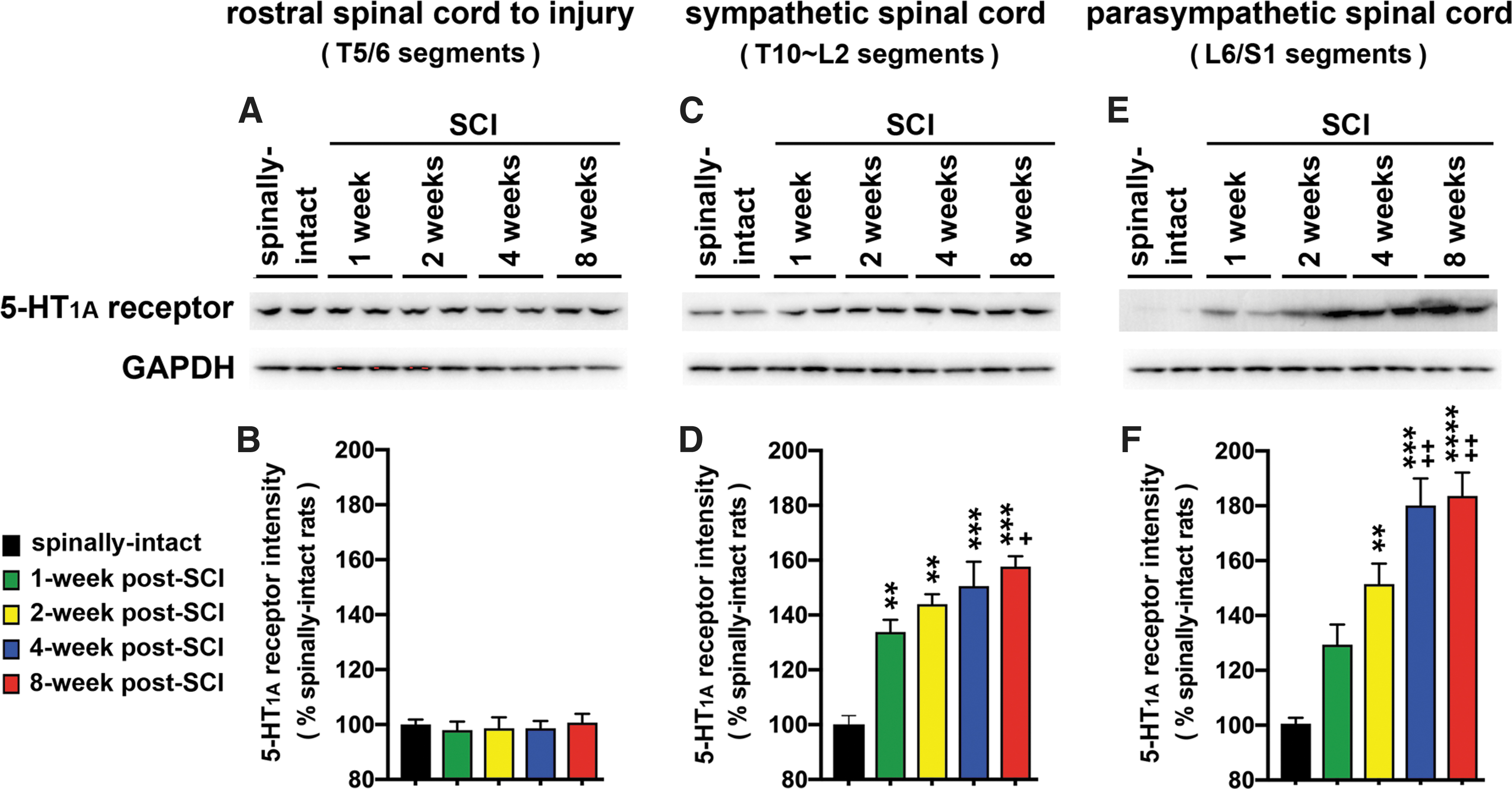
The spatiotemporal expression of the 5-HT1A receptor in the spinal cord after T8 complete spinal cord injury (SCI). At the designated time points after SCI, spinal cords were harvested and total tissue lysate was extracted for Western blot analyses of 5-HT1A receptors. The expression levels of 5-HT1A receptor in both the sympathetic (T10∼L2 spinal segments,
In addition, we further studied the anatomical distribution of upregulated spinal 5-HT1A receptor. The L6/S1 segment of spinal cords from spinally intact animals and SCI animals at 8 weeks post-injury were collected for this study. We analyzed the immunoreactivity of 5-HT1A receptor with double-staining of 5-HT1A receptor and a neuronal marker (NeuN) from laminae I to X of the gray matter (Fig. 3A–L). There was a significant increase in the immunoreactivity of 5-HT1A receptor from laminae I to V, and a trend toward increased immunoreactivity of 5-HT1A receptor in lamina VI, VII, and X. These increases were ∼250% (lamina I), 70% (lamina II and III), and 30% (lamina IV and V) following SCI when compared with the spinally intact animals (Fig. 3M, N). The overall increase in 5-HT1A receptor immunoreactivity is consistent with the outcomes of the Western blot analyses, and supports the upregulation of spinal 5-HT1A receptor in the chronic phase of SCI. These results indicated an upregulation of 5-HT1A receptor in the areas caudal to the injured site, including both sympathetic and parasympathetic segments, which are involved in the regulation of micturition reflex pathways.
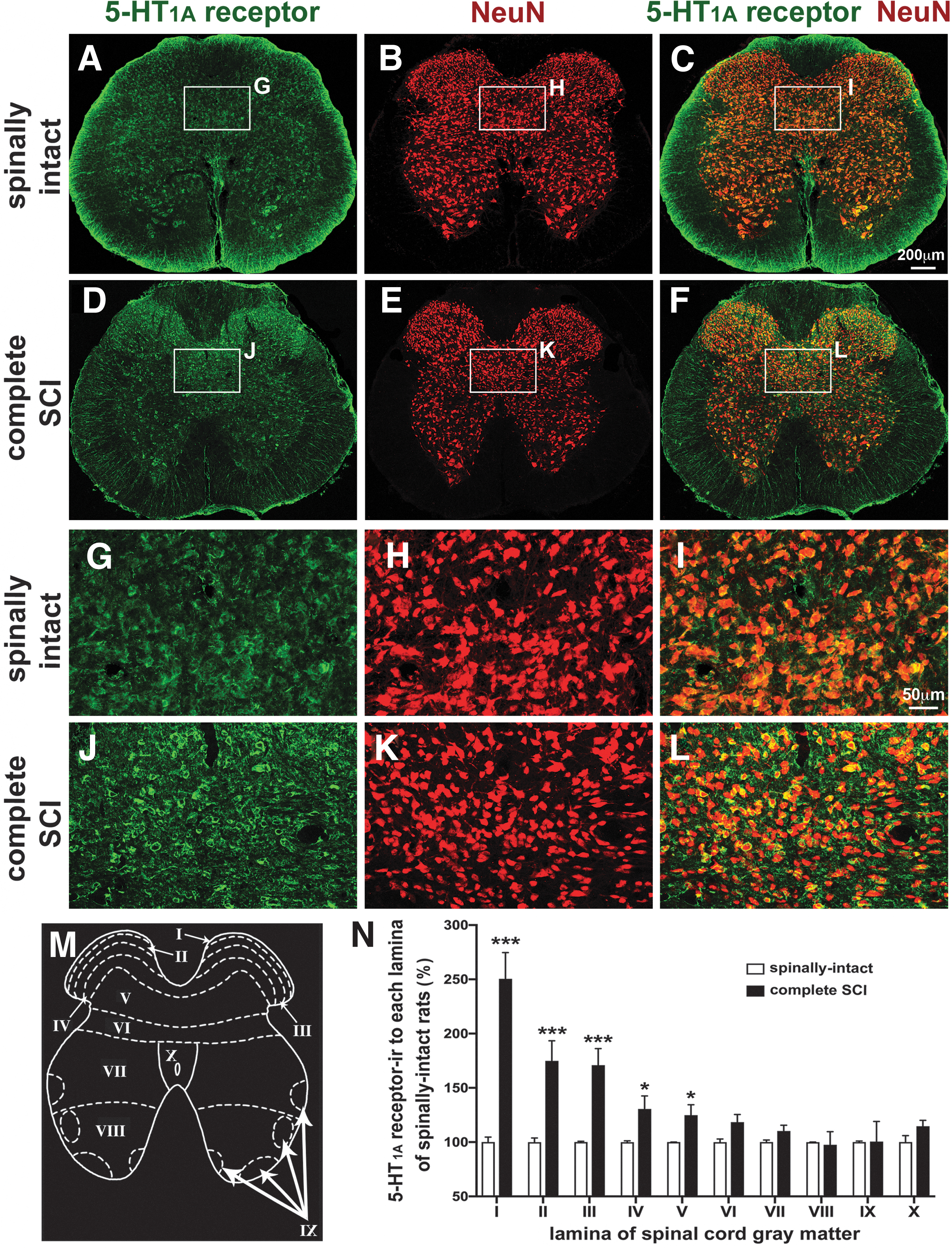
Immunoreactivity of 5-HT1A receptor in L6/S1 segments. Representative confocal images of double staining with antibodies for 5-HT1A receptor and NeuN from a spinally intact animal (
Pharmacological blockade of spinal 5-HT1A receptor regulates detrusor and EUS activities in awake spinally intact animals
Previous studies showed the inhibitory effects of the 5-HT1A receptor antagonist (WAY100635) on detrusor activity in anesthetized spinally intact rats. 15,16 We next investigated the dose-response effect of WAY100635 on both detrusor and EUS activity in awake animals. The CMG data showed continuous infusion of saline into the bladder without generating non-voiding activity prior to the micturition event (Fig. 4A). The application of WAY100635 demonstrated significant inhibition of detrusor activity in a dose-response manner in different CMG parameters (Figs. 4A and 5A). Trends toward a decrease in voiding efficacy (Fig. 5A, left graph of first row) and voiding interval (Fig. 5A, graph column of second row) were observed following the higher doses of WAY100635, including the highest dose (2 mg/kg) in voiding efficacy and the two higher doses (1 mg/kg and 2 mg/kg) in voiding interval shown the significant difference among them while compared with the baseline (Fig. 5A).

Dose-dependent effects of an intravenously injected 5-HT1A receptor antagonist (WAY100635) on the lower urinary tract (LUT) function of spinally intact animals. (
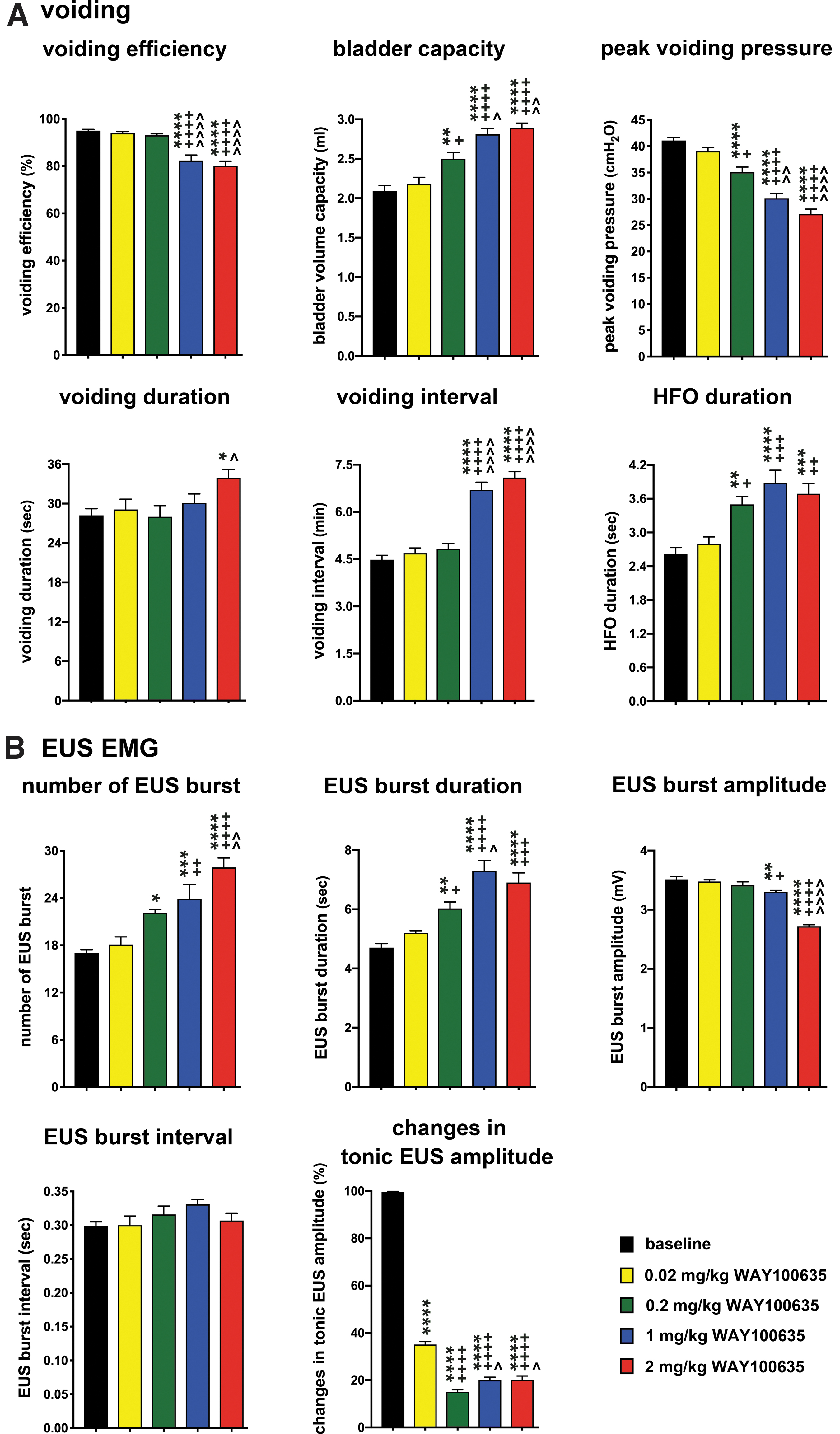
The effects of an intravenously injected 5-HT1A receptor antagonist (WAY100635) on the micturition reflex of spinally intact animals. Statistical analyses of parameters of voiding (
The measurement of peak voiding pressure (Fig. 5A, right graph of first row) demonstrated a significant dose-dependent decrease following WAY10065 (from 0.2 mg/kg, 1 mg/kg to 2 mg/kg) compared with baseline and the smaller dose of 0.02 mg/kg WAY100635. Measurements of bladder capacity (Fig. 5A, middle graph of first row) and HFO during the voiding period (Fig. 5A, right graph of second row) showed significant increases following the higher doses of WAY100635 (from 0.2 mg/kg, 1 mg/kg to 2 mg/kg) compared with the baseline and the smaller dose of 0.02 mg/kg WAY100635. Voiding duration (Fig. 5A, left graph of second row) showed a significant increase following 2 mg/kg WAY100635 compared with baseline and 0.2 mg/kg WAY100635.
In addition to the inhibitory effect on detrusor activity, we also found alterations in EUS recording following WAY100635 (Fig. 4B and 5B). There were significant increases in both EUS burst number and duration (Fig. 5B, left and middle graphs of first row) with the higher doses of WAY100635. The significant difference can be observed from 0.2 mg/kg, 1 mg/kg to 2 mg/kg. The EUS burst interval (Fig. 5B, left graph of second row) did not show any significant difference with or without WAY100635. The amplitude of EUS burst activity (Fig. 5B, right graph of first row) showed a significant decrease with the higher dose of WAY100635 (1 mg/kg and 2 mg/kg) among them. We further analyzed the tonic activity of EUS EMG during the filling phase (Fig. 5B, right graph of second row) and found a significant reduction (65–85%) following all doses of WAY100635 compared with baseline (Fig. 4C, 5B). We did not observe any incontinence or leakage of urine during the filling phase, even with such alterations in EUS activity.
Pharmacological blockade of spinal 5-HT1A receptor regulates detrusor and EUS activities in awake animals with T8 complete lesion
We next investigated the effects of WAY100635 on LUT function after complete SCI. In the CMG recordings, SCI animals showed typical neurogenic bladders, with many NVCs prior to the occurrence of a micturition event (Fig. 6A). In the voiding parameters of CMG analysis (Fig. 7A), the bladder-capacity analyses (Fig. 7A, second graph) demonstrated a significant increase with the higher dose of WAY100635 (1 mg/kg and 2 mg/kg) when compared with the baseline. The analyses of voiding duration (Fig. 8A, fourth graph) showed a significant increase at the highest dose of WAY100635 (2mg/kg) compared with the other lower doses and baseline. The HFO-duration analyses (Fig. 7A, sixth graph) showed a significant decrease following WAY100635 (from 0.2 mg/kg, 1 mg/kg to 2 mg/kg) compared with the baseline. There were no significant differences in either peak voiding pressure (Fig. 7A, third graph) or voiding efficacy (Fig. 7A, first graph) for any dose of WAY100635. For the non-voiding parameters (Fig. 7B), including the number of NVCs, the percentage of NVCs during the filling phase, and NVC amplitude, there were significant differences in the number of NVCs (Fig. 7B, first graph) and in the amplitude (Fig. 7B, third graph) of NVCs for the three highest doses of WAY100635 compared with baseline. There were significant differences in the percentage of NVCs during filling phase (Fig. 7B, second graph) at the two highest doses of WAY100635 when compared with the lower dose and baseline. For single NVC duration (Fig. 7B, fourth graph), both 1 mg/kg and 2 mg/kg WAY100635 caused a significantly longer duration than 0.2 mg/kg WAY100635 and baseline.

Dose-dependent response of an intravenously injected 5-HT1A receptor antagonist (WAY100635) on lower urinary tract (LUT) function in T8 complete spinal cord injury (SCI) animals. (

Effects of an intravenously injected 5-HT1A receptor antagonist (WAY100635) on micturition reflexes of T8 complete spinal cord injury (SCI) animals. Statistical analyses of parameters of voiding (

Dose-dependent response of an intrathecally delivered 5-HT1A receptor antagonist (WAY100635) on lower urinary tract (LUT) function in T8 complete spinal cord injury (SCI) animals. (
In addition to detrusor activity, we also investigated EUS activity during the voiding period (Fig. 7C). There was EUS burst activity observed in awake baseline recordings after T8 complete SCI (Fig. 6B). In the EUS EMG analysis (Fig. 7C), both the number of EUS bursts (Fig. 7C, first graph) and EUS burst duration (Fig. 7C, second graph) showed significant increases following 2 mg/kg WAY100635 when compared with the other doses and baseline. There were no significant differences found in either EUS burst amplitude (Fig. 7C, third graph) or EUS burst interval (Fig. 7C, fourth graph) for any dose of WAY100635.
To further test the findings of systemic delivered WAY100635 on LUT function, we performed the intrathecal injection of WAY100635 in two different doses (0.002 mg/kg and 0.02 mg/kg) with CMG and EUS recordings in the SCI animals (Fig. 8). In general, we have observed the similar results in parameters of voiding, non-voiding, and EUS EMG activity in a dose-dependent manner as what was found by systemic delivery. The significant increase in bladder capacity and the significant decrease in HFO duration were observed in the higher dose of WAY100635 treatment (Fig. 9A). The number, percentage, and amplitude of NVC were significantly decreased, but the single NVC duration was significantly increased with the higher dose of WAY100635 (Fig. 9B). The number and duration of EUS bursts were also significantly increased with the increase of the dose of WAY100635 (Fig. 9C). These data indicated that the acting sites of WAY100635 were mainly on the spinal cord, wherein WAY100635 regulated LUT function in SCI animals.
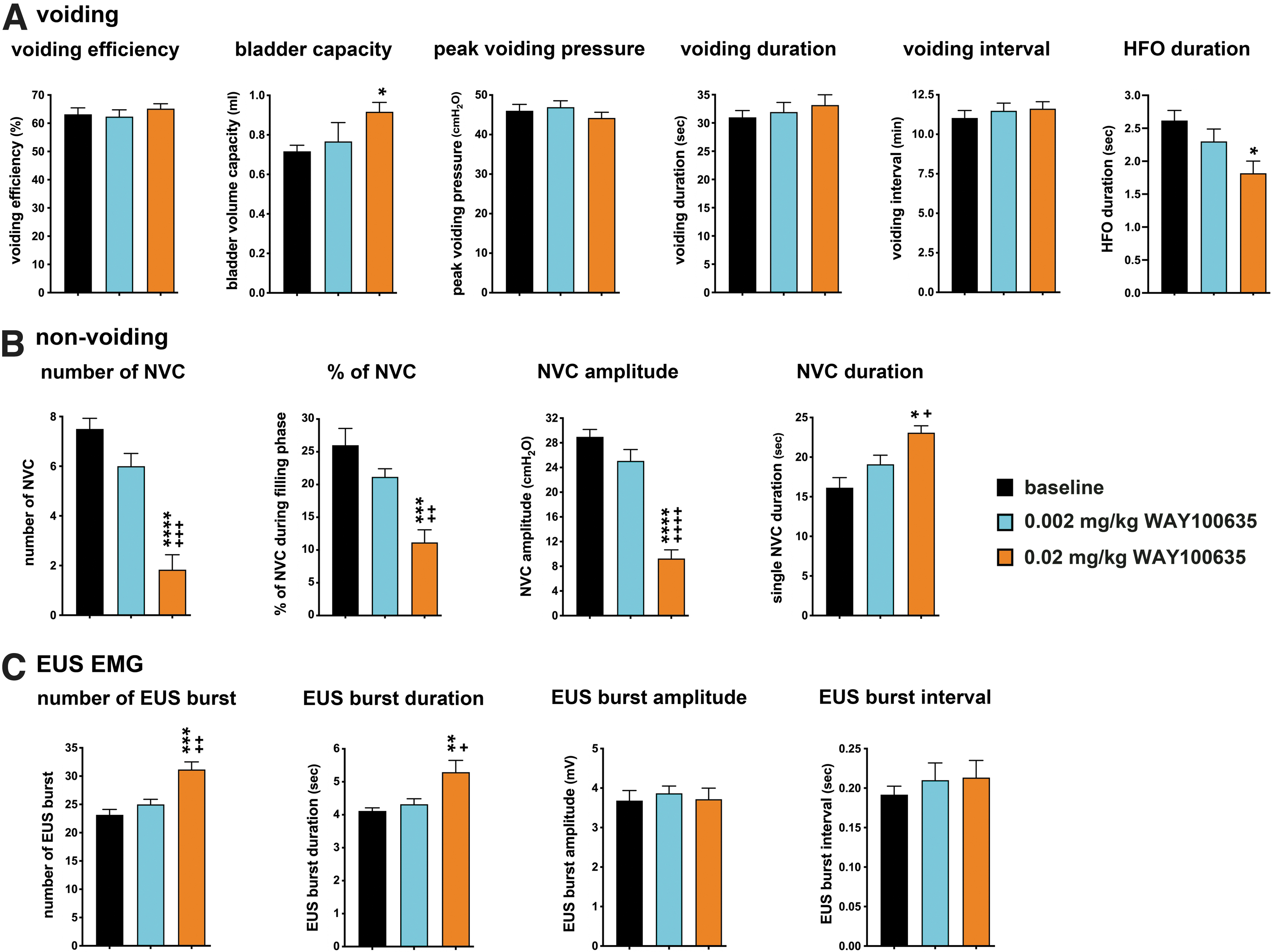
Effects of an intrathecally delivered 5-HT1A receptor antagonist (WAY100635) on micturition reflexes of T8 complete spinal cord injury (SCI) animals. Statistical analyses of parameters of voiding (
Application of propranolol diminishes WAY100635 regulation of LUT activity after T8 complete SCI
We next investigated whether the reduction in NVC activity produced by WAY100635 after complete SCI is caused bt activation of the sympathetic pathway, which relaxes the detrusor during the filling phase of micturition. The application of propranolol (a β1/β2-adrenergic receptor antagonist with weaker affinity for β3-adrenergic receptors; 2 mg/kg) diminished the effects of WAY100635 on both detrusor and EUS activities during the urodynamic and EUS EMG recordings (Fig. 10A, B). For the voiding parameters (Fig. 11A), the voiding interval (Fig. 8A, fifth graph) and HFO duration (Fig. 11A, sixth graph) were significantly decreased, and the peak voiding pressure (Fig. 11A, third graph) was significantly increased, as compared with 2 mg/kg WAY100635 alone or baseline. The propranolol did not alter the effects of WAY100635 on volume capacity (Fig. 11A, second graph) or voiding efficiency (Fig. 11A, first graph). For the non-voiding parameters (Fig. 11B), propranolol diminished the effects of WAY100635 on the number of NVCs (Fig. 11B, first graph), the amplitude of NVCs (Fig. 11B, third graph), and the percentage of NVCs during the filling phase (Fig. 11B, second graph). For the EUS EMG parameters (Fig. 11C), the increased number (Fig. 11C, first graph) and burst duration (Fig. 11C, second graph) of EUS produced by WAY100635 were diminished by propranolol. In contrast, there were no significant differences in EUS burst amplitude (Fig. 11C, third graph) or interval (Fig. 11C, fourth graph) after propranolol treatment.

Propranolol diminished the effects of WAY100635 on lower urinary tract (LUT) function after T8 complete SCI. (

Effects of a 5-HT1A receptor antagonist (WAY100635) and a beta-adrenoceptor antagonist (propranolol) on micturition reflexes of T8 complete spinal cord injury (SCI) animals. Statistical analyses of parameters of voiding (
Discussion
The serotonergic system plays an important role in regulating LUT function by acting through several receptor subtypes, including 5-HT1A, 5-HT2A, 5-HT2C, and 5HT7. Previous studies have shown compensatory overexpression of these spinal 5-HT receptors below the injured site after SCI, which may the result of the reduction of serotonergic inputs from the brainstem in mammals or lampreys. 23 –25 The upregulation of 5-HT1A receptor has not been reported rostral to the injured site after SCI, indicating that the overexpression of 5-HT receptors below the injured site could be part of the plasticity that regulates functional outcomes.
The present study investigated the spinal expression of 5-HT1A receptor rostral to the injured site and caudal to the injury, including both sympathetic and parasympathetic segments, at various time points following complete SCI. The Western blot data showed that the levels of 5-HT1A receptor were significantly upregulated below, but not rostral, to the injured site after complete SCI. These results are generally consistent with the previous report. 26 –28 We investigated further and found that the upregulation of 5-HT1A receptor was in both the T10-L2 and L6/S1 segments over a period of time after complete SCI. The increase in 5-HT1A receptor expression below the injured site may correlate to the time and location of 5-HT+ fiber degeneration after T8 complete SCI. A previous study 29 showed significant loss of 5-HT+ fibers from 7 days post-injury, and then continuing to 21 days with few remaining fibers, with fibers being completely absent at 30 days. The significant loss of 5-HT+ fibers was observed in the lower thoracic to upper lumbar segment, but not in the lower lumbar/sacral segment, at 7 days post-SCI.
These histological observations may support our findings of (1) the overall increase in 5-HT1A receptor expression at the later time points after SCI in both the sympathetic (T10-L2) and parasympathetic (L6/S1) segments, and (2) much higher expression of 5-HT1A receptor in the L6/S1 segment at the later time points (4 weeks and 8 weeks) than at the early time point (1 week) post-SCI. These data support the hypothesis that 5-HT1A receptors below the injured site are upregulated in response to the loss of serotonergic inputs after SCI.
An anatomical study from a previous report indicated that the distribution of overexpressed 5-HT1A receptor was in the dorsal or ventral horn of the gray matter, depending on the antibodies used against different epitopes of 5-HT1A receptor in the L3-L5 segment after complete T8 SCI. 28 In addition, the upregulation of 5-HT1A receptor below the injury site was dependent upon the sensory inputs after SCI. 28 In the present study, we have shown significant upregulation of 5-HT1A receptor in the dorsal portion of gray matter in the L6/S1 segment, which normally plays a role in LUT parasympathetic networks involved in bladder contraction and relaxation of EUS during voiding. The sensory afferent pathway contributes to the involuntary micturition reflex after SCI. A previous study indicated the activation of bladder c-fiber afferents contributing to NVC activity after complete SCI under deep anesthetic conditions. 5 The role of overexpressed 5-HT1A receptor in sensory afferents of parasympathetic circuits in micturition reflexes after SCI remains to be determined.
Several pharmacological interventions to activate or inhibit 5-HT1A receptor by using a selective agonist (8-OH-DPAT) or antagonist (WAY100635), respectively, indicated the important role of 5-HT1A receptor in modulating micturition reflexes in spinally intact animals under anesthesia. 16,17 In general, activation of 5-HT1A receptor can facilitate voiding reflexes by both detrusor and EUS activity, and inhibition of 5-HT1A receptor can inhibit detrusor activity. Particularly, the inhibition of 5-HT1A receptor at the L6/S1 level, but not the cervical or thoracic level, could affect micturition reflexes. 16 The present study demonstrated the inhibitory effects of WAY100635 on detrusor activity under awake conditions, which is consistent with previous studies performed in spinally intact animals under anesthesia 16 or awake. 15 In addition, we observed alterations in EUS activity by WAY100635, including increases in the number and duration of bursting, which may be caused by the increases in bladder capacity and HFO duration. However, there was no alteration of EUS burst interval induced by WAY100635, leading to a decrease in voiding efficiency. Interestingly, we also demonstrated reduced amplitude of EUS tonic activity during the bladder filling phase, although there is no report of this finding in anesthetized conditions. The proposed inhibitory effect of bladder activity by systematic administration of WAY100635 may relate to blocking 5-HT1A receptor on raphe neurons and increasing descending serotonin levels to activate inhibitory interneurons in the LUT network in spinally intact animals. 30 A similar inhibitory mechanism may help to explain the alteration of EUS tonic activity by WAY100635 during the bladder filling phase in the present study, as we did not observe the same alteration of EUS activity after complete SCI, which has no descending serotonergic control from the brainstem. These data indicate that the descending serotonergic system controls EUS tonic activity through 5-HT1A receptor, but it remains to be determined whether other subtypes of 5-HT receptor are involved in this regulation. For example, a recent study 31 showed that 5-HT2A receptor agonist could increase the tonic activity of EUS after complete SCI. Further study will be needed to clarify the EUS tonic activity regulated by either 5-HT1A receptor agonist in SCI animals or 5-HT2A receptor antagonist in spinally intact animals, respectively.
In the present study, we further investigated the effects of 5-HT1A receptor on LUT function by application of WAY100635 after T8 complete SCI. We found that WAY100635 had little effect on the majority of voiding parameters or EUS activity, except at the highest dose (2 mg/kg) applied. The presence of EUS phasic activity after SCI has been reported in several studies under awake conditions with or without restraint. 32 –34 In addition, there is a potential central pattern generator (CPG) for EUS phasic activity in the spinal circuit (between the T8 and L4 segments) that is independent of supraspinal control after SCI. 35 The presence of EUS burst activity in awake EUS EMG recording after SCI is consistent with previous reports. 32 –34 Our Western blot data and anatomical data indicated upregulation of 5-HT1A receptor in the dorsal region of spinal cord L6-S1 segments, which is involved in the neural circuits with EUS activity. It is unclear if increases in the number and duration of EUS bursts at the highest dose of WAY100635 (2 mg/kg) resulted from the action of L6/S1 segments or the potential CPG to EUS phasic activity. Another possibility to consider is increased bladder capacity. However, these alterations did not lead to the improvement of voiding efficiency, as HFO was decreased and the EUS burst interval was not altered. In addition, the observation of the increased number and duration of EUS burst from both activation of 5-HT1A receptor by agonists 17,22,35 and inhibition of 5-HT1A receptor by WAY100635 in the current study indicates the high plasticity of 5-HT1A receptor and the complexity of regulating EUS activity after SCI. The main difference is that 5-HT1A receptor agonist but not WAY100635 can improve voiding efficiency. One possibility is that these EUS burst activity altered by WAY100635 may be the result of the increase in bladder capacity. The other possibility is the interaction of WAY100635 with dopamine receptors.
A previous study 36 showed that WAY100635 has strong high affinity for D4 receptors but binds weakly to D2 receptors. In addition, the activation of D2 receptor can increase EUS burst activity after SCI, 37 whereas there is little information regarding D4 receptor regulation of lower urinary tract function. Further study will be needed to clarify this possibility.
One interesting finding in this study was the alteration of non-voiding contractions by WAY100635 during the bladder filling phase after T8 complete SCI. Normally, the bladder filling phase is the period of time to relax or inhibit the detrusor prior to micturition. Our data demonstrated a significant reduction in non-voiding activity by WAY100635, indicating that there were certain relaxation effects on the detrusor during the bladder filling phase that resulted from the activation of the adrenergic pathway in the sympathetic system. We found that upregulation of 5-HT1A receptor in the spinal cord segments involved in sympathetic neural activity. It is possible that the constitutive expression of 5-HT1A receptor may be associated with non-voiding activity, and that pharmacological inhibition by WAY100635 may lead to the activation of the sympathetic pathway to relax the detrusor, which in turn resulted in the reduction of non-voiding activity. We further tested this possibility by the application of propranolol to inhibit beta-adrenergic receptors following WAY100635 intervention. The diminishment of WAY100635-induced changes in non-voiding activity by propranolol supports this possibility. However, it remains to be determined if the effect of propranolol could directly reverse the effects of WAY100635 when former one is strong enough, and to explore the mechanism underlying interaction between the serotonergic and adrenergic systems. In addition, a future study can be considered to cut the hypogastric nerve (sympathetic pathway innervation to the detrusor) to determine whether WAY100635 reduction of non-voiding activity still occurred after SCI. This will help clarify if the effects of WAY100635 are mediated by the sympathetic system.
Conclusion
To summarize these findings, upregulation of 5-HT1A receptor was observed in spinal cord segments for both sympathetic and parasympathetic pathways after T8 complete SCI. The upregulated 5-HT1A receptors are involved in the regulation of LUT functions such as non-voiding activity and EUS activity.
Footnotes
Acknowledgments
We thank Dr. Christopher Nelson (Cleveland Clinic) for editorial assistance and critical reading of the manuscript.
Funding Information
This study was supported by National Institute of Neurological Disorders and Stroke Grant 2R01NS069765 and Craig H. Neilsen Foundation Grant# 597265 to Y.-S. Lee; as well as Department of Defense grant SC170117 to C.-Y. Lin.
Author Disclosure Statement
No competing financial interests exist.