
Abstract
Following spinal cord injury (SCI), fibrotic scar inhibits axon regeneration and impairs neurological function recovery. It has been reported that T cell–derived interferon (IFN)-γ plays a pivotal role in promoting fibrotic scarring in neurodegenerative disease. However, the role of IFN-γ in fibrotic scar formation after SCI has not been declared. In this study, a spinal cord crush injury mouse was established. Western blot and immunofluorescence showed that IFN-γ was surrounded by fibroblasts at 3, 7, 14, and 28 days post-injury. Moreover, IFN-γ is mainly secreted by T cells after SCI. Further, in situ injection of IFN-γ into the normal spinal cord resulted in fibrotic scar formation and inflammation response at 7 days post-injection. After SCI, the intraperitoneal injection of fingolimod (FTY720), a sphingosine-1-phosphate receptor 1 (S1PR1) modulator and W146, an S1PR1 antagonist, significantly reduced T cell infiltration, attenuating fibrotic scarring via inhibiting IFN-γ/IFN-γR pathway, while in situ injection of IFN-γ diminished the effect of FTY720 on reducing fibrotic scarring. FTY720 treatment inhibited inflammation, decreased lesion size, and promoted neuroprotection and neurological recovery after SCI. These findings demonstrate that the inhibition of T cell-derived IFN-γ by FTY720 suppressed fibrotic scarring and contributed to neurological recovery after SCI.
Introduction
Spinal cord injury (SCI) is one of the most severe public health problems in the world, leading to irreversible sensorimotor dysfunction. 1 In addition, there are about 0.6-0.9 million new patients with SCI every year. 2 Generally, a primary physical injury occurs after SCI, and secondary injury is rapidly triggered, including T cell infiltration, fibrotic scar formation, axonal demyelination, and neuronal apoptosis. 3,4 In particular, fibrotic scar, as a major obstacle to axonal regeneration and neurological recovery after SCI, 5 is considered as a therapeutic target for SCI.
Perivascular fibroblasts migrate, proliferate and deposit extracellular matrix to form a fibrotic scar in the lesion core to block axon regeneration after SCI. 4,6 -8 Fibrotic scar plays a beneficial role in maintaining tissue integrity and limiting inflammation response after SCI, 7 while excessively deposited fibrotic scar is detrimental to axon regeneration and neurological recovery, 9 indicating that moderate inhibition of fibrotic scar is necessary for function recovery after SCI. Under the direct action of external mechanical forces, the blood–spinal cord barrier is destroyed, then lymphocytes infiltrate the lesion site, leading to an inflammatory reaction. 10 After SCI, flow cytometry showed that T cells infiltrate and persist until 180 days post-injury (dpi), 11 while single-cell transcriptomic analyses showed that T cells appear in the spinal cord at 3 dpi and increased until 38 dpi. 12 Both methods indicted that there are no resident T cell populations in uninjured spinal cord. Recent studies show that T cells aggravate axon demyelination, neuronal apoptosis, blood–spinal cord barrier damage, and locomotor dysfunction in central nervous system (CNS) diseases. 13 -15 However, whether T cells are involved in fibrotic scar formation after SCI remains poorly understood.
IFN-γ is well known for potential pro-inflammatory and anti-viral functions. 16,17 IFN-γ has deleterious effects on CNS repair by activating M1 macrophages and microglia. 18,19 After SCI, γδ T cell provide the early source of IFN-γ and has harmful effect on functional recovery. 20 In addition, it has been reported that IFN-γ can cause fibrosis in peripheral organs such as the heart, lung and kidney. 21 –24 Otherwise, T cell-derived IFN-γ promotes fibroblast proliferation and deposition of extracellular matrix to form fibrotic scar in multiple sclerosis (MS). 25 Hence, we hypothesize that IFN-γ secreted by T cells promotes fibrotic scar formation after SCI.
Fingolimod (FTY720), as a modulator of the sphingosine 1-phosphate receptor (S1PR), has been approved in clinics for the treatment of MS 26 and can hinder fibrotic scar formation in MS. 25 The mechanism of the therapeutic effect of FTY720 is through the prevention of the egress of T cell out of lymph node, thus reducing the T cell infiltration in the CNS. 27 -29 W146 is a sphingosine-1-phosphate receptor 1 (S1PR1) antagonist and has been proven to reduce peripheral lymphocyte infiltration, including T cells. 30,31 In MS, FTY720 treatment not only reduced T cell count but also T cell activation. The presence of FTY720 also reduced IFN-γ production. 32 Previous study showed that the inhibition of IFN-γ by FTY720 was mediated via the SET/protein phosphatase 2A (PP2A) pathway. 33 A recent study showed that FTY720 reduces IFN-γ production in innate lymphoid cell (ILC) 1 and ILC3 inhibition of SIPR1. 34 FTY720 treatment in SCI achieves positive results, including electrophysiological recovery, reduction of lymphocyte infiltration, bladder function recovery, attenuation of vascular permeability and pathological astrocyte accumulation, less cavitation volume and neuron loss, maintenance of the blood–brain barrier, and reduction of neuropathic pain. 35 –39 However, whether FTY720 can attenuate fibrotic scarring after SCI is still unclear.
Here, we showed that platelet-derived growth factor receptor beta (PDGFRβ)+ fibroblasts surrounded the IFN-γ in spatiotemporal distribution after SCI. Moreover, IFN-γ is mainly secreted by T cells. In situ injections of IFN-γ in the normal spinal cord induced the aggregation of PDGFRβ+ fibroblasts and inflammation. FTY720 and W146 hindered CD3+ T cell infiltration and reduced fibrotic scar formation by inhibiting IFN-γ/ IFN-γR signaling. In comparison, in situ injection of IFN-γ diminished the effect of FTY720 on reducing fibrotic scarring. FTY720 treatment eliminated inflammation, decreased lesion size, and promoted neuroprotection and locomotor function recovery after SCI. These results suggest that the FTY720 treatment hinders INF-γ-mediated fibrotic scar formation and facilitates neurological recovery after SCI.
Methods
Animals and ethical approval
The Ethics Committee of Anhui Medical University approved all animal-related experiments (Approval No. LLSC20160052); 8- to 10-week-old female C57BL/6J mice (weighing 20 ± 2 g) from the Animal Experiment Center of Anhui Medical University were used. Animals were housed in a controlled environment (25℃, 50% humidity, and a 12-h light-dark cycle) and were free accessed to food and water.
Spinal cord crush model
The mice were anesthetized by intraperitoneal injection of 50 mg/kg of pentobarbital. The T10 level of spinal cord was exposed completely and clamped with calibrated Dumont #5 forceps (11252-20, Fine Science Tools, Germany) for 5 sec from both sides. 40 The wound was rinsed with saline and then sutured layer by layer with sutures. The red blood marks at the spinal cord clamp site and the violent swing of both lower limbs and tail indicate the successful establishment of the spinal cord crush model. The mice were allowed to recover in an incubator until they were fully awake. The mice with SCI received anti-infection treatment and auxiliary urination nursing twice daily.
FTY720 treatment
FTY720, W146, and saline (vehicle) was injected intraperitoneally daily after the SCI. The mice in the FTY720 group received 2 mg/kg FTY720 (No. 10006292; Cayman Chemical, U.S.) diluted in sterile saline, the mice in the W146 group received 10 mg/kg W146 (No. 10009109; Cayman Chemical) diluted in sterile saline, while the control group received the same volume of sterile saline until end of study for tissues analysis. 36,38,39
In situ injection of IFN-γ
The T10 spinal cord was exposed as described above. Then, the mice were fixed on the stereotaxic device, with the insertion site of a microinjection needle (7634-01 and 7803-05; Hamilton, Switzerland) located 0.3 mm lateral to the midline and at a depth of 0.8 mm from the dorsal spinal cord of the mice. 41 2 μL of 0.1 μg/μL recombinant human IFN-γ (C014; Novoprotein, China) dissolved in sterile phosphate-buffered saline (PBS) and injected into the normal spinal cord at 0.5 μL/min using a stereotactic syringe (KDS LEGATO 130; RWD, China). The control mice were injected with 2 μL of 0.1 μg/μL human serum albumin (HSA; A8230; Solarbio, China) dissolved in sterile PBS. All mice were sacrificed after 7 days post-injection. After SCI, the insertion site of a microinjection needle located to the center of lesion site and at a depth of 0.8 mm from the dorsal spinal cord of the mice. The mice received IFN-γ injections immediately after the spinal cord was clamped.
Tissue preparation and immunofluorescent staining
After cardiac perfusion with 0.01 M PBS (Servicebio, China) followed by 4% paraformaldehyde (PFA; Servicebio), the 0.5 cm of spinal cord tissue containing the injured core was subsequently fixed in 4% PFA at 4℃ for 5 h. Then the tissue was dehydrated in a 30% sucrose solution at 4°C for 24 h. Finally, the tissue was cut into 16-μm-thick continuous sagittal sections using a cryostat (NX50; Thermo Fisher Scientific, U.S.). The sections were blocked in 5% donkey serum (SL050; Solarbio) supplemented with 0.3% TritonX-100 (T8200; Solarbio) for 1 h at room temperature. Next, the sections were incubated with primary antibodies at 4°C overnight. The primary antibodies included goat anti-PDGFRβ (5 μg/mL; AF1042; R&D Systems, U.S.), rabbit-anti-IFN-γ (1:100, 15365-1-AP; Proteintech), rat-anti-IFN-γ (1:200, MM700; Invitrogen, U.S.), rabbit-anti-IFN-γR (1:100, PA586322; Intrivogen), rat-anti-CD3 (1:100, 14-0032-82; Intrivogen), rabbit anti-neurofilament (NF; 1:500, ab207176; Abcam, U.S.), rat anti–glial fibrillary acidic protein (GFAP; 1:200, 13-0300; Intrivogen), rat anti-CD68 (1:200, MCA1957; Bio-Rad, U.S.).
Subsequently, the sections were incubated with corresponding secondary antibodies at room temperature for 1 h, including donkey anti-goat Alexa Fluor 555, donkey anti-rabbit Alexa Fluor 488, donkey anti-rabbit Alexa Fluor 555 and donkey anti-rat Alexa Fluor 488 (1:500, A-21432, A-21206, A-21428, A-21208; Intrivogen). Finally, the sections were stained with 4′,6-diamidino-2-phenylindole (DAPI; C1005; Beyotime Biotechnology, China) to label the nuclei. The images were acquired through an A1 fluorescence microscope system (Zeiss).
Image analysis
All quantitative analyses were performed under double-blind conditions. Three to five sections per animal were used for quantitation using immunofluorescences staining. The proportions of CD3+IFN-γ+ cells and CD68+IFN-γ+ cells were quantified and expressed as the numbers of marker+ cells divided by all IFN-γ+ cells with 10 × magnification, and only DAPI+ cells were counted. 42 The number of IFN-γ+ cells was quantified and normalized to the area of the spinal cord segment spanning the injured core with 10 × magnification, and only DAPI+ cells were counted. The number of CD3+ cells was quantified with 4 × magnification, and only DAPI+ cells were counted. To evaluate the area of the fibrotic scar, the area of PDGFRβ at the lesion site was measured, and then divided the area of PDGFRβ by the area of the entire spinal cord with 4 × magnification. 43 Similarly, the GFAP- area and CD68+ area were normalized to the area of the spinal cord segment spanning the injured core with 4 × magnification. To evaluate axon preservation, the number of NF+ axons longer than 1 μm in the GFAP- region was counted and normalized to the area of the GFAP- region, and the immunofluorescence of 5-HT was normalized to the area of the spinal cord segment spanning the injured core with 10 × magnification. To evaluate neuronal survival, The sections were divided into three parts (Z1: 0-250 μm, Z2: 250-500 μm, Z3: 500-750 μm) according to the straight distance from the lesion core, the number of NeuN+ cells in Z1-Z3 was quantified and normalized to the area of the divided part area separately with 10 × magnification.
Western blot analysis
The proteins of the spinal cord were extracted by radioimmunoprecipitation assay (RIPA) buffer (P0013B; Beyotime) supplemented with Protease Inhibitor Cocktail (04693124001; Roche, Switzerland) and Phosphatase Inhibitor (04906845001, Roche). After being loaded onto a sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) gel, the proteins were separated by electrophoresis and transferred to a polyvinylidene difluoride (PVDF) membrane (Millipore, Schwalbach, Germany). Then, 5% dried skim milk was used to block the nonspecific binding sites of membranes at 37℃ for 1.5 h. Afterwards, the membranes were incubated with primary antibodies at 4℃ overnight, including mouse anti-glyceraldehyde 3-phosphate dehydrogenase (GAPDH; 1:5000, ab8245; Abcam, China) and rabbit anti-IFN-γ (1:3000, 15365-1-AP, Proteintech). After being washed, secondary antibodies, including goat anti-mouse secondary antibody (1:10000, A4416; Sigma, U.S.) and goat anti-rabbit secondary antibody (1:10000, A0545, Sigma), were added and incubated with the membranes at 37℃ for 1 h. The band signals were detected using enhanced chemiluminescence (32209; Intrivogen). ImageJ software version 1.53 (National Institutes of Health, U.S.) was used for quantitation analysis.
Behavioral assessment
All mice were scored in an open field using the Basso Mouse Scale (BMS) scoring protocol. 44 Two independent observers assessed each mouse at pre-injury, and 3, 7, 14, and 28 dpi. The final BMS value is the average of the two observers' scores.
Footprint analysis was used to assess the neurological recovery at 28 dpi. Forelimbs were stained with green dye, and hindlimbs were stained with red dye. The mice walk on a narrow track (80 cm long, 4 cm wide) lined with white paper to measure paw rotation, stride length, and width.
The above behavioral assessments were performed in a blind manner.
Statistical analysis
The data were presented as mean ± standard error of the mean. One-way or two-way analysis of variance (ANOVA) with a Tukey's post hoc test was used for multiple comparisons, and Student's t-test was used for comparisons between two groups. GraphPad Prism version 8.0 (GraphPad Software, U.S.) was used for statistical analysis. A p < 0.05 was seen as statistical significance.
Results
The spatiotemporal distribution and expression of IFN-γ and fibroblasts after SCI
PDGFRβ, as a fibrotic scar-forming cells marker, is mainly expressed in the lesion core and form a characteristic honeycomb-like circular structure after SCI. 45 Therefore, PDGFRβ was used to track fibroblasts in the lesion core. To clarify the spatiotemporal distribution and expression level changes of IFN-γ after SCI, the immunofluorescence staining and Western blot assay for IFN-γ were performed. The immunofluorescence staining results showed that the expression of IFN-γ increased from 3 to 7 dpi and peaked at 14 dpi after SCI. Meanwhile, the population of PDGFRβ+ fibroblasts increased significantly from 3 to 7 dpi and aggregated to the lesion site, forming continuous fibrotic scar boundary in the lesion core at 14 and 28 dpi, which was consistent with previous studies. 4,6,7 These results indicated that IFN-γ was mainly surrounded by PDGFRβ+ fibroblasts in the lesion core after SCI (Fig. 1A, 1B). The Western blot results showed that IFN-γ was not expressed in the uninjured spinal cord, and its expression level increased from 3 to 7 dpi and gained a peak at 14 dpi, which was consistent with the immunofluorescence staining results (Fig. 1C, 1D). These results showed that the fibrotic scar was closely correlated with IFN-γ in spatiotemporal distribution and expression, indicate that IFN-γ may play a vital role in fibrotic scar formation after SCI.
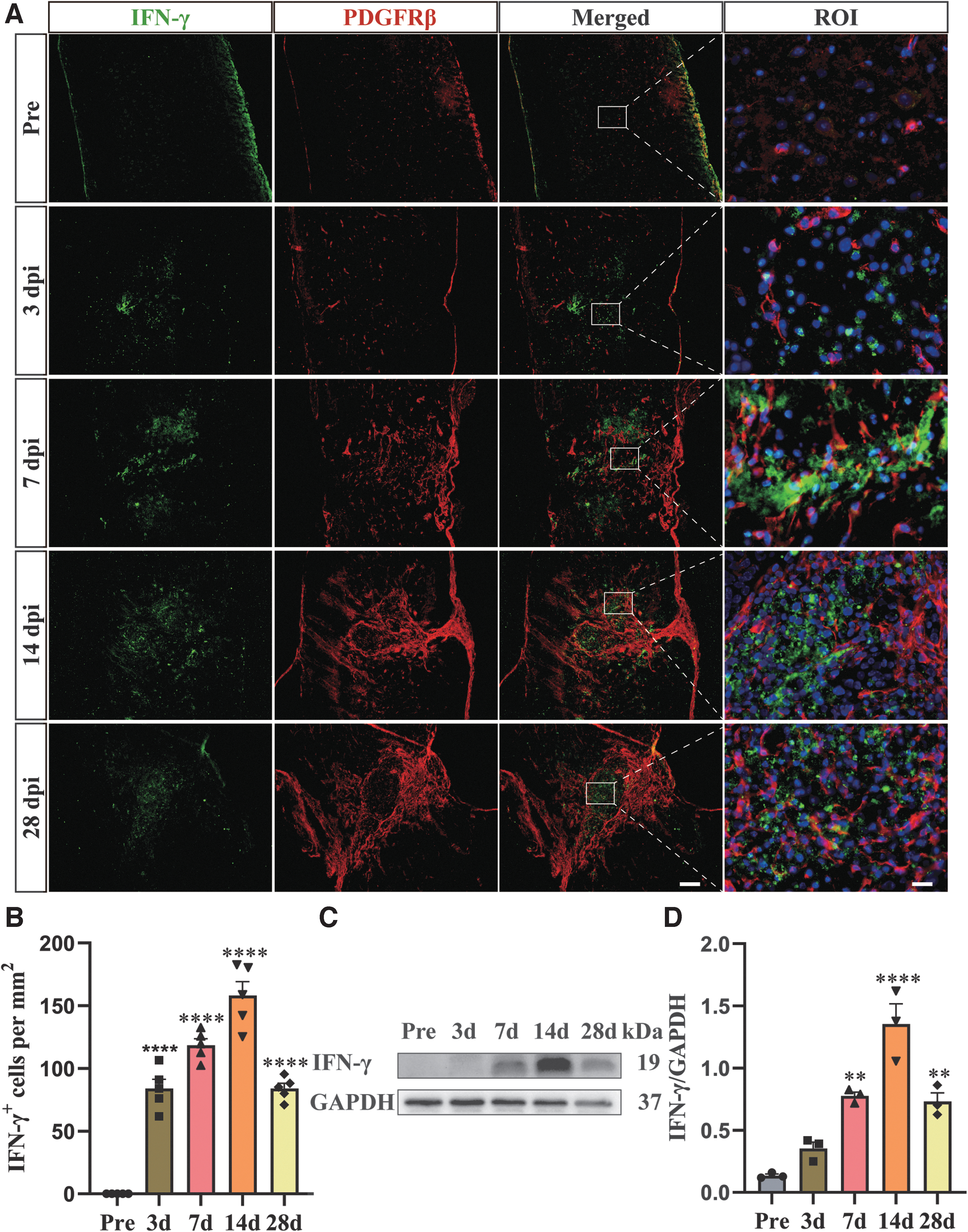
Spatiotemporal distribution of interferon (IFN)-γ and platelet-derived growth factor receptor beta (PDGFRβ)+ cell after spinal cord injury (SCI).
IFN-γ is mainly secreted by CD3+T cell after SCI
After SCI, peripheral monocytes rapidly infiltrate into the lesion core, mediate inflammatory responses, and participate in the construction of spinal cord micro-environment. To further determine the cellular source of IFN-γ after SCI, immunofluorescence staining of IFN-γ with CD3+ T cell and CD68+ macrophages/microglia in the lesion core was performed at 14 dpi. The results showed that IFN-γ was mainly co-localized with CD3+ T cells. The percentage of IFN-γ co-localized with CD3 was as high as about 60%. In addition, CD68+ macrophages/microglial was also expressed INF-γ in the lesion core, which represented that IFN-γ had multiple cellular origins (Fig. 2A-C). This result was consistent with previous report that T cells were the main source of IFN-γ after SCI. 20

Interferon (IFN)-γ is mainly secreted by CD3+ T cell after spinal cord injury (SCI).
IFN-γ induces fibrosis and inflammation in normal spinal cord
To directly reveal the effect of IFN-γ on fibrotic scarring after SCI, we injected IFN-γ in situ into the T10 segment of the normal spinal cord. For 7 days after injection, immunofluorescence staining was performed to detect the fibrotic scar formation and inflammation response. As shown in Figure 3A, IFN-γ was successfully injected into the spinal cord compared with the HSA group. IFN-γ induced the aggregation of PDGFRβ+ fibroblasts in the injection site of normal spinal cord, while the HSA group showed that the injection of saline did not lead to fibrotic scarring. Notably, a sharp glial scar was also formed at the edge of the fibrotic scar after IFN-γ injection (Fig. 3B). IFN-γ also caused a great number of CD68+ macrophages/microglia accumulation in the injection site of normal spinal cord, while no obvious CD68+ macrophages/microglia infiltration was observed in the HSA group (Fig. 3C), indicating that IFN-γ has a strong pro-inflammatory effect. In addition, infiltrating macrophages and microglia were confined to the IFN-γ injection site and tightly surrounded by fibrotic scar. Therefore, our results suggest that IFN-γ can lead to fibrotic scaring and inflammation response in normal spinal cord by inducing PDGFRβ+ fibroblast aggregation and CD68+ macrophages/microglia accumulation.

In situ injection of interferon (IFN)-γ induces fibrosis and inflammation in normal spinal cord.
FTY720 attenuates fibrotic scarring through IFN-γ/ IFN-γR signaling after SCI
During the early phase after SCI, T cells infiltrate the lesion site and hinder functional recovery by secreting IFN-γ. 15,20 To examine the effect of T cells in fibrotic scar formation after SCI, we used FTY720 and to inhibit T cells infiltration. Immunofluorescence staining revealed that the population of CD3+ T cells in the FTY720 treatment group was significantly decreased compared with the control group at 28 dpi (Fig. 4A, 4B). Meanwhile, the area of fibrotic scar was significantly decreased in the FTY720 treatment group compared with the control group at 28 dpi (Fig. 4C). And the S1PR1 antagonist, W146, could also exert a similar therapeutic effect to FTY720, suggesting that FTY720 acts by blocking the S1P/S1PR1 pathway on T cells. Therefore, our results suggest that FTY720 can hinder T cell infiltration by blockade the S1P/S1PR pathway and attenuate fibrotic scarring.

Fingolimod (FTY720) attenuates fibrotic scarring through interferon (IFN)-γ/ IFN-γR signaling after spinal cord injury (SCI).
The IFN-γ/ IFN-γR signaling pathway has been demonstrated to be involved in pathological fibrosis in a variety of tissues. 21 –24 To further clarify whether the mechanism of FTY720 inhibiting fibrotic scarring is related to IFN-γ/ IFN-γR signaling pathway, we detected the distribution and expression level change of IFN-γ after FTY720 treatment and IFN-γR costaining with the PDGFRβ+ fibroblasts at 14 dpi by immunofluorescence staining. Immunofluorescence staining confirmed that FTY720 and W146 significantly reduced IFN-γ production compared with the control group (Fig. 4D, 4E). And the costaining results showed partial co-localization between IFN-γR and PDGFRβ+ fibroblasts (Fig. 4F). Hence, these results suggest that the inhibition of IFN-γ/IFN-γR signaling by FTY720 can reduce fibrotic scar formation after SCI.
In situ injection of IFN-γ diminished the effect of FTY720 on reducing fibrotic scarring
To further demonstrate that the effect of FTY720 on fibrosis is mediated by IFN-γ, we injected IFN-γ in situ into the T10 segment of the injured spinal cord after SCI. Immunofluorescence staining revealed that the area of fibrotic scar was significantly decreased in the FTY720 + HSA group compared with the control group (saline + HSA), and the area of fibrotic scar in the FTY720 + IFN-γ group was significantly increased compared with the control and FTY720 + HSA group at 28 dpi (Fig. 5A, 5B). Altogether, these results indicate that IFN-γ diminished the effect of FTY720 on attenuated fibrosis.

In situ injection of interferon (IFN)-γ diminished the effect of fingolimod (FTY720) on reducing fibrotic scarring.
FTY720 inhibits the inflammation after SCI
Fibrotic scar has been reported to limit the extent of inflammation. 7,43 Therefore, we used immunofluorescence staining to explore the effect of FTY720 on inflammation at 28 dpi. Compared with the control group, FTY720 treatment significantly reduced CD68+ macrophages/microglial accumulation (Fig. 6A). As shown in Figure 5B, the CD68+ macrophages/microglia area was found to be 40% less than the control group after FTY720 treatment, which may be attributed to the inhibition of IFN-γ expression by FTY720.
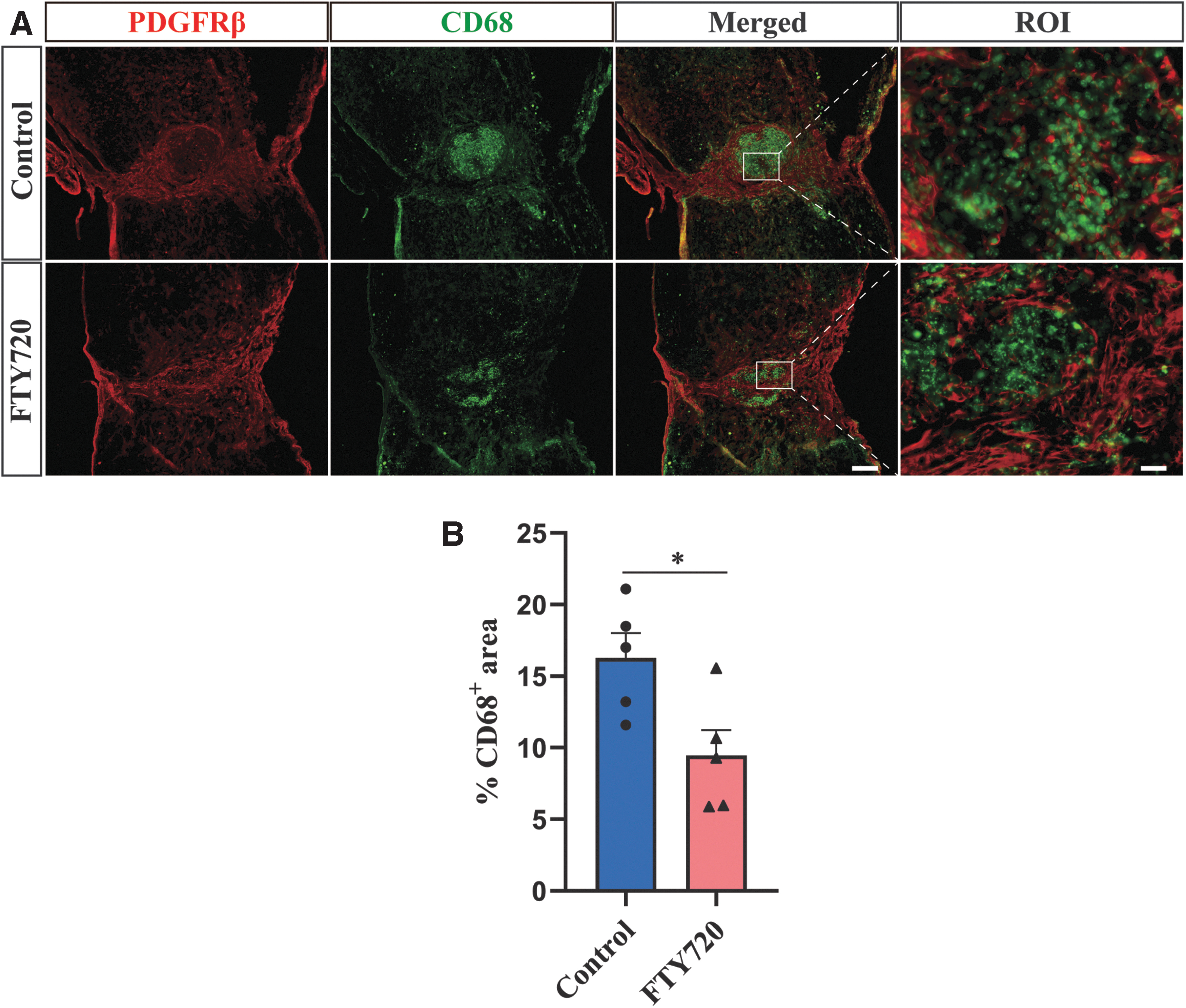
Fingolimod (FTY720) inhibits the inflammation after spinal cord injury (SCI).
FTY720 promotes neuroprotection and neurological recovery after SCI
Fibrotic scar and inflammatory responses are major obstacles to axon preservation and neurological recovery after SCI. To confirm whether FTY720 attenuates fibrotic scarring and inflammation after SCI to promote axon preservation and neurological recovery, we first assessed the NF+ and 5-HT+ axon preservation, GFAP- lesion size and NeuN+ neuron survival by immunofluorescence staining at 28 dpi. The results showed that compared with the control group, the number of NF+ axons in the lesion core significantly increased with the area of GFAP- lesion size obviously decreased in the FTY720-treated group (Fig. 7A-C). There were significantly more 5-HT axons crossing the lesion site in the FTY720 group than in the control group (Fig. 7D, 7E). Moreover, compared with the control group, the number of viable NeuN+ neurons in the Z1-Z3 region was significantly increased in the FTY720 group (Fig. 7F, 7G). These results suggest that FTY720 reduces lesion size and promotes axon preservation and neuron survival.

Fingolimod (FTY720) promotes axon preservation after spinal cord injury (SCI).
To further assess the neurological recovery after SCI in mice, BMS score and footprint analysis were performed (Fig. 8A, 8B). Compared with the control group, the mice injected with FTY720 had better recovery of hindlimb locomotor function and corresponding higher BMS scores at 14 and 28 dpi (Fig. 8B). In addition, footprint analysis further demonstrated a better recovery of locomotor function in the FTY720 group than in the control group (Fig. 8C), including longer stride length, shorter stride width, and minor paw rotation at 28 dpi (Fig. 8D-F). Overall, our results indicate that FTY720 treatment promotes neurological recovery after SCI.

Fingolimod (FTY720) promotes neurological recovery after spinal cord injury (SCI).
Discussion
Neuron loss and impaired locomotor function after SCI are severe health problems further exacerbated by immunocyte infiltration and inflammatory response. Our study shows that PDGFRβ+ fibroblasts are closely correlated with IFN-γ in spatiotemporal distribution after SCI, and the primary source of IFN-γ is CD3+ T cell. In situ injections of IFN-γ induced fibrosis and inflammation in the normal spinal cord. In addition, FTY720 treatment could inhibit T cell infiltration, attenuate fibrotic scar formation through inhibiting IFN-γ/ IFN-γR signaling, suppress inflammation, decrease lesion size, promote axon preservation, and facilitate neurological recovery in mice, indicating the deleterious effects of IFN-γ after SCI. Overall, the present study reveals a new therapeutic role of FTY720 by inhibiting the IFN-γ-mediated fibrotic scarring after SCI (Fig. 9A, 9B).
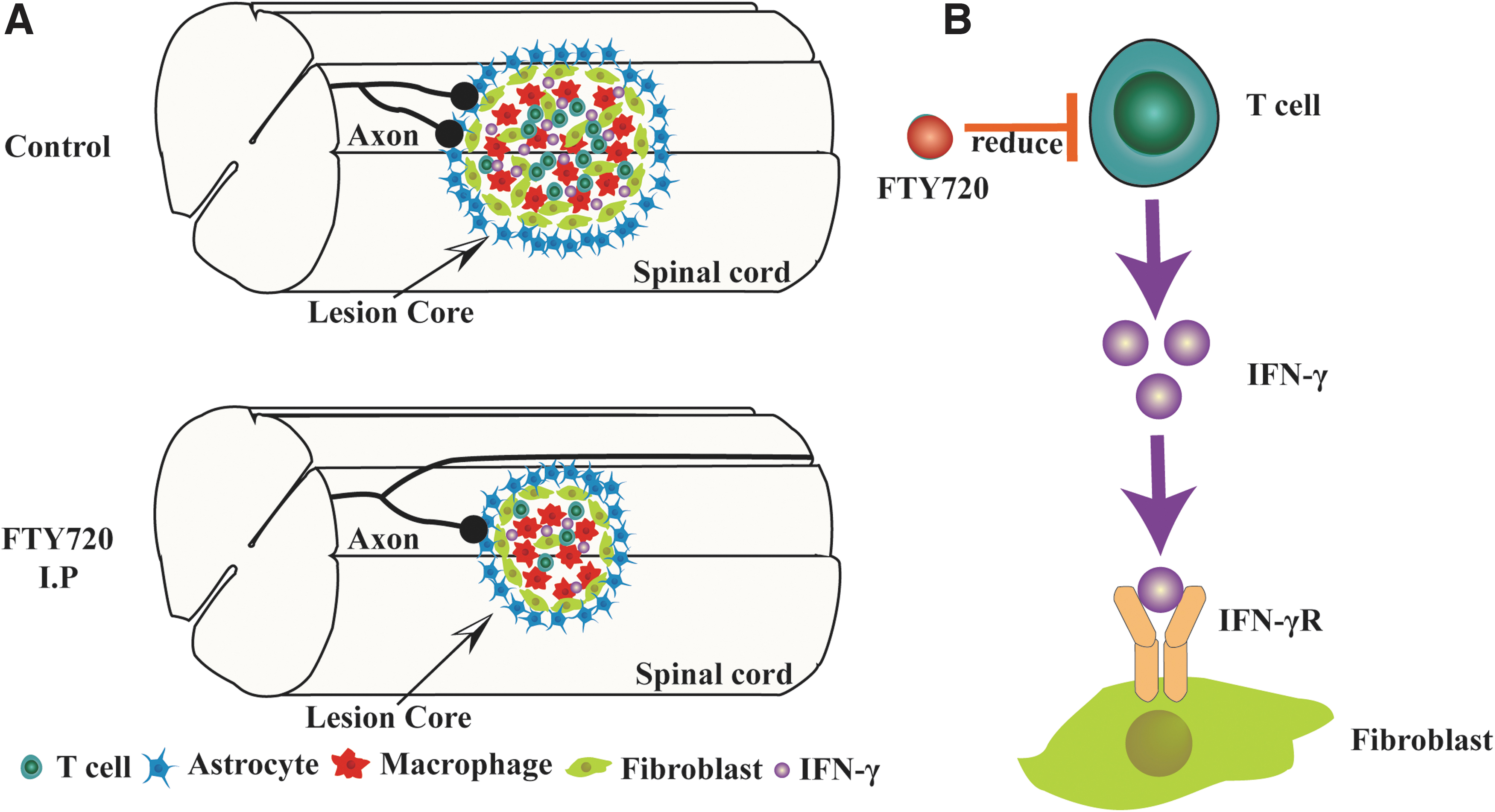
A pattern diagram of fingolimod (FTY720) to reduce fibrotic scar formation after spinal cord injury (SCI). Intraperitoneal injection of FTY720 reduced T cells-derived interferon (IFN)-γ, which promotes the proliferation and migration of fibroblasts, thereby attenuating the formation of fibrotic scar, promoting neuroprotection and locomotor function recovery after SCI.
The scar tissue formed after SCI includes glial scar and fibrotic scar. 46 The fibrotic scar is formed by fibroblasts migrating, proliferating, and encasing large numbers of macrophages in the injured core. 4,6,8 Fibrotic scar maintains tissue integrity and limits inflammation, 7 but persistent fibrotic scar and inflammation prevent axon regeneration and impair functional recovery. 47,48 Thus, a modest reduction of fibrotic scar can significantly promote axon regeneration and neurological recovery after SCI. 7,43,47 It suggests that fibrotic scar may be an important therapeutic target after SCI.
T cells play an essential role in CNS diseases. It has been reported that T cells exacerbate axonal damage and demyelination, promote neuronal apoptosis and locomotor and cognitive dysfunction in CNS diseases. 13,14 After SCI, T cells begin to appear and persist in the spinal cord for several months. 11,12 Some studies have reported that T cells are recruited to the lesion site via CCL2/CCR2 signaling after SCI, 49 and T cells provide the early source of IFN-γ to impair functional recovery after SCI. 20 In our study, we further confirmed that IFN-γ is mainly secreted by T cells after SCI. However, there is also a tiny fraction of IFN-γ that is of other cellular origins, and in the present study, we did not exclude the effect of this fraction of IFN-γ. To further confirm the effect of IFN-γ in fibrotic scarring after SCI, we need to block IFN-γR directly in next study.
The role of IFN-γ signaling in fibrosis remains controversial in peripheral tissues. 21 –24,50 -52 In MS, IFN-γ can promote fibroblast proliferation and deposition of extracellular matrix resulting in fibrotic scar formation. 25 The dual role of IFN-γ in fibrosis may depend on the disease stage and receptor type. To investigate the role of IFN-γ in the formation of fibrotic scar, we chose to inject IFN-γ directly into the uninjured spinal cord to avoid the influence of the complex microenvironment of the injured spinal cord. Our results show that IFN-γ can lead to fibrotic scar formation and inflammation in the uninjured spinal cord. These suggest that IFN-γ signaling may be essential in forming a fibrotic scar. Our previous studies have revealed that activation of the PDGFRβ pathway can induce fibrotic scarring directly. 53 It is still not sure whether the effect of IFN- γ in fibrotic scarring is through direct activation of the PDGFRβ pathway or indirect activation by inducing macrophages/microglia accumulation and secreting PDGFRD. The detailed mechanisms of IFN-γ in fibrotic scarring need to be explored by further research. In addition to fibroblasts, IFN-γR is expressed by astrocytes, neurons, and macrophages/microglia in CNS. 14,25,54,55 However, in the present study, we only focused on the association of IFN-γ with fibrotic scar and inflammation. Next, we will further explore the effect of IFN-γ on macrophages/microglia phagocytosis and myelin regeneration after SCI.
FTY720 is a non-selective sphingosine 1-phosphate receptor (S1P1, S1P3, S1P4, and S1P5) modulator phosphorylated by sphingomyelin kinase. 56 Phosphorylated FTY720 can bind to S1PR1 in the initial stage after entering the lymph node and activate S1PR1 as S1P, after which it rapidly leads to internalization of S1PR1 on T cells and becomes a highly efficient S1PR1 functional antagonist, so that T cells no longer respond to changes in S1P concentration and are retained in the lymph node. 57 In CNS diseases, it can also reduce the infiltration of peripheral T cells into the CNS by blocking the SIP/SIPR signaling pathway to keep T cells in the lymph nodes. 27 -29 FTY720 has been shown to play a beneficial role in restoring neurological function in SCI, such as reducing T cell number in the lesion site, improving bladder function in contusion SCI rats, 35 decreasing glial scarring, cavitation volume and neuron loss in right hemisection SCI rats and dogs. 36,37 Previous studies showed that FTY720 could inhibit fibrosis in peripheral organs or tissues, such as skin and renal. 58 -60 However, the effect of FTY720 in fibrotic scarring after SCI is still undefined.
To investigate the role and mechanism of FTY720 in fibrotic scar formation after SCI, we examined the IFN-γ/ IFN-γR pathway and fibrotic scar formation. Our study found that IFN-γ receptors were partly expressed on PDGFRβ+ fibroblasts, while treatment with FTY720 after SCI reduced T cell infiltration, inhibited IFN-γ signaling, and attenuated fibrotic scar formation. Further, in situ injection of IFN-γ significantly diminished the effect of FTY720 on reducing fibrotic scarring, indicating that IFN-γ plays a role in fibrotic scar formation. Previous studies have shown that a moderate fibrotic scar can prevent the spread of inflammation and that complete removal of fibrotic scar leads to the spread of inflammation at 14 days. 6,7 In our study, FTY720 reduced the inflammation area. Considering that in situ injections of IFN-γ could induce fibrosis and inflammation in normal spinal cord in this study, it may be due not only to the integrity of the fibrotic scar but also to the reduction of IFN-γ. At the same time, reducing fibrotic scar promotes neuroprotection and neurological recovery, which may also benefit from the reduction of IFN- γ.
Conclusion
In summary, our study confirms that IFN- γ, mainly from T cells, could facilitate the formation of fibrotic scar after SCI. Moreover, in situ injections of IFN-γ induced fibrosis and inflammation in the normal spinal cord. FTY720 treatment also could hinder T cell infiltration, attenuate fibrotic scar formation through inhibiting IFN-γ/ IFN-γR signaling, and suppress inflammation after SCI, thereby decrease lesion size, promote neuroprotection and facilitate neurological recovery. These findings suggest that FTY720 is a promising therapy for reducing fibrotic scar formation and neurological recovery after SCI.
Footnotes
Authors' Contributions
JJ designed the experiments. YL and YC wrote the manuscript. YL, XH, FO, JL, JX, JY, and FS performed the experiments. LC, MZ, and YL administrated the project. SY, ZL, FY, YL, and YS analyzed the data. All authors read and approved the final manuscript.
Funding Information
Financial support for this research was provided by the Provincial Natural Science Research Key Project of Colleges and Universities of Anhui Province (Grant number KJ2021A0310 and KJ2021A0316) and the Key Research and Development Projects of Anhui Province (Grant number 202004j07020042).
Author Disclosure Statement
No competing financial interests exist.