
Abstract
Increasing concerns have been raised about the long-term negative effects of subconcussive repeated head impact (RHI). To elucidate RHI injury mechanisms, many efforts have studied how head impacts affect the skull-brain biomechanics and have found that mechanical interactions at the skull-brain interface dampen and isolate brain motions by decoupling the brain from the skull. Despite intense interest, in vivo quantification of the functional state of the skull-brain interface remains difficult. This study developed a magnetic resonance elastography (MRE) based technique to non-invasively assess skull-brain mechanical interactions (i.e., motion transmission and isolation function) under dynamic loading. The full MRE displacement data were separated into rigid body motion and wave motion. The rigid body motion was used to calculate the brain-to-skull rotational motion transmission ratio (Rtr ) to quantify skull-brain motion transmissibility, and the wave motion was used to calculate the cortical normalized octahedral shear strain (NOSS) (calculated based on a partial derivative computing neural network) to evaluate the isolation capability of the skull-brain interface. Forty-seven healthy volunteers were recruited to investigate the effects of age/sex on Rtr and cortical NOSS, and 17 of 47 volunteers received multiple scans to test the repeatability of the proposed techniques under different strain conditions. The results showed that both Rtr and NOSS were robust to MRE driver variations and had good repeatability, with intraclass correlation coefficient (ICC) values between 0.68 and 0.97 (fair to excellent). No age or sex dependence were observed with Rtr , whereas a significant positive correlation between age and NOSS was found in the cerebrum, frontal, temporal, and parietal lobes (all p < 0.05), but not in the occipital lobe (p = 0.99). The greatest change in NOSS with age was found in the frontal lobe, one of the most frequent locations of traumatic brain injury (TBI). Except for the temporal lobe (p = 0.0087), there was no significant difference in NOSS between men and women. This work provides motivation for utilizing MRE as a non-invasive tool for quantifying the biomechanics of the skull-brain interface. It evaluated the age and sex dependence and may lead to a better understanding of the protective role and mechanisms of the skull-brain interface in RHI and TBI, as well as improve the accuracy of computational models in simulating the skull-brain interface.
Introduction
Subconcussive repeated head impact (RHI) exposure has been recognized as a public health issue and an important component of traumatic brain injury (TBI). 1 –3 Growing evidence suggests that people with a history of RHI are more vulnerable to subsequent TBI and can have long-term neurological problems, such as headaches, balance problems, and memory impairments. 4 It has been suggested that chronic traumatic encephalopathy (CTE) may in part be a result of RHI. 5,6 Early diagnosis of RHI changes in the brain can thus help track associated neurological impairments and provide preventative strategies to avoid future damage. Patients, however, usually show no clinical symptoms of RHI and in vivo identification remains challenging. Advanced neuroimaging techniques (e.g., functional magnetic resonance imaging [fMRI] or diffusion tensor imaging [DTI]) have shown brain functional and structural changes correlated with RHI, but the findings are not consistent across studies. 7 There is an urgent need for improved detection and characterization of the effects of RHI exposure and its consequences with non-invasive and quantitative tools.
The main risk of RHI is external force-induced brain deformation during head acceleration and rotation. 1 To elucidate RHI injury mechanisms, many efforts seek to study how exposure to head impacts affects the skull-brain biomechanics. Relative skull and brain motions occurring during an impact have been the subject of intense interest for decades with animal experiments, 8 –10 cadaver experiments, 11 –14 physical and in silico models, 15 –17 and human studies using MR tagging techniques. 18 –22 It has been found that the mechanical coupling characteristics at the skull-brain interface, especially involving the meninges, cerebrospinal fluid (CSF), arachnoid trabeculae (AT), and subarachnoid vasculature, hereafter the “pia-arachnoid complex (PAC),” are pivotal in protecting the brain from external mechanical trauma. To isolate the brain from potential subconcussive damage, the PAC interface dampens the motion transmitted into the brain and tethers the brain in suspension within the skull. Specifically, the CSF fills the subarachnoid space functioning as a thin cushion against external forces, and the ATs effectively “stitch” the pia and arachnoid together to provide mechanical stability for the brain. 23 An ex vivo animal study compared the separated and the PAC-overlaid brains under indentation loading and demonstrated that the skull-brain interface substantially affects the mechanical response of brain tissues. 24 Considering that the skull-brain interface acts as a fundamental layer of protection for the brain, in vivo measurements of the biomechanics at the interface could provide insight into the response of the brain-protective system to subconcussive RHI exposures. Further, such measurements have the potential to increase the precision of computational models in modeling the skull-brain interface, which currently presents a challenge because of limited experimental data. 25
Although the skull-brain interface cannot be directly visualized with conventional imaging modalities, its functionality can be evaluated by analyzing the biomechanical changes at the point of interface. In particular, the skull-brain interface plays a crucial role in dampening the motions by acting as a shock absorber. This can be evaluated by calculating the transmission of motion at the interface. Moreover, the skull-brain interface functions as a stabilizing mechanism, limiting the brain's motion and preventing excessive deformation. This stabilizing, or isolation capability, can be assessed by measuring the strain field of the brain close to the skull-brain interface.
To study the motion transmissibility of the skull-brain interface, a non-invasive imaging technique, magnetic resonance elastography (MRE), has been employed to measure the brain's responses from dynamic loadings. 26 –28 In brain MRE, vibrating the head via an MRE head driver transmits dynamic mechanical vibrations (∼tens of microns) into the brain. A phase-contrast MR pulse sequence is then performed to encode the brain displacement into MR phase signals. In previous studies, the brain motion was directly measured from MRE, but the skull motion was indirectly assessed using accelerometers or pressure sensors, which is uncomfortable and can introduce measurement errors related to mouthguard fit and even saliva accumulation. 28,29 In this study, we utilized a recently developed dual saturation and dual-sensitivity motion encoding (DSDM)-MRE sequence to simultaneously image the skull and brain motion. 30 Based on this technique, the motion transmission and attenuation from the skull to the brain were measured by calculating the rotational motion transmission ratio (Rtr ). The Rtr is defined as the brain-to-skull amplitude ratio of the rigid-body rotation. Our preliminary study has shown a significant difference in Rtr between the ice-hockey players and healthy volunteers. 31
Besides the motion transmission, strain-based analysis has been applied to study the isolation function of the skull-brain interface by analyzing the brain deformation patterns. Tagged MRI sequences measured the brain deformation and strain under impulsive loadings and reported regions of smaller displacements reflecting the skull-brain isolation effects. 20,32,33 Our previous MRE study also showed that the skull and brain interface does not exhibit a complete slip behavior, but rather a weak and heterogeneous connection, based on qualitative visual inspection. 30 In this study, the cortical Normalized Octahedral Shear Strain (NOSS), an MRE-based parameter, has been calculated to quantify the brain surface deformation near the skull-brain interface, serving as a useful indicator of the skull-brain isolation capability. Octahedral Shear Strain (OSS) is the norm of the deviatoric strain tensor for quantifying the magnitude of deformation, which is used because of its consideration of shear deformation in all three dimensions and its capability to yield the maximum shear stress in any plane, independent of the orientation of the coordinate system. 34 NOSS normalizes the OSS to the wave displacement to reduce bias from the wave amplitude variations. Together with the Rtr , these variables may be able to quantify the mechanical interactions of the skull-brain interface.
The goals of this study were (1) to apply two MRE-based parameters, Rtr and cortical NOSS, to assess the transmissibility and isolation capabilities of the skull-brain interface; (2) to perform a test-retest study to determine the MRE driver dependence and repeatability of the proposed acquisition and analysis techniques; and (3) to investigate the effects of age and sex on Rtr and cortical NOSS in healthy volunteers.
Methods
Study participants
With Institutional Review Board approval and written informed consent, 47 healthy volunteers (age range: 18–56 years, 32.6 ± 10.3 years, 15F/32M) with (1) no history of TBI and (2) no self-reported history of contact-sports participation were recruited. To evaluate the performance of our proposed method regarding MRE driver dependence and repeatability, 17 of 47 healthy volunteers (age range: 18–56 years, average age: 35.6 ± 11.2 years, 5F/12M) were scanned three times under vertical (anterior-posterior direction, AP) vibrations: the first scan was collected during the first visit, and the following two scans were collected after a one-month follow-up, where these two scans were separated by a 5-min break with the volunteer leaving the scanner. Ten participants received three additional scans with horizontal (left-right direction, LR) vibrations to evaluate the direction dependence of the proposed technique.
MRE/MRI scans and data acquisition
All MRE/MRI examinations were performed on a recently developed high-performance compact 3T MR scanner. 35,36 Mechanical vibrations at 60 Hz were introduced into the participant's head using a commercially available pneumatic active driver (Resoundant, Rochester, MN) and a custom-made, soft, pillow-like multi-excitation head passive driver. The multi-excitation driver introduced two distinct vibration directions: a vertical (AP direction) vibration with the driver positioned under the participant's head; and a horizontal (LR direction) vibration with the driver positioned along the right side of the participant's head (Fig. 1A). The position of the side LR driver was adjusted by an attached plastic rod to ensure contact between the head and the driver. Memory foam pads (not shown) were placed between the head and the inside of the brain coil to prevent bulk movement of the head during the scan.
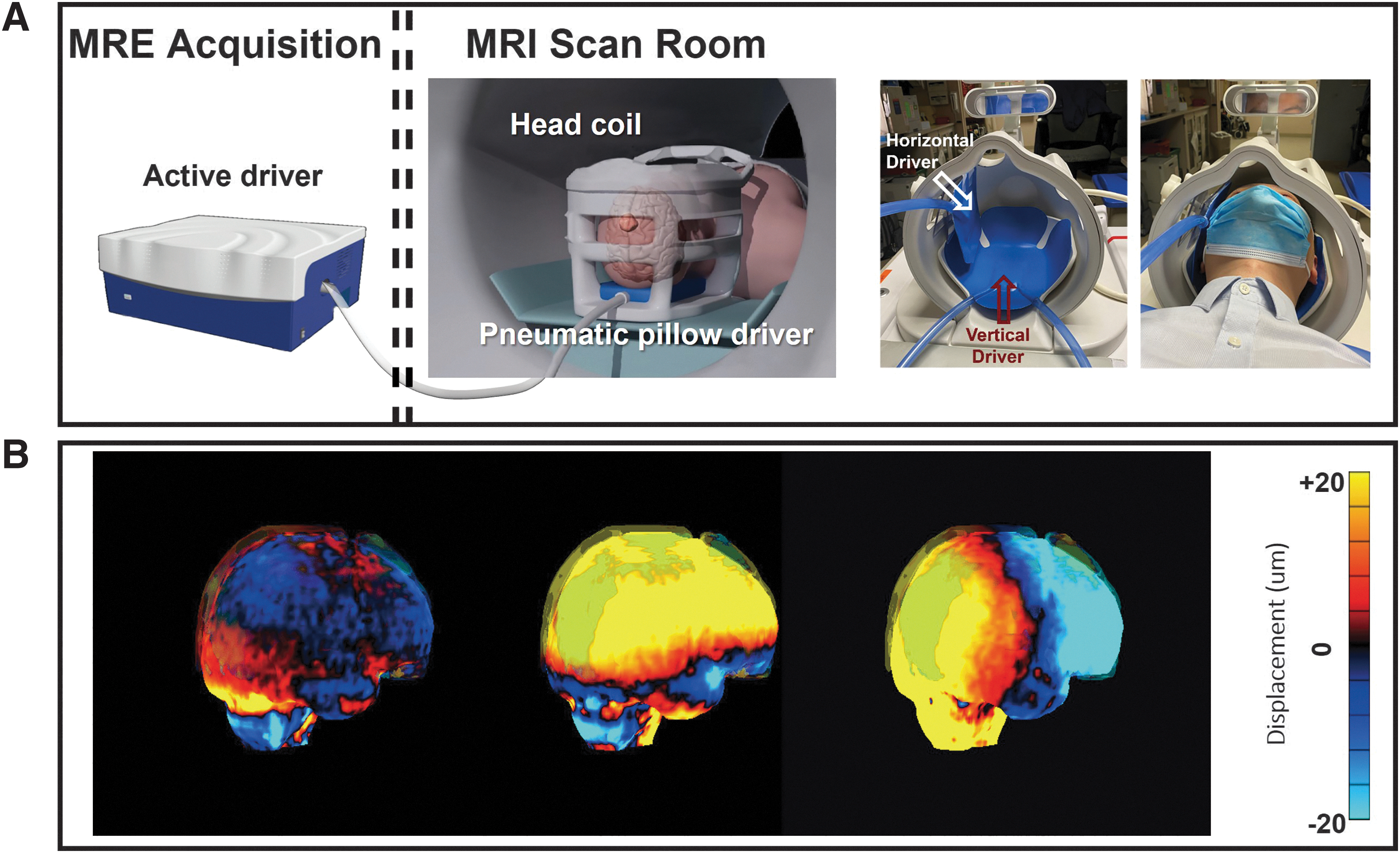
(
The resulting full three-dimensional (3D) displacement vector field (Fig. 1B) was acquired using a single shot, spin-echo (SE), echo-planar imaging (EPI) based DSDM-MRE pulse sequence 30 with the following parameters: repetition time (TR)/ echo time (TE) = 4000/58.7 msec; field of view (FOV) = 24 cm; matrix size = 80 × 80; 48 contiguous 3-mm-thick axial slices; 2 × ASSET acceleration; four phase offsets sampled over one period of excitation; and total acquisition time for each MRE measurement: 3:20 min. The motion encoding gradients (MEGs) were employed in the x- (frequency encoding, left-to-right), y- (phase encoding, anterior-to-posterior), and z- (slice selection, superior-to-inferior) directions to record the full displacement field. A slightly different encoding efficiency between positive and negative encoding procedures was used to generate dual-sensitivity phase images. 30
The 3D T1-weighted and axial T2-weighted sequences were also performed for brain segmentation and to guide the atlas transformation between the T1 and MRE image volumes. The 3D T1-weighted magnetized prepared rapid gradient echo (MP-RAGE) pulse sequence was acquired with the following parameters: sagittal plane, TR/TE = 6.1/2.5 msec, 8 degree flip angle, inversion time (TI) = 900 msec, FOV = 26 cm, 256 × 256 imaging matrix, and 166 slice locations with 1.2-mm spacing. The T2-weighted image was acquired with a fast SE pulse sequence with the following parameters: axial plane, TR/TE = 4785/102 msec, 125 degree flip angle, echo train length (ETL) = 16, FOV = 24 cm, 256 × 256 imaging matrix, 2 NEX, and 72 slice locations with 2-mm spacing.
Data processing and analysis
Skull and brain displacement calculation
The full 3D skull and brain displacement was calculated as described previously. 30 Briefly, the skull (fat) and the brain (water) images were combined in the complex image domain. Dual sensitivity phase images were calculated by adding or subtracting the signals from positive and negative MEGs. The high-sensitivity phase data (with phase wrap) were unwrapped with the guidance of the low-sensitivity phase images (no phase wrap), because the ratio between the high and low encoding sensitivity is known. The x-, y-, and z- motion displacements at each MRE phase offset were then calculated from the unwrapped high-sensitivity MRE phase.
Brain-to-skull rotational motion transmission ratio (Rtr) calculation
Skull and brain segmentation
The automatic skull and brain segmentation was performed using a customized MATLAB program (The MathWorks, Natick, MA) that integrates the FSL brain extraction tool from the functional MRI of the Brain (FMRIB) Software Library (London, UK) 37 and mathematical morphological operations 38 to segment the MRE magnitude data into the skull, scalp, and brain. Visual evaluations were performed to ensure accurate segmentation of the skull and brain in all 47 volunteers.
Rigid-body fitting and region of interest (ROI) selection
Given the segmentation results, the displacement vectors in the brain and skull were separately fitted to a rigid body model with the origin set as the center of mass of the cerebrum (Fig. 2A). 30 To study the mechanical interactions at the skull-brain interface, a 9-mm-thick shell ROI was used for the rigid-body fitting of the brain, located immediately interior to the edge of the brain mask, and slices covering the entire cerebrum were taken.
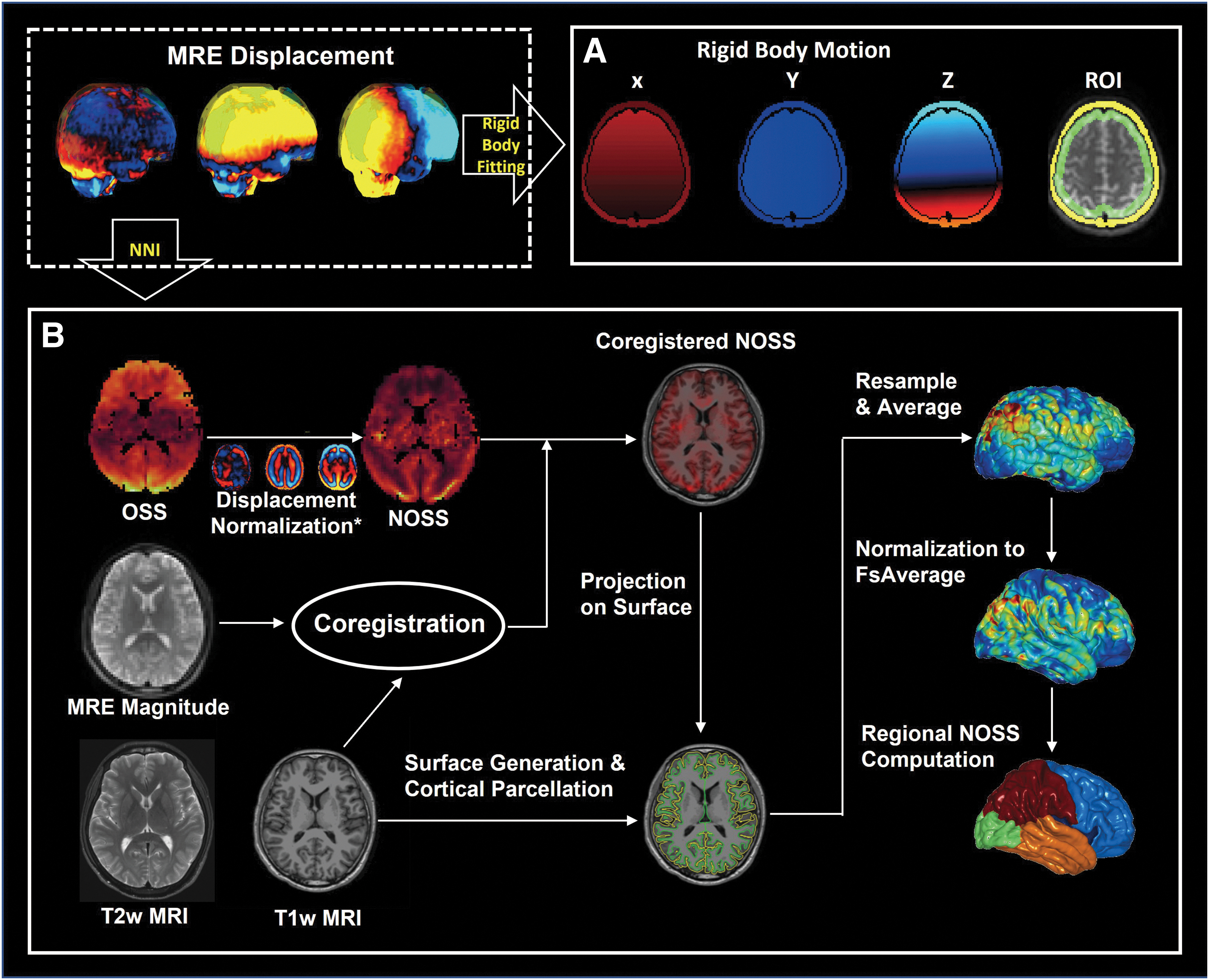
The pipeline of (
Brain-to-skull Rtr calculation
Given that rotational motion is the primary cause of brain deformation and injuries, 39 we focused mainly on the rotational part of the motion to evaluate the functionality of the skull-brain interface in damping the motions. As the trajectory of the rotation motion with the harmonic vibration was an ellipse, 30 we used the dominant component (square of the major axis of the ellipse) to represent the motion amplitude. The Rtr (defined as the brain-to-skull amplitude ratio of the rigid-body rotational motion) was calculated for each volunteer.
Cortical normalized octahedral shear strain (NOSS) calculation
The deformation of the brain near the skull-brain interface was assessed by calculating the NOSS across the cortical surface. The NOSS has demonstrated great potential in representing the adhesive status between two adjacent layers. 40 Hence, the NOSS was calculated at the cortical level here to assess the deformation of the brain surface.
NOSS calculation using artificial neural network
To address the challenge of strain computation near the skull-brain interface, where noise and partial volume effect are prevalent, a neural network-based algorithm (DiffNet) was developed to compute the partial derivatives of harmonic MRE displacements for OSS calculation. DiffNet uses the wave equation
With DiffNet, the full-volume OSS maps were generated with the partial derivatives computed by the neural network. The NOSS was calculated to reduce the amplitude variations by normalizing the time-averaged OSS to the wave motion amplitude (square root of the sum of squares of the amplitude of wave motion in each direction) at each voxel. 40 Here, the wave motions were obtained by applying a Gaussian low-pass filter with a cutoff = 1 wave/FOV to the full MRE displacement data to remove bulk motion.
To determine the degree of correlations between traditional numeric calculations and the DiffNet, we performed a comparison study where the NOSS values were computed using the central difference and the DiffNet, respectively. The results are summarized in the supplemental materials (Fig. S2).
Cortical segmentation and cortical NOSS estimation
To evaluate the shear deformation in cortical regions, we used FreeSurfer (version 6.0) 44 to generate and visualize cortical NOSS distribution with the following steps (Fig. 2B):
(1) Surface generation and cortical parcellation. The recon-all pipeline was used with both T1- and T2-weighted images for each subject to generate cortical surfaces and do the cortical segmentation.
(2) MRE magnitude images and T1-weighted MRI images coregistration. The registration was implemented by a rigid body registration with six degrees of freedom. The transformation matrix was saved for next-step NOSS resampling.
(3) NOSS maps to structure alignment and surface NOSS resample. The NOSS maps corresponding to the MRE magnitude images were aligned to the anatomical images with the transform matrix, and the NOSS values were sampled equidistantly along the surface normal. The mean cortical NOSS was obtained by averaging the sampled NOSS values along the cortical ribbon and was then overlayed on the cortical surface for visualization.
(4) Normalization and smoothing. To allow intra-subject comparison, we normalized the cortical NOSS maps to the standard space FsAverage. The NOSS value at each vertex was then averaged with its neighbors by setting the smoothing iteration to 5.
(5) Regional analysis. The Desikan-Killiany atlas 45 was used for the regional NOSS calculation, comparison, and statistical analysis. The frontal, temporal, parietal, and occipital cortical regions were selected for analysis.
Statistical analysis
The OSS-SNR 46 was calculated for each subject to exclude the cases with low wave amplitude. The cutoff of the OSS-SNR was set as 6 to ensure the high-quality phase data were included, resulting in the exclusion of four cases, although 3 is commonly used for stiffness estimation. The mean cortical NOSS values in the cerebrum, frontal, temporal, parietal, and occipital lobes were calculated for each volunteer. A paired two-sided Wilcoxon signed-rank test was performed on Rtr and cortical NOSS to identify differences under two vibration orientations. The within-day and between-day repeatability were analyzed by calculating intraclass correlation coefficients (ICCs) with 95% confidence intervals (CI). An ICC between 0.6 and 0.75 is considered fair, and ≥0.75 is considered excellent. 47 To study the age/sex effect, the two measures (NOSS and Rtr ) were separately fitted to a multiple linear regression model, both using the independent parameters of age and sex. Statistical significance was defined as p < 0.05.
Results
Rtr and cortical NOSS are independent of driver orientation
Figure 3A shows the comparison results of the Rtr under AP and LR vibrations. There is no significant difference between the two distinct vibration orientations (mean Rtr with AP = 0.59, mean Rtr with LR = 0.61, p = 0.85), demonstrating that the brain-to-skull rotational transmission is independent of the driving mechanism.
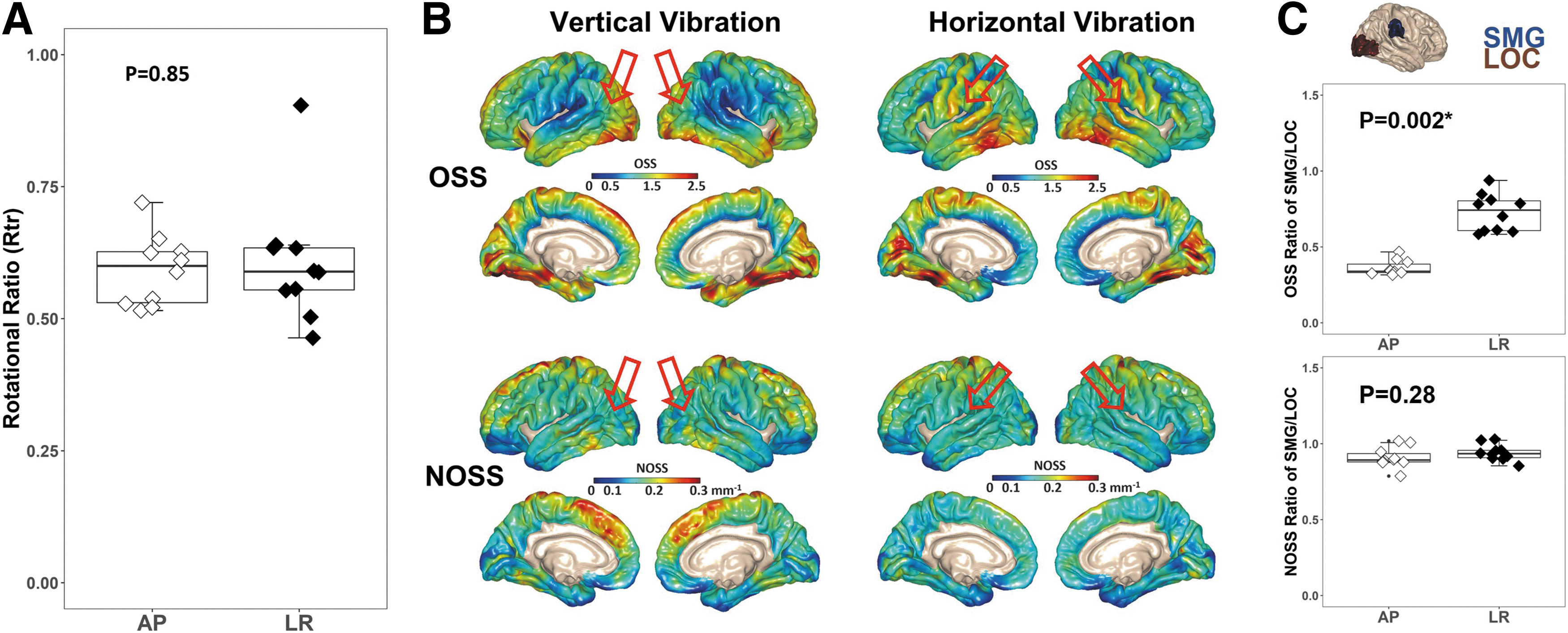
Driver effects on rotational motion transmission ratio (Rtr
) and cortical strain calculation. (
Figure 3B shows the averaged OSS and NOSS distributions across all subjects under two vibration orientations. Although the driver location dominates the OSS distribution, where higher OSS values are present in the areas corresponding to passive driver locations, the regional variations of NOSS are similar across the brain surface between two vibrations, showing that normalization reduces the bias induced by the driver orientation. To further test the effects of the vibration orientation, strain ratios between two subcortical regions (supramarginal gyrus [SMG], lateral occipital cortex [LOC]) that are close to the driver locations were calculated and compared (Fig. 3C). As expected, there is a significant difference between the two vibration orientations in the OSS ratio of the SMG/LOC (mean OSS ratio with AP = 0.36, mean OSS ratio with LR = 0.73, p = 0.002), whereas the difference is much smaller after normalization (mean NOSS ratio with AP = 0.91, mean NOSS ratio with LR = 0.94, p = 0.28).
High repeatability of Rtr and cortical NOSS characterization
The test-retest study of Rtr shows excellent repeatability for between-day (ICC ranges from 0.84 to 0.95) and within-day (ICC ranges from 0.95 to 0.96) measurements either under AP or LR vibration (Table 1). Figure 4 shows the voxel-wise ICC values of cortical NOSS across the three different MRE scans with AP vibrations. The within-day (D2-1 vs. D2-2) and between-day (D1 vs. D2-1, D1 vs. D2-2) results show good to excellent repeatability in most regions. Table 2 summarizes the regional ICC values of cortical NOSS in different brain lobes, further indicating a high test-retest repeatability for between- (ICC ranges from 0.68 to 0.91) and within-day (ICC ranges from 0.80 to 0.94) measurements.
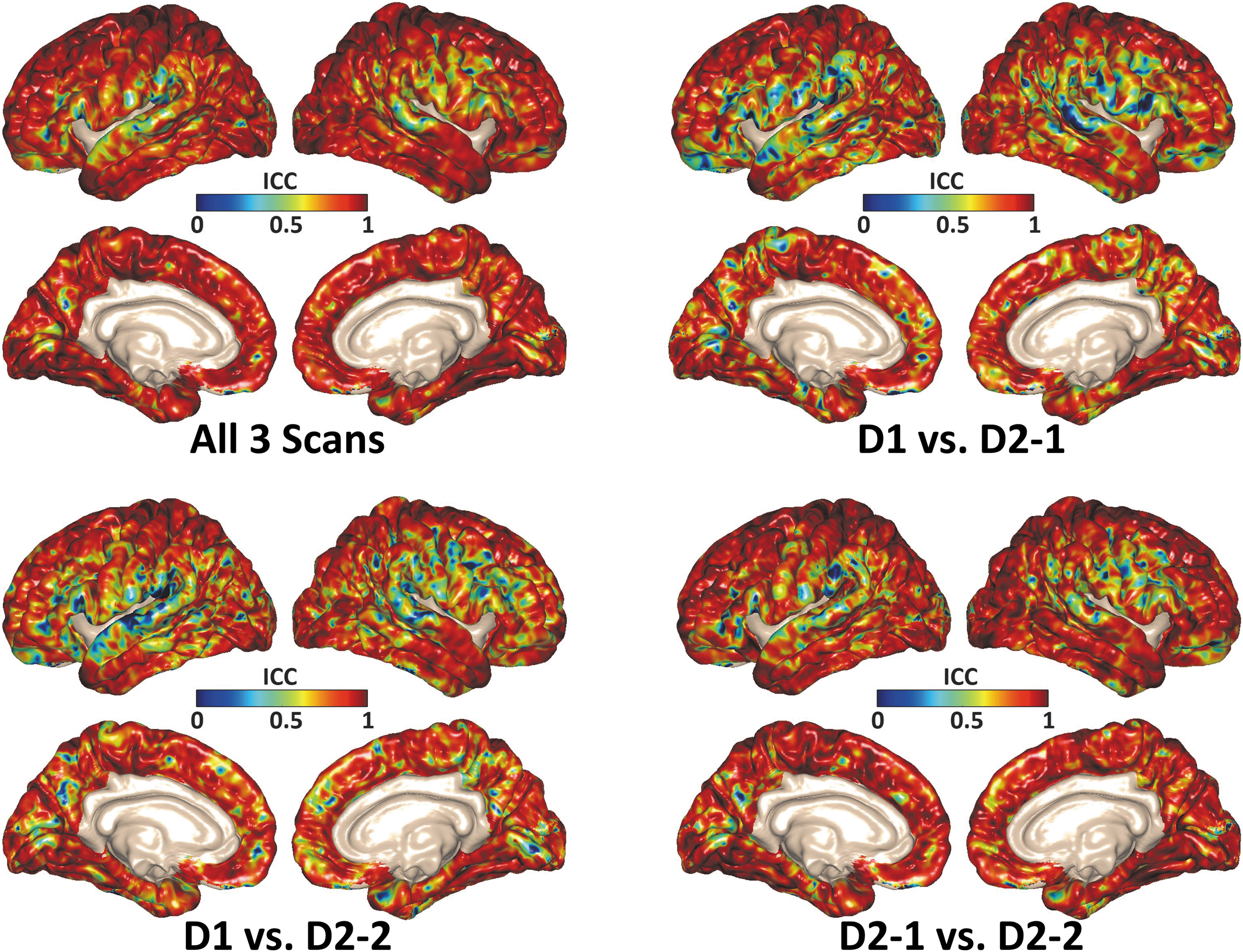
Pixel-wise intraclass correlation coefficient (ICC) values of cortical normalized octahedral shear strain (NOSS) calculated through all three scans, between-day scans (D1 vs. D2-1, D1 vs. D2-2) and within-day scans (D2-1 vs. D2-2). These results demonstrated the excellent repeatability of the proposed cortical NOSS calculation.
Repeatability Statistics of Rtr Under Different Vibration Modes
Rtr, rotational motion transmission ratio; ICC, intraclass correlation coefficient; CI, confidence interval.
Repeatability Statistics of Cortical NOSS Among Different Brain Lobes
NOSS, normalized octahedral shear strain; ICC, intraclass correlation coefficient; CI, confidence interval.
Sex/Age dependence of Rtr and cortical NOSS
Figure 5A illustrates the correlation between the Rtr and age/sex. Overall, we found no significant age or sex effects on the Rtr . Figure 5B shows the cerebrum cortical NOSS changes across different age groups. There is no significant NOSS change among individuals younger than 30 years. Meanwhile, for individuals >30 years, the cerebrum cortical NOSS is positively correlated with age for both men and women, as shown in Figure 5C. A plot of cortical NOSS as a function of age and sex is shown for the cerebrum (Fig. 5C), frontal lobes, temporal lobes, parietal lobes, and occipital lobes (Fig. 5D) for volunteers over 30 years old. Except for the occipital lobe, we observed an increasing trend of the mean NOSS with respect to age.

Age/sex dependence of rotational motion transmission ratio (Rtr) and cortical normalized octahedral shear strain (NOSS). (
The detailed multiple linear regression model results are summarized in Table 3. A sex effect was found in the temporal lobe, where females had relatively higher NOSS than males, but not observed in the other regional NOSS analyses. For visualization of NOSS changes with age, the fitted slope rates in four lobes are projected on the cortical surface (Fig. 5E). We observed that the region with the most rapid age-dependent changes in NOSS is the frontal lobe (slope rate = 0.0005 ± 0.0002 mm^-1/year), and the NOSS values were not significantly associated with age in the occipital lobe (slope rate = 0 ± 0.0001 mm^-1/year). We also found that NOSS in superior regions was significantly higher than NOSS in inferior regions (Fig. S3).
Parameters of NOSS Linear Regression Model for People Over 30 Years
NOSS, normalized octahedral shear strain.
Discussion
In this study, we have developed an MRE framework to calculate two MRE-assessed parameters, Rtr and cortical NOSS, that represent two important features of the skull-brain mechanical interactions—i.e., motion transmission and isolation capability, respectively. Calculation emphasis was placed on regions near the skull-brain interface, because changes in its functionality can significantly impact cortical biomechanics. In contrast, the mechanics of deeper brain regions are more intricate, potentially impacted by deep structures (e.g., ventricles) and brain properties (e.g., wave attenuation from the brain's viscosity). We have demonstrated the robustness of these two parameters with a repeatability study using two vibration orientations. We also investigated their correlations with age and sex in a cohort of healthy volunteers.
When the head moves, the relative motion between the inner skull and brain as well as the deformation on the cortical surface of the brain result in a mechanical (compression and shear) loading on the skull-brain interface. Conversely, the mechanical properties of the skull-brain interface will significantly affect the measured skull/brain motion and the deformation of the cortical surface.
Our previous preliminary work has shown significantly elevated Rtr values among ice-hockey players compared with healthy controls. 31 The higher the Rtr , the more harmful rigid body rotation is directly transmitted to the brain. Given the age difference between the hockey group (18 ± 1 y) and healthy controls (43 ± 18 y), it was not known whether the differences arose from RHI or an age effect, or both. In this current study, we found no age effect in Rtr . Based on the above evidence, Rtr may have clinical potential for RHI detection: the high Rtr values (among ice-hockey players) may indicate the attenuation capability of the skull-brain interface is degenerated because of RHI exposure. Regarding translational motion transmission, our previous study showed that the amplitude of translational motion was barely changed through the skull-brain interface. 30
Cortical NOSS was found to increase with aging, which may suggest that the isolation of the skull-brain interface is degraded in the older population. In such a situation, the brain surface may deform more easily and thus undergo a larger shear strain. A softer cortex in aging people 48 may also partly contribute to the high NOSS, because soft tissue will deform more under the same impact level. It is widely acknowledged that the frontal lobe is more vulnerable to injury and deformation because of its location at the front of the cranium, proximity to the sphenoid wing, and large size. A recent in situ cadaver experiment revealed a 12% increase in the density of arachnoid near the vertex (primarily comprising the frontal lobe) compared with inferior brain regions. 49 This disparity in density may suggest an evolutionary adaptation to protect the frontal lobe. It is speculated that the degradation of the trabeculae's mechanical properties, potentially resulting from impaired arachnoid trabeculae, may lead to the degeneration of the skull-brain interface's isolation capability, particularly in areas with a high density of arachnoid like the frontal lobe, which could lead to a faster rate of age-related NOSS changes in the frontal lobe. Directly measuring the mechanical properties of the in vivo PAC, however, is quite challenging, and further investigation is warranted. We are currently recruiting volunteers with a history of RHI exposure to investigate the feasibility of NOSS as a biomarker for RHI exposure.
Regional strain variations have been reported in other studies. Chan and coworkers 33 calculated strain distributions in the cortical gray matter using tagged MRI. They reported greater strains in the temporal lobe as well as the frontal and parietal cortex in the opposite hemisphere of the applied loading, but lower strains in the occipital lobe. The strains in Chan's study were not normalized to the wave amplitude, and as a result, the unnormalized strain distribution they observed could have been affected by the impact location.
In the early stage of CTE, phosphorylated tau (p-tau) accumulation is found at the depths of the sulci, and some research has suggested that this phenomenon may be caused by higher strains in the sulci. 50 Conversely, we found that the gyral regions exhibit high NOSS values instead of the sulcal regions. Okamoto and colleagues 26 also reported no elevated strain magnitudes in the sulci compared with other cortical areas. This may be because of the different impact modes (dynamic vs. transient) and scales (microns vs. centimeters) between the MRE and the actual sub-concussive/concussive level impact.
The vibration amplitude, wave propagation direction, and transmission efficiency of the external loading may vary substantially between examinations, which could lead to inconsistent estimates of OSS, as shown in Figure 3B. The test-retest results showed that NOSS was less sensitive to the experimental setup, suggesting NOSS is a more robust biomarker.
A neural network-based partial differential method was proposed in this study to mitigate the calculation errors near the brain surface from noise and partial volume effect. In particular, we expect the network to produce less biased estimates of the strain in tissue adjacent to the falx cerebri, which acts as a shear wave source and results in a displacement discontinuity. The network, by computing the partial derivatives with data from only one hemisphere at a time, avoids this discontinuity and is substantially less affected by missing measurements than the finite difference estimate. It is also worth noting that the data used in this study were acquired from a high-performance compact 3T MRI scanner, which provides higher SNR images compared to a standard 3T scanner. Given the superior performance of DiffNet in simulation studies, DiffNet could perform better when dealing with noisy data (Fig. S4, S5). In addition, increasing spatial resolution could be a potential approach to improve strain estimates. However, this would also result in a reduction of image SNR and an associated increase in bias when calculating derivatives. Our future studies will explore the potential of DiffNet to effectively address these limitations.
The findings of this study have the potential to contribute to the development of more accurate models for the skull-brain interface in computational models of brain injury. Our findings support the widely accepted notion that the skull-brain interface can be modeled as a single layer of viscoelastic tissue, as evidenced by a Rtr measurement of less than 1. In addition, the results suggest that there is a non-uniform distribution of cortical NOSS, implying a heterogeneous skull-brain interface that may contribute to distinct patterns of brain deformation during impact events. To enhance the accuracy of TBI computational model predictions, the spatial variability of the skull-brain interface across the brain may need to be taken into account. Our previous research has also indicated that the skull–brain interface cannot be considered as a completely frictionless connection, but rather as a weak connection with spatial heterogeneity. 30 Based on these findings, it is recommended that the skull-brain interface should be modeled as a viscoelastic tissue with spatial heterogeneity in the skull-brain “connections.”
Several limitations of this study must be considered. (1) Relatively low resolution (3-mm isotropic) was used in this study. (2) The voxel-wise ICCs in the temporal lobe were lower than in other regions, which may be partly because of susceptibility-induced distortion in the EPI pulse sequence. These could be addressed in future studies by acquiring high-resolution distortion-free MRE images with multi-band DIADEM-MRE 51 to improve the accuracy of cortical registration between MRE and anatomical images. (3) The aging study was limited to healthy volunteers <60 years old. A larger cohort of participants with a wider age range will further advance our findings. (4) Participants with RHI exposures are needed in future studies to test the clinical performance of the two proposed parameters in identifying early RHI exposure. (5) All experiments were performed under a single frequency (60 Hz) vibration, and further studies using multi-frequency MRE would be beneficial in exploring the frequency-dependent response of the skull-brain interface.
Conclusion
MRE-assessed measurements of the mechanical dampening and isolation capability of the skull-brain interface (Rtr and cortical NOSS) demonstrated high repeatability and independence of driver mechanisms. Cortical surface shear (NOSS) is positively correlated with age, consistent with gradual degradation of the isolation capacity of the pia-arachnoid complex over time. This work provides quantitative biomarkers for evaluating the biomechanical isolation function of the skull-brain interface to facilitate the study of RHI effects and potential susceptibility to subsequent acute TBI.
Footnotes
Acknowledgment
The authors wish to acknowledge all volunteers who participated in the study.
Authors' Contributions
Xiang Shan: Software, Investigation, Formal analysis, Writing - Original Draft. Matthew C. Murphy: Software, Writing - Review & Editing. Yi Sui: Writing - Review & Editing. Emanuele Camerucci: Writing - Review & Editing. Keni Zheng: Writing - Review & Editing. Armando Manduca: Writing - Review & Editing. Richard L. Ehman: Funding acquisition, Project administration, Writing - Review & Editing. John Huston III: Funding acquisition, Project administration, Writing - Review & Editing, Resources. Ziying Yin: Conceptualization, Software, Supervision, Funding acquisition, Project administration, Writing - Review & Editing.
Funding Information
This work was supported by grants from the NIH (R01 NS113760, R01 EB001981, and U01EB024450) and Mayo Clinic Imaging Awards CIM-92541587 and CIM-92541650.
Author Disclosure Statement
M.C.M., A.M., J.H., R.L.E., and Mayo Clinic have a financial interest in MRE technology. For the remainng authors, no competing financial interests exist.
Transparency,Rigor and Reproducibility Summary
The study was registered after the study began. The study is registered at the Mayo Clinic Clinical Trials (
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.