
Abstract
Cerebrovascular dysfunction following traumatic brain injury (TBI) is a well-characterized phenomenon. Given the therapeutic potential of xenon, we aimed to study its effects after localized delivery to the brain using microbubbles. We designed xenon-containing microbubbles stabilized by dibehenoylphosphatidylcholine (DBPC) and polyethylene glycol (PEG) attached to saturated phospholipid (DPSE-PEG5000). Using a pig model of TBI, these microbubbles were intravenously injected, and ultrasound was used to release xenon at the level of the carotid artery. The control group received perfluorobutane containing microbubbles. Diffusion tensor imaging (DTI) showed areas of higher fractional anisotropy for pigs receiving xenon microbubbles compared to the control group at 1 day after injury. Radial diffusivity analysis showed that this effect was mainly the result of acute edema. Pigs were euthanized at 5 days, and the brain tissues of xenon-treated animals showed reduction of perivascular inflammation and blood–brain barrier disruption. Endothelial cell culture experiments showed that glutamate reduces tight junction protein zona occludens-1 (ZO-1), but treatment with xenon microbubbles attenuates this effect. Xenon treatment protects cerebrovasculature and reduces astroglial reactivity after TBI. Further, these data support the future use of localized delivery of various therapeutic agents for brain injury using microbubbles in order to limit systemic side effects and reduce costs.
Introduction
Traumatic brain injury (TBI) is one of the leading causes of death and disability throughout the world. Various pharmacological and neuromodulatory strategies have been proposed over the last 30 years of TBI research, but no consistent therapeutic effect has been demonstrated in the clinical trials.
1
One of the promising potential therapeutic agent is xenon, a noble gas that was first reported to have anesthetic properties in 1951.
2
It has low reactivity and stability as a noble gas, and it is eliminated quickly from the body because of its low blood-gas partition coefficient.
3
Xenon has neuroprotective effects via the N-methyl-
Methods
Microbubble formulation
As previously described, we have developed a method of encapsulating xenon gas in phospholipid microbubbles (MB) of sizes <10
Animal surgery
All animal experiments were approved by the Institutional Animal Care and Use Committee at the University of Pennsylvania, in accordance with the Guide for the Care and Use of Laboratory Animals. These experiments were in compliance with the Animal Research: Reporting of in Vivo Experiments (ARRIVE) guidelines. We performed controlled cortical impact (CCI) injury with ∼7–10 kg, 4-week-old pigs as previously described. 12 After intramuscular (IM) injection of ketamine (20 mg/kg) and xylazine (2 mg/kg), pigs were induced with 4% inhaled isoflurane. They were subsequently intubated and maintained at 1% isoflurane throughout the experiment. For analgesia, bupivacaine was injected in the subcutaneous tissue at the intended incision site. Pigs also received cefazolin IM for prophylaxis. Pigs were subjected to central venous catheter (CVC) placement in cephalic veins, terminating in the superior vena cava. They then underwent right-sided craniotomy overlying the rostral gyrus. This was followed by moderate severity TBI with a spring-loaded impactor at 4 m/sec. Prior to the end of the procedure, one dose of Buprenorphine-SR was administered for analgesia. They were treated with 4 mg/kg of intravenous diphenhydramine 20–30 min prior to injection of MB in order to prevent any allergic reaction. They were randomized into either CtMB or XeMB groups, and they received either type of MBs at 1, 3, and 24 h after TBI using continuous infusion pump at ∼1 ± 0.5 x 108/mL concentration. Infusion was performed three times with freshly diluted MBs at each time point, with the infusion rate raised steadily from 0.2 mL/min to 0.6 mL/min over 8 min each time, with ∼4 mL being delivered. This administration schedule of three administrations within 24 h was devised to simulate a potential treatment plan for a clinical TBI subject, as multiple treatments within an acute window after TBI would potentially have the most efficacy. A portable ultrasound transducer (Lumify, Philips Healthcare) was placed at the level of carotid artery with a mechanical index of 1.0 following CVC injection of the MBs in order to release the xenon contents. Additionally, in the post-operative period, pigs were monitored daily for signs of infection and pain. Additional doses of Buprenorphine were administered as needed based on pain assessment. For euthanasia on day 5, animals were anesthetized with an IM injection of ketamine/xylazine followed by inhaled isoflurane. They then received an intracardiac injection of pentobarbital at 150 mg/kg.
Diffusion imaging
As previously described, 13 diffusion tensor imaging (DTI) was performed using a 3T Tim Trio whole body magnetic resonance (MR) scanner (Siemens, Germany) with a 12-channel phased array head coil. The sequence parameters were as specified: repetition time (TR) = 4200 ms, echo time (TE) = 103 ms, flip angle = 180 degrees, bandwidth = 1186 Hz/pixel, field of view (FOV) = 192 mm, slices thickness = 2mm, number of slices = 24, voxel size = 2 x 2 x 2 mm, b-values = 0, 1000, 2000 sec/mm2, with 64-non-collinear/non-coplanar directions. A composite skeleton was made for the entire cohort, followed by outlining of the region of interest (ROI) for the corona radiata (both ipsilateral and contralateral), cerebral peduncle (both ipsilateral and contralateral), genu of corpus callosum, and splenium of corpus callosum by a blinded analyzer. Fractional anisotropy (FA), mean diffusivity (MD), radial diffusivity (RD), and axial diffusivity (AD) were calculated for reach ROI.
Tissue histology
On day 5, anesthetized animals were euthanized, and the brain was removed whole. The brains were cut into standardized 5 mm coronal sections and fixed in 10% neutral buffered formalin. Following a fixation period of at least 2 weeks, whole brain slices were processed, paraffin embedded, cut into 5 μm sections, and stained with hematoxylin and eosin (H&E) and immunohistochemical stains for GFAP (Sigma-Aldrich, AB5804, 1:300) and Iba1 (Fisher Scientific, NC9288364, 1:500). All histopathology studies were performed by a blinded, board-certified neuropathologist (A.N.V.). H&E-stained histological sections of both hemispheres were examined and used to identify the lesion (ipsilateral = same side as the lesion) and to score perivascular inflammation and reactive vascular changes. Vascular changes including endothelial proliferation were scored as follows: 0 = no vascular changes; 1 = reactive endothelial cells; 2 = endothelial cell proliferation (more than one cell layer thick). Perivascular inflammation was scored as follows: 0 = no to rare inflammatory cells; 1 = a few inflammatory cells; 2 = cuff of inflammatory cells 1 cell layer thick; 3 = cuff of inflammatory cells more than one cell layer thick. Perivascular inflammation and vascular changes were scored in the ipsilateral cortex surrounding the lesion and in the contralateral cortex. GFAP and Iba1 staining were quantified as the total percent area stained using ImageJ software. Because one of the animals in the CtMB group had problems with histological processing, it was not used for the analysis. Therefore, the number of animals for histology displayed in this manuscript was four for the CtMB group, and eight for the XeMB group. The percent area stained was measured in the cortical gray matter and underlying white matter for both the ipsilateral and contralateral hemispheres.
For fibrinogen staining, tissue was deparaffinized in xylene and rehydrated in a series of ethanol solutions. Antigen retrieval was performed in sodium citrate buffer pH 6.0 in a Biocare Medical Decloaking Chamber (SKU: DC 2012) for 10 min at 110°C, followed by a 3% bovine serum albumin (BSA) block for 1 h at room temperature and a 30 sec block in 1% Sudan Black. Blocking was followed by incubation with the primary antibody Recombinant Anti-Fibrinogen Antibody [1F2] (1:700, Cat# ab58207, abcam) diluted in 3% BSA/1x PBS/0.1% Triton-X for 1 h at room temperature. After washing in 1x PBS, an anti-mouse Alexa Fluor 568 secondary antibody (1:500, Cat.# A11031, Invitrogen) diluted in 3% BSA/ 1xPBS/0.1% Triton-X was applied for 1 h at room temperature. Slides were mounted and cover-slipped using ProLong Gold Antifade Mountant with DAPI (Cat.#: P36941, Invitrogen) and imaged on a Leica DMi8 microscope.
Endothelial cell culture and immunocytochemistry
Mouse brain endothelial cells (BEND3, CRL-2299) were purchased from American Type Culture Collection (ATCC, Manassas, VA, USA), They were cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco, Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS; Gibco, Waltham, MA, USA) and 100 U/mL penicillin as well as 0.1 mg/mL streptomycin (Gibco, Waltham, MA, USA), maintained at 37°C in a humidified atmosphere with 5% CO2. The rabbit polyclonal anti-ZO-1 tight junction protein antibody (ab216880), FITC-conjugated goat anti-rabbit antibody (ab6717), and normal goat serum (ab138478) were purchased from abcam (Cambridge Biomedical Campus, Cambridge, UK). The expression levels of Zonula occludens (ZO)-1 tight junction proteins in BEND3 cells were assayed by immunocytochemistry. Briefly, BEND3 cells were grown on cover-slips until 80% confluent. The cells were treated with or without glutamate (80 mM) for 24 h. The cells were then treated with XeMB or CtMB (20% of the total volume of cell culture media) for 24 h. After completion of the treatment, the cells were washed three times with PBS (each 5-min wash).
For immunocytochemistry, the cells were incubated in 10% goat serum in PBS for 30 min followed by anti-ZO-1 antibody (1:100 dilution) overnight at 4°C. The cells were washed three times in PBS, 5 min each wash. Then the cells were incubated in FITC-conjugated goat anti-rabbit antibody (1:1000 dilution) for 1 h at room temperature in the dark. The cells were then washed three times in PBS, 5 min each wash, and mounted on glass slides with a mounting medium counterstained with DAPI. The cells were visualized under confocal microscopy. Images were acquired and analyzed for ZO-1 expression levels using ImageJ.
Statistical analysis
GraphPad Prizm (GraphPad Software, Inc.) was used for statistical analysis of the data. Data were first assessed for normality by the Shapiro–Wilk test. If normality could be assumed, Student's t test was used to compare the CtMB and XeMB groups in each condition. If data were deemed non-parametric, then a Mann–Whitney U test was used. For DTI data (FA, MD, RD, AD), a Mann–Whitney U test was used to compare the CtMB and XeMB groups at each time point and region. Vascular reaction and perivascular inflammation scores were also compared between the CtMB and XeMB groups using a Mann–Whitney U test. For comparison of groups in an immunocytochemistry experiment for endothelial cells, one-way analysis of variance (ANOVA) was performed followed by the Tukey's test as a post-hoc test using MATLAB software.
Results
This study followed the schematic shown in Figure 1 (D). After randomization into either the control group or the xenon group, pigs underwent CCI to simulate TBI.
12
They were then subjected to injection of MBs encapsulating xenon (XeMB) or control MBs encapsulating biologically inert PFB (CtMB). MBs were formulated as described previously with a phospholipid shell using sonication and differential centrifugation, followed by microscopy characterization to yield population diameters in the sub-10

Xenon Delivery to Cerebral Vasculature via Microbubbles. View is magnified from the level of the animal
Analysis of DTI parameters at these two time points showed that at day 1, there is significantly higher FA for the XeMB group than for the CtMB group as shown in Figure S2. These changes were noted in bilateral corona radiata, bilateral cerebral peduncles, genu, and splenium of the corpus callosum at day 1. However, at day 5, most of this difference was eliminated and only a small region in the right cerebral peduncle (ipsilateral to the CCI) showed significantly higher FA in the XeMB than in the CtMB treated group as shown in Figure 2. Quantification of the day 1 FA at bilateral corona radiata, genu, and splenium, as well as the right cerebral peduncle showed significantly elevated values in the XeMB group (p < 0.05) as shown in Figure 3. MD showed generally lower levels for the XeMB group than for the CtMB group, but it was only significantly lower at the contralateral corona radiata at day 1 (p < 0.05). At day 5, there were no significant differences between the two groups for FA and MD. Although FA elevation may indicate the level of white matter integrity, other processes such as vasogenic edema and demyelination can alter FA. Therefore, we analyzed other parameters of DTI such as RD and AD. The CtMB group had significantly higher day 1 RD values than the XeMB group at bilateral corona radiata, genu and splenium of the corpus callosum, and the contralateral cerebral peduncle. There were no differences in RD at day 5, and AD at either day 1 or day 5 showed no differences (data not shown).
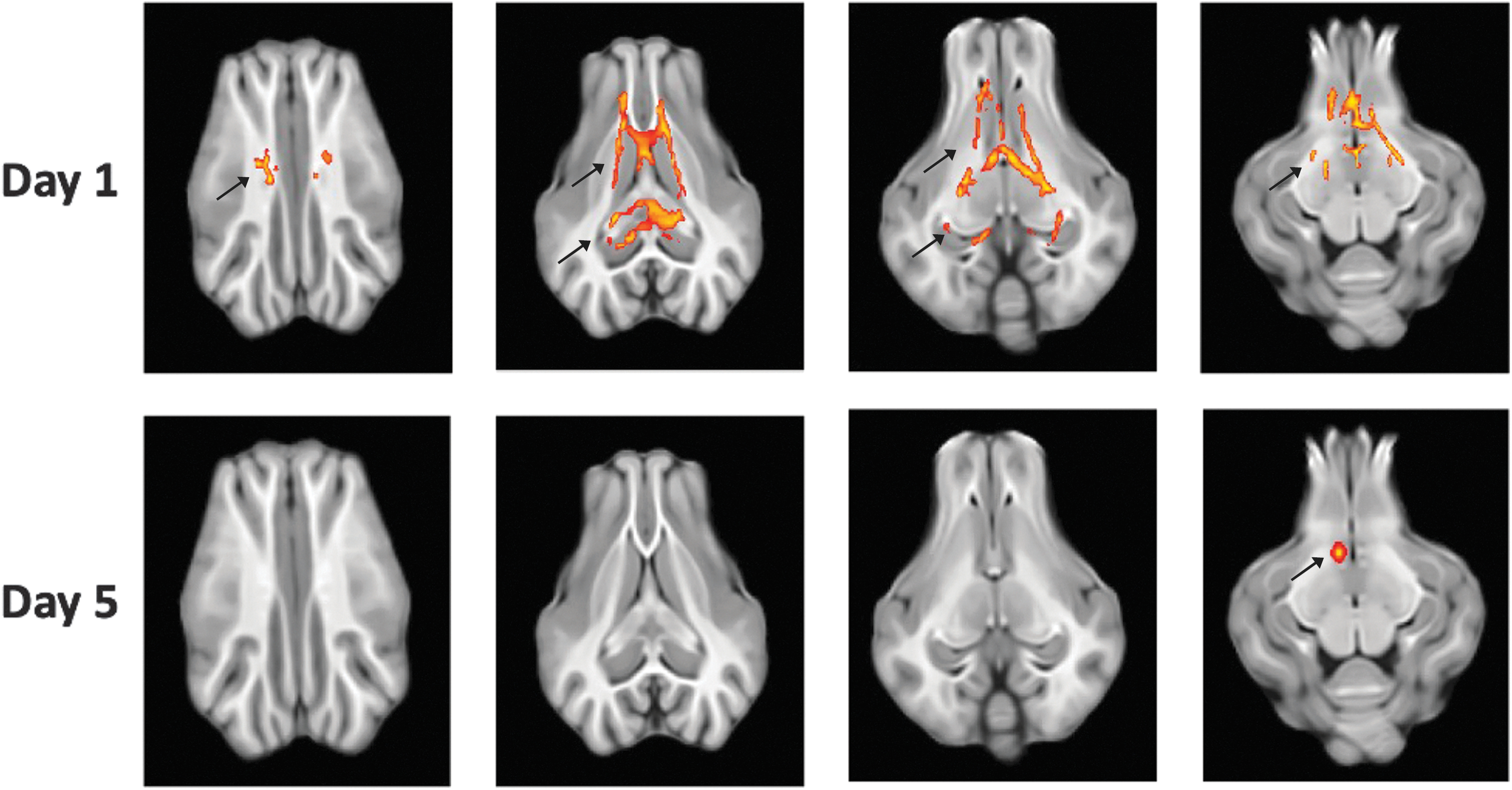
Composite DTI map showing areas of significant FA elevation in XeMB treated pigs compared to CtMB treated pigs. From left the right, superior to inferior axial cuts are displayed. Red-yellow regions indicate areas that have higher FA in XeMB group compared to CtMB group with (P < 0.05).

DTI parameters at 1 day and 5 days after TBI. Ipsi, ipsilateral side to CCI, Contra, contralateral side to CCI; CR, corona radiata; CC, corpus callosum; CP, cerebral peduncle. *P < 0.05 compared to CtMB group.
As further explored in the Discussion section, these findings of FA and RD are consistent with the XeMB group having lower vasogenic edema than the CtMB group at day 1, with subsequent attenuation of this effect at day 5. We explored the tissue effects of xenon post-TBI by histological analysis of the brain parenchyma near the site of injury. At day 5, we euthanized the animals and analyzed the reactive changes at the brain tissue level as shown in Figure S2. At the ipsilateral side to the injury, the XeMB group showed significantly lower perivascular inflammation and vascular reactive changes than the CtMB group (p < 0.05). The contralateral side also showed a close trend of having lower perivascular inflammation for the XeMB group than for the CtMB group (p = 0.052). Part of these data on perivascular inflammation and vascular reactive changes with a smaller group of animals (3 for XeMB and 3 for CtMB) was reported in our prior work. 14
We then studied the reactivity of glial cells across multiple brain regions. The histological sections were stained for GFAP and Iba1 to analyze the degree of astrocytic and microglial reactivity as shown in Figure 4. This showed that the XeMB group had significantly lower GFAP staining than the CtMB group in the contralateral cortex and ipsilateral white matter. Also, the ipsilateral cortex and contralateral white matter showed notable trends for the XeMB group having lower staining with p values 0.08 and 0.06, respectively. However, there were no significant differences between the two groups when Iba1 staining was performed.

GFAP
To understand the degree of blood–brain barrier (BBB) disruption, we compared the degree of fibrinogen extravasation between the CtMB and XeMB groups as shown in Figure 5. On the injury side, there was a statistically significant reduction in staining at the level of the white matter in the xenon-treated group compared with the control group (p = 0.01). This trend was also seen in the pericontusional cortex, but the results were not statistically significant (p = 0.17). On the contralateral side, staining was minimal in both groups and there was no statistically significant difference.

Fibrinogen staining (green) within with blue nuclear stain in the white matter ipsilateral to the injury site. The CtMB animal demonstrated multifocal areas of extravascular fibrinogen staining
We then explored the mechanism of XeMB's protection of the cerebrovasculature in the setting of TBI. Given that TBI results in acute surge of glutamate (Glu) from excitotoxicity, we tested the effect of XeMB on tight junction protein ZO-1 in the setting of Glu insult. The expression levels of ZO-1 tight junction protein were evaluated in endothelial cell culture (BEND3 cells) by confocal microscopy as shown in Figure 6. The control group (without any treatment) showed a baseline level of ZO-1 expression at 53.68 ± 4.91 a.u. When treated with 80 mM Glu, the level of ZO-1 was significantly lower at 21.42 ± 2.53 a.u. (p < 0.001).

Confocal microscopy
When the endothelial cells were treated with the Glu and PFB MBs, the ZO-1 level was similar to that of cells treated with Glu. The mean green fluorescence in the Glu + PFB (22.37 ± 2.27 a.u.) was comparable with that in Glu (21.42 ± 2.53 a.u.), p = 0.997. However, when treated with XeMB (Glu + Xe), the expression level of ZO-1 was similar to the expression level of the control group. Additionally, there was a higher level of ZO-1 expression in Glu + Xe (54.12 ± 3.01) than in Glu (21.42 ± 2.53) treated endothelial cells (p < 0.001).
Discussion
Here, we present the data from a drug delivery strategy for xenon gas using a clinically relevant large animal model of TBI. Compared with CtMB, which encapsulated biologically inert PFB, treatment with XeMB caused acute increase in FA, an imaging biomarker for white matter integrity. Further analysis by multiple diffusivity parameters showed that there was specific reduction in RD consistent with XeMB reducing vasogenic edema. Histologically, XeMB's protective effect on the cerebrovasculature was clarified, as the reactive changes of the vascular endothelial cells and inflammatory cells near the vasculature are significantly decreased in animals treated by XeMB. Moreover, astrocytic reactivity in the XeMB-treated animals was reduced, showing that there is overall reduction of reactive changes from injury. In-vitro endothelial cell imaging showed that XeMB can attenuate the reduction in tight junction protein ZO-1 caused by an acute elevation in glutamate, further supporting the mechanistic basis of this protection. These data indicate that xenon's primary therapeutic action in TBI may be to maintain the integrity of cerebrovasculature rather than its direct action on neurons.
As a monoatomic chemical with low reactivity, xenon has no toxicity in cardiac, renal, hepatic, or hematological function. 15 As previously reviewed, 16 it can also be used as an anesthetic agent, but its high cost makes this agent prohibitive: at a current cost of $10 per liter, a 2 h procedure using 70% xenon gas would lead to 34 L of xenon gas being wasted into the atmosphere. 17 Given this background, this report demonstrates how localized xenon delivery using MBs could be utilized in future brain injury studies to make this therapeutic approach more efficient. MB-induced drug delivery has been explored in various settings in the last 15 years. MB-encapsulating drugs can deliver them locally by ultrasound-induced oscillations, leading to a phenomenon called sonoporation. 18 For expensive agents such as xenon, this local delivery technique can reduce the amount of xenon needed to be delivered to target tissue. For other agents that have concerns for peripheral side effects, MBs can also limit exposures to non-target tissues. Design of lipid MBs that encapsulates xenon and releases them on ultrasonic stimulation has been previously demonstrated by our group. 8,14 The current report is about the utilization of this method in an in-vivo setting. Moreover, we utilized a pig model of TBI given its gyrencephalic anatomy, size, and gray–white matter ratio that much more closely replicates the human brain than do rodent brains. 19 The physiological relevance of the pig model can be evidenced also by the number of neurons: the human brain possesses ∼86 billion neurons, whereas the porcine brain possesses 2.2 billion neurons. 20 In comparison, the mouse brain contains 67,000,000 neurons.
The therapeutic effect of xenon in stroke, perinatal asphyxia, and TBI has been associated with its NMDA receptor antagonist effect. 4,21 –23 Specifically, in TBI animal models, xenon treatment can lead to reduction of injury volume, reduced astrogliosis and microglia, and attenuation of hippocampal neuronal loss. 5 However, the therapeutic effect of xenon may reach beyond preventing excitotoxicity. Prior research on inhalational xenon's effect on blood flow showed that it can increase perfusion in various regions of the brain, such as the cerebral cortex, brainstem, and cerebellum. 24 This treatment also showed potential therapeutic benefit in the setting of anoxic injury in pigs that underwent cardiac arrest. 25 Glycerol, a marker of damage collected via microdialysis, showed reduction during the reperfusion period when xenon was administered. Therefore, additional therapeutic effects may have been via optimization of cerebral perfusion following TBI, given that dysregulation of perfusion has been well demonstrated in the past. 26,27
The findings from the current article show the protective effect of xenon in measures associated with cerebrovasculature. Diffusion imaging parameters are consistent with XeMB-treated animals having reduced vasogenic edema, and histological findings showed a reduction of reactive changes within endothelial cells, decreased leakage of inflammatory cells into perivascular spaces within the brain, and reduced permeability of cerebral vasculature. Additionally, our cell culture results support BBB protection by XeMB as the potential mechanism of reducing perivascular inflammation after TBI. Xenon treatment reversed the BBB permeability effects of glutamate and demonstrated restoration of tight junction protein expression in endothelial cells. In support of this, NMDA receptor antagonists have previously been shown to reduce BBB permeability after glutamate treatment, 28 although it has not been shown that xenon similarly protects BBB integrity.
One of the other potential mechanisms of xenon's therapeutic effect is by activating adenosine triphosphate (ATP) sensitive K+ channels. Xenon has been shown to activate ATP-sensitive K+ channels, 29 and ATP-sensitive K+ channels are important contributors to neuroprotection in ischemic insult. 30 Specifically, Kir6.2 subunit activation by xenon can induce a neuroprotective effect, because Kir6.2 knockout mice have increased susceptibility to acute hypoxia. 31 As evidenced by our finding in Figure 4, xenon may act to reduce astroglial reactive changes. Reduction of astroglial proliferation by xenon was shown in a Parkinson's disease model, by potentially delaying the cell cycles of astrocytes. 32,33 Aside from drug delivery, MBs have been demonstrated to be effective agents in gene vector delivery in vivo. 34 Moreover, this delivery can be targeted for specific molecular markers by embedding antibodies for targets such as E-selectin, P-Selectin, vascular cell adhesion molecule (VCAM)-1, and vascular endothelial growth factor (VEGF)-R2. 35 Therefore, in future studies, specific intracerebral targets may be chosen for MB delivery of a therapeutic agent such as xenon.
In this study, we also describe an efficient delivery of xenon, which can be cost prohibitive. Other methods of delivery in animal studies use a much larger volume of gas: conventional chambers used for gas delivery are ∼40–50 L in size, 36 with additional delivery of xenon 200 mL/min over 1–3 h, which can range from ∼10 to 40 L of additional xenon use. 37,38 Also in our prior review, we compared the use of xenon via MBs to inhalational methods. 16 Most studies here explored xenon as an anesthetic agent at 70% concentration. For a 2-h anesthetic procedure that uses 70% xenon at 0.5 L/min, there is efficiency of <20%; therefore 34 L of xenon gas is wasted. 17 Xenon at 70% is likely not necessary for TBI treatment, because 35–50% xenon is still neuroprotective in hypoxia experiments, 38 but the point of low efficiency is still valid even at lower concentrations. The use of a closed-circuit system can significantly reduce xenon gas usage: a 2-h anesthetic procedure using 70% xenon with this system may use 5.8–11.5 L of xenon. However, this is still much higher than the volume of xenon we utilized (three treatments of 1.5 mL = 4.5 mL).
Limitations
This is an initial study showing the therapeutic potential for localized xenon delivery. However, this study is limited by a lack of dose comparison. A side-by-side comparison at multiple doses to see which dose has the largest therapeutic effect should be performed in the future. Additional characterization of the imaging and histological changes after XeMB or inhaled xenon exposure will also be helpful to justify the importance of MB delivery. Although MB encapsulation definitely reduces the volume of xenon gas that is utilized, XeMB use can only be argued for if the therapeutic effect of both means of xenon delivery is equivalent. Moreover, long-term effects of xenon treatment up to weeks or months after injury needs to be assessed in future studies, as the current report only focused on changes at 5 days. Moreover, functional assessment of pigs after xenon treatment is necessary to further support the need for a clinical trial using xenon in human TBI patients.
Conclusion
Data from the current study support the use of targeted xenon delivery to the brain to protect cerebrovasculature following TBI. Given the favorable safety profile of xenon gas and potential toxicities reported with NMDA antagonist drugs, there is exciting future potential for clinical trials using targeted xenon delivery to the brain for neuroprotection and BBB protection.
Footnotes
Transparency,Rigor,and Reproducibility Summary
The study and analysis plan were registered at the Center for Open Science (
Acknowledgments
We thank Sara Morton for the surgical treatment of the animals.
Authors' Contributions
S.S.S. was responsible for conceptualization, writing–original draft, review and editing, and formal analysis R.C. was responsible for review and editing, data curation, and formal analysis. A.N.V. was responsible for data curation. M.B.K. was responsible for data curation and formal analysis. S.H. was responsible for data curation. R.D. was responsible for data curation. A.S. was responsible for data curation. C.S. was responsible for conceptualization and visualization. D.L. was responsible for conceptualization and supervision. T.J.K. was responsible for conceptualization and supervision. M.H. was responsible for supervision, review and editing, and funding acquisition.
Funding Information
Funding for this report was provided by the Junior Faculty Pilot Grant (7225290000-05), Children's Hospital of Philadelphia, and the 2019-20 Health Research Formula Fund Grant Award 581526 from the Commonwealth of Pennsylvania.
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.