
Abstract
Neonatal spinal cord injury (SCI) shows better functional outcomes than adult SCI. Although the regenerative capability in the neonatal spinal cord may have cues in the treatment of adult SCI, the mechanism underlying neonatal spinal cord regeneration after SCI is unclear. We previously reported age-dependent variation in the pathogenesis of inflammation after SCI. Therefore, we explored differences in the pathogenesis of inflammation after SCI between neonatal and adult mice and their effect on axon regeneration and functional outcome. We established two-day-old spinal cord crush mice as a model of neonatal SCI. Immunohistochemistry of the spinal cord revealed that the nuclear translocation of NF-κB, which promotes the expression of chemokines, was significantly lower in the astrocytes of neonates than in those of adults. Flow cytometry revealed that neonatal astrocytes secrete low levels of chemokines to recruit circulating neutrophils (e.g., Cxcl1 and Cxcl2) after SCI in comparison with adults. We also found that the expression of a chemokine receptor (CXCR2) and an adhesion molecule (β2 integrin) quantified by flow cytometry was lower in neonatal circulating neutrophils than in adult neutrophils. Strikingly, these neonate-specific cellular properties seemed to be associated with no neutrophil infiltration into the injured spinal cord, followed by significantly lower expression of inflammatory cytokines (Il-1β, Il-6 and TNF-α) after SCI in the spinal cords of neonates than in those of adults. At the same time, significantly fewer apoptotic neurons and greater axonal regeneration were observed in neonates in comparison with adults, which led to a marked recovery of locomotor function. This neonate-specific mechanism of inflammation regulation may have potential therapeutic applications in controlling inflammation after adult SCI.
Introduction
The mature mammalian spinal cord does not regenerate once damaged. 1 Therefore, spinal cord injury (SCI) causes permanent motor, sensory, and autonomic nerve damage, which impose physical, psychological, and economic burdens on patients. 2 A SCI can occur even in neonates at birth because of head malposition during delivery or the use of suction or forceps for delivery assistance, potentially resulting in quadriplegia. 3
Interestingly, after SCI, neonates often achieve remarkable functional recovery. Through this recovery, even neonates with complete paralysis at the time of injury (i.e., American Spinal Cord Injury Association Impairment Scale A) may regain the ability to walk. 4 This fact suggests that neonates may have greater spinal cord regenerative capacity than adults.
Injury to the spinal cord causes neuroinflammation leading to neuronal apoptosis or demyelination, which worsens the functional prognosis. 5,6 For this reason, suppressing inflammation after SCI is important for improving functional outcomes. The inflammatory response after SCI, however, involves complex cellular networks, including several types of blood-derived cells (e.g., neutrophils, monocytes/macrophages, lymphocytes, and mast cells), as well as spinal cord-intrinsic astrocytes and microglia. 6 –9
Considering the inflammatory response at the injured spinal cord caused by blood-derived cells, neutrophils are the first to infiltrate into the injured spinal cord as the main initiator of a complex inflammatory cascade, 10,11 suggesting that the suppression of neutrophil infiltration would be quite effective for inflammation control.
In addition to infiltrating blood-derived inflammatory cells, astrocytes also contribute to enhancing the inflammatory response at the injured spinal cord, secreting the neutrophil attractant chemokine CXCL1. 12 Therefore, elucidation of the cellular properties of both neutrophils and astrocytes in the injured spinal cord is important for controlling acute phase inflammation after SCI. 13,14
Neutrophils and astrocytes in the injured spinal cord also affect axon regeneration. When inflammatory cytokines secreted from infiltrated neutrophils act on astrocytes, naïve astrocytes are activated and phenotypically changed to become reactive astrocytes. Reactive astrocytes then form glial scars that physically and chemically inhibit axon regeneration. 14 –16 Once formed, the glial scar remains until the chronic phase of SCI; 17 thus, the suppression of astrocyte activation is important for preventing glial scar formation.
To address this problem of astrocyte interference in axon regeneration, the neutrophil-astrocyte interaction in the injured spinal cord should be elucidated. How neutrophils interact with astrocytes in neonates after SCI, however, is unclear, whereas neonates regain better locomotor function after SCI than adults.
Here, we established a spinal cord crush injury model using post-natal day 2 (P2) mice and adult mice to investigate the spinal cord regenerative capacity and pathophysiology in the acute phase of SCI, focusing on neuroinflammation from astrocytes and neutrophils, axon regeneration, and gait functions.
Cell sorting from the injured spinal cord and gene expression analysis revealed a different pattern of inflammatory cell infiltration between adults and neonates. The neonate-specific inflammatory cell infiltration patterns were related to chemokine-chemokine receptor interaction properties between astrocytes and neutrophils.
Further, we found a difference in the neutrophil adhesion molecule expression states that was also associated with neonate-specific inflammatory responses. Neonatal spinal cord regeneration seemed to be associated with neonate-specific inflammation regulatory mechanisms.
Methods
Mice
Wild-type C57/BL6N mice (Japan SLC, Japan) and Aldh1l1-EGFP transgenic mice were used in the study. Post-natal day 2 pups (P2) and eight-week-old mice were used as neonatal and adult mice, respectively. A total of 156 mice were assigned randomly to the sham group or the SCI group with matching for age. Randomization was performed using computer-generated random numbers. All mice were housed with humidity controlled to 30–70%, temperature controlled to 21–23°C, and a 12h light-dark cycle.
All experimental procedures were conducted in compliance with animal protocols approved by the Committee of Ethics on Animal Experimentation in the Faculty on Medicine, Kyushu University (A22-238-0). All measurements were performed with the experimenter blinded to group identity.
Spinal cord crush injury
Mice were anesthetized with isoflurane inhalation (4% induction, 3% maintenance). A 5 mm skin incision was made along the sagittal midline of the spine, and laminectomy of the T9 laminae was performed to expose and crush the spinal cord at the T10 spinal segment. The spinal cord was fully compressed for 2 sec with forceps (width: 100 μm). The tips of the forceps were continuously attached on the back wall of the vertebrae during the crush procedure. After crush injury, the skin was closed with 6-0 nylon thread.
Meloxicam (0.2 mg/kg) was administered subcutaneously for analgesia and mice were placed into a temperature-controlled chamber during recovery from anesthesia to regain thermoregulation. Sham-operated controls were also anesthetized with isoflurane and subjected to laminectomy only. We confirmed that the dura mater was not affected by crush injury in either adults or neonates (Supplementary Fig. S1).
Analysis of the locomotor function
Open field locomotor function was evaluated via the Basso Mouse Scale scoring system at 7, 14, 28, and 112 days after injury. 18 A footprint analysis was performed as described previously. 19 Briefly, the forelimbs and hindlimbs were dipped in red and green dyes, respectively. The mice walked on a paper-covered runway 4 cm in width and 80 cm in length. A bright box was placed at the beginning of the runway, and a dark box was also placed with food at the end of the runway.
Flow cytometry
At 12h after crush injury, animals were reanesthetized with a lethal dose of isoflurane and perfused with ice-cold normal saline for circulating blood removal. A spinal cord specimen (6 mm in length) centered on the epicenter of the lesion was rapidly dissected from the vertebral column and placed into type 1 collagenase solution (100 U/mL; Invitrogen).
The spinal cord was minced using scissors and incubated in the solution for 25 min at 37°C for gradual dissociation. Dissociated cells were centrifuged at 4000 rpm for 1 min, suspended in Dulbecco modified Eagle medium containing 10% fetal bovine serum and passed through a 40 μm nylon strainer.
For blood sample collection, cardiac puncture was conducted on mice at 12h after crush injury. After removing red blood cells by BD Pharm Lyse (BD Biosciences, San Jose, CA), the resulting suspension was centrifuged and washed twice in FACS buffer. These suspensions were pelleted by centrifugation and incubated at 4°C for 30 min with the following fluorescent antibodies: phycoerythrin-Cy7-conjugated CD45, fluorescein isothiocyanate-conjugated Gr-1, allophycocyanin-conjugated CD11b, phycoerythrin-conjugated CXCR2, and phycoerythrin-conjugated β2 integrin (BioLegend, San Diego, CA).
Before the analysis, 7-amino-actinomycin D (BD Biosciences) was added to determine the cell viability. All samples were suspended in 500 μL of FACS buffer and analyzed using a FACSfusion flow cytometer (BD Biosciences). The data were analyzed using the FACSDiva software program (BD Biosciences).
Quantitative real-time polymerase chain reaction (PCR)
Total ribonucleic acid (RNA) was isolated from each injured spinal cord specimen (length: 6 mm; centered on the epicenter of the lesion) using an RNeasy Mini Kit (Qiagen, Hilden, Germany). The RNA was reverse transcribed using PrimeScript Reverse Transcriptase (TaKaRa, Shiga, Japan). Quantitative real-time PCR was conducted on 20 μL of reaction mixture with primers specific to the genes of interest (Table 1) and TB Green Premix DimmerEraser (TaKaRa).
Primers Used in the Quantitative Polymerase Chain Reaction Analyses
The mRNA levels in each sample were normalized to those of glyceraldehyde-3-phosphate dehydrogenase. For the mRNA expression analysis of neutrophils and astrocytes, these cells were individually isolated from the spinal cord via FACSfusion, and total RNA isolation was performed using the RNeasy Micro Kit (Qiagen).
Immunohistochemistry
Mice were reanesthetized and transcardially perfused with ice-cold normal saline, followed by fixation with 4% paraformaldehyde. The spinal cord was removed and immersed in the same fixative for 24h. The spinal cord was dehydrated in 10% sucrose solution for 1h and subsequently in 30% sucrose solution for 24h.
The spinal cord was embedded in an optimal cutting temperature compound. Frozen sections were cut on a cryostat in the sagittal plane at 16 μm. The sections were stained with primary antibodies at 4°C overnight, followed by a 1h incubation with fluorescence-conjugated secondary antibodies and Hoechst 33342 (1:1000; Invitrogen).
The applied primary antibodies were as follows: Ly6B.2 (rat, MCA771GA; 1:400; BIO RAD), glial fibrillary acidic protein (GFAP, rat, no.13-0300; 1:400; Invitrogen), nuclear factor kappa B (NF-κB p65, rabbit, sc-372: 1:400; Santa Cruz Biotechnology), NeuN (mouse, MAB377; 1:400; Sigma‒Aldrich), cleaved caspase-3 (rabbit, no.9579T; 1:400; Cell Signaling Technology), and 5-hydroxytryptamine (5-HT, goat, no.20079; 1:400; ImmunoStar).
For the evaluation of apoptotic cells in the injured spinal cord, a terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) assay was performed using an ApopTag red in situ kit (Chemicon, Temecula, CA).
Quantitative analysis for immunohistology
For the quantification of Ly6B.2+ neutrophils, intranuclear p65+/GFAP+ cells, TUNEL+ cells, cleaved caspase-3+/NeuN+ cells, and NeuN+ cells in the spinal cord at 12h after injury, 10-mm-long sagittal sections at 200-μm intervals were viewed on a light microscope (BZ-9000; Keyence, Osaka, Japan), and each cell was counted using the Dynamic Cell Count BZ-H1C software program (Keyence). The numbers of 5-HT + areas per 1 mm2 segment were calculated using ImageJ (National Institutes of Health).
For the comparison of astrocyte process lengths of adult and neonatal pups at 12h after injury, 20 astrocytes were selected randomly per field of view in each section, and the average length of the longest projection of each astrocyte was calculated.
Statistical analysis
Statistical analyses to compare the means of data between two groups were performed using unpaired t tests. For analyses of the repeated measures and multiple group comparisons, a two-way repeated-measures analysis of variance (ANOVA) with the Tukey-Kramer post hoc test was performed. All tests were two-tailed, and p values of <0.05 were considered statistically significant.
Data in the graphs are presented as the mean ± standard error of the mean. All analyses were conducted using the JMP PRO 16.0.0 software program (SAS Institute, Cary, NC). We performed a post hoc power analysis to confirm that each analysis had sufficient statistical power (Table 2).
Results of Post-Hoc Power Analyses
SD, standard deviation.
Results
Expression of inflammatory cytokines and chemokines in the acute phase of SCI is lower in neonates than in adults
We have reported previously that inflammation after SCI is milder in young mice than in adult mice. 20 Therefore, we hypothesized that the pathophysiology of neuroinflammation after SCI changes in an age-dependent manner and investigated whether there is a difference in the inflammatory response after SCI between adults and neonates.
The gene expression analysis of the spinal cord at 12h after injury, when the expression of inflammatory cytokines peaks, 20 revealed that there was a statistically significant interaction between the effects of mouse age and crush injury on the expression levels of interleukin (Il-1β) (F(1, 28) = 38.85, p < 0.0001), Il-6 (F(1, 28) = 12.24, p = 0.0016) and tumor necrosis factor-alpha (TNF-α) (F(1, 28) = 51.00, p < 0.0001).
The simple main effects analysis showed that the expression levels of Il-1β, Il-6 and TNF-α in neonates after SCI were significantly lower than those in adults (p < 0.0001, = 0.0002, and <0.0001, respectively), whereas no significant differences were found in the spinal cord of sham-operated neonates and adults (p > 0.9999, p > 0.9999, and p = 0.9890, respectively) (Fig. 1A).

Low expression of inflammatory cytokines and chemokines in the neonatal spinal cord after spinal cord injury. (
The expression of these inflammatory cytokines is associated with the infiltration of inflammatory cells, such as neutrophils and monocytic cells, which migrate toward chemotactic factors. 21 Therefore, we examined the expression level of several chemokines in the spinal cord at 12h after injury and found a statistically significant interaction between the effects of mouse age and crush injury on the expression levels of Cxcl1 (F(1, 28) = 16.54, p = 0.0004), Cxcl2 (F(1, 28) = 11.08, p = 0.0024), Cxcl5 (F(1, 28) = 37.85, p < 0.0001), Ccl2 (F(1, 28) = 19.80, p = 0.0001), and Ccl3 (F(1, 28) = 41.32, p < 0.0001), which promoted the migration of neutrophils and macrophages.
The simple main effects analysis showed that the expression levels of Cxcl1, Cxcl2, Cxcl5, Ccl2, and Ccl3 in neonates after SCI were significantly lower than those in adults (p < 0.0001, p = 0.0003, p < 0.0001, p < 0.0001, and p < 0.0001, respectively), whereas no significant differences were found in the spinal cord of sham-operated neonates and adults (p > 0.9999, p > 0.9999, p > 0.9999, p > 0.9999, and p = 0.9975, respectively) (Fig. 1B).
Few neutrophils infiltrate the injured spinal cord in neonatal mice
Neonatal spinal cord specimens showed lower chemokine levels than adult spinal cord specimens at 12h after SCI. This difference may have affected the number of infiltrating neutrophils or macrophages that migrate under the influence of chemokines. Therefore, the numbers of inflammatory cells infiltrating into the spinal cord at 12h after injury were investigated by flow cytometry.
In adults, both monocytic cells (CD45posi/CD11bposi/Gr-1nega-int) and neutrophils (CD45posi/CD11bposi/Gr-1high) infiltrated the spinal cord at 12h after injury. Surprisingly, the neonatal spinal cord had no neutrophil infiltration at 12h after injury in contrast to the adult spinal cord (t = 3.495, df = 8, p = 0.0081, two-tailed), whereas neutrophils were present in the peripheral blood (Fig. 2A,B). Further, immunostaining showed that Ly6B.2+ neutrophils infiltrated into the adult spinal cord at 12h after injury, but not into the neonatal spinal cord (t = 7.841, df = 8, p < 0.0001, two-tailed), similar to the flow cytometry results (Fig. 2C, D).

Neutrophils scarcely infiltrate the neonatal spinal cord after spinal cord injury. (
Astrocyte-derived neutrophil chemokine expression in neonates is lower than that in adults after SCI
We showed that the expression levels of several chemokines in the injured spinal cord of neonates were lower than those in adults and that neutrophils did not eventually infiltrate the spinal cord. It is still unclear why the expression of neutrophil migration factor is low in neonates. Because astrocytes secrete chemokines in mammalian central nervous system (CNS) injury and promote the infiltration of neutrophils beyond the blood‒brain barrier into the CNS, 12,22 –24 we decided to compare astrocyte-derived chemokine expression between adults and neonates.
Using Aldh1l1-EGFP transgenic mice, a reporter mouse for astrocytes (Fig. 3A), EGFP⁺ astrocytes were collected by flow cytometry (Fig. 3B). The gene expression analysis of enhanced green fluorescent protein (EGFP⁺) astrocytes at 12h after SCI revealed that there was a statistically significant interaction between the effects of mouse age and crush injury on the expression levels of Cxcl1 (F(1, 20) = 8.817, p = 0.0096) and Cxcl2 (F(1, 20) = 13.23, p = 0.0016), both of which are neutrophil migration factors. 12,21,25,26
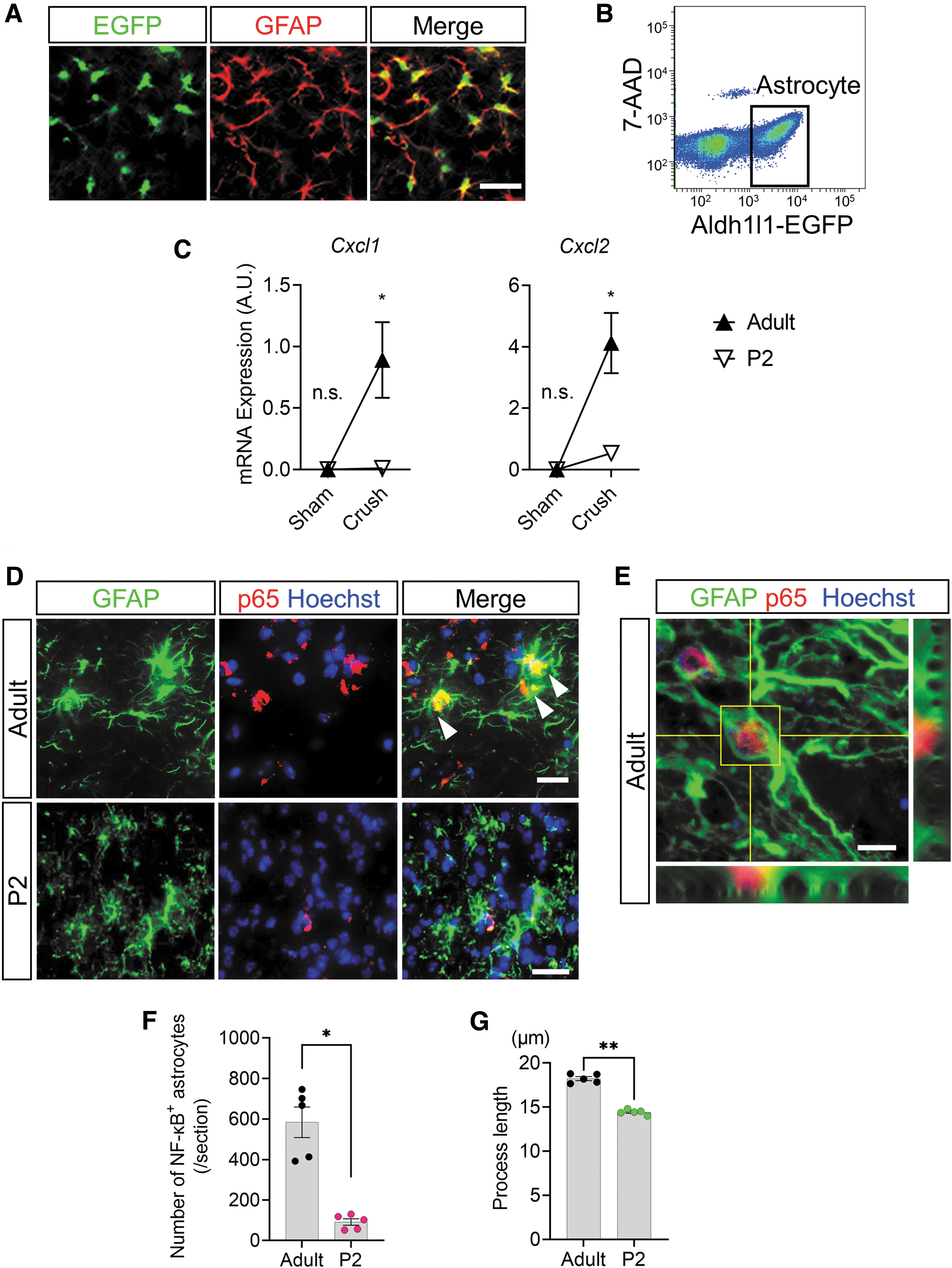
The expression of astrocytic chemokines via nuclear factor kappa B (NF-κB) activation rarely occurs in neonatal mice after spinal cord injury. (
The simple main effects analysis showed that the expression levels of Cxcl1 and Cxcl2 in neonatal astrocytes after SCI were significantly lower than those in adults (p = 0.0033 and 0.0003, respectively), whereas no significant differences were found in the astrocytes of sham-operated neonates and adults (p > 0.9999, respectively) (Fig. 3C).
The expression of Cxcl1 and Cxcl2 from spinal cord astrocytes is regulated by p65, a transcription factor of the NF-κB family. 23,27 The comparison of p65 expression in astrocytes by immunostaining revealed a significant decrease in the intranuclear translocation of p65 in astrocytes of neonates compared with that of adults (t = 6.399, df = 8, p = 0.0002, two-tailed) (Fig. 3D,F). Interestingly, adult astrocytes colocalized with p65 showed reactive astrocyte-like morphology with significantly elongated processes in comparison with astrocytes from neonates (t = 14.59, df = 8, p < 0.0001, two-tailed) (Fig. 3G).
Circulating neutrophils express fewer chemokine receptors and adhesion molecules in neonates than in adults
Because neutrophil migration and infiltration are tightly controlled by interactions between chemokines and chemokine receptors, as well as adhesion molecules and the endothelium, 28,29 the lack of neutrophil infiltration in the injured spinal cord in neonates may also be related to these neutrophil surface antigen expression levels.
We used flow cytometry to quantify CXCR2 (a receptor for CXCL1 and CXCL2) 28 and β2 integrin (an adhesion molecule required for neutrophil infiltration outside of blood vessels) 30,31 in neonatal circulating neutrophils. As a result, the level of CXCR2 in circulating neutrophils of neonates was significantly lower than that of adults (t = 6.452, df = 8, p = 0.0002, two-tailed) (Fig. 4A, 4B). Further, β2 integrin expression in neonatal circulating neutrophils was significantly lower than that in adult circulating neutrophils (t = 13.67, df = 8, p < 0.0001, two-tailed). (Fig. 4C, 4D).
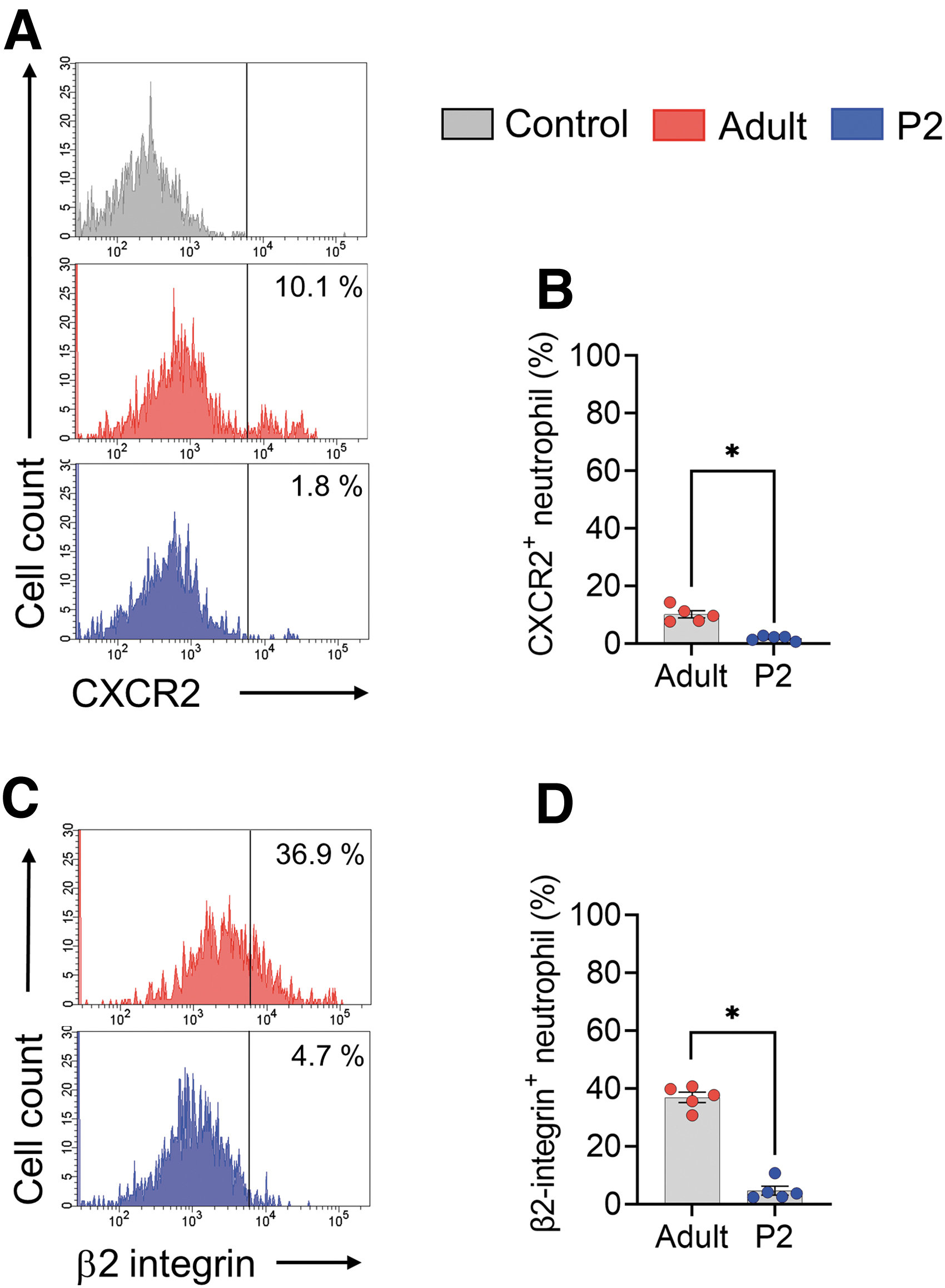
Circulating neutrophils in neonatal mice express lower levels of chemokine receptors and adhesion molecules. (
Secondary injury after SCI is attenuated in neonatal mice
The inflammatory responses after SCI cause neuronal apoptosis and demyelination (called secondary injury) and significantly affect the functional outcome. 5,6,8 Because the acute inflammatory response differs between neonatal and adult mice, we decided to compare the magnitude of secondary injury between the two groups by performing TUNEL staining, which reflects apoptosis throughout the spinal cord (Fig. 5A). As a result, there was a statistically significant interaction between the effects of mouse age and crush injury on the number of TUNEL-positive apoptotic cells (F(1, 16) = 35.05, p < 0.0001).
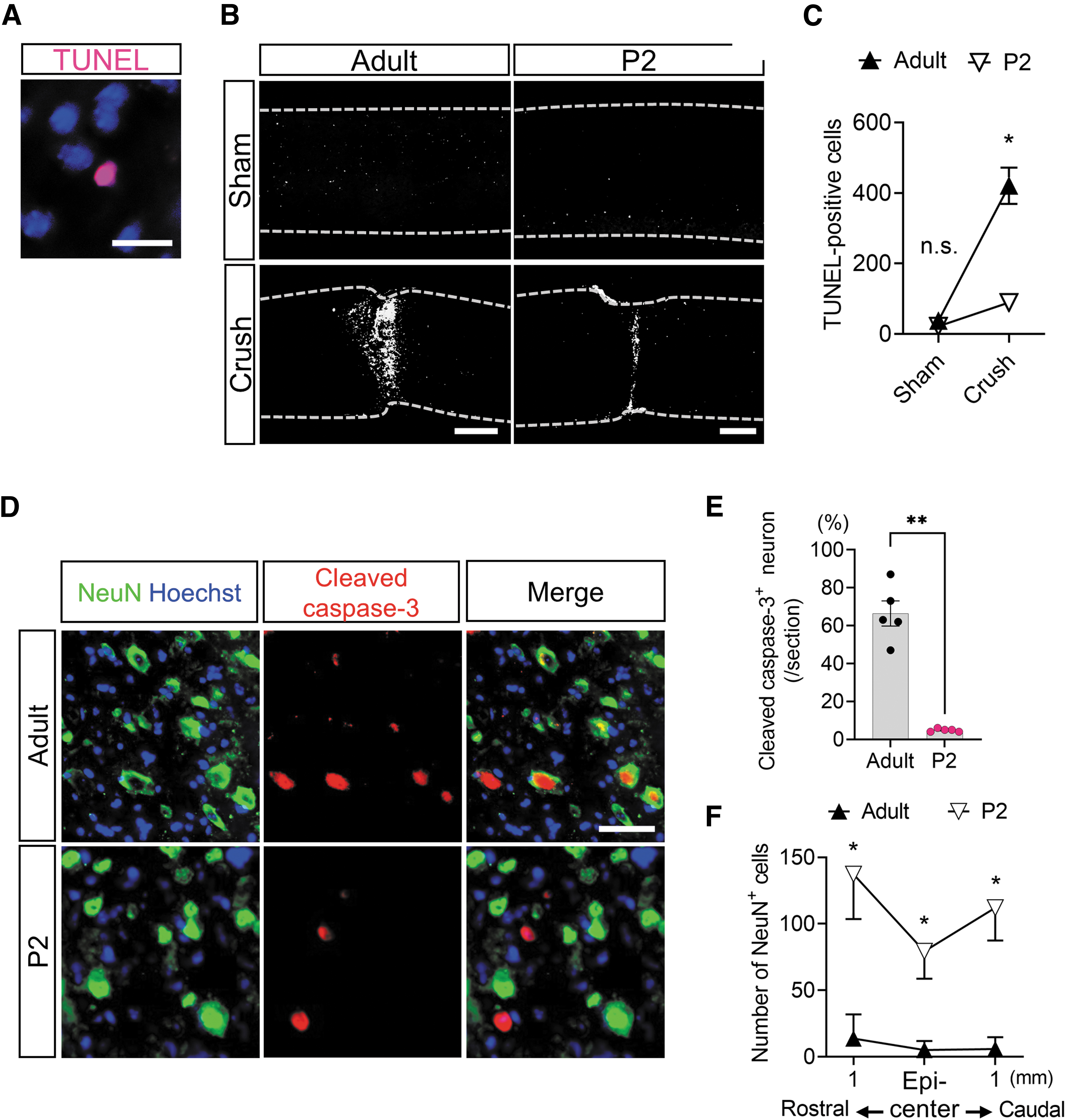
Neuronal apoptosis following the inflammatory response after spinal cord injury is attenuated in neonatal mice. (
The simple main effects analysis showed that the number of TUNEL-positive apoptotic cells in the neonatal spinal cord after SCI was significantly lower than that in adults (p < 0.0001), whereas no significant differences were found in the spinal cord of sham-operated neonates and adults (p = 0.9763) (Fig. 5B, 5C).
We subsequently investigated whether apoptosis was induced by the extrinsic pathway because TNF-α causes caspase-3-mediated apoptosis of neurons and oligodendrocytes. 6 Immunostaining revealed a greater number of intranuclear translocated cleaved caspase-3 proteins colocalized with neurons in adults than in neonates (t = 9.297, df = 8, p < 0.0001, two-tailed) (Fig. 5D, 5E), suggesting that neuronal apoptosis induced by the extrinsic pathway is less likely to occur in neonates after SCI.
As a result of attenuated neuronal apoptosis, the number of neurons on day 3 after injury was significantly higher in the neonates than in the adults (F(2, 30) = 4.286, p = 0.0230) at the epicenter of the lesion (p < 0.0001) in addition to the rostral side (p < 0.0001) and caudal side (p < 0.0001) of the lesion (Fig. 5F).
Axon regeneration and good recovery of motor function in neonatal mice after SCI
Because IL-1β in the injured spinal cord has been reported to exacerbate axon regeneration, 32 we decided to examine the effects of the different inflammatory cytokine expression levels between adults and neonates on axon regeneration. Immunostaining revealed that 5-HT+ axons disappeared caudal to the injured area on day 3 after SCI in both adults and neonates (Fig. 6A).
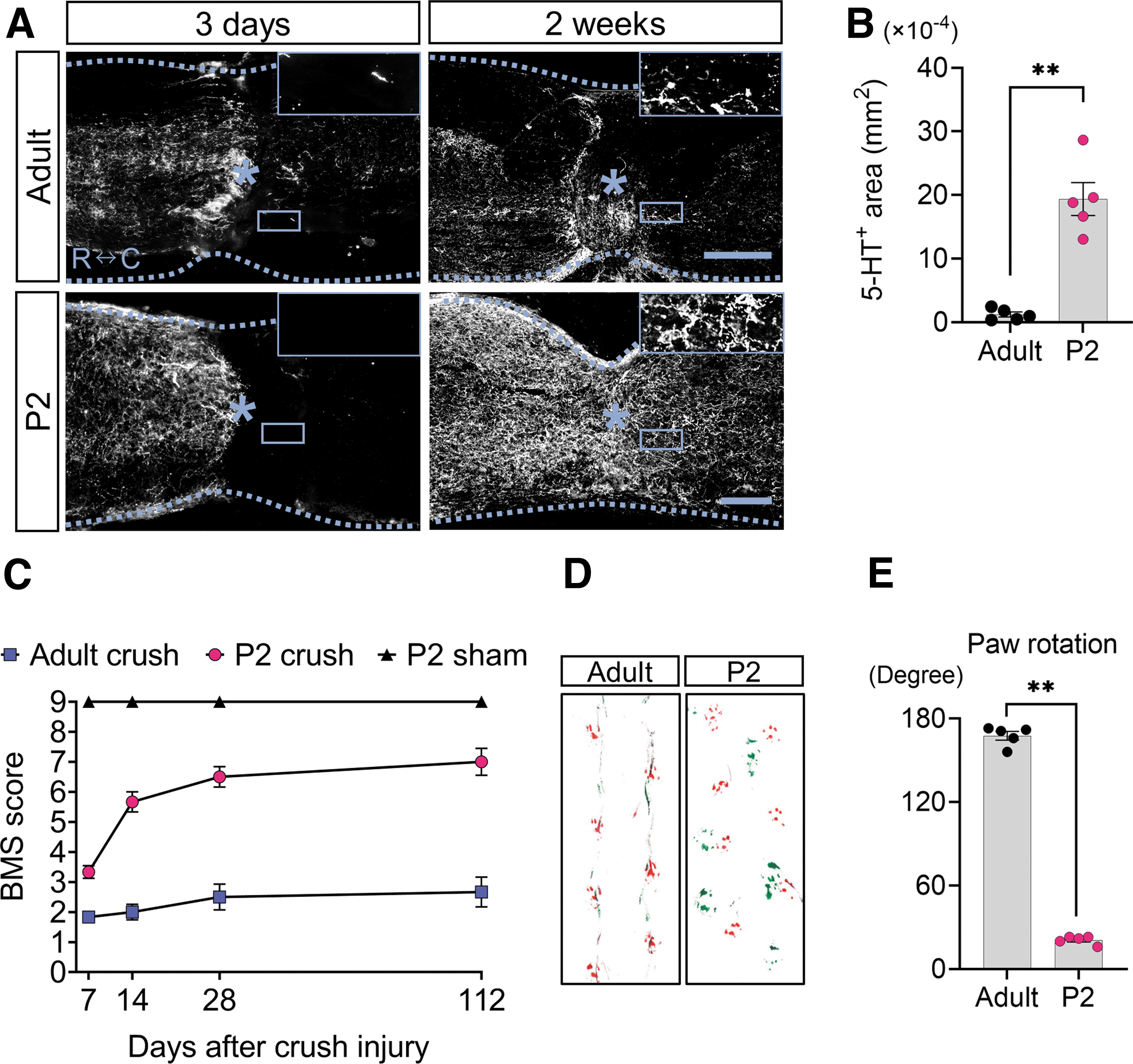
Serotonergic axon regeneration accompanied by outstanding functional recovery is observed in neonatal mice. (
The 5-HT+ axons, however, were vigorously extended on the caudal side of the injured area in neonates at two weeks after injury, whereas little recovery was observed in adults (Fig. 6A). Consequently, the number of 5-HT+ axons on the caudal side of the injured area was significantly greater in neonates than in adults (t = 6.929, df = 8, p < 0.0001, two-tailed) (Fig. 6B). Locomotor function, as assessed by the Basso Mouse Scale, which is related to the magnitude of regenerated 5-HT+ axons, 33,34 was significantly better in neonates than in adults (F(6, 60) = 9.397, p < 0.0001: Tukey‒Kramer post hoc test, p < 0.0001) (Fig. 6C).
The footprint analysis showed that adult mice walked with their green pigmented hindlimbs dragging, while neonate hindlimbs and forelimbs were placed in coordination (Fig. 6D). The footprint analysis also revealed that the hindlimb rotation of neonates was improved, while adult mice suffered residual rotation (t = 43.10, df = 8, p < 0.0001, two-tailed) (Fig. 6E).
Discussion
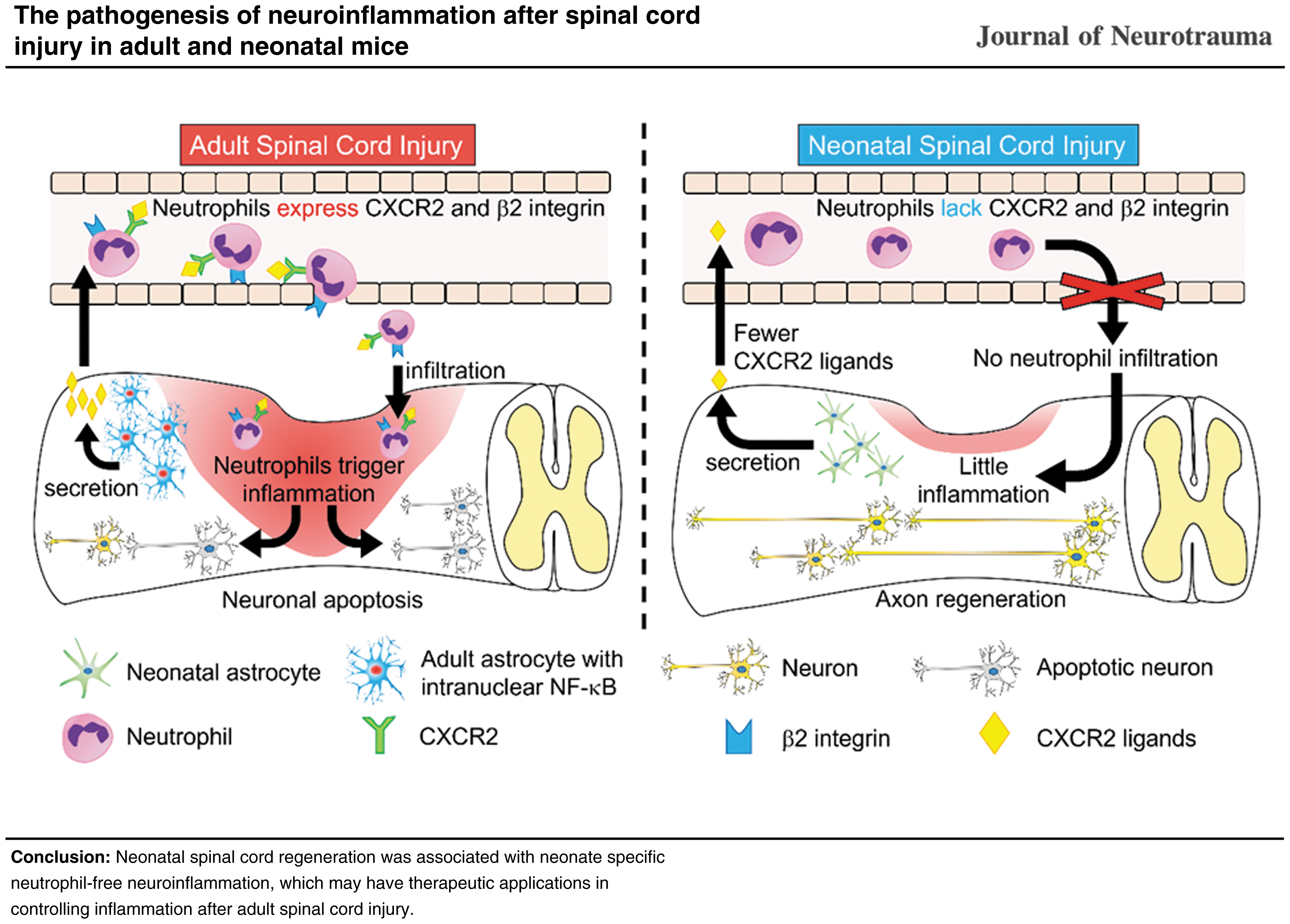
In this study, we discovered that the expression levels of intranuclear NF-κB and associated chemokines in the astrocytes of neonatal mice were significantly lower than those in adult mice after SCI. Moreover, the expression levels of chemokine receptors and adhesion molecules in the neutrophils in neonatal mice were markedly lower than those in adult mice. These neonate-specific cellular properties were accompanied by no neutrophil infiltration into the spinal cord and little inflammation, mild secondary injury, and remarkable spinal cord repair with good recovery of locomotor functions after SCI.
In regard to the functional properties of circulating neutrophils in neonates, the expression levels of both CXCR2 and β2 integrin at 12h after injury were significantly lower than those in adults (Fig. 4A-4D). In a study using neutrophil-specific CXCR2-depleted adult mice with experimental autoimmune encephalomyelitis, the infiltration of neutrophils into the spinal cord was observed. 35 From this result, the neutrophil-free inflammation seen in this study could not be solely caused by the reduced expression of CXCR2.
β2 Integrin combined with CD11a and CD11b forms the adhesion molecules LFA-1 and MAC-1, respectively. 36 –38 LFA-1 and MAC-1 play roles in stopping on, crawling on, and transmigrating through vessel walls in the series of neutrophil migration. 37,38 Thus, the low expression of β2 integrin has a negative effect on several steps of neutrophil infiltration into the spinal cord.
Taken together, the neutrophil-free inflammation in the neonates of this study was from the simultaneous lower expression of CXCR2 and β2 integrin. This fact suggests that combined inhibition of CXCR2 and β2 integrin can be considered to suppress post-SCI inflammation in adults as much as in neonates.
To date, no study has evaluated the properties of neutrophils in neonates after SCI in comparison with adults. Through a comprehensive understanding of the age-dependent differences in the functional properties of neutrophils, we may be able to achieve neonatal-like reduced inflammation and vigorous axon regeneration with favorable functional recovery, even in the adult spinal cord.
Spinal cord regeneration in mature mammals has never been reported. In opossums soon after birth, however, spinal cord regeneration after injury has been observed, although they belong to a mammalian class. 39 It has also been reported that in neonatal mice with SCI, the spinal cord is histologically repaired after microglia-mediated fibronectin secretion. 40 These facts suggest that the mammalian spinal cord has regenerative potential soon after birth.
In the present study, we have advanced the research on this topic by confirming that neonatal mice with SCI demonstrate axon regeneration in the context of extremely low inflammation that is characterized by a lack of neutrophil infiltration. Further, we proved that neutrophil-free inflammation in neonates results from the lower expression of chemokines from astrocytes and the lower expression of chemokine receptors and adhesion molecules on neutrophils in comparison with adults.
In a study using young (age: 4 weeks) mice with SCI, fewer neutrophils infiltrated into the injured spinal cord of young mice in comparison with adult mice. 20 It has also been reported that when IL-1β was administered to the brain parenchyma of rodents of varying ages, the vascular permeability and the number of infiltrating neutrophils differed according to the maturity of the animals, indicating that the sensitivity to IL-1β depends on age. 41 These facts, coupled with our results, suggest that aging is a factor that affects the amount of neutrophil infiltration after SCI.
Astrocytes may be related to the age-dependent change in neutrophil migration. Because the astrocyte nuclear NF-κB binding capacity in the brain changes in an age-dependent manner, 42 NF-κB-mediated chemokine release from astrocytes and neutrophil infiltration after SCI may be affected by aging. Taken together, elucidation of the age-dependent pathophysiology of inflammation after SCI is important for optimal inflammation control and the effective prevention of secondary injury.
Considering the fact that neutrophil infiltration has been observed—to some extent—after CNS injury in 9- and 10-day-old mice, 10,43 the 2-day-old mice with SCI used in this study are an extremely optimal model for analyzing the mechanisms of successful suppression of neutrophil infiltration and subsequent inflammation.
Although astrocytes induce neutrophil infiltration through NF-κB-mediated chemokine release after CNS injury, 12,23,27 the functional state of NF-κB in neonatal astrocytes after SCI had remained unclear. Our study revealed that NF-κB nuclear translocation and the resulting chemokine release after SCI were milder in neonatal astrocytes. Interestingly, the nuclear translocation of NF-κB in adult astrocytes was accompanied by morphological changes, such as hypertrophy and process elongation, indicating a reactive astrocyte phenotype. 14
This NF-κB-mediated morphological change in astrocytes has also been observed in damaged brain white matter of a mouse model of vascular dementia, where astrocytes with transgenic NF-κB inhibition did not show the reactive astrocyte phenotype. 44 In addition, the gene expression profile of reactive astrocytes in LPS-stimulated brain or hypoxia-damaged brain contains higher NF-κB levels in comparison with naïve astrocytes. 45
Thus, NF-κB in astrocytes after CNS injury has a dual role in the activation of astrocytes and the expression of chemokines. When naïve astrocytes become reactive astrocytes, they form a glial scar through interaction with type 1 collagen in the injured spinal cord, 14 whereas astrocytes before activation do not interact with type 1 collagen. For this reason, the inhibition of astrocyte activation is important to prevent glial scar formation. 14 Further, mice with selective transgenic inhibition of astrocyte NF-κB show reduced lesion volumes and a reduction in chondroitin sulfate proteoglycans, which participate in glial scar formation. 46
These facts suggest that NF-κB signaling in astrocytes is the optimal therapeutic target for the prevention of glial scar formation after astrocyte activation, as well as neutrophil infiltration by chemokine secretion. Accordingly, reproduction of neonate-like inflammatory regulation in adult SCI may be beneficial for preventing glial scar formation as well as secondary injury after SCI.
The present study was associated with several limitations. First, we did not examine the effect of sex differences on the results of the study because it is often difficult to determine the sex of mice on post-natal day 2. Second, while we showed a correlation between juvenile astrocytes and spinal cord repair in neonatal mice, we did not show direct causality. To confirm their direct causality, a method to juvenilize astrocytes in the adult spinal cord or to make astrocytes in neonatal mice senescent might be established.
Conclusion
Astrocytes in neonatal mice after SCI show low activation and release low levels of NF-κB-mediated chemokines, such as CXCL1 and CXCL2. In addition, in neonatal neutrophils, the expression of the chemokine receptor CXCR2 and the adhesion molecule β2 integrin is lower than that in adult neutrophils. These neonatal specific cellular properties were correlated with a neutrophil-free inflammatory response with the reduced expression of inflammatory cytokines (e.g., IL-6, IL-1b, and TNF-α) after SCI accompanied with attenuated secondary injury and remarkable spinal cord regeneration.
Our findings provide deeper insight into the role of astrocytes and neutrophils in neuroinflammation and axon regeneration, highlighting the importance of a neonate-like neutrophil-free inflammatory response after traumatic SCI.
Transparency, Rigor, and Reproducibility Summary
Five to eight mice per group were used to detect both an overall significant effect of the repeated-measures ANOVA (the primary statistical analysis) and a post hoc difference between the best performing experimental group vs. control with p < 0.05 after correction for multiple comparisons. Overall, 173 mice were subjected to experimental injury, including neonates and adults. Five were excluded for technical reasons, and 12 neonates died after SCI. Complete data were obtained from 156 mice. Both neonates and adults were randomly assigned to the naïve group or SCI group by using a random number generator.
Ethics Approval
All surgical procedures and experimental manipulations were approved by the Committee of Ethics on Animal Experimentation in the Faculty on Medicine, Kyushu University (A-22-238-0). Experiments were conducted under the control of the Guidelines for Animal Experimentation.
Availability of Data and Materials
All data generated or analyzed during this study are included in this published article.
Footnotes
Acknowledgments
Aldh1l1-EGFP transgenic mice were kindly provided by Y. Gotoh (Tokyo University, Tokyo, Japan). We thank The Research Support Center, Research Center for Human Disease Modeling, Kyushu University Graduate School of Medical Sciences, for technical assistance regarding flow cytometry. We are also grateful to J. Kishimoto (Center for Clinical and Translational Research, Kyushu University Hospital) for supervision of the statistical analyses.
Authors' Contributions
K. Kitade: Conceptualization (lead); investigation; formal analysis; writing – original draft (lead). K. Kobayakawa and H. Saiwai: Supervision; funding acquisition; writing – original draft (supporting). K. Kijima, H. Iura, T. Tamaru, Y. Haruta, and G. Ono: Conceptualization (supporting); writing – review and editing (equal). Other authors: Writing – review and editing (equal).
Funding Informaton
This work was supported by JSPS KAKENHI Grant Number JP22K19587, JP22K09426, and JP22H03200; The General Insurance Association of Japan; ZENKYOREN (National Mutual Insurance Federation of Agricultural Cooperatives). The funders had no role in the study design, data collection, data analyses, interpretation, or writing of the article.
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.