
Abstract
The hippocampus plays a prominent role in learning and memory formation. The functional integrity of this structure is often compromised after traumatic brain injury (TBI), resulting in lasting cognitive dysfunction. The activity of hippocampal neurons, particularly place cells, is coordinated by local theta oscillations. Previous studies aimed at examining hippocampal theta oscillations after experimental TBI have reported disparate findings. Using a diffuse brain injury model, the lateral fluid percussion injury (FPI; 2.0 atm), we report a significant reduction in hippocampal theta power that persists for at least three weeks after injury. We questioned whether the behavioral deficit associated with this reduction of theta power can be overcome by optogenetically stimulating CA1 neurons at theta in brain injured rats. Our results show that memory impairments in brain injured animals could be reversed by optogenetically stimulating CA1 pyramidal neurons expressing channelrhodopsin (ChR2) during learning. In contrast, injured animals receiving a control virus (lacking ChR2) did not benefit from optostimulation. These results suggest that direct stimulation of CA1 pyramidal neurons at theta may be a viable option for enhancing memory after TBI.
Introduction
The hippocampus plays a critical role in learning and memory and spatial navigation. 1,2 The functional integrity of the hippocampus has been shown to be compromised by traumatic brain injury (TBI), with the degree of cognitive impairments correlating with injury severity. 3 –5
One aspect of hippocampal function that has received attention in recent years is hippocampal place cells and the modulation of their activity by theta oscillations (6–12 Hz). 6,7 It is thought that theta oscillations function to coordinate and organize hippocampal neuronal activities that are encoding spatial and other information. 8 –11
Studies have shown that experimental TBI can reduce theta power in rats and mice. 12 –15 Further, it has been reported that neuronal entrainment to theta is compromised after TBI, and that the natural recovery of theta power correlates with improved cognitive function. 14,16 We therefore questioned whether optogenetic stimulation of CA1 pyramidal neurons at theta could be used to improve memory in brain injured animals.
Results/Methods
Using a novel environment task, Ondek and colleagues reported that both memory and theta power normalize by seven days after lateral fluid percussion injury (2.1 atm FPI) 16 . In contrast, Fedor and associates reported a lasting suppression of theta power (at least six weeks post-injury) in lateral FPI (2.16 atm) rats performing the Barnes maze. 17 Cohen and coworkers have observed that 1.5 atm lateral FPI delivered to mice weakens broadband power, and that theta power is not selectively reduced. 12
To address these discordant findings, male rats (12–16 weeks old; n = 5/group) were given a lateral FPI (2.0 atm) or sham surgery. Sham animals received anesthesia but did not receive a craniectomy or hub implantations. Seven days after injury, sham and injured rats were implanted with 32-microwire arrays (BioSignal, Dallas, TX) directed at the dorsal CA1 subfield of the hippocampus (3.8 mm posterior to bregma, 2.0 mm lateral from the midline, and a depth of 2.2 mm).
Local field potentials (LFPs) from a total of 160 channels were isolated over three sessions of the novel location (NL) task per animal (Fig. 1a). For each raw LFP trace, the power spectral density (PSD) was computed using a Welsh periodogram in Neuroexplorer (Nex Technologies, Colorado Springs, CO). 18 The raw local field potential was split into data segments of 2*sampling frequency, with a 10% overlap.
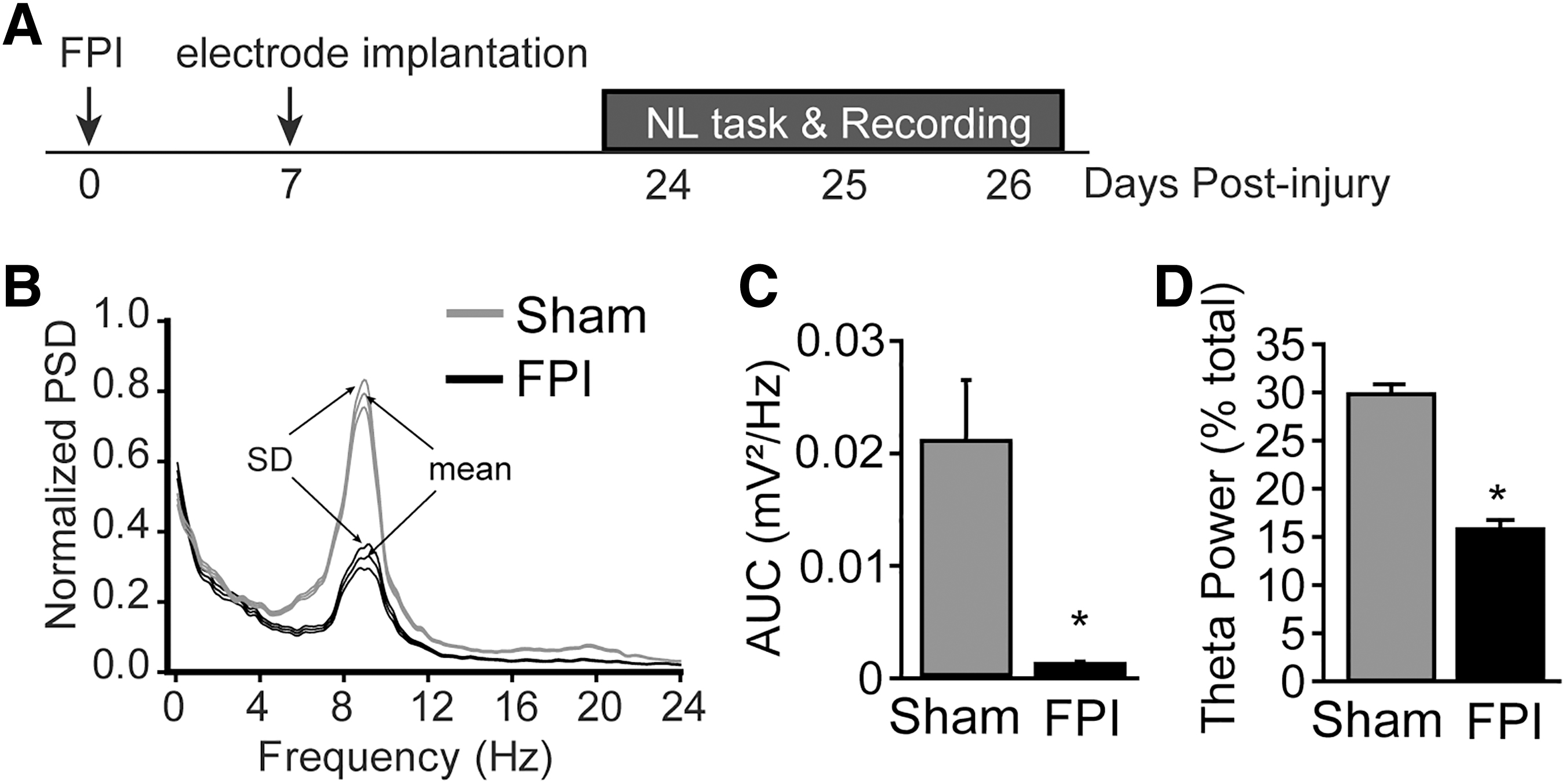
Lateral fluid percussion injury (LFPI) reduces hippocampal theta power that persists for at least three weeks. (
A discrete Fourier transform was then applied to each segment, the squared magnitude of the result computed, and averaged. Figure 1B shows the normalized PSD (as a % of total) from 0–24 Hz for sham (gray line) and mFPI (black line) rats while the animals were actively engaged in the NL task.
To quantify changes in hippocampal theta, the area under the curve (AUC, between 6-12 Hz) was calculated using the trapezoidal method in Microsoft Excel in which we binned the PSD values and summed the bins. For bin x, the PSD amplitude is added to the PSD amplitude of the previous bin, and then the result is divided by 2. The sum of these values is the estimate of the AUC.
When the integration of PSD values within the theta band (6–12 Hz) was compared across groups, a significant reduction was observed in FPI animals (Fig. 1C; F = 4.96, p < 0.05; n = 5/group). This difference was still observed when theta power was normalized by total broadband power (0–250 Hz) (Fig. 1D; F = 57.10, p < 0.01; n = 5/group).
Optogenetic stimulation improves recognition memory
To bypass the upstream pathologies that may contribute to reduced theta generation, we tested whether direct optogenetic stimulation of hippocampal CA1 pyramidal neurons at theta could be used to improve recognition memory. The timeline for these experiments is shown in Figure 2A. Seven days after FPI, injured rats (n = 8) received bilateral, stereotaxic injections of AAV5-CaMKIIα-ChR2-mCherry (University of North Carolina Viral Vector core, 4.9

Optogenetic stimulation of pyramidal neurons at theta frequency improves memory in lateral fluid percussion injury (LFPI) rats. (
To control for non-specific effects of viral infection or optostimulation, a group of injured rats (n = 8) was bilaterally injected with the same number of vg lacking the ChR2 gene (AAV5-CamKIIα-mCherry). The LED probes (10–12 mW; Amuza, San Diego, CA) were implanted bilaterally above the hippocampi immediately after virus injections.
Because FPI has been shown previously to disrupt theta in both the ipsilateral and contralateral hippocampi, 17 we reasoned that bilateral optogenetic stimulation would be required to restore hippocampal function and improve learning and memory. Two weeks after viral injection (to allow for cargo gene expression), rats were trained in a NL task by placing them in a chamber containing two identical objects for a period of 5 min. 19,20
Figure 2B shows that the injured rats receiving either the AAV5-CaMKIIα-ChR2-mCherry virus or the control virus (AAV5-CaMKIIα-mCherry) explored the two objects equally during training, indicating no bias for the objects or their positions in the arena. Location memory was tested 24h later by moving one of the objects to a new position within the chamber and monitoring the time the rat spent exploring each of the two objects. In the absence of optogenetic stimulation, no difference in exploration time between the stationary and displaced objects was observed in either group, indicating impaired location memory.
Using a new set of objects and positions within the chamber, training was again performed. A 5 msec, 472 nm optical pulse was delivered at theta frequency (7.7 Hz) for a total duration of 1 sec each time an animal approached an object during exploration (30 ± 3.98 stimulation events/session). The stimulation frequency (7.7 Hz) was chosen because it represented the peak of theta in our recordings. The pulse duration (5 msec) was based on the minimal optical stimulation to depolarize pyramidal neurons. 21
Figure 2C shows that optostimulation during training did not have any influence on the exploration of the two objects in either group. When tested for their memory 24h later, injured animals expressing the control virus did not benefit from the stimulation during training and still displayed impaired location memory. In contrast, mFPI rats that received the AAV5-CaMKIIα-ChR2-mCherry virus showed a significant preference for the object in the new location (Fig. 2C; t = 3.12; p < 0.05), indicating enhanced NL memory.
One week later, we tested the same animals in a novel object recognition (NOR) task in the absence and presence of optostimulation. Similar to that seen in the NL task, both groups showed no preference for the novel object during memory testing in the absence of theta stimulation during training (Fig. 2D). In contrast, when tested for NOR memory 24h after optostimulation, the group receiving the AAV5-CaMKIIα-ChR2-mCherry virus, but not the group receiving the control virus, displayed improved recognition memory as indicated by significantly more time spent exploring the novel object (Fig. 2E; t = 3.12; p < 0.05).
Interestingly, the performance of the injured rats receiving the AAV5-CaMKIIα-ChR2-mCherry virus and optogenetic stimulation was similar to that seen in uninjured rats (Fig. 2F).
After the completion of memory testing, animals were humanely euthanized, and 40 μm brain sections were prepared and used for immunohistochemical analysis. Figure 2G shows the expression of mCherry in the CA1 pyramidal (pyr) layer in an AAV5-CaMKIIα-ChR2-mCherry-injected animal. The outlined area (above the hippocampus) shows the location of the LED probe.
Staining for NeuN (green) showed that all mCherry-positive (red) cells were neurons. No expression was observed in either GFAP-positive astrocytes (blue) or parvalbumin (PV)-positive inhibitory neurons (purple). This staining pattern was observed in all animals examined.
Discussion
A large body of experimental evidence has demonstrated a role for the medial septal area in generating hippocampal theta oscillations (7–12 Hz), with pharmacological inactivation of the medial septum blocking theta oscillations in the hippocampus. 22,23 Electrophysiological recordings indicate that PV-expressing inhibitory neurons in the medial septum are responsible for generating hippocampal theta. 24 These PV-expressing inhibitory neurons give rise to long-range projections that terminate on hippocampal PV-expressing inhibitory neurons, 25 –27 which in turn, provide strong synchronous perisomatic inhibition of CA1 pyramidal neurons. 28 –30
The modulation of CA1 pyramidal neurons by theta oscillations may facilitate the formation of place fields and organize them in temporal sequences that allow the animal to determine its position in space, to encode memories, and for navigational planning. Consistent with the previous study by Fedor and associates, 17 we found that 2.0 atm FPI resulted in a significant and persistent reduction in hippocampal theta power that lasted at least three weeks after injury. This reduction in theta was observed even after the theta power was normalized to total broadband power. Although the reason for this reduction is not known at present, previous studies have reported the loss of septal cholinergic neurons and hippocampal parvalbumin neurons following FPI in rats, changes that could give rise to compromised theta power. 20, 31
A few studies have used electrical stimulation of the medial septum and fornix and have observed improved spatial memory when theta burst stimulation was employed. 13,15,32,33 It is not known, however, whether direct stimulation of hippocampal excitatory neurons is sufficient to improve hippocampus-dependent memory.
The present study employed an optogenetic approach to directly stimulate CA1 pyramidal neurons at theta to bypass the consequence of TBI-triggered loss of cholinergic and inhibitory neurons. Our results indicate that optogenetic stimulation can improve memory in two different tasks—the novel location and novel object recognition tasks. This nootropic effect was not observed in injured rats receiving a control virus, indicating that it is not the result of increased experience in the task or other influences of the light pulses. Although we observed enhanced memory as a result of optogenetic stimulation, the mechanism by which neuroplasticity was increased is not known at present.
It has been recently reported that direct stimulation of CA1 pyramidal neurons can trigger these cells to become place cells. 34 Thus, in addition to strengthening place cell ensemble activity, theta stimulation may have recruited additional cells to form place fields in the immediate vicinity of the objects. Because some place cells (i.e., landmark-vector cells) have been shown to link to objects in an environment, 35,36 this may have allowed the theta-stimulated animals to better recognize changes within their environment.
The present studies have a number of weaknesses. One weakness of this study is that a group of sham animals were not included for optogenetic stimulation. Optogenetic experiments manipulating theta rhythms in the hippocampus of normal rodents have been reported previously, however. Specifically, stimulating at theta rhythms during encoding has been shown to enhance recall in a spatial memory. 37,38 Conversely, optogenetically interfering with hippocampal theta has been shown to impair spatial memory in normal mice. 39
Because the purpose of this study was to determine whether optogenetic stimulation of pyramidal neurons at theta could be used to bypass the reduction in theta seen after mFPI, we did not perform similar experiments in uninjured animals.
Another limitation of this study was that we did not perform simultaneous recordings of the LFP and spiking neurons concurrent during optogenetic stimulation. Future studies using optoelectrodes that can simultaneously stimulate and record from neurons 40 will be used to address the specific neurophysiological changes that underlie the memory-enhancing effects we observed.
Finally, it remains to be determined whether the present strategy has direct clinical utility. Because the approach utilized herein involved viral delivery of the channelrhodopsin gene and optical stimulation during learning, an alternative means of stimulating pyramidal neurons may be required for use in humans. Currently, six clinical trials are investigating the utility of opsin-based optogenetic therapy, but these are restricted to the restoration of retinal function. 41 Coupling optogenetic approaches to deep brain stimulation in humans provides a unique set of challenges and one estimate suggests that clinical trials are still 5–10 years in the future. 42
Although still in a pre-clinical stage, our study provides mechanistic support for optogenetic stimulation of the hippocampus for improving learning and memory in neurodegenerative states. 43
Transparency, Rigor, and Reproducibility Summary
The study and analysis plan were not preregistered, as the data used in the manuscript had been collected prior to policy implementation. A sample size calculation in a two-group comparison indicated that 5 rats per group would be sufficient to detect a difference in theta power that is 2.5 times the standard deviation with 90% power and an alpha of 0.05. Based on previous studies examining theta power after brain injury, this difference (2.5 times the standard deviation) was anticipated to be satisfied in the present study. Thus, 5 animals/group was used to examine theta power between injured and uninjured animals. For the behavioral studies, comparisons are made within groups to determine if animals display a significant preference for one object over another. Using the recorded deviation for the behavioral parameters to be used in this study (i.e. object exploration), it was found that the standard deviation of the residuals could be anticipated to be between 16% and 21% of the mean. Assuming a minimally detectable effect size of 30% between 2 groups, the number of animals required to obtain statistical significance (with a power of 90% and an alpha value of 0.05) was found to range between 8 to 12 animals/group. All analysis was performed by team members blinded to the injury condition or the viral vectors injected. All rats were tested between 0800 and 1300 in a fed state. Histological analyses were performed in 2 batches, with samples assigned to batches randomly. Whenever possible, antibodies that had been validated against knock-out animals were utilized. The specificity of each antibody was confirmed by comparing the resultant immunoreactivity to published reports. Upon publication, the data from this study will be deposited into Synapse. The analytical codes used for LFP analysis are components of Neuroexplorer (available from
Footnotes
Authors' Contributions
J.I.B., J.B.R., J.Z., and P.K.D. were involved in the conceptualization of this study, and review, and approval of the analysis and manuscript. J.Z. performed surgery. J.I.B. conducted neurophysiological recordings and optogenetic testing. R.W. performed behavioral testing. R.H. and J.I.B. conducted statistical analysis on the neurophysiological data. J.B.R. performed immunohistochemistry and microscopy. P.K.D performed the editing and drafting of the manuscript.
Funding Information
This work was supported by funds made available to PKD by NIH (NS118329) and to JB by TIRR Foundation.
Author Disclosure Statement
No competing financial interests exist.