
Abstract
Blast-induced traumatic brain injury (bTBI) has been identified as the signature injury of Operation Iraqi Freedom and Operation Enduring Freedom. Although the incidence of bTBI increased significantly after the introduction of improvised explosive devices, the mechanism of the injury is still uncertain, which is negatively impacting the development of suitable countermeasures. Identification of suitable biomarkers that could aid in the proper diagnosis of and prognosis for both acute and chronic bTBI is essential since bTBI frequently is occult and may not be associated with overtly detectable injuries to the head. Lysophosphatidic acid (LPA) is a bioactive phospholipid generated by activated platelets, astrocytes, choroidal plexus cells and microglia and is reported to play major roles in stimulating inflammatory processes. The levels of LPA in the cerebrospinal fluid (CSF) have been reported to increase acutely after non-blast related brain injuries. In the present study, we have evaluated the utility of LPA levels measured in the CSF and plasma of laboratory rats as an acute and chronic biomarker of brain injury resulting from single and tightly coupled repeated blast overpressure exposures. In the CSF, many LPA species increased at acute time-points, returned to normal levels at 1 month, and increased again at 6 months and 1 year post-blast overpressure exposures. In the plasma, several LPA species increased acutely, returned to normal levels by 24 h, and were significantly decreased at 1 year post-blast overpressure exposures. These decreases in LPA species in the plasma were associated with decreased levels of lysophosphatidyl choline, suggesting a defective upstream biosynthetic pathway of LPAs in the plasma. Notably, the changes in LPA levels in the CSF (but not plasma) negatively correlated with neurobehavioral functions in these rats, suggesting that CSF levels of LPAs may provide a suitable biomarker of bTBI that reflects severity of injury.
Introduction
Blast-induced traumatic brain injury (bTBI) has been identified as a significant concern for both military and civilian populations due to the widespread, repeated use of improvised explosive devices (IEDs). In the military, 80% of the brain injuries with mild TBI (mTBI) are related to blast exposure. 1 The effects of mild TBI include symptoms such as headache, dizziness, fatigue, fogginess, as well as impairments in cognitive, vestibular, oculomotor, and psychological functions, which may evolve into chronic symptoms in 15 to 30% of the cases. 2 –6
Many of the chronic symptoms of mTBI overlap with those of post-traumatic stress disorder (PTSD), which are reported in up to 40% of service members who have had mTBI. 7 War-related mTBI often is associated with chronic neurocognitive impairments and symptoms of PTSD, which are exacerbated by previous exposures to blast. 8 Moreover, blast exposure has also been reported to cause chronic white matter abnormalities, which are associated with long-term memory impairment. 9,10 Causes of bTBI are multi-faceted in that the often invisible wound to the brain may result from direct and/or indirect (reflective blast waves) exposure to blast. Use of IEDs, occupational blast training, and heavy munitions firing are common sources of blast injuries experienced in the military. Reported data suggest that there are differences in the pathophysiology between blast and non-blast related TBI. 11 -13 One of the difficulties in assessing blast injuries is the different spectra of clinical manifestations that can appear due to injury from a direct blow to the head, transmission of blast overpressure waves through the brain, or a combination of both. 14,15
While the acute symptoms of bTBI are typically readily apparent and can be treated symptomatically, chronic sequelae are harder to recognize. The acute symptoms in victims exposed to blast are treated symptomatically and the patients are typically discharged after a few days/weeks, but many of those patients then develop significant neurological deficits during ensuing months or years. 16 The proper diagnoses for individuals with bTBI is greatly impaired in large part due to the lack of sensitive and reliable biomarkers, which correlate with the severity of injury. Identifying suitable biomarkers that could aid in the diagnosis of and prognosis for chronic bTBI is urgently needed in the population of military veterans who are receiving assessment and care often years after blast exposure.
Several protein biomarkers have been proposed for bTBI diagnosis and prognosis. 17 –23 However, most of those previous pre-clinical studies that focused on identifying biomarkers for the diagnosis of bTBI terminated at acute and sub-acute time-points after blast exposure because the measured biomarker changes were transient and returned to near normal levels at sub-acute time-points, such as a few weeks post-blast. Recently, using a rat model of bTBI, we have shown that the levels of phosphorylated neurofilament heavy chain in the cerebrospinal fluid can be a suitable biomarker of bTBI since its levels correlated with the acute and chronic neurobehavioral changes observed in the experimental animal subjects from which the samples were obtained. 24
Lysophosphatidic acid (LPA), which is reported to play major roles in stimulating inflammatory processes, 25,26 is a bioactive metabolite of phospholipids (i.e., phosphatidyl choline) generated by the sequential actions of phospholipase A2 followed by lysophospholipase D (autotaxin) within activated platelets, astrocytes, choroidal plexus cells and microglia in response to stress and/or injuries to the body. LPAs stimulate astrocytic proliferation, neuroinflammation, neuropathic pain, demyelination and, depending upon their concentration, can promote death of neurons by apoptosis or by necrosis. 27 –33 Multiple LPA species have been identified in the cerebrospinal fluid (CSF), plasma, and serum. LPAs mediate a series of signaling events through specific G-protein coupled LPA receptors. Increased levels of LPAs in the CSF and modulation of LPA receptors occur in different regions of the brain acutely after non-blast related TBI. 34,35 No prior studies evaluated the levels of LPAs in the CSF or plasma at chronic time-points after either blast or non-blast-induced TBI. In the present study, using a rat model of bTBI utilizing an advanced blast simulator, we have evaluated the levels of LPAs in the CSF and plasma for their suitability as a sensitive and reliable biomarker for bTBI diagnosis and prognosis.
Methods
Animals
Research was conducted under an Institutional Animal Care and Use Committee (IACUC)-approved animal use protocol in an Association for Assessment and Accreditation of Laboratory Animal Care (AAALAC) International–accredited facility with a Public Health Services Animal Welfare Assurance and in compliance with the Animal Welfare Act and other federal statutes and regulations relating to laboratory animals. Male Sprague Dawley rats, 9-10 weeks old that weighed 300-350 g (Charles River Laboratories, Wilmington, MA) were housed at 20-22°C (12-h light/dark cycle). Rats were given free access to nutritious rat chow (Prolab IsoPro RMH 3000 from LabDiet, St. Louis, MO) and water ad libitum till 1 month after the blast exposure, when they reached a body weight of 400-450 g. Rats including sham controls were provided restricted diet after 1 month post-blast exposures so that the weights of the rats were maintained between 450 and 500 g until the completion of the study (1 year). Body weights were recorded 3 days a week and adjustments were made in the quantity of diet to maintain body weights within the range identified above since these animals were also used for neurobehavioral functional tests summarized in a previous report. 36
Primary blast exposure
For primary blast overpressure exposure, the previously described advanced blast simulator (ABS) was used. 24,36 For blast overpressure exposure, the rats were anesthetized with 4% isoflurane for 8 min and secured in a longitudinal prone orientation (i.e., rat facing the oncoming shockwave) in the test section of the ABS. To produce moderate injury in rats in these experiments, we used Valmax membranes yielding peak positive static pressures of approximately 19 psi with a positive phase duration of 4-5 msec. For tightly coupled repeated blast overpressure exposures, the rats were subjected to two 19 psi blast overpressure waves separated by 2 min as described previously. 24,36 Sham control rats were subjected to isoflurane anesthesia as above but were not exposed to primary blast waves.
Collection of plasma and CSF samples
After blast overpressure exposure, the rats were euthanized at 1 h, 4 h, 24 h, 1 month, 6 months, or 1 year to collect plasma and CSF samples. Animal euthanasia was carried out in accordance with 2020 American Veterinary Medical Association Guidelines for the Euthanasia of Animals. For the collection of plasma, rats were anesthetized with 5% isoflurane for 8 min. The absence of toe pinch reflex was used to judge if the animal was deeply anesthetized. The blood samples were collected by cardiac puncture under anesthesia into vacutainer tubes containing ethylenediaminetetraacetic acid. Plasma was separated by centrifugation at 3000 rpm for 10 min. CSF samples were collected using the procedure described previously. 21 After the collection of blood and euthanasia by exsanguination, the animal carcasses were secured in a stereotaxic frame (Stoelting Inc, Wood Dale, IL), and after a midline incision, the cervicospinal muscle was separated to expose the atlanto-occipital membrane. More than 100 μL of CSF was removed from each rat using a needle puncture of the atlanto-occipital membrane. The plasma and CSF samples were stored at -80°C until analyses.
LPA measurements
LPA measurements in the plasma and CSF samples were carried out as described earlier. 37,38 Briefly, lipids were extracted from plasma using acidified organic solvents. Different LPA species were measured by Ultra High Performance Liquid Chromatography (UHPLC) coupled electrospray ionization tandem mass spectrometry using an AB Sciex 6500 Q-Trap mass spectrometer operated in multiple reaction monitoring mode to identify the molecules based upon their specific precursor and product ion pairs. As an internal standard, 17:0 LPA was used. Plasma/CSF LPA values are reported as μmol per liter.
Lysophosphatidyl choline assay
Levels of lysophosphatidyl choline (LPC), the precursor of LPA, was measured in the plasma at 1 h and 1 year post-blast exposures using the diagnostic kit received from BioVision Inc. (Milpitas, CA), according to manufacturer's instructions. BioVision's LPC assay kit utilizes LPC-specific enzymes to generate an intermediate that then reacts with a probe, yielding a signal that can be quantified either colorimetrically or fluorometrically, and is proportional to the amount of LPC present in the sample.
Statistical analysis
Statistical analysis was performed using a two-way analysis of variance (ANOVA) followed by Tukey's post hoc test using multiple comparisons (GraphPad Prism 6 software). Values are expressed as mean ± standard error of the mean (SEM). A p value less than 0.05 was considered significant and a p value less than 0.01 was considered highly significant.
Results
LPA levels in the CSF after blast overpressure exposure
UHPLC followed by mass spectrometry analysis of CSF samples collected at different intervals post-blast indicated that the levels of several LPA species were increased at acute time-points (1, 4, and 24 h) after single and tightly coupled repeated blasts compared with sham controls (Table 1). At 1 and 4 h post-blast(s), the increases in different LPA species in the CSF were more pronounced in the single blast overpressure exposed rats compared with repeated blast overpressure exposed animals. No significant changes in any LPA species were observed in the CSF at 1 month after single or repeated blast overpressure exposures compared with sham controls. Statistically significant increases in multiple LPA species were observed in the CSF again at 6 months after repeated blast overpressure exposures compared with both sham controls and single blast overpressure exposed rats. At 1 year, the levels of multiple LPA species showed statistically significant increases in the CSF after both single and repeated blast overpressure exposures compared with sham controls.
LPA Levels in the CSF at Different Intervals After Single (B) and Repeated (BB) Blast Exposures
Values are expressed as mean ± standard error of the mean. Values of sham animals were compared to those of single and repeated blast exposed animals (* p < 0.05; ** p < 0.01; n = 6). Values of single blast exposed animals were compared to those of repeated blast exposed animals (# p < 0.05; ## p < 0.01; n = 6).
LPA, lysophosphatidic acid; CSF, cerebrospinal fluid.
LPA levels in the plasma after blast overpressure exposure
Results shown in Table 2A and Table 2B indicate that several LPA species were increased in the plasma at 1 and 4 h after single as well as repeated blast overpressure exposures compared with sham controls. No significant differences in plasma LPA species were observed between the single and repeated blast overpressure exposed rats at 1 or 4 h, nor were statistically significant changes in any plasma LPA species observed among any of the treatment groups at 24 h, 1 month, or 6 months. At 1 year after single and repeated blast overpressure exposures, several LPA species were decreased in the plasma compared with sham controls. Multiple LPA species were significantly lower in the plasma of repeated blast overpressure exposed rats compared with those exposed to a single blast.
LPA Levels in the Plasma at Different Intervals After Single (B) and Repeated (BB) Blast Exposures
Values are expressed as mean ± standard error of the mean. Values of sham animals were compared to those of single and repeated blast exposed animals (* p < 0.05; ** p < 0.01; n = 6). Values of single blast exposed animals were compared to those of repeated blast exposed animals (# p < 0.05; # p < 0.01; n = 6).
LPA, lysophosphatidic acid.
LPA Levels in the Plasma at Different Intervals After Single (B) and Repeated (BB) Blast Exposures
Values are expressed as mean ± standard error of the mean. Values of sham animals were compared to those of single and repeated blast exposed animals (* p < 0.05; ** p < 0.01; n = 6). Values of single blast exposed animals were compared to those of repeated blast exposed animals (# p < 0.05; # p < 0.01; n = 6).
LPA, lysophosphatidic acid.
Lysophosphatidyl choline levels in the plasma
The data shown in Figure 1 indicate that the levels of LPC did not significantly change in the plasma at 1 h after single or repeated blast overpressure exposures compared with sham controls. However, plasma LPC levels decreased significantly in the plasma at 1 year after single as well as repeated blast overpressure exposures compared with sham controls. No statistically significant changes in plasma LPC levels were observed between the blast overpressure-exposed groups at 1 h or 1 year after the exposure.
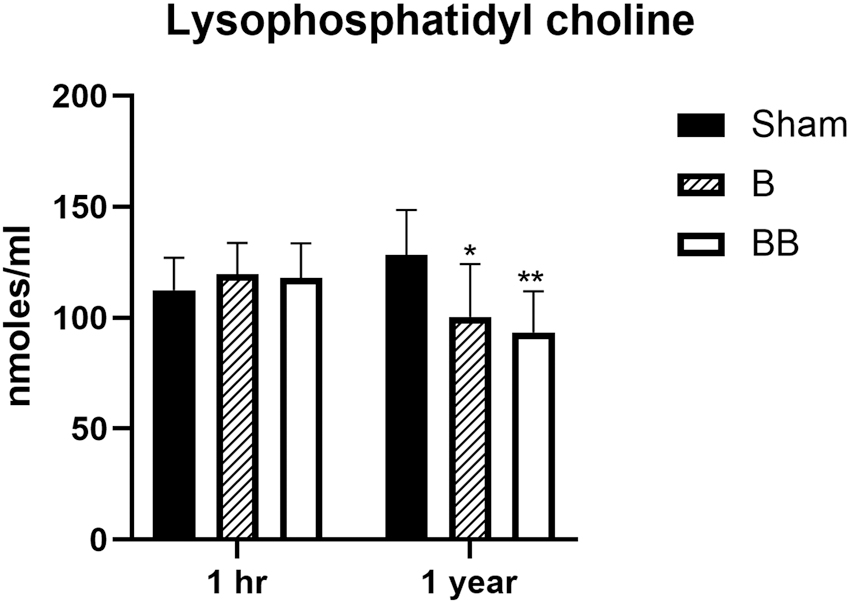
Concentration of lysophosphatidyl choline in the plasma at 1 h and 1 year after single and repeated blast exposures. Values are expressed as mean ± standard error of the mean. Values of sham animals were compared with those of single and repeated blast exposed animals (*p < 0.05; **p < 0.01; n = 6).
Discussion
The results of the current study suggest that CSF levels of LPAs can provide potentially suitable biomarkers for bTBI diagnosis and prognosis. Although prior studies evaluated the utility of LPAs in the CSF as acute biomarkers of non-blast–related TBIs, no studies addressed their suitability with regard to bTBI. Moreover, no previous studies explored the utility of CSF levels of LPA as a chronic biomarker of any forms of TBI. Acute increases in different LPA species were observed in the brain and CSF in a rodent controlled cortical impact (CCI) model of TBI, prompting consideration of CSF levels of LPAs as suitable biomarkers of TBI at acute time-points. 34,39 Additionally, in a mouse CCI model of TBI, a single intravenous administration of antibodies against LPA improved the neuropathological and neurobehavioral outcome measures, implicating the potential role of LPA in the pathogenesis of TBI. 34 The increase in LPA levels in different brain regions acutely after the CCI was found to be associated with cell death and secondary axonal injury, supporting the notion that CSF levels of LPAs could provide an acute biomarker of TBI. 39 LPAs also appear to be important contributors in the pathogenesis of spinal cord injury since treatment with antibodies specific to LPA provided significant neuropathological and functional improvements after spinal cord injury. 40
Total and individual LPA species measured in the CSF collected on Days 1 and 5 from patients who had severe TBI (non-blast and non-penetrating) showed significant increases on Day 1 but not on Day 5 post-injury. 34 LPA(18:0) and LPA(20:4) were the two LPA species found to be increased in the CSF of those TBI patients within 24 h post-injury. Except at 1 month, those two LPA species were significantly elevated at all the time-points up to 1 year in the CSF of rats exposed to blast in the current study (Table 1). In addition to those two LPA species, blast exposure also increased the CSF levels of LPA(16:0), LPA(18:1), LPA(18:2), and LPA(20:3) at all the time-points evaluated except 1 month (Table 1). LPA(18:0) and LPA(18:2) species were also increased at 3 h in the CSF of mice subjected to CCI. 34 Analysis of postmortem brains from normal individuals and patients who died following non-blast related acute closed-head injuries revealed that upregulation of LPA receptors occurs in the brain acutely after the brain injury. 35 The effect of blast overpressure exposure on LPA receptors was not explored in the current study.
Several protein biomarkers have been proposed for bTBI diagnosis and prognosis. 17 –23 However, most if not all, pre-clinical attempts to identify biomarkers suited for bTBI diagnosis terminated at acute and sub-acute time-points since the changes were transient and returned to near normal levels at sub-acute time-points after injury. In the clinical setting, the acute symptoms are addressed and patients are discharged after a few days/weeks, but a subset of those patients then develop significant neurological abnormalities after a few months or years. 16 Detection of biomarkers that could aid in the diagnosis and assessment of chronic mTBI is urgently needed, as little progress has been made toward identification of biomarkers of bTBI during the chronic state.
We have shown previously that blast overpressure exposure increased the levels of neuron-specific phosphorylated neurofilament heavy chain protein (pNFH) in the CSF of rats acutely, followed by a return to normal levels by 1 month and an increase again at 6 months and 1 year, and suggested that CSF levels of pNFH might be a suitable biomarker for acute and chronic bTBI diagnosis and prognosis. 24 The CSF samples used for the analysis of LPAs in the current study were obtained from the same rats used for the pNFH analysis in this previous study. Comparable to pNFH, the levels of LPAs also showed an acute increase followed by a return to normal levels at 1 month, followed by an increase again at 6 and 12 months post-blast (Table 1).
The assessment of neurobehavioral functions in the same rats used for the analysis of pNFH in the previous study 24 and LPAs in the current study indicated that several neurobehaviors changed acutely, followed by a recovery at 1 and 3 months post-blast, after which functional deficits were observed again at 6 and 12 months. 36 Thus, since the time-dependent changes in CSF levels of LPAs parallel the temporal neurobehavioral functional changes in the rats, LPA levels in the CSF appear to be promising candidates as another suitable biomarker of acute and chronic bTBI. Due to the small size of LPAs, they can relatively easily enter the blood circulation from the injured brain without a marked disruption of the blood–brain barrier (BBB). In contrast, due to its substantially larger molecular weight and size, detection of pNFH in the blood circulation using conventional methods requires significant axonal degeneration in combination with a more severely disrupted BBB. It is worth noting that we were unable to detect pNFH in the plasma of these rats at any of the time-points evaluated after blast exposure. 24
At 1 and 4 h, CSF levels of LPAs increased less after tightly coupled repeated blast compared with a single blast overpressure exposure (Table 1). One potential reason for this could be the likelihood that repeated blast overpressure exposure induces more disruption of cellular membranes and BBB, leading to the release of more LPAs into the blood circulation. 41 -43 This speculation is supported by the plasma levels of LPAs at 1 and 4 h. Although statistically not significant, the average values of different LPA species were greater at 1 and 4 h post-blast in the plasma of repeated blast overpressure exposed rats compared with those exposed to a single blast overpressure (Table 2A and Table 2B). The statistical insignificance was due to widely ranging LPA levels among rats within the repeated blast exposed group, which might be attributable to similarly varied disruptions to the cell membranes and BBB in that group of rats. Such an increase in LPAs in the plasma after repeated blasts was not observed at later time-points likely due the fact that the integrity of cell membrane and BBB return to normal levels at later time-points after blast overpressure exposure. 41 -43
Plasma levels of multiple LPA species were increased at 1 and 4 h post-blast, but no significant changes were subsequently observed at 24 h, 1 month, or 6 months (Table 2A and Table 2B). Further, we observed significantly decreased levels of different LPA species in the plasma of rats exposed to blast overpressure waves compared with shams at 1 year (Table 2A and Table 2B). To consider whether this decrease in LPA species might be due to the decreased availability of its precursor, we estimated the levels of lysophosphatidyl choline (LPC) in the plasma at both 1 h and 1 year after blast overpressure exposure. Data shown in Figure 1 indicate that the levels of LPC did not change at 1 h, but statistically significant decreases were observed at 1 year after single and repeated blasts, suggesting that the observed decrease in LPAs at 1 year post-blast could be due to the decreased plasma levels of LPC. In contrast, LPC levels were shown to be increasing in the brain regions at acute and sub-acute time-points after non-blast related injuries. 39,44 The higher levels of LPC in the brain after injury also contribute to the increased CSF levels of LPAs. 39
In a repeated cortical impact model of TBI using humanized Tau transgenic mice, it has been reported that several phospholipids including LPC increased in the cortex and hippocampus at acute time-points post-injury and significant increase in LPC and lysophosphatidylethanolamine observed at 9 months post-TBI. 44 Plasma levels of LPC or other phospholipids were not evaluated in those studies. In another longitudinal study using mouse model of repeated mild TBI, several phospholipids were found to be decreasing in the plasma at chronic time-points, especially at 3, 12, and 24 months post-injury, supporting our current observation. 45
Since blast-induced changes in plasma LPA levels at 24 h, 6 months, and 1 year did not correlate to those measured in the CSF or to the concomitant neurobehavioral changes observed at those time-points post-blast(s), the plasma levels of LPAs do not appear to be suitable candidates as a chronic biomarker of bTBI. This is not surprising, recognizing that blast is a whole–body insult and that the changes in LPAs in the plasma after blast overpressure exposure will also be greatly influenced by the effect of blast overpressure exposure on the peripheral organs and blood cells, especially platelets (where significant quantities of LPAs are being synthesized). Consequently, CSF, more selectively and closely reflecting the ongoing neurobiological changes associated with bTBI, provides a clearer means to gauge these CNS responses.
One of the requirements of this study was the use of food restriction to maintain the body weight of the rats from 1 month post-blast to conduct the behavioral functional tests up to 1 year. Restriction of diet before injury has been shown to promote significantly higher levels of brain-derived neurotrophic factor around the injury site along with improvements in spatial memory functions in a weight drop model of TBI. 46 Post-TBI diet/caloric restriction have also been shown to elicit significant improvements in neuropathological features and neurobehavioral outcome measures in weight drop models of TBI. 47,48 Although sham control rats were also subjected to diet restriction in the current study, future studies are warranted to determine if the diet restriction may have had salutary effects in injured subjects and possibly significantly affected the plasma and CSF levels of LPAs with and without blast overpressure exposures.
Footnotes
Acknowledgments
We acknowledge Zhilin Liao for assistance with euthanasia, and Ondine Eken and Irene Gist for assistance with general laboratory procedures.
Disclaimer: Material has been reviewed by the Walter Reed Army Institute of Research. There is no objection to its presentation and/or publication. The opinions or assertions contained herein are the private views of the author, and are not to be construed as official, or as reflecting true views of the Department of the Army or the Department of Defense. Research was conducted under an IACUC-approved animal use protocol in an AAALAC International–accredited facility with a Public Health Services Animal Welfare Assurance and in compliance with the Animal Welfare Act and other federal statutes and regulations relating to laboratory animals.
Authors' Contributions
Conceptualization: P.A., R.S., and J.B.L. Formal analysis: P.A. and A.J.M. Funding acquisition: J.B.L. and P.A. Investigation: P.A., J.B.L., R.S., and A.J.M. Methodology: P.A., D.M.W., and A.J.M. Project administration: J.B.L. and P.A. Resources: J.B.L. and P.A. Supervision: J.B.L. and P.A. Visualization: P.A. Writing-review and editing: P.A., R.S., A.J.M., and J.B.L. All authors have read and agreed to this version of the manuscript.
Funding Information
This study was funded by the Military Operational Medicine Research Program at United States Army Medical Research and Development Command.
Author Disclosure Statement
No competing financial interests exist.