
Abstract
Traumatic brain injury (TBI) is known to affect the physiology of neural circuits in several brain regions, which can contribute to behavioral changes after injury. Disordered sleep is a behavior that is often seen after TBI, but there is little research into how injury affects the circuitry that contributes to disrupted sleep regulation. Orexin/hypocretin neurons (hereafter referred to as orexin neurons) located in the lateral hypothalamus normally stabilize wakefulness in healthy animals and have been suggested as a source of dysregulated sleep behavior. Despite this, few studies have examined how TBI affects orexin neuron circuitry. Further, almost no animal studies of orexin neurons after TBI have included female animals. Here, we address these gaps by studying changes to orexin physiology using ex vivo acute brain slices and whole–cell patch clamp recording. We hypothesized that orexin neurons would have reduced afferent excitatory activity after injury. Ultimately, this hypothesis was supported but there were additional physiological changes that occurred that we did not originally hypothesize. We studied physiological properties in orexin neurons approximately 1 week after mild traumatic brain injury (mTBI) in 6–8-week-old male and female mice. mTBI was performed with a lateral fluid percussion injury between 1.4 and 1.6 atmospheres. Mild TBI increased the size of action potential afterhyperpolarization in orexin neurons from female mice, but not male mice and reduced the action potential threshold in male mice, but not in female mice. Mild TBI reduced afferent excitatory activity and increased afferent inhibitory activity onto orexin neurons. Alterations in afferent excitatory activity occurred in different parameters in male and female animals. The increased afferent inhibitory activity after injury is more pronounced in recordings from female animals. Our results indicate that mTBI changes the physiology of orexin neuron circuitry and that these changes are not the same in male and female animals.
Introduction
Traumatic brain injury (TBI) is very common; in the United States alone, an estimated 2.5 million emergency department visits are due to TBIs each year. 1 Mild TBI (mTBI) accounts for 75% of these cases, representing a significant number of patients. 2,3 There are a variety of negative symptoms associated with TBI and mTBI, including cognitive dysfunction, memory impairments, and mood disorders. 4,5 Previous research has shown that TBI can have a profound effect on the neural circuitry that underlies these negative symptoms. 6 –13 However, there are some behavioral symptoms that have not been well studied from a physiological perspective. Sleep disorders are a common symptom after TBI; approximately 30-70% of TBI patients will experience sleep disorders after injury. 14 –18 There are known neural circuits that regulate sleep 19 but relatively little work has investigated how TBI affects those circuits.
The physiology of orexin/hypocretin neurons is at the heart of sleep/wake regulation. Orexin/hypocretin (hereafter referred to as orexin) is a neuropeptide produced by neurons in the lateral hypothalamus (LH). 20,21 Reduced levels of orexin cause narcolepsy 22 –25 or hypersomnias 26 and injecting orexin neuropeptide into the brain causes wakefulness, 27,28 showing it is necessary and sufficient to promote wake behavior. In normal animals, orexin neurons stabilize wakefulness by activating wake-promoting neurons 29,30 and it is thought that orexin neurons regulate an animal's arousal threshold. 31,32 28-50% of TBI patients report disorders of hypersomnolence as part of their sleep disruptions after injury 16,33 –36 and it is believed that orexin dysregulation may be a source of this excessive sleepiness after TBI.
There is evidence that orexin is disrupted after TBI. Studies have shown that TBI patients have reduced orexin neuropeptide in their cerebral spinal fluid (CSF). 37 This reduction can last 6 months after injury 17 and correlates with increased excessive daytime sleepiness. In rodents, there is reduced orexin neuropeptide in the cerebral spinal fluid of the hypothalamus immediately following TBI. 38 Some studies suggest that it is a reduction of the number of orexin neurons that explains these decreases in orexin. 38 –43 However, the study that found decreased orexin in the CSF of animals after injury did not observe a decrease in the number of orexin neurons meaning that these two factors do not necessarily correlate with one another. 38
Additionally, our lab reported sleep disturbances after mTBI even when the number of orexin neuron numbers did not decrease, 44 which means that the number of orexin neurons is not the only consideration after injury. Our lab used cFos as a marker of neuronal activity to demonstrate that the activity of orexin neurons is reduced after mTBI, not the number of cells. This shows that it is important to consider orexin neuron function after injury rather than only counting the number of orexin neurons.
Only two previously published studies have examined of the activity of orexin neurons after TBI. The first was the previously mentioned study, Lim and colleagues. 44 The other reported that TBI reduces presynaptic glutamate in dendritic afferents onto orexin neurons. 45 The current study expands on this work and uses electrophysiological techniques together with a transgenic orexin EGFP mouse to determine the mechanism of altered orexin neuronal function after mTBI.
Importantly, sleep is different in males and females. 46,47 Female sex is considered a risk factor for sleep disorders after TBI. 48 Previous studies report that TBI induces changes in sleep in both male 38 –41,44,49 –56 and female animals. 55,57,58 Despite this, little research on orexin after TBI has been done in female animals. Previously, it has been demonstrated that disruptions in homeostasis, such as stress 59 can affect orexin neurons in a sex dependent manner. Sex-dependent changes in orexin could occur with TBI as well since injury disrupts homeostasis. Therefore, this study specifically examines orexin physiology of males and females after mTBI. Based on the two previous functional studies of orexin neurons after injury, we hypothesized that excitatory afferent activity onto orexin neurons would be reduced after mild traumatic brain injury.
Methods
Animals
All experiments were performed in accordance with protocols approved by the Children's Hospital of Philadelphia's Institutional Animal Care and Use Committee (IACUC; Protocol 0694) and the guidelines established by the U.S. Public Health Service's Guide for the Care and Use of Laboratory Animals. Animals were given access to ad libitum food and water. A total of 79 animals had data analyzed for these experiments. 41 Males, 19 Sham and 22 Injured; 35 Females, 17 Sham and 18 Injured, and 3 Male Naïve animals only used for neurobiotin filling and no other data collection. Mice were between 6-8 weeks old at the time of injury. Animals were bred in-house to contain EGFP on the prepro-orexin promoter. 60 Each mouse was genotyped to check for the Orexin-EGFP marker prior to injury and recording. Female animals were checked for estrus cycle stage on the day of sacrifice. There were not enough animals to appropriately compare between estrus cycle stages (Supplementary Table S1); therefore, the female animals were analyzed as one group. For all experiments, the experimenter was blinded to the injury condition of the animal until after analysis of the data.
Surgical procedures in preparation for lateral fluid percussion injury
Animals were anesthetized with a mixture of ketamine (2.6 mg/kg) and xylazine (0.16 mg/kg) via intraperitoneal injection. Animals were given meloxicam (2 mg/kg) as an analgesic prior to surgery. Once fully anesthetized, animals were placed in a stereotaxic frame (Stoelting, Wood Dale, IL, USA), the scalp was incised and pulled away to fully expose the right parietal bone. An ultra-thin Teflon disk, (3-mm diameter) was glued to the skull with Vetbond (3M, St. Paul, MN, USA) between lambda and bregma sutures, and between the sagittal suture and the lateral ridge over the right hemisphere. Guided by the Teflon disk, a trephine was used to perform a 3-mm diameter craniectomy over the right parietal area. Following craniectomy, a Luer-lock needle hub (3-mm inner diameter) was secured above the skull opening with superglue (Loctite, Düsseldorf, Germany) and dental acrylic (Stoelting), filled with saline and capped. Lastly, animals were removed from stereotaxis, placed on a heating pad until fully recovered from anesthesia as assessed by the animal's spontaneous ambulation. Once the animals can walk on their own, they are then returned to their respective home cage. Animals were monitored for 24 h after surgery.
Lateral fluid percussion injury
Twenty-four hours following craniectomy, animals were placed under isoflurane anesthesia (2% oxygen in 500 mL/min) in a chamber and respiration was visually monitored until animals reached a surgical plane of anesthesia (one respiration per 2 sec). At this point, animals were removed from isoflurane, the needle hub was refilled with saline and connected to the fluid percussion injury device (Department of Biomedical Engineering, Virginia Commonwealth University, Richmond, VA, USA, and Custom Design and Fabrication, Sanston, VA, USA) via high-pressure tubing. The animal was placed onto a heating pad on its left side and upon resumption of normal breathing pattern but before sensitivity to stimulation, the injury was induced by a 20-msec pulse of saline onto the intact dura. The pressure transduced onto the dura was monitored with a pressure transducer connected to an oscilloscope, with injury severity ranging between 1.4 and 1.6 atmospheres. This is considered a mild injury. Immediately after injury, the hub was removed from the skull and the animal was placed in a supine position to assess righting reflex. After righting, the animal was subjected to inhaled isoflurane to suture the scalp.
Animals were allowed to recover on a heating pad until mobile, at which point they were returned to their home cage. Animals were given meloxicam post-surgery (5 mg/kg) for pain relief. Sham animals underwent all surgical procedures and anesthesia, including attachment to the fluid percussion injury device, with exclusion of the actual fluid pulse. Average righting times and standard deviations for animals in seconds: Shams: 41.39 ± 31.77; Injured: 275.97 ± 160.53. Divided by sex, the righting times were not significantly different: Male Sham: 30.29 ± 10.76; Male Injured: 233.12 ± 166.50; Female Sham: 50.53 ± 40.02; Female Injured: 321.5 ± 145.3. Injured animals that showed no delay in righting time were not included in the analysis. The injury used is classified as mild, so animals with hemorrhaging, or severe motor dysfunction after injury were not included in the analysis.
Slicing details
All animals used for electrophysiological recordings were sacrificed 6-10 days after the injury or sham-injury. Animals were anesthetized with inhaled isoflurane in a bell jar, decapitated and the brain was removed into an ice-cold bath of sucrose cutting solution. Animals were sacrificed between zeitgeber time (ZT) ZT4 - ZT5 after lights-on in the animal facility, between 10
Electrophysiological recording and recording parameters
After incubation, slices were moved to a slice chamber (Warner Instruments RC-26, Holliston, MA, USA) superfused with aCSF and kept at 29.6°C with an inline heater (Warner Instruments TC-324B). Patch electrodes with resistances of 2-8 MΩ were pulled from borosilicate glass (1B150F-4; World Precision Instruments, Sarasota, FL, USA). Series resistance was monitored throughout the experiment and recordings were not used if a cell exceeded 35 MΩ in series resistance. Series resistance was compensated for at 70%. All recordings were made using a Multiclamp 700B (Molecular Devices, Palo Alto, CA, USA) sampled at 20 kHz, filtered at 3 kHz.
Orexin neurons were initially identified using 2% neurobiotin (Vector Laboratories, Newark, CA, USA) in K-Gluconate internal solution (n = 9 cells; Fig. 1 as an example). After the accuracy of our EGFP label was confirmed, neurons were identified using EGFP at the time of recording. All orexin neurons recorded were selected from the side ipsilateral to the brain injury.
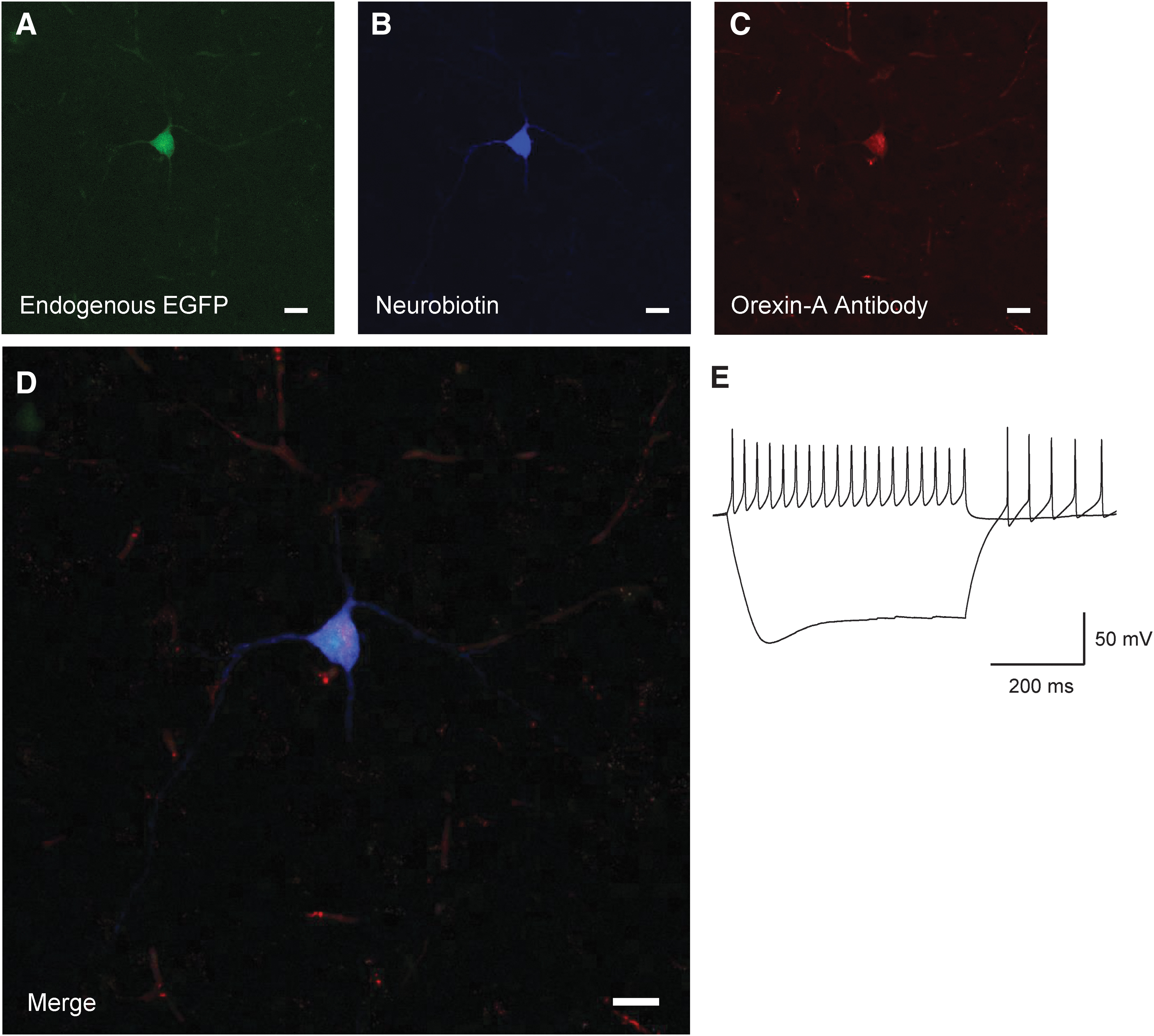
Identification of orexin/hypocretin neurons. Post hoc neuronal staining after recording in Orexin-EGFP mice. EGFP Neuron
Passive membrane properties, action potential properties, and excitatory post-synaptic currents (EPSC) recordings were collected with a K-Gluconate based internal pipette solution. Composition of the K-Gluconate internal solution was (all in mM): K Gluconate 125, KCl 5, EGTA 1.1, HEPES 10, MgCl2 1, Mg-ATP 5, Na-GTP 0.5. Inhibitory post-synaptic current (IPSC) recordings were collected with KCl based internal pipette solution to achieve isotonic chloride recordings. Composition of the KCl internal solution was: KCl 145, EGTA 1.1, HEPES 10, MgCl2 1 Mg-ATP 2, Na-GTP 0.5. In both internals, KOH was used to titrate pH between 7.2 and 7.3. The osmolality of both internals was between 280 and 290 mmol/kg. All measurements described in this study were compensated for the liquid junction potentials of the internal solutions estimated at 14 mV for the K-Gluconate Internal and 3.4 mV for the KCl Internal (Clampex; Molecular Devices).
To isolate excitatory events, EPSCs were recorded in the presence of the Gamma-Aminobutyric Acid (GABA) receptor antagonist (-)- bicuculline methiodide (Abcam, 30 μM, Cambridge, UK). To isolate inhibitory events, IPSCs were recorded in the presence of the N-methyl-D-aspartate (NMDA) channel antagonist DL-2-amino-5-phosphonopentanoic acid sodium salt (DL-AP5, sodium salt; Abcam, 50 μM, Cambridge, U.K.) and the α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) channel antagonist (1,2,3,4-Tetrahydro-7-nitro-2,3-dioxoquinoxaline-6-carbonitrile disodium; DS-CNQX; Abcam, 20 μM, Cambridge, UK). To isolate miniature EPSCs and IPSCs, events were recorded in the presence of the sodium channel antagonist tetrodotoxin citrate (TTX; Abcam, 0.4 μM, Cambridge, U.K.). To test drug efficacy and determine current identity in orexin neurons, IPSCs were blocked with bicuculline and EPSCs were blocked with DS-CNQX and DL-AP5 and no currents were observed (n = 10 cells, data not shown).
Recording measurements
Intrinsic membrane properties and action potential properties were recorded in current clamp. Resting membrane potential was determined by reading the membrane voltage in current clamp between 1-5 sec after cell break in. To measure action potential (AP) firing frequency versus injected current, cells were injected with current steps from -200 pA to 200 pA in increments of 25 pA for a duration of 500 msec. The frequency of AP firing was determined by counting the number of APs in the first 100 msec of the 500 msec pulse during each current step. Input resistance was determined by using the change in voltage during the -100 pA step and calculated with Ohm's law,
All voltage clamp recordings were made while holding cells at -67 mV after adjusting for the internal solution's liquid junction potential. Synaptic events were determined via the Template Search algorithm in Clampfit 11.1 (Molecular Devices). Current events with a 10-90% rise time greater than 1.5 msec were excluded from the analysis. MATLAB (2021a, MathWorks, Natick, MA, USA) code was used to randomly select 75 current events per cell to ensure equal weighting of cells in the cumulative probability histogram analysis. All events from all cells in one group (e.g., Sham vs. Injured) were then pooled to create a cumulative probability histogram. Statistical analyses were performed in Prism 9 (GraphPad, San Diego, CA, USA).
Staining to confirm orexin/hypocretin identity after recording
For cells filled with 2% neurobiotin, post hoc staining was used to confirm the identity of the EGFP neurons. Post-recording, slices were fixed in 4% paraformaldehyde overnight. Slices were washed four times with 0.01M phosphate-buffered saline (PBS) and then blocked in blocking solution of 10% bovine serum albumin, NGS, 0.3% Triton-X and PBS for 3 h. All washes were done with 0.01M PBS. After blocking, slices were washed and incubated in Mouse-On-Mouse Blocker (1:40 concentration; Vector Labs, Newark, CA, USA) for 2 h at room temperature. Slices were washed and Orexin-A Antibody (1:1000; KK09, Santa Cruz Laboratories, Dallas, TX, USA) was added to incubate overnight at room temperature. Slices were washed and incubated with 594 goat-anti rabbit (1:200) and 405 Dylight Conjugated Streptavidin (1:300; Jackson ImmunoResearch, West Grove, PA, USA) at room temperature for 3 h and then at 4°C overnight. Slices were washed and mounted in Fluoromount-G (Southern Biotech, Birmingham, AL, USA) and imaged with a Leica Sp8 confocal microscope (Leica, Deer Park, IL, USA).
Statistical analysis
Statistical tests for current clamp recordings including intrinsic properties and membrane potential properties included unpaired Student's t-tests comparing means of these properties, or mixed-effects analysis where appropriate. For these tests, values of p < 0.05 were considered significant (Table 1). For voltage clamp recordings, Kolmogorov-Smirnov tests were performed on the pooled cumulative probability histograms and compared between sham and injured groups. Significant differences in distributions were set at a threshold p value of p < 0.001 or less to be considered significant due to the large number of events (Table 2).
Current Clamp Values
Bolded data indicates a significant finding.
All properties and significance values recorded in current clamp of orexin/hypocretin neurons. Values reported as means with standard error of the mean.
AP, Action potential.
Voltage Clamp Significance
Bolded data indicates a significant finding.
Significance of cumulative probability histograms of orexin/hypocretin neurons recorded in voltage clamp. Values computed with the Kolmogorov-Smirnov test with a value of p < 0.001 to be considered significant.
sEPSC, spontaneous excitatory post-synaptic current; mEPSC, miniature excitatory post-synaptic current; sIPSC, spontaneous inhibitory post-synaptic current; mIPSC, miniature inhibitory post-synaptic current.
Results
Identification of orexin/hypocretin neurons for electrophysiological recording
We used transgenic mice with enhanced green fluorescent protein (EGFP) downstream of the prepro-orexin promoter to visually identify orexin neurons. 60,62 To initially confirm their identity, we performed visualized whole–cell patch clamp on EGFP labeled neurons (Fig. 1), filled a subset with neurobiotin (n = 9 cells) and co-stained with orexin-A antibody (Fig. 1D) post hoc. Recorded EGFP labelled neurons show the previously established electrical properties of orexin neurons. 63 -65 That is, spontaneously firing action potentials, non-accommodating action potentials and a hyperpolarizing sag in response to injection of negative current (Fig. 1E).
Increased afterhyperpolarization in female animals and reduced action potential firing threshold in male animals after mild traumatic brain injury
We next examined if mTBI altered the intrinsic membrane properties or action potential firing of orexin neurons (Fig. 2). In current clamp recordings from female animals, we observed a larger after hyperpolarization after mTBI (Fig. 2B). This result was not observed in neurons from injured male animals (Fig. 2A). We also observed a reduction in the action potential firing threshold in orexin neurons from male animals after mTBI (Fig 2C), but we did not see this reduction in firing threshold in neurons from female animals (Fig 2D). No other significant differences between intrinsic properties in slices from sham and injured animals were detected (Table 1; Males: Sham = 9 animals, 19 cells; Injured = 11 animals, 23 cells; Females: Sham = 7 animals, 13 cells; Injured = 8 animals, 14 cells]
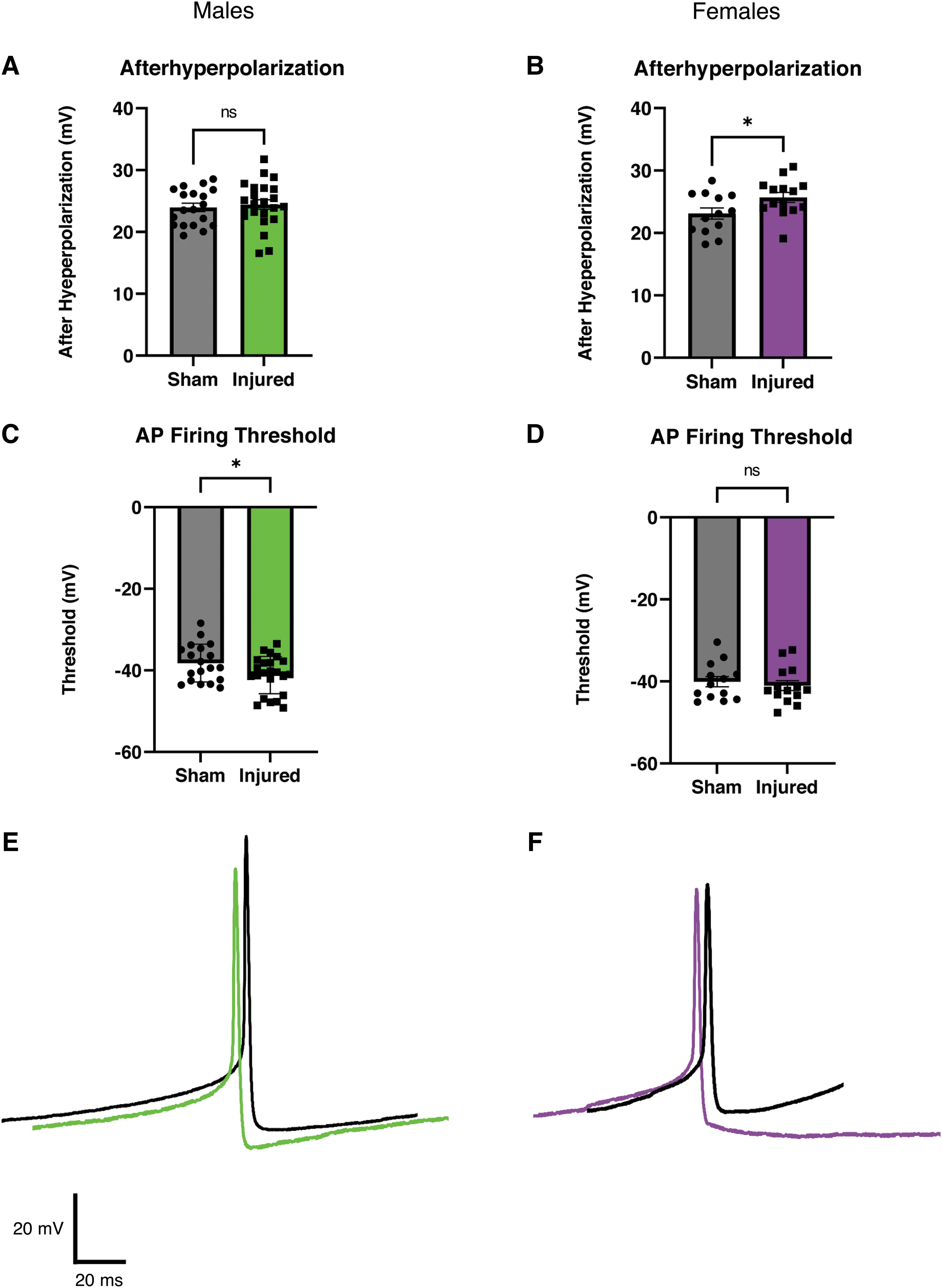
Afterhyperpolarization is increased after injury in female animals but not in male animals. Action potential firing threshold is reduced after injury in male animals, but not in female animals.
Orexin neurons receive local afferents from within the LH 66 as well as from a variety of other brain regions. 62,67,68 Due to the extensive afferent connections onto orexin neurons, we examined whether mTBI alters afferent synaptic activity onto orexin neurons.
Mild traumatic brain injury reduces afferent excitatory activity onto orexin/hypocretin neurons in both male and female mice
Orexin neurons are innervated by both excitatory and inhibitory afferents. 69 Therefore, we recorded EPSCs in orexin neurons in bicuculline (30 μM) to isolate excitatory currents. We recorded both spontaneous excitatory post-synaptic currents (sEPSCs) as well as miniature excitatory post-synaptic currents (mEPSCs) where miniature excitatory events were isolated by recording in bicuculline plus tetrodotoxin (TTX; 0.4 μM).
We examined if injury would affect the properties of EPSCs onto orexin neurons from male animals (Fig. 3). For sEPSCs in slices from male animals, mTBI increased the time between events, or the interevent interval, indicating a decrease in frequency of afferent excitatory activity (Fig. 3C). For mEPSCs in male animals, the amplitude was decreased after mTBI (Fig. 3D). These results match our expected hypothesis that there would be diminished afferent excitatory activity onto orexin neurons after mTBI (sEPSC Sham: n = 7 animals, 16 cells; Injured: n = 7 animals, 18 cells; mEPSC Sham: n = 5 animals, 18 cells; Injured: n = 5 animals, 17 cells).
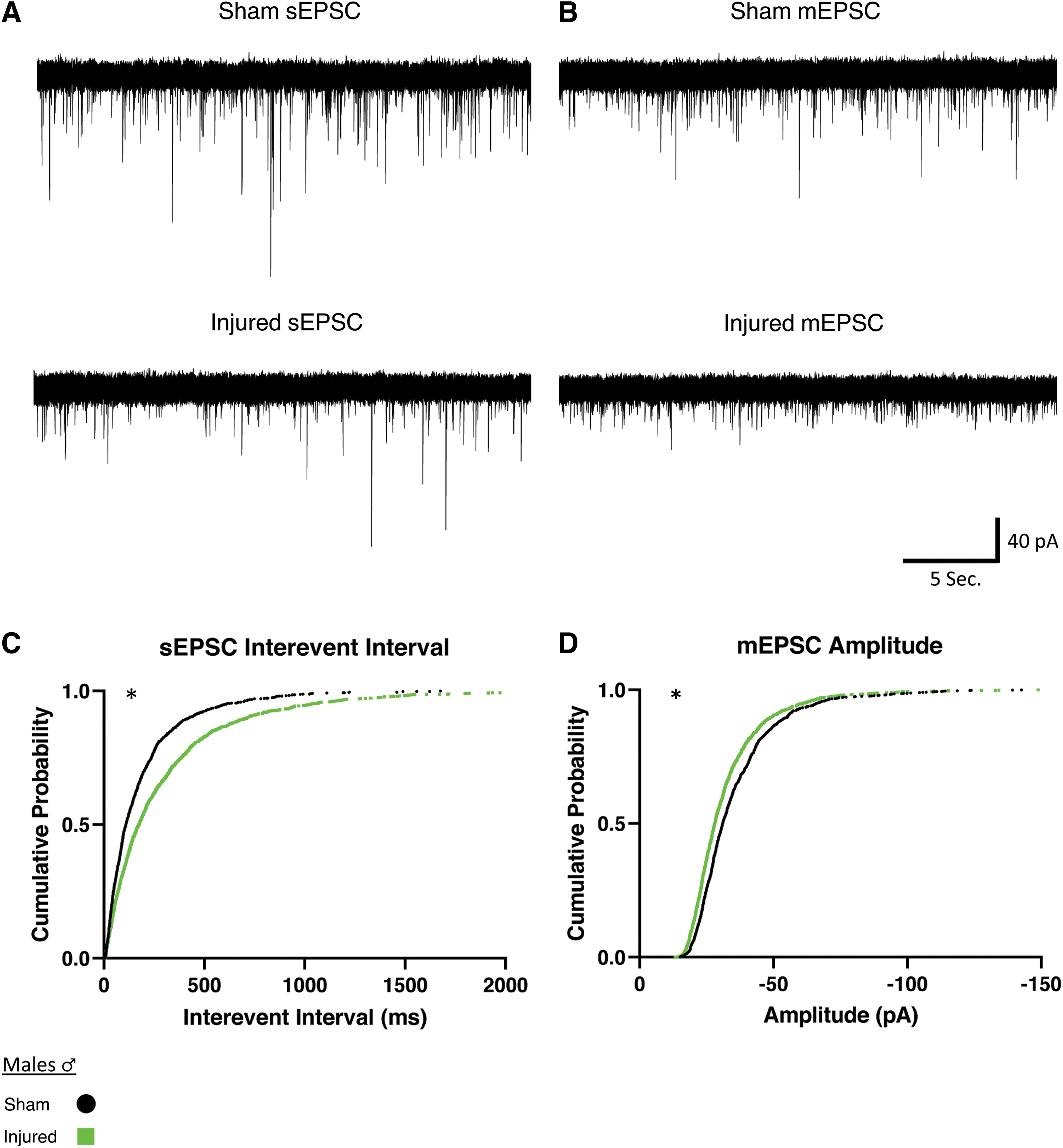
Males show reduced afferent excitatory activity onto orexin/hypocretin neurons after mild traumatic brain injury.
We next examined whether this reduced afferent excitatory activity after injury would hold true for female animals (Fig. 4). We observed in slices from female animals that sEPSCs decreased in amplitude after mTBI (Fig. 4C). Miniature EPSCs in orexin neurons in slices from females had an increase in interevent interval (Fig. 4D), which indicates a decrease in the frequency of miniature afferent excitatory activity. Similar to the male animals, these results indicate a reduction of afferent excitatory signaling after injury (sEPSC Sham: n = 8 animals, 29 cells, Injured: n = 5 animals, 18 cells. mEPSC Sham: n = 8 animals, 32 cells. Injured: n = 4 animals, 17 cells).
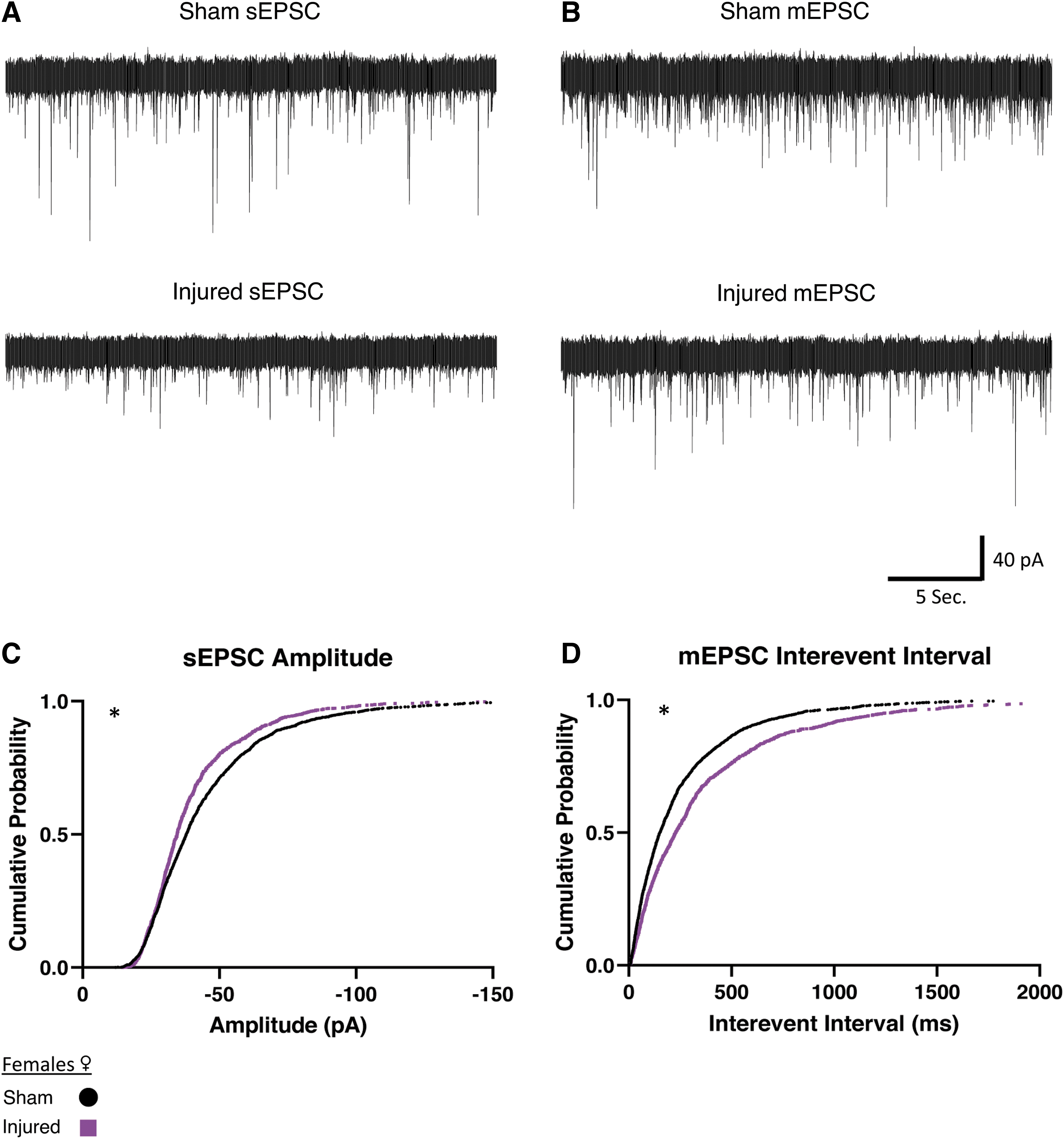
Females show reduced afferent excitatory activity onto orexin/hypocretin neurons after mild traumatic brain injury.
Mild traumatic brain injury increases afferent inhibitory activity onto orexin/hypocretin neurons
Since changes in inhibitory synaptic transmission could also affect orexin neuron activity, and because inhibitory regulation of orexin neurons is crucial for reducing arousal at appropriate times, 70 –74 we next investigated whether there were injury-induced changes in the inhibitory synaptic activity onto orexin neurons. IPSCs in orexin neurons were recorded in the presence of AP5 (50 μM) and CNQX (20 μM) to eliminate excitatory currents from NMDA and AMPA receptors respectively. We recorded both spontaneous (sIPSCs) and miniature currents (mIPSCs) where miniature events were again isolated by recording in the presence of TTX (0.4 μM).
In slices from male animals, injured animals showed a decrease of the interevent interval between sIPSCs, (i.e., an increased frequency of inhibitory activity onto orexin neurons; Fig. 5B). There were no significant differences in mIPSCs in male animals after injury (Table 2; sIPSC Sham: n = 7 animals, 17 cells; Injured: n = 6 animals, 19 cells).
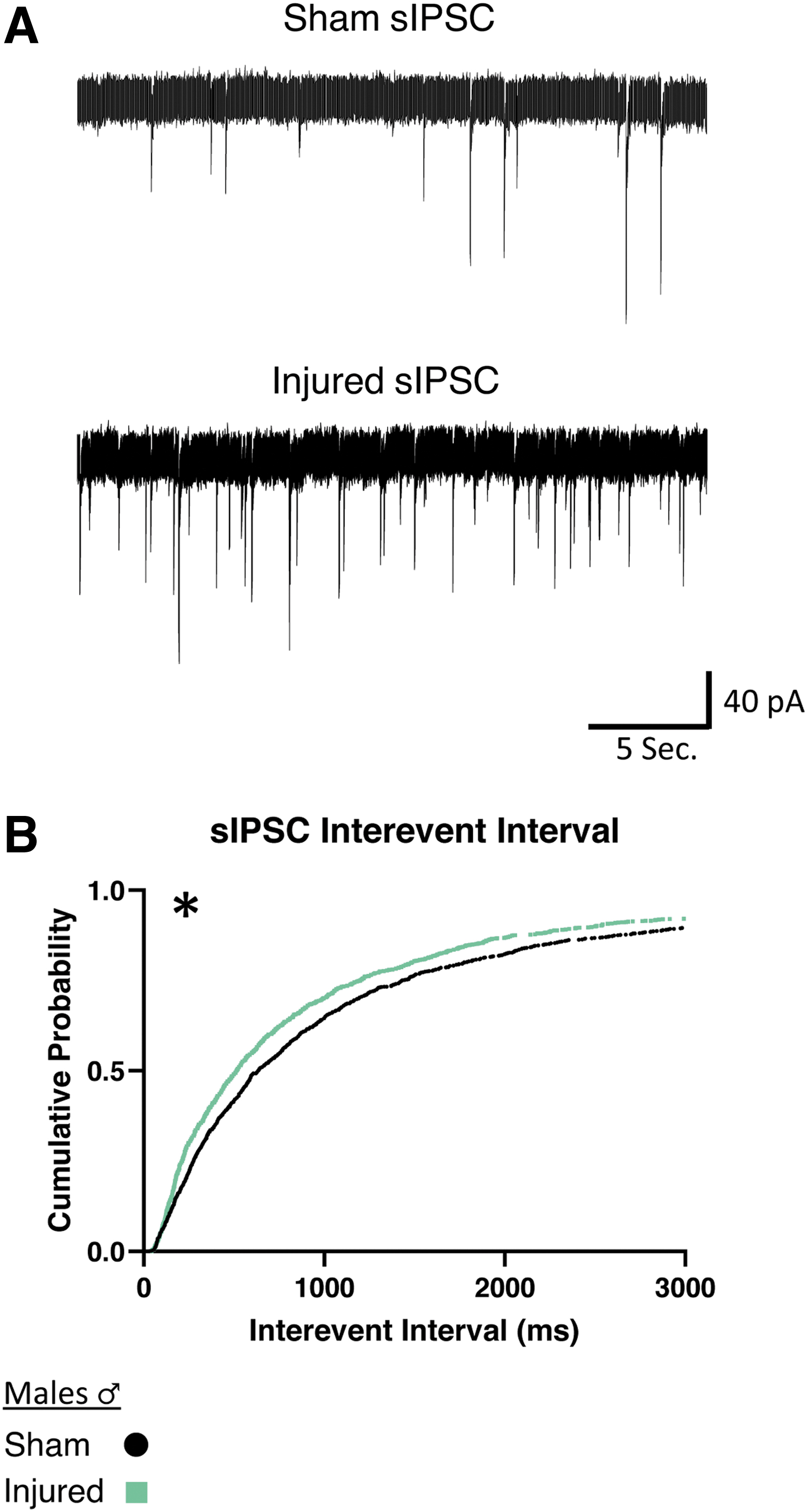
Males show increased frequency of afferent spontaneous inhibitory activity onto orexin/hypocretin neurons after mild traumatic brain injury.
We then wanted to determine if increased afferent inhibitory activity would occur in female animals. In slices from female animals, we also found a reduction of the interevent interval in sIPSCs (Fig. 6E), indicating that injury also increases spontaneous inhibitory activity in female animals. Interestingly, after injury in female animals, both sIPSCs and mIPSCs show an increase in amplitude (Fig. 6C, 6D) that was not present in male animals (Table 2). Taken together, this data suggests that in both males and females there is an increase in afferent inhibitory activity onto orexin neurons after injury. Interestingly, the increase in inhibitory activity is more robust in slices from females (sIPSC Sham: n = 7 animals, 16 cells; Injured: n = 6 animals, 13 cells; mIPSC Sham: n = 5 animals, 13 cells; Injured: n = 6 animals, 14 cells).
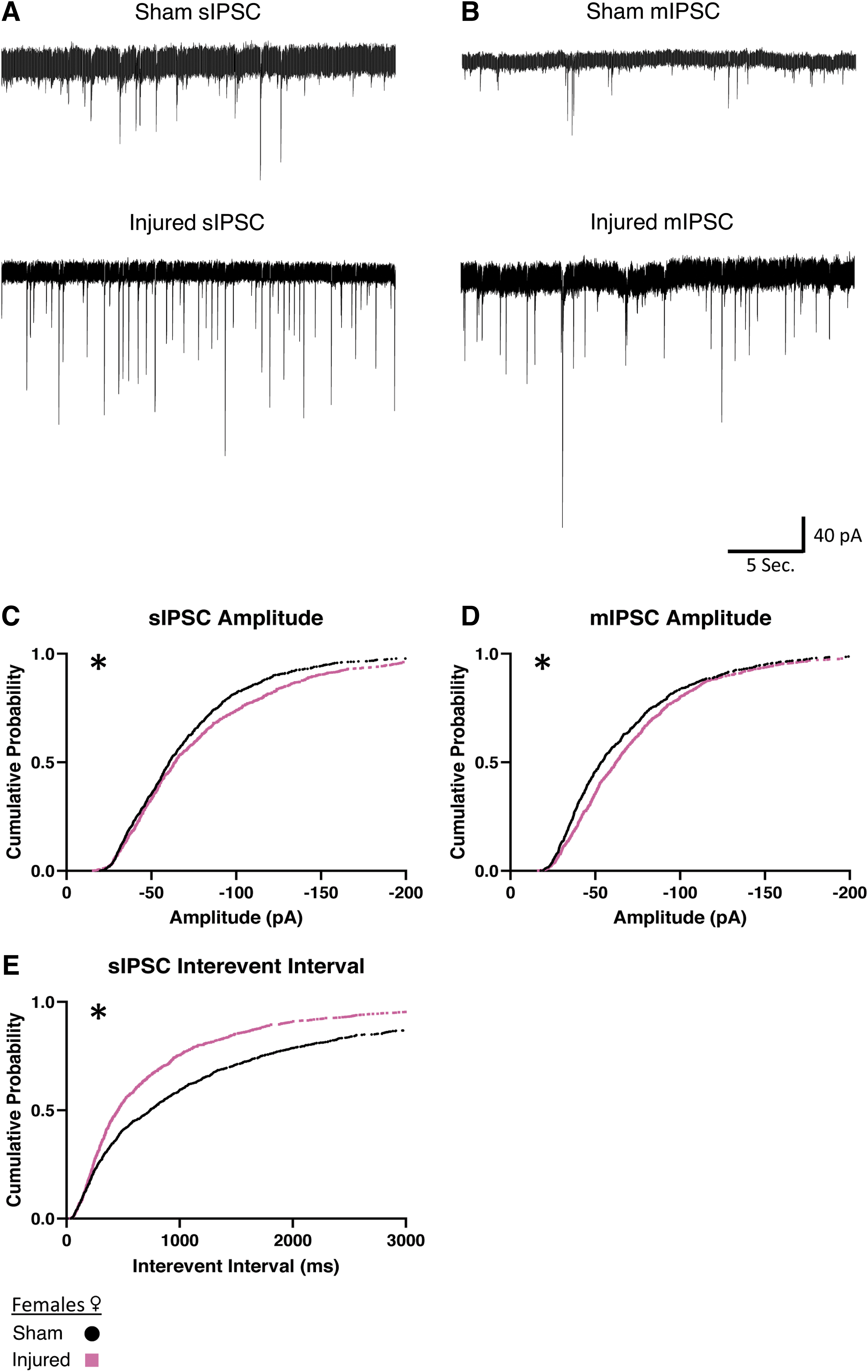
Females show increased afferent inhibitory activity onto orexin/hypocretin neurons after mild traumatic brain injury.
Discussion
While previous work has shown that mild traumatic brain injury affects orexin neurons, how injury affects their activity has not been studied in depth. This work addresses this gap in knowledge by employing whole–cell patch clamp recordings from identified orexin neurons to study the intrinsic properties and afferent synaptic activity onto orexin neurons. We found reduced afferent excitatory activity and increased inhibitory activity onto orexin neurons after injury, suggesting reduced orexin activity. These findings are consistent with others in the field that have shown that there is reduced orexin neuropeptide in CSF after injury, 37,38 as well as studies of orexin neurons that showed reduced orexin neuron activity after injury. 44 We observed an increase of the size of afterhyperpolarization in neurons from female animals and a reduction of action potential threshold in male animals. Injury affecting intrinsic neuronal properties is a known consequence of injury in some brain regions as alterations of action potential threshold have also been observed in neurons in layer 2/3 of the prefrontal cortex after injury. 9 The changes presented in this study provide potential mechanisms for the observations of altered orexin neuron activity after TBI.
We observed that there were different changes in spontaneous and miniature synaptic current events after mTBI. Miniature events are exclusively action-potential independent, while spontaneous events include currents that are action potential dependent. It has been demonstrated that different vesicular pools can contribute to these currents. 75 –82 The injury-induced alterations in current activity may occur in miniature or spontaneous events, which means that mTBI may affect synaptic pools differently. In addition, it is thought that changes in the frequency of currents are related to presynaptic alterations and changes in the amplitude of currents are related to postsynaptic alterations. Together with the model that spontaneous and miniature events contribute to different vesicular pools, we can conceptualize how injury may alter synaptic activity in a mechanistic way (see Fig. 7 for a summary of these ideas).
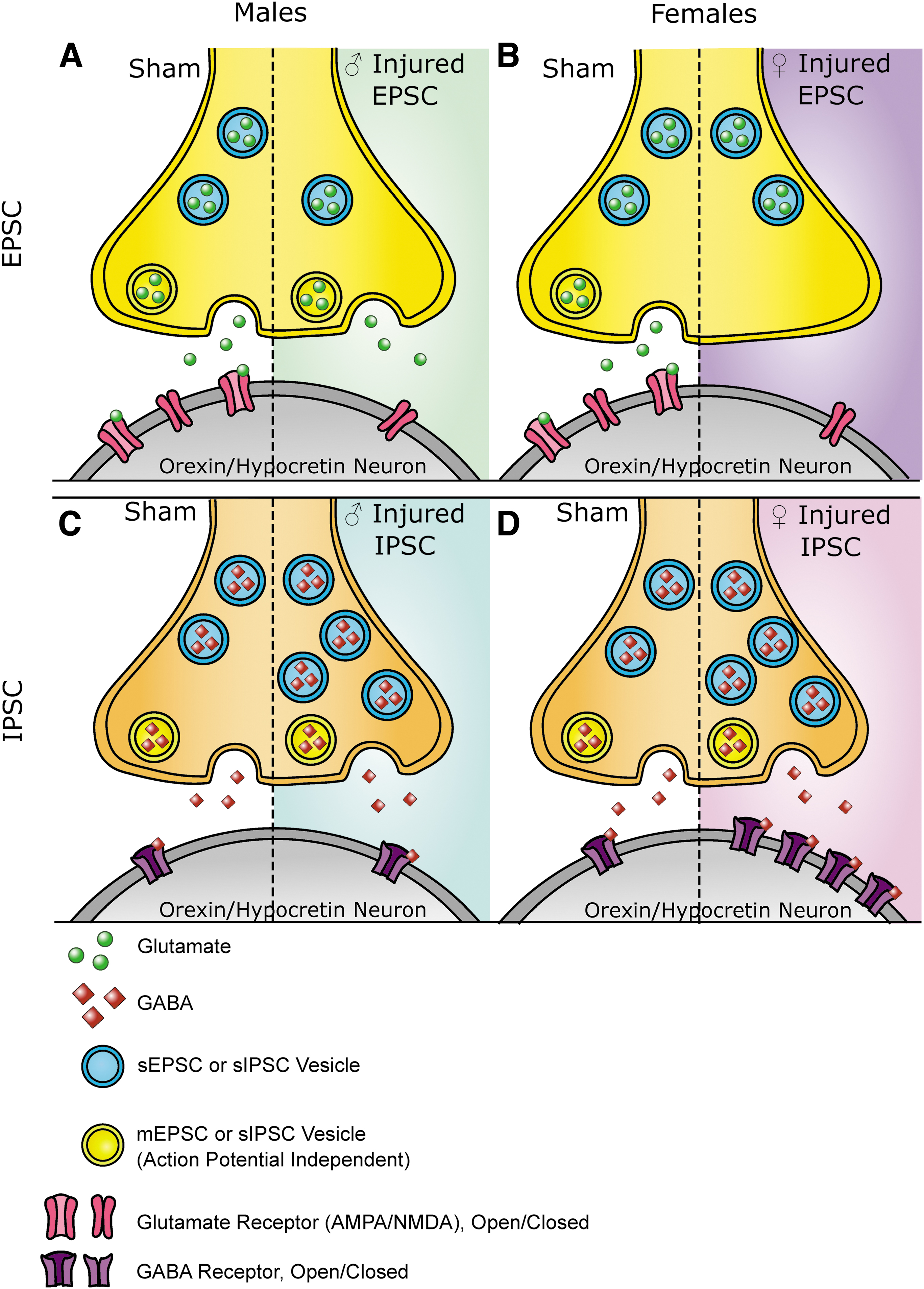
Summary of findings.
Overall, the physiological alterations presented here contribute to a more mechanistic understanding of the changes to orexin circuitry after mTBI. The data presented here may also point to other changes in synapses not summarized in Figure 7, which could include injury induced changes in the number of synapses, changes in the proteins involved in vesicle fusion, altered activity of the receptors or changes in the amount of neurotransmitter within vesicles. Each of these ideas provides interesting and testable avenues for future research.
Based on previous studies, we had originally hypothesized that a majority of observed changes would come from decreased presynaptic glutamate. We did observe reduced afferent glutamatergic activity but we also observed significant increases in afferent inhibitory activity, especially in slices from female animals. The upregulation of GABAergic activity may be a result of compensatory mechanisms after mTBI. After injury, it is known that there is a transient increase in glutamate release 83 and increased inhibition may be an attempt to compensate for this initial transient increase in excitatory activity to restore overall balance. Unfortunately, any potential compensation now appears maladaptive. The reduced afferent excitatory activity and increased inhibitory afferent activity were both measured approximately 1 week after injury. It appears that the increased inhibition is no longer compensating for an initial transient glutamatergic surge. Instead, increased inhibition onto orexin neurons may further reduce their activity following injury and potentially impair their ability to properly integrate signals from across the brain.
An interesting and important outcome of this study is that there are sex differences in how TBI affects orexin physiology after injury. While a majority of the injury induced alterations in the circuitry presented in this work imply reduced orexin neuron activity, the mechanisms are not the same between sexes. The circuitry may be differentially vulnerable to injury in males versus females. Previously published work regarding the effects of TBI on orexin neurons has been conducted only in male animals 38 –41,44,45,52,84 with just one exception, Saber and colleagues, 55 who reported no change in the number of orexin neurons after injury. Due to this paucity of evidence, it has been difficult to speculate further regarding the possibility that injury could affect orexin circuitry differently between sexes. The data presented here will be important for future research and highlights specific alterations in pre- and post-synaptic sites for potential treatment of sleep disorders in men and women after mTBI.
Broadly, the orexin system is not identical between males and females. There is evidence for more prepro-orexin in female animals 85 and higher levels of orexin in females. 86 Another study has reported more orexin neurons in males compared with females, 87 although this study was done in older rats as compared with the work reported from Taheri and colleagues. 86 In either case, males and females measured at similar times and with similar techniques in the same lab show differences between sexes in the orexin neurons in the lateral hypothalamus. This is an indication that the system might be structured differently between sexes at baseline.
The orexin system in males and females also responds differently to stressors. Females have higher levels of prepro-orexin after restraint stress, 88 males and females have different changes in orexin neuron synaptic structure after restraint stress, 59 and female animals show a significantly greater correlation between prepro-orexin messenger RNA (mRNA) expression and corticotrophin releasing hormone, an indicator of stress, compared with males after unpredictable mild stress. 89 Women with major depressive disorder have fewer orexin neurons. 89 Even aging can affect the orexin system differently with one study showing that older female animals have fewer orexin neurons than males. 87 This note about the orexin system differing in the sexes during aging may be particularly important since some research suggests that TBI “ages” the brain. 90 Thus, the stressor of TBI may emphasize sex differences in the orexin system.
In addition to how the orexin system is different in males and females in terms of cells, circuitry, and response to stress, one can also consider whether circulating sex hormones may influence orexin responses to injury since those are also different in males and females. There is evidence that estrogens can influence the orexin system 91 -93 ; therefore, higher levels of estrogen in females may cause injury responses in the orexin system that would be different than males, where estrogen is lower. There is also interest in the female sex hormone cycling and whether that could influence outcomes in female animals. In this current study, there were not enough animals per estrus stage to appropriately compare them (Supplementary Table S1), but comparisons of orexin neurons after injury with a consideration of hormone levels or estrus cycle stage could be a potentially interesting future direction for this work.
As of now, only two published studies have studied electrophysiology to understand mechanistic changes to physiology in the hypothalamus after TBI. 94,95 One of these studies did not find differences in physiology after injury 94 and the other did. 95 In other brain regions, there is evidence that TBI can affect brain regions and cell types differently, even in the same circuit or cells in close proximity. 6 –9 The structure, location, and properties of neurons within a brain region contribute to how TBI affects its physiology. The hypothalamus is a highly heterogeneous structure composed of many nuclei that perform a variety of essential homeostatic functions. 96,97 TBI may very well affect each region of the hypothalamus differently. The work presented in this paper investigating the LH is part of the foundation of exploring how TBI affects the physiology of neural circuits throughout the hypothalamus.
The current study does have several limitations. This study measured the activity of orexin neurons after mTBI, but the complete circuitry of arousal and sleep is more complicated than any one cell subtype. Further investigations of the other connections within the sleep/wake circuitry could provide valuable insights into how the system is affected by injury. Additionally, while whole–cell patch clamp recording can provide detailed information regarding the summed synaptic input onto a neuron, there are still other mechanistic questions that cannot be answered with this technique. One of these questions includes the source of the afferent inputs onto orexin neurons, which could not be determined in this study. An exciting future direction of this work is to determine these potential inputs and isolate which of them are particularly susceptible to mTBI.
Conclusions
In conclusion, orexin circuitry is a critical part of the network in the brain that regulates wakefulness, but there has not been much direct study of how TBI affects this circuitry. As in other brain regions, mTBI does have an effect on the balance between excitation and inhibition in orexin circuitry. This study is among a handful that explore this phenomenon in the hypothalamus. This study is also one of the few that examines orexin neurons after TBI and shows that the physiological differences may appear similar but are not the same between sexes. Future mechanistic studies of how TBI affects neuronal function may reveal important changes in the brain after injury that could be missed with other techniques.
Transparency, Rigor, and Reproducibility Summary
This study and analysis plan were not formally preregistered. Based on extensive experience with patch clamp electrophysiology, sample size was set at a minimum of three animals and 10 cells per group. A total of 186 mice were bred for these experiments, 79 animals were the right age and genotype and had data collected for analysis. Technical animal exclusions included: hemorrhage, herniation, motor deficits, and no delay in righting time for injured animals. The experimenter performing patch clamp experiments was blinded to the animals' injury condition by the technician who performed the injury. The experimenter was blinded to injury condition until data analysis was complete. Animals were heterozygotes for Orexin-EGFP. All animals were 6-8 weeks old at the time of injury, except for one sham female that was 10 weeks old but whose data were not significantly different than other sham females. Animals were sacrificed between 6-10 days after the mild traumatic brain injury. Animals were brought from the animal facility at 9
Footnotes
Acknowledgments
We would like to thank Dr. Robert Greene of UT Southwestern for graciously providing the Orexin-EGFP breeder mice with permission from Dr. Masashi Yanagisawa, creator of the mouse line, and Dr. Theresa Bjorness for helping with their transfer. We would like to thank the Intellectual and Developmental Disabilities (IDDRC) research center at CHOP/Penn and the Robinson lab for the use of the confocal microscope (NIH/NICHD P50HD105354). We would also like to thank Dr. Brian Johnson for thoughtful comments and input.
Authors' Contributions
Rebecca Somach: Conceptualization, data curation, formal analysis, software, investigation, methodology, project administration, resources, visualization, writing-original draft, writing—review and editing.
Ian Jean: Investigation, methodology, resources.
Anthony Farrugia: Investigation, methodology, resources.
Akiva Cohen: Conceptualization, funding acquisition, project administration, supervision, writing—review and editing.
Funding Information
Provided by training grant 5T32HL007953, R01 NS120099, R37HD059288 and the Children's Hospital of Philadelphia Frontier Program.
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.