
Abstract
Many military veterans who experienced blast-related traumatic brain injuries in the conflicts in Iraq and Afghanistan currently suffer from chronic cognitive and mental health problems that include depression and post-traumatic stress disorder (PTSD). Male rats exposed to repetitive low-level blast develop cognitive and PTSD-related behavioral traits that are present for more than 1 year after exposure. We previously reported that a group II metabotropic receptor (mGluR2/3) antagonist reversed blast-induced behavioral traits. In this report, we explored mGluR2/3 expression following blast exposure in male rats. Western blotting revealed that mGluR2 protein (but not mGluR3) was increased in all brain regions studied (anterior cortex, hippocampus, and amygdala) at 43 or 52 weeks after blast exposure but not at 2 weeks or 6 weeks. mGluR2 RNA was elevated at 52 weeks while mGluR3 was not. Immunohistochemical staining revealed no changes in the principally presynaptic localization of mGluR2 by blast exposure. Administering the mGluR2/3 antagonist LY341495 after behavioral traits had emerged rapidly reversed blast-induced effects on novel object recognition and cued fear responses 10 months following blast exposure. These studies support alterations in mGluR2 receptors as a key pathophysiological event following blast exposure and provide further support for group II metabotropic receptors as therapeutic targets in the neurobehavioral effects that follow blast injury.
Introduction
Traumatic brain injury (TBI) happens frequently in both civilian and military life. Public awareness of military-related TBI has increased recently because of the conflicts in Iraq and Afghanistan. 1 While TBI in war zones occurs through various mechanisms, blast-related injuries were the major cause of TBI in Iraq and Afghanistan. 1 While symptoms following particularly mild TBI (mTBI) tend to resolve with time, it is not uncommon that they persist and evolve into a chronic post-concussion syndrome that can last for years. Besides static effects, new symptoms may develop, or existing symptoms may worsen. 2 -4 How much of this worsening is driven by blast-related mechanisms versus mental health related factors including depression and post-traumatic stress disorder (PTSD) remains to be elucidated. 1 Supporting a role for blast-related mechanisms, male rats exposed to repetitive low-level blast exposure develop a variety of PTSD-related behavioral traits that are chronic and persistent being present for more than 1 year after injury. 5 –8
Metabotropic glutamate receptors (mGluRs) play crucial roles in regulating glutamatergic neurotransmission. 9 Agents that modulate mGluR2/3 activity have gained attention as having potential therapeutic roles in a range of mental health related disorders. 10,11 Previously, we treated male rats exposed to repetitive low-level blast with BCI-838 (MGS0210), a group II metabotropic glutamate receptor (mGluR2/3) antagonist prodrug. 12 BCI-838 improved anxiety and fear-related behaviors as well as long-term recognition memory. 12 The ability of an mGluR2/3 antagonist to reverse blast-induced behavioral traits suggested that either blast was disturbing signaling through mGluR2/3 related pathways or that mGluR2/3 antagonism could compensate for other blast-related effects. Further support for a role of mGluR2 in blast-related behavioral effects was obtained in our recent transcriptomic study showing that Grm2 (mGluR2) was among a set of differentially regulated genes whose expression increased in multiple brain regions between 6 weeks and 12 months following blast exposure. 13
Here, we demonstrate that blast exposure induces chronic increased expression of mGluR2 in male rats following blast injury. Extending the evidence that group II metabotropic antagonists may be a promising target for treatment, we also show that another mGluR2/3 antagonist (LY341495) can reverse blast related behavioral traits even if administered after those traits are established. Taken together, these findings point to alterations in mGluR2 expression as a key pathophysiological event following blast injury. They provide further support for mGluR2 signaling as a molecular pathway affected by blast exposure and mGluR2/3 antagonists as potential therapies for PTSD-related symptoms that follow blast injury.
Methods
Animals
Adult male Long-Evans hooded rats (250-350 g; 10 weeks of age; Charles River Laboratories International, Wilmington, MA, USA) were used. All studies involving animals were reviewed and approved by the Institutional Animal Care and Use Committees of the Walter Reed Army Institute of Research (WRAIR)/Naval Medical Research Command (NMRC) and the James J. Peters VA Medical Center (Bronx, NY, USA). Studies were conducted in compliance with the Public Health Service policy on the humane care and use of laboratory animals, the National Institutes of Health Guide for the Care and Use of Laboratory Animals, and all applicable federal regulations governing the protection of animals in research.
Blast overpressure exposure
Rats were exposed to overpressure injury using the WRAIR/NMRC shock tube, which simulates the effects of air blast exposure under experimental conditions. 14 The shock tube has a 12-inch circular diameter and is a 19.5 ft.-long steel tube divided into a 2.5 ft. compression chamber that is separated from a 17 ft. expansion chamber. The compression and expansion chambers are separated by polyethylene Mylar TM sheets (Du Pont, Wilmington, DE, USA) that control the peak pressure generated. The peak pressure at the end of the expansion chamber was determined by piezoresistive gauges specifically designed for pressure-time (impulse) measurements (Model 102M152, PCB, Piezotronics, Depew, NY, USA). This apparatus has been used in multiple prior studies to deliver blast overpressure injury to rats. 5 –8,12,13,15 –22
Individual rats were anesthetized using an isoflurane gas anesthesia system consisting of a vaporizer, gas lines and valves, and an activated charcoal scavenging system adapted for use with rodents. Rats were placed into a polycarbonate induction chamber, which was closed and immediately flushed with 5% isoflurane in air mixture for 2 min. Rats were placed into a cone shaped plastic restraint device and then placed in the shock tube. Movement was further restricted during the blast exposure using 1.5 cm-diameter flattened rubber tourniquet tubing. Three tourniquets were spaced evenly to secure the head region, the upper torso and lower torso while the animal was in the plastic restraint cone. The end of each tubing was threaded through a toggle and run outside of the exposure cage where it was tied to firmly affix the animal and prevent movement during the blast overpressure exposure without restricting breathing.
Rats were randomly assigned to sham or blast conditions and were placed in the shock tube lying prone with the plane representing a line from the tail to the nose of the body in line with the longitudinal axis of the shock tube, with the head placed more upstream. Further details of the physical characteristics of the blast wave are described in Ahlers and colleagues. 14 Blast exposed animals received 74.5 kPa (equivalent to 10.8 psi, duration 4.8 msec, impulse 175.8 kPa*msec) exposures administered one exposure per day for 3 consecutive days (3X blasts). Sham exposed animals were treated identically, including anesthesia administration and placement inside the blast tube, but did not receive a blast exposure (3X sham). Within 10 days after the last blast or sham exposure animals were transported in a climate-controlled van from the WRAIR to the James J. Peters VA Medical Center. Animals left in the morning from the NMRC and arrived in the afternoon of the same day at the James J. Peters VA Medical Center, where all other procedures were performed.
Animal housing
Animals were housed at a constant 70-72°F temperature with rooms on a 12:12 h light cycle with lights on at 7
Regional brain dissection
Animals were sacrificed by CO2 inhalation and the brain was removed. To dissect the various brain regions the cerebellum was removed, and the brain was placed ventral side up and coronal cuts were made through the optic chiasm and anterior commissure. The cortical tissue surrounding this piece of tissue was defined as the anterior cortex. The amygdala was obtained by dissecting the tissue lateral to the hypothalamus between its caudal and rostral border and ventral to the rhinal sulcus on either side. After dissection of the amygdala, the tissue was turned dorsal side up, the cerebral hemispheres were reflected out, and the hippocampus was dissected based on its typical morphology. The posterior cortex included the remainder of the cerebral tissue after the removal of the caudate-putamen. The tissues were flash frozen and stored at -80°C until used.
Western blot analysis
Tissue was homogenized in 0.1 M Tris HCl buffer pH 7.4, containing 0.15 M NaCl, 5 mM ethylenediaminetetraacetic acid (EDTA), 1% Triton X100, 0.1% sodium dodecyl sulfate (SDS) and a protease and phosphatase inhibitor cocktail (Halt, Pierce, Rockford IL, USA), centrifuged at 14,000 rpm, and the supernatant saved and stored at -80°C. Protein concentration was determined with the bicinchoninic acid reagent (ThermoFisher). Protein samples (50 μg) were mixed with loading buffer but not boiled, separated by SDS-polyacrylamide gel electrophoresis (PAGE) and blotted onto polyvinylidene difluoride (PVDF) membranes (Millipore Corporation, Billerica, MA, USA). Blots were blocked with 50 mM Tris HCl, pH 7.6, 0.15 M NaCl, 0.1% Tween-20 (Tris-buffered saline with Tween), 5% nonfat dry milk and probed overnight with the relevant primary antibody diluted in blocking solution. Blots were then incubated for 1.5 h with the appropriate horseradish peroxidase (HRP) conjugated secondary antibody (Cytiva, Piscataway, NJ, USA) diluted in blocking solution (1:7500-1:10000) and the bands were visualized by ECL Prime (Cytiva) and imaged with Imager 600 imaging station or by exposure to HyBlot CL film (Denville Scientific). The primary antibodies utilized are indicated in Table 1. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as loading control. For reprobing, the membranes were stripped with Re-Blot Plus strong stripping solution (Millipore) or ReblotPlus western blot stripping buffer (ThermoFisher) according to the manufacturer's instructions. Quantification was performed using Image Quant TL software (Cytiva).
Primary Antibodies Used for Western Blotting and Immunohistochemistry
WB, Western blot; IHC, immunohistochemistry; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
RNA isolation and quantitative polymerase chain reaction
RNA was isolated using the Qiazol reagent (Qiagen, Germantown. MD, USA) and further purified by RNeasy mini kit according to the manufacturer's instructions (Qiagen). RNA quality was assessed using an RNA2100 Agilent bioanalyzer. Reverse transcription was performed with the High-capacity cDNA reverse transcription kit (ThermoFisher) using 1 μg of RNA. Quantitative polymerase chain reaction (qPCR) was performed using TaqMan gene expression assays and TaqMan universal master mix (ThermoFisher). The following assays were used: Rn01447672_m1 (mGluR2), Rn01755349_m1 (mGluR3), Rn01775763 (GAPDH). Expression levels were normalized to GAPDH and expression changes relative to control samples calculated using the 2-ΔΔCt method.
Immunohistochemistry of rat brain
Rats were perfused with 4% paraformaldehyde in phosphate-buffered saline, and the brains dissected and post-fixed overnight in 4% paraformaldehyde. The brains were sectioned into 50 μ-thick coronal sections with a Vibratome (Leica, Wetzlar, Germany). Immunohistochemical staining was performed as previously described 21 using the antibodies listed in Table 1. Stained sections were imaged with a Zeiss 700 or LSM980 confocal microscope (Zeiss, Thornwood, NY, USA) or a Keyence BZ-X810 fluorescence microscope (Keyence Corporation, Itasca, IL, USA). Images were processed with Adobe Photoshop (CC 20.0.6, Photoshop 23, Adobe Systems, San Jose, CA, USA).
Drug administration
LY341495 (2S-2-amino-2-[1S,2S-2-carboxycycloprop-1-yl]-3-[xant-9-yl] propanoic acid; Bio- Techne Corporation/Tocris Biosciences, Minneapolis MN, USA) was dissolved in saline. Rats were treated with doses of 3 mg/kg administered intraperitoneally. Animals were divided into three experimental groups: 1) sham exposed rats treated with vehicle (sham + veh; n = 13); 2) blast-exposed rats treated with vehicle (blast + veh; n = 6); and 3) blast-exposed rats treated with LY341495 (blast + LY341495; n = 6). At 10 months following blast exposure, we administered 3 mg/kg IP on 2 consecutive days before initial novel object recognition (NOR) training. Recognition memory was tested 24 h later. The following week rats were administered an additional dose of 3 mg/kg LY341495 or vehicle on Day 1. On Day 2, fear training was conducted and on Day 3 rats were tested for cued fear learning.
Novel object recognition
Rats were habituated to the arena (90 cm length × 60 cm width × 40 cm height) for 20 min, 24 h before training. On the training day, two identical objects were placed on opposite ends of the empty arena, and the rat was allowed to explore the objects freely for 7 min. After a 24-h delay during which the rat was held in its home cage, one of the two familiar objects (FOs) was replaced with a novel one (NO), and the rat was allowed to freely explore the FO and NO for 5 min to assess recognition memory. Raw exploration times for each object were expressed in seconds. Object exploration was defined as sniffing or touching the object with the vibrissae or when the animal's head was oriented toward the object with the nose placed at a distance of less than 2 cm from the object.
All sessions were recorded by video camera (Sentech, Carrollton TX, USA) and analyzed with ANYMAZE software (San Diego Instruments). In addition, offline analysis by an investigator blind to the blast-exposed status of the animals was performed. Objects to be discriminated were of different size, shape and color and were made of plastic or metal material. The objects consisted of a 330-mL soda can, a metal box, a cup, and a plastic tube. All objects were cleaned with 70% ethanol between trials.
Cued fear conditioning
Sound-attenuated isolation cubicles (Coulbourn Instruments, Holliston, MA, USA) were utilized. Each cubicle was equipped with a grid floor for delivery of the unconditioned stimulus (US) and overhead cameras. All aspects of the test were controlled and monitored by the Freeze Frame conditioning and video tracking system (Actimetrics, Coulbourn Instruments). During training the chambers were scented with almond extract, lined with white paper towels, had background noise generated by a small fan and were cleaned before and between trials with 70% ethanol. Each subject was placed inside the conditioning chamber for 2 min before the onset of a conditioned stimulus (CS; an 80 dB, 2 kHz tone), which lasted for 20 sec with a co-terminating 2-sec footshock (0.7 mA; US). A total of three tone/shock pairings were administered with the first/second and second/third pairings separated by 1 min. Each rat remained in the chamber for an additional 40 sec following the third CS-US pairing before being returned to its home cage. Freezing was defined as a lack of movement (except for respiration) in each 10-sec interval. Minutes 0-2 during the training session were used to measure baseline freezing.
Animals were returned to their home cage for 24 h at which time cued conditioning was tested. To create a new context with different properties, the chambers were free of background noise (fan turned off), lined with blue paper towels, scented with lemon extract, and cleaned before and during all trials with isopropanol. Each subject was placed in this novel context for 2 min and baseline freezing was measured, followed by exposure to the CS (20-sec tone) at 120, 240 and 360 sec.
Statistical analysis
Values are expressed as mean ± standard error of the mean (SEM). Comparisons were performed using one-way analysis of variance (ANOVA), repeated-measures ANOVA, or two-tailed unpaired t-tests where appropriate. When repeated-measures ANOVA was used, statistical significance was determined using the Greenhouse-Geisser correction. 23 Due to the three-group design, post hoc tests following a significant ANOVA were performed using Fisher's least significant difference (LSD). 24 For all tests, statistical significance was set at a level of 0.05. Statistical tests were performed using the program GraphPad Prism 9.4.1 (GraphPad Software) or SPSS v29 (IBM).
Results
mGluR2 expression increases in a delayed manner following blast injury in rats
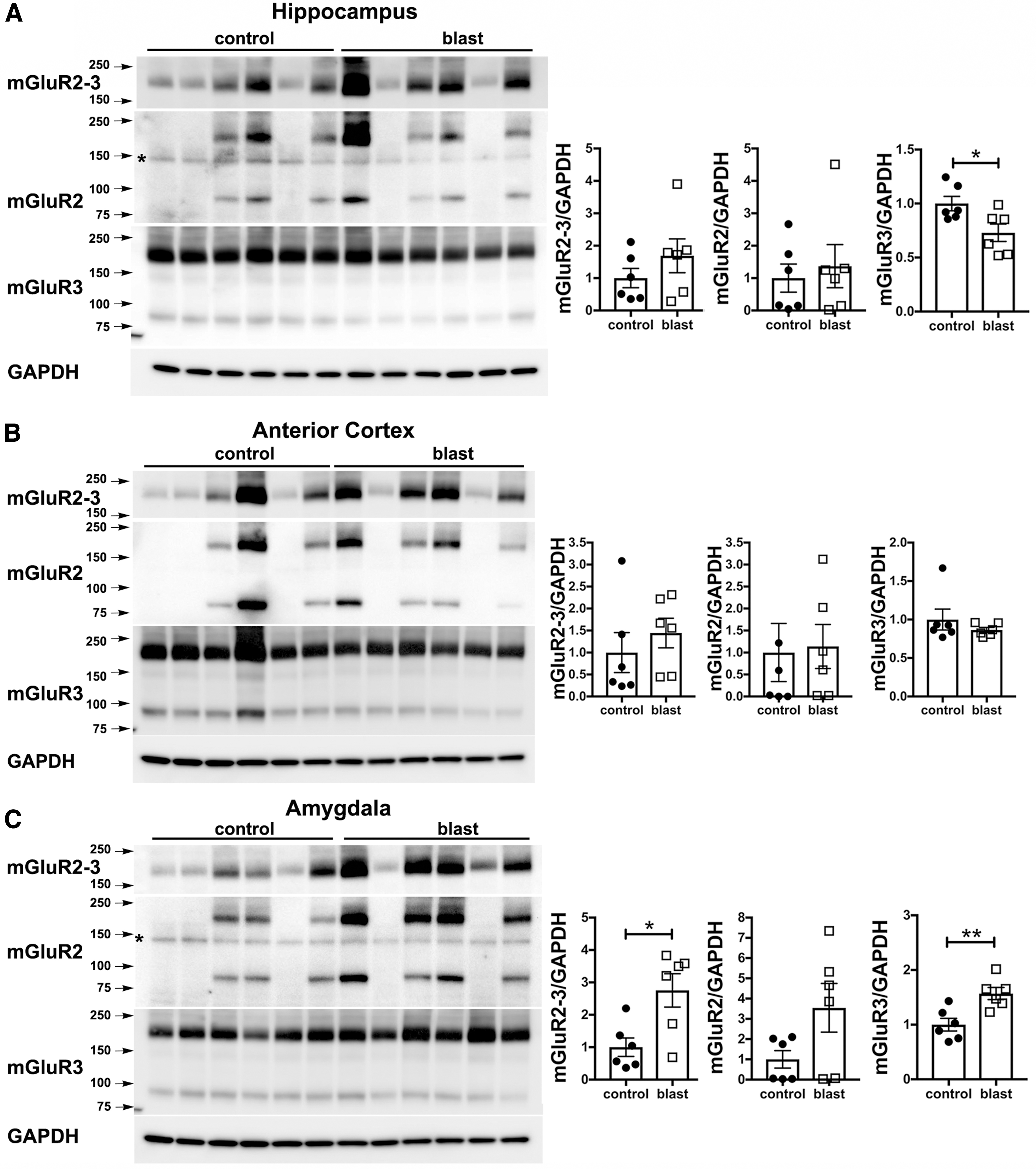
Because treatment with an mGluR2/3 receptor antagonist reversed many of the PTSD-related behavioral traits in blast-exposed male rats, 12 we examined mGluR2/3 expression by Western blotting after blast exposure. The study design and time-points for tissue collection are shown in Figure 1. We initially examined expression at 43 weeks following three blast exposures using shock wave parameters consistent with mTBI or low-level military occupational exposure. Samples were studied from a cohort that had previously been behaviorally tested and exhibited the chronic behavioral phenotype. 6 mGluR2/3 levels were increased in blast-exposed rats in all three brain regions examined (Fig. 2). Increases were confirmed in a second cohort of rats examined at 52 weeks following exposure (Supplementary Fig. S1).

Experimental design.

Metabotropic glutamate receptor (mGluR)2/3 expression at 43 weeks following blast injury. mGluR2/3 expression was analyzed by Western blotting in hippocampus
Since the antibody initially used to generate these data recognizes both mGluR2 and mGluR3, to determine which isoform was responsible for increased mGluR2/3 levels, we used mGluR2 or mGluR3 specific antibodies. This analysis showed that the increase in mGluR2/3 expression was driven by elevated mGluR2 (Fig. 2 and Supplementary Fig. S1). There were no changes in mGluR3 levels at 43 weeks following blast (Fig. 1), while at 52 weeks the only change in mGluR3 was a decrease in hippocampus, but not anterior cortex or amygdala (Supplementary Fig. S1).
In keeping with the protein data, quantitation of mRNA levels by qPCR in the rats studied at 52 weeks following blast exposure found elevated mGluR2 RNA in all three regions examined (Fig. 3A-C). mGluR3 RNA was modestly decreased in amygdala (Fig. 3F), a change not reflected in protein levels (Supplementary Fig. S1C). mGluR3 mRNA was unchanged in hippocampus and anterior cortex (Fig. 3D, 3E), which suggested that decreases in mGluR3 at the protein level in hippocampus could be post-translational.
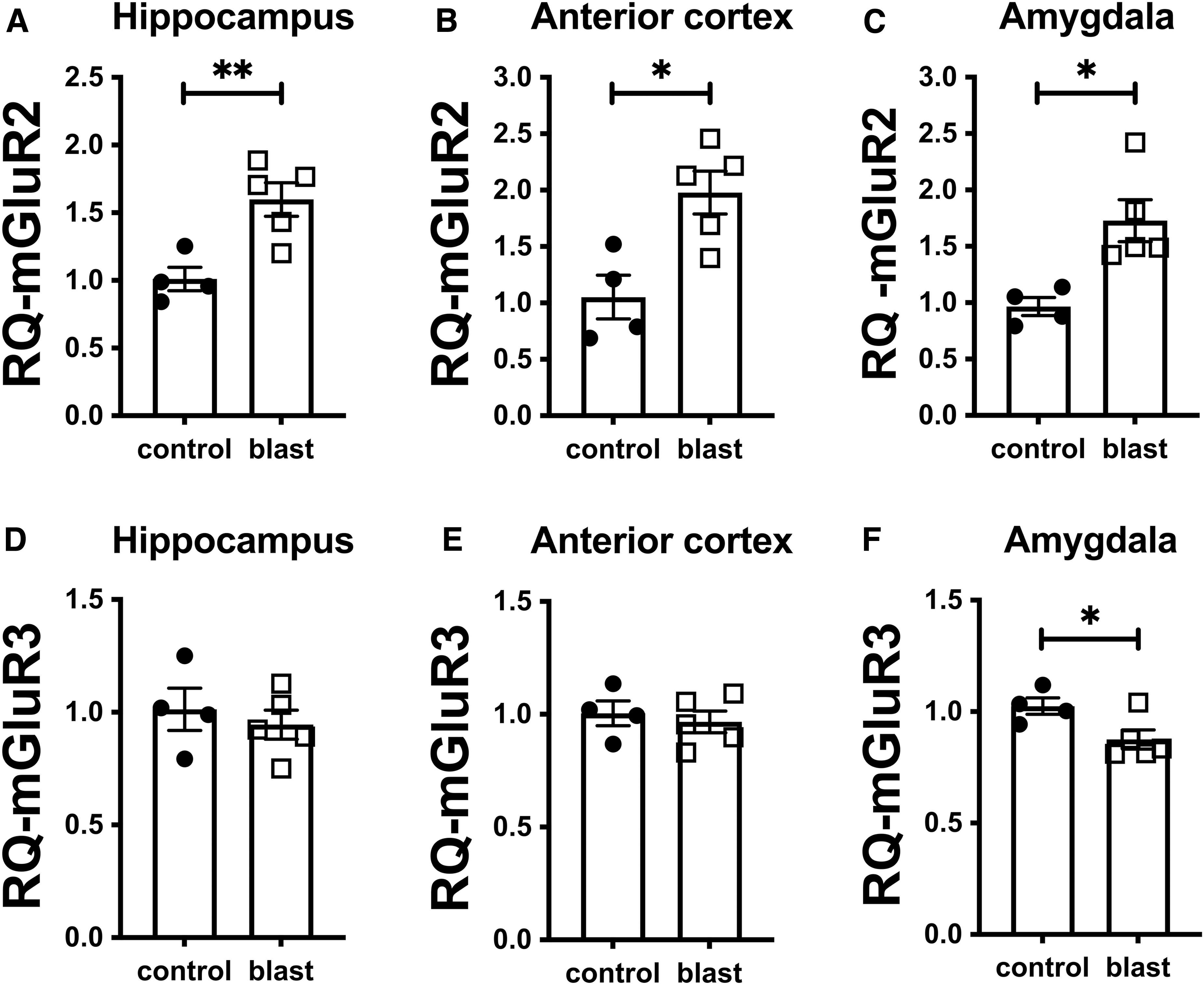
Levels of metabotropic glutamate receptor (mGluR)2 and mGluR3 RNA at 52 weeks after blast exposure. Quantitative polymerase chain reaction for mGluR2
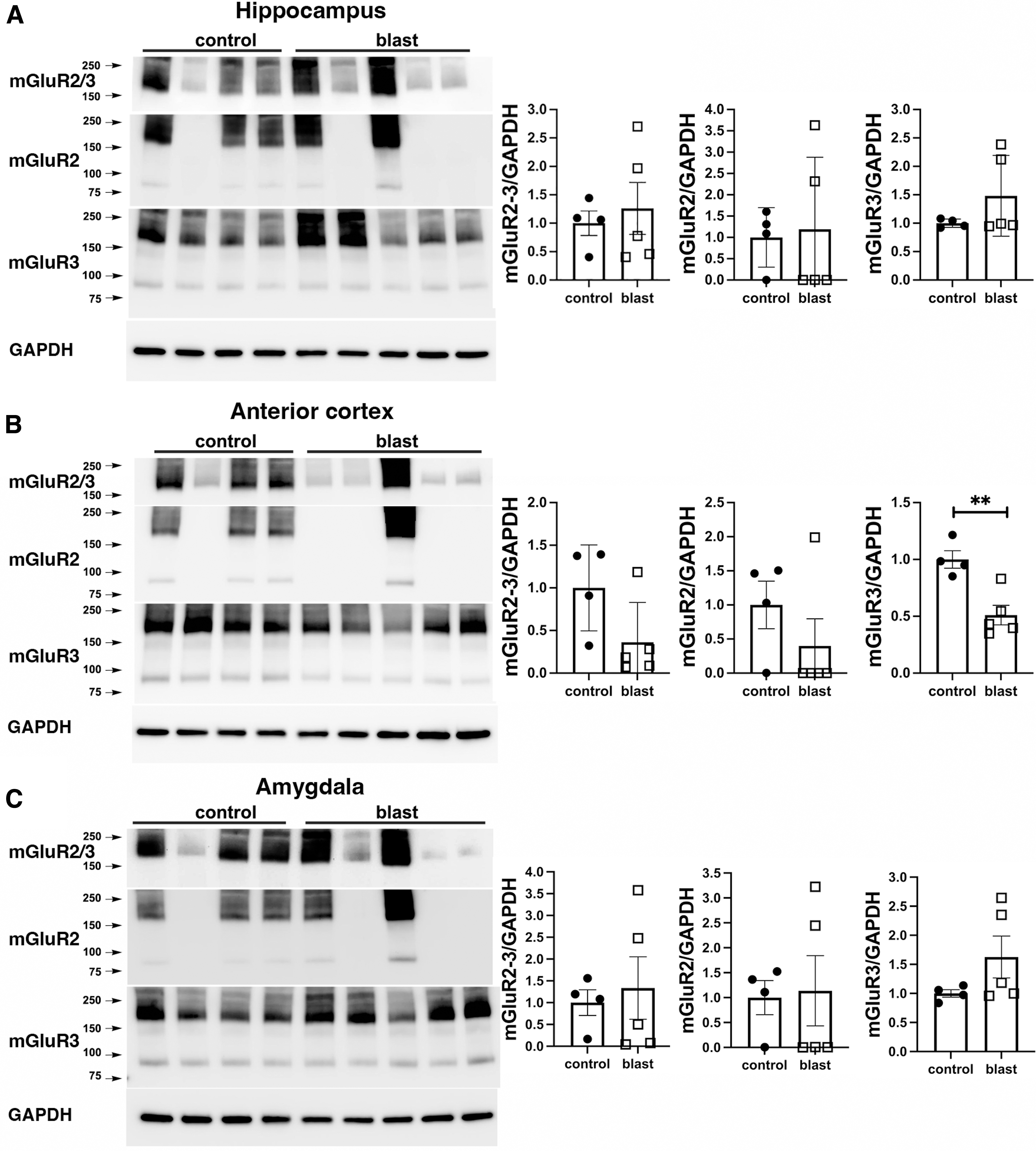
To examine the time course of mGluR2/3 changes we performed Western blots on rats harvested at 2 weeks or 6 weeks after blast exposure. At both time-points, sample to sample variability, particularly in mGluR2 expression was larger than at later time-points. Two weeks following blast exposure, mGluR2/3 expression was increased in amygdala (Fig. 4C) while there were no changes in mGluR2/3 levels in hippocampus or anterior cortex (Fig. 4A, 4B). When analyzed with specific antibodies, mGluR2 levels were not changed in any region while mGluR3 was decreased in hippocampus (Fig. 4A) and increased in amygdala (Fig. 4C). At 6 weeks, mGluR3 was decreased in anterior cortex (Fig. 5B). However, there were no differences in mGluR3 in other regions and there were no statistically significant differences in mGluR2/3 or mGluR2 in any region (Fig. 5).
Metabotropic glutamate receptor (mGluR)2/3 expression at 2 weeks following blast injury. mGluR2/3 expression was analyzed by Western blotting in hippocampus
Metabotropic glutamate receptor (mGluR)2/3 expression at 6 weeks following blast injury. mGluR2/3 expression was analyzed by Western blotting in hippocampus
Immunostaining shows wide-spread mostly presynaptic mGluR2 expression in blast exposed and control samples
To examine whether the pattern of mGluR2 expression was altered following blast exposure, we examined by immunohistochemistry tissue sections from blast-exposed and control rats using the same antibodies as used for Western blotting (Table 1). Figure 6 shows immunohistochemical staining of hippocampus for mGluR2 at 43 weeks following blast exposure and Figure 7 shows staining of the amygdala and frontal neocortex at 52 weeks. Except for an apparent increase in expression, there was no change in the overall pattern of mGluR2 expression.
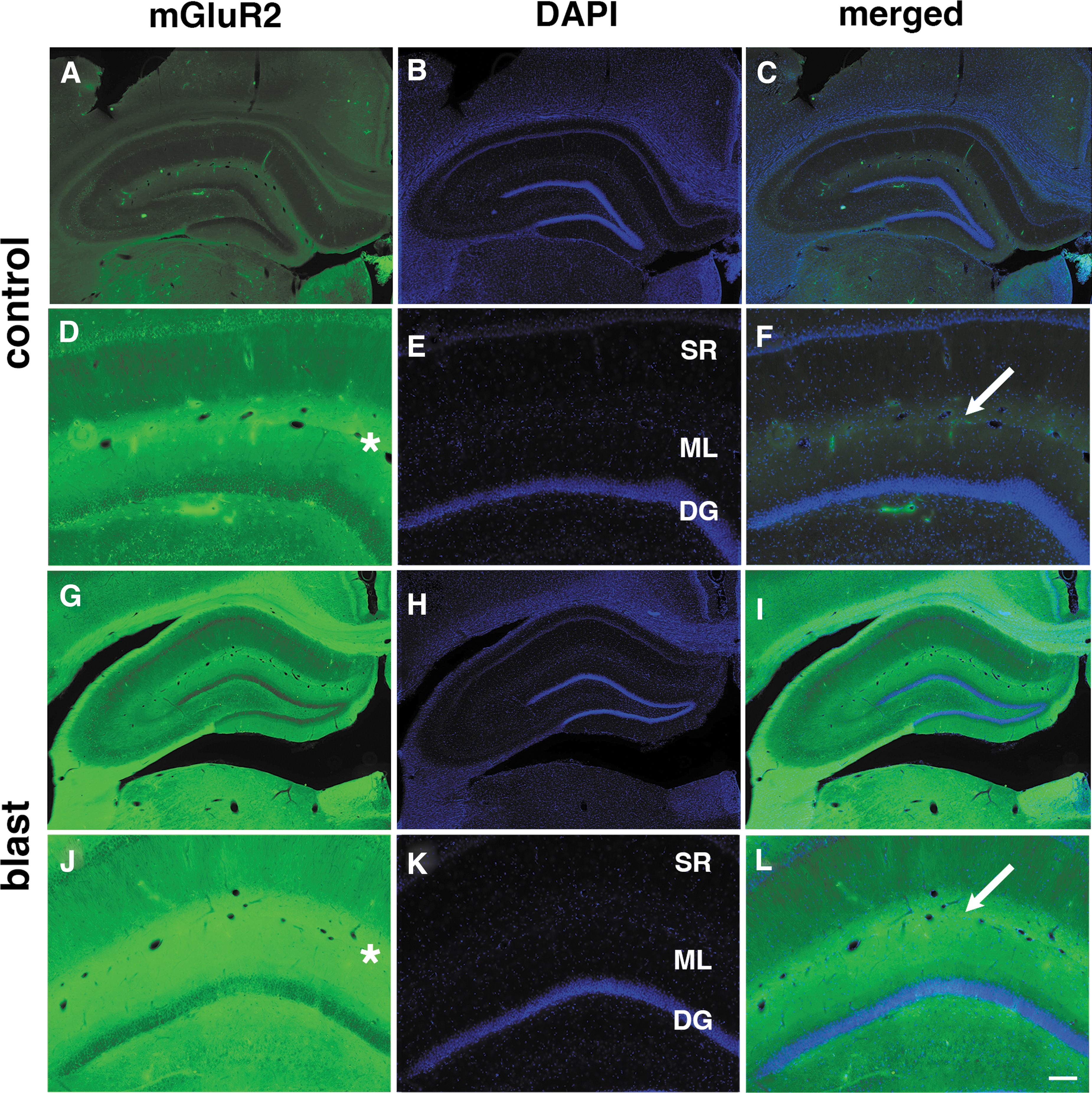
Immunostaining of metabotropic glutamate receptor (mGluR)2 in the rat hippocampus at 43 weeks following blast exposure. Shown are sections of hippocampus from control

Immunostaining of metabotropic glutamate receptor (mGluR)2 in rat amygdala and neocortex at 52 weeks following blast exposure. Shown are sections of amygdala
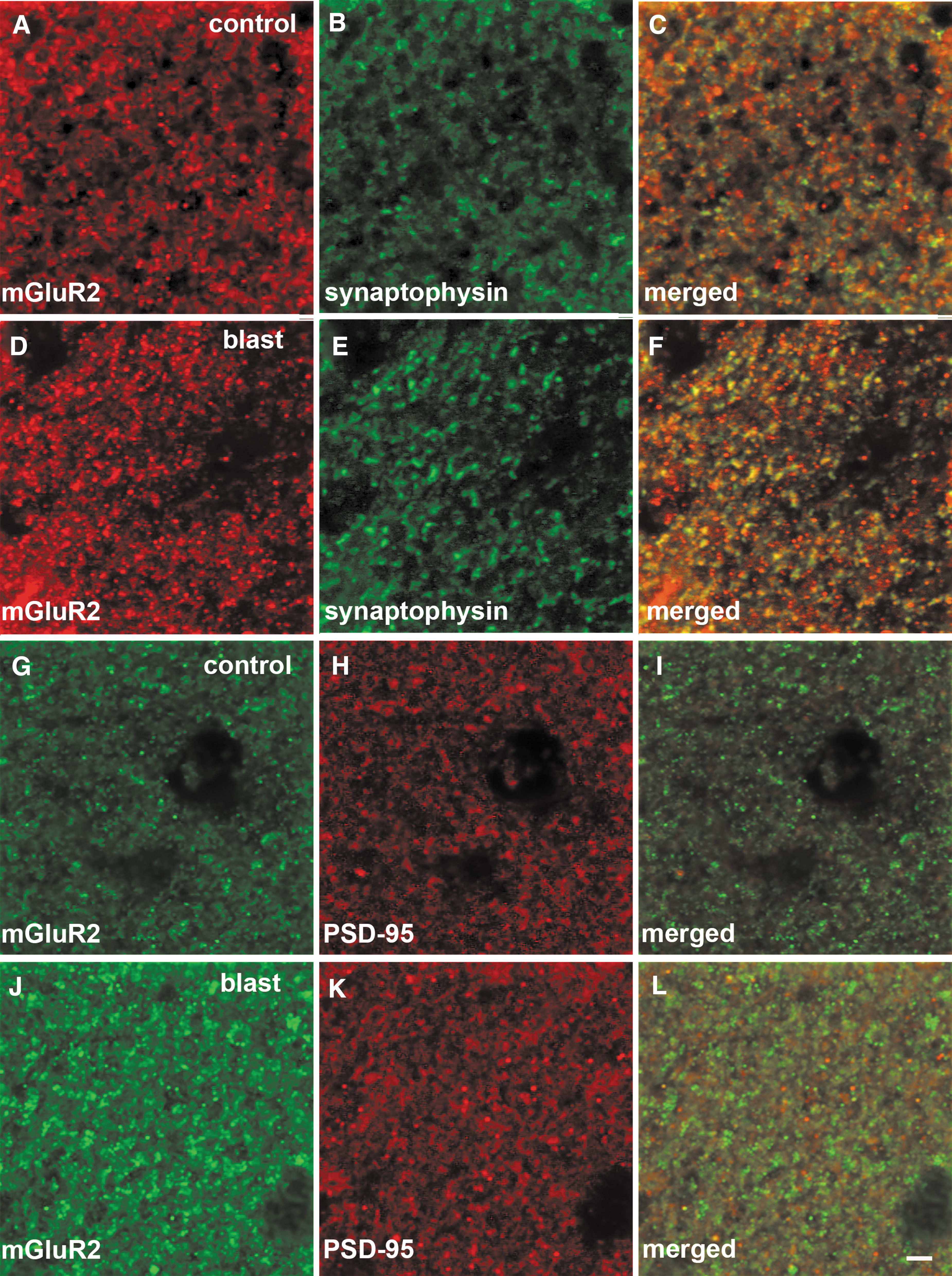
Immunostaining and distribution of metabotropic glutamate receptor (mGluR)2 in relationship to synaptophysin and PSD-95 in rat hippocampus at 43 weeks following blast exposure. Shown are sections of hippocampus from control
mGluR2 receptors are synaptic proteins primarily expressed in perisynaptic locations on the presynaptic side 11 although expression on postsynaptic membranes has been reported. 25 To determine whether the principal subcellular localization of mGluR2 was disturbed by blast, we performed immunostaining combining antibodies against mGluR2 with those against the presynaptic protein synaptophysin or the postsynaptic protein PSD-95. Figure 8 illustrates staining of the stratum lacunosum moleculare of the rat hippocampus at 43 weeks following blast exposure. The stained region corresponds to the area marked with asterisks in Figure 6. Substantial overlap was seen between mGluR2 and synaptophysin in the merged images of Figure 8 (yellow/orange in panels C and F). By contrast, mGluR2 and PSD-95 staining was largely non-overlapping suggesting the primarily presynaptic localization of mGluR2. The general pattern of staining appeared comparable in control and blast-exposed animals, thus suggesting that localization of the receptor was not influenced by blast exposure.
The mGluR2/3 antagonist LY341495 rapidly reverses blast-induced effects on behavior
We previously reported that BCI-838 an orally administered mGluR2/3 antagonist prodrug could prevent development of PTSD-related behavioral traits if given for 8 weeks starting 2 weeks after blast exposure, 12 that is, beginning treatment before the behavioral phenotype emerges. 8 Here, we tested whether the parenteral mGluR2/3 antagonist LY341495, could reverse PTSD-related behavioral traits if administered after the behavioral phenotype was established.
We tested LY341495 in blast-exposed rats at 10 months following exposure. The design of the study is shown in Figure 9. A dose of 3 mg/kg LY341495 or vehicle was administered IP on two consecutive days before NOR training. Recognition memory was tested 24 h later. Animals were divided into three experimental groups: 1) control rats treated with vehicle (control + vehicle); 2) blast-exposed rats treated with vehicle (blast + vehicle); and 3) blast-exposed rats treated with LY341495 (blast + LY341495). Comparison of sham + vehicle-treated controls with blast + vehicle treated animals provided a positive control for development of the blast-related behavioral phenotype. Comparing blast + saline with blast + drug allowed effects of LY341495 on the blast-related phenotype to be determined.
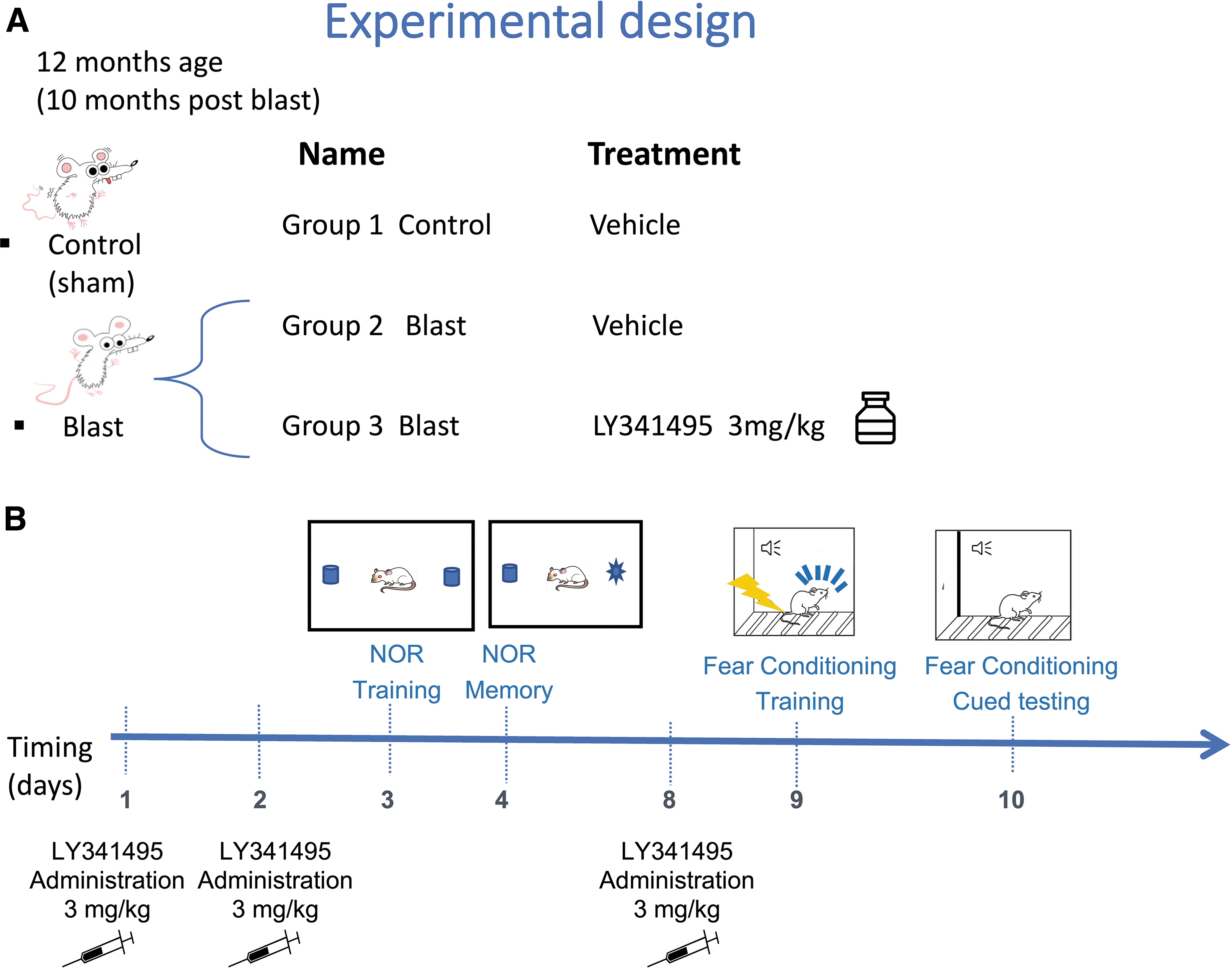
Experimental design for LY341495 treatment of blast-exposed rats. At 10 months after blast exposure, control or blast-exposed rats were treated with vehicle (n = 13 control; n = 6 blast) and blast-exposed rats were treated with 3 mg/kg LY341495 (n = 6) for two days. Panel
During NOR training (Fig. 10A), all groups explored the two objects equally. However, blast-exposed rats treated with LY341495 spent more total time exploring the objects than either control or blast-exposed rats treated with vehicle (Fig. 10B). Twenty-four hours later subjects were given the choice of exploring a novel object (NO) or one of the previously presented familiar objects (FO). Figure 10C shows a discrimination index (DI) calculated for the 24 h testing. Control + saline explored the NO more than FO and exhibited an increased DI compared with blast + saline who explored the NO and FO roughly equally. However, blast + LY341495-treated rats explored the NO more than the FO and did not differ in the calculated DI from control + saline indicating that deficits in NOR in blast-exposed rats were rescued by LY341495.

Effect of LY341495 treatment on behavior of blast-exposed rats. Panel (
The following week, the same rats examined in NOR testing (Fig. 10A-C) were administered an additional dose of 3 mg/kg LY341495 or vehicle (Day 8 in Fig. 9B). The following day (Day 9) fear training was conducted and on Day 10 rats were tested for cued fear learning. Blast-exposed and control groups responded similarly during training with increased freezing following pairing of the tone with the shock (Fig. 10D) but without between group differences. In the cued phase when the tone was presented in a novel environment without a foot shock (Fig. 10E), blast-exposed rats treated with saline responded with increased freezing compared with saline-treated controls. However, in blast-exposed rats LY341495 restored responses to those of control + saline (Fig. 10E). Taken in combination with our previous studies, 26 these results support the relevance of mGluR2 elevation to development of the behavioral phenotype.
Discussion
A history of TBI is frequently found in veterans seeking mental health treatment in U.S. Department of Veterans Affairs medical centers. 27 Various mental health problems have been linked to TBI including anxiety, depression, impulsivity, and suicidality. 1,28,29 One striking feature in the recent Iraq and Afghanistan veterans has been the co-existence of TBI and PTSD. 1
In this study we used a well-established animal model, which employs male rats and mimics the type of open field low-level blast exposure associated with human mild TBI or subclinical blast exposure in humans. 5 –8,12,13,15 –22 Male rats were subjected to 74.5 kPa exposures (equivalent to 10.8 psi), which represent a level of blast that is transmitted to brain 30 but which does not produce major gross neuropathological nor systemic effects. 14
Blast exposures were delivered at 10 weeks of age, with rats lying prone in the shock tube with the plane representing a line from the tail to the nose of the body in line with the long axis of the shock tube and the head upstream. Head motion was restricted during exposure to minimize damage from rotational/acceleration injury. 31,32 Lack of evidence for gross coup/contrecoup injuries supports the relatively mild nature of the brain injury. 5,14 To mimic the multiple blast exposures commonly experienced by veterans in Iraq and Afghanistan, 33 rats were subjected to three 74.5 kPa exposures delivered once a day for 3 consecutive days.
Male rats subjected to repetitive low-level blast exposure as described above show cognitive and PTSD-related behavioral traits including anxiety, enhanced acoustic startle, and impaired recognition memory. 5 –8 These traits develop in a delayed manner being absent in the first eight weeks after blast exposure but consistently present 3-4 months and longer after exposure. Once established, these traits remain present for more than 1 year after blast exposure 5 -7,12,34 likely being present for the lifetime of the animal. These animals thus model the chronic neurobehavioral syndromes that veterans often suffer following blast exposure. 2,3,35
Herein, we show that mGluR2 expression is chronically increased long after blast exposure. Initially we examined time-points in the period when the behavioral phenotype is well established. By Western blotting, mGluR2 expression was increased in all brain regions examined at 43 or 52 weeks following blast injury. Elevation was further supported by mGluR2 RNA increases in animals studied at 52 weeks. Next, we examined time-points before the behavioral phenotype emerges which occurs between 8-12 weeks. 8 mGluR2 levels were not increased at 2 weeks or 6 weeks after blast exposure.
Further support for mGluR2 elevation being a delayed consequence of repetitive blast injury is provided by our recent transcriptomic studies using this model showing that Grm2 (mGluR2) was one of a set of differentially regulated genes whose expression increased in amygdala, hippocampus, and anterior cortex between 6 weeks and 12 months following blast exposure. 13 Now knowing that mGluR2 elevation occurs over a similar time frame as the behavioral phenotype, it will be important in future studies to examine intermediate time-points between 6 and 43 weeks, in particular that critical 8-12 week time period when the behavioral phenotype emerges.
Support for a functional relevance of mGluR2 elevation to the behavioral phenotype can be found in our previous studies showing that the mGluR2/3 antagonist BCI-838 ameliorated anxiety and fear-related behaviors as well as long-term recognition memory if treatment was initiated before the behavioral phenotype emerged. 8,12 Herein, we show that the mGluR2/3 antagonist LY341495 rapidly reversed deficits in NOR and fear learning even if administered after blast-induced behavioral traits had emerged months after mild injury.
LY341495 dosing for this study was arbitrarily chosen but based on previous work showing that one or a few doses of LY341495 exerts sustained behavioral effects that can last for at least 7 days. 36 –42 Two doses of LY341495 were administered prior to NOR training and testing and then one additional dose, rationalized as a booster dose, was given prior to fear training and testing the following week. Elimination half-life of LY341495 in brain has been reported to be 4.75 h. 43 mGluR2/3 antagonists like LY341495 are often compared with drugs such as ketamine 44 and like ketamine, they seem to exert biological effects that extend well beyond their pharmacological half-life.
Indeed, it is interesting to compare the current findings with our recent studies using (2R,6R)-hydroxynorketamine [(2R,6R)-HNK] to treat blast-exposed rats. 22 (2R,6R)-HNK like its parent ketamine has biological effects that extend well beyond its pharmacological half-life, which is on the order of hours. 45 Like LY341495, (2R,6R)-HNK exerts antidepressant actions at least in part through mGluR2-dependent mechanisms. 46,47 In blast-exposed rats, a single dose of (2R,6R)-HNK still exerted effects on NOR 27 days after treatment, suggesting that modulation of mGluR2 activity may exert prolonged effects well beyond the pharmacological half-lives of LY341495 or (2R,6R)-HNK.
How these compounds exert such long-lasting effects is incompletely understood but likely occur through second messenger systems able to modulate growth factor and synaptic protein related pathways. 44 Future studies will be needed to optimize LY341495 dosing as well as determine its length and mechanism of prolonged action in blast-exposed rats. Studies have also suggested that subtherapeutic doses of LY341495 effects can enhance effects of (R) ketamine in a mouse model of chronic unpredictable stress. 48 Similar studies combining LY341495 and ketamine in blast-exposed rats seem warranted.
Why mGluR2 expression should be elevated following blast exposure is unclear. Indeed, little is known about regulation of mGluR2 expression. One well-studied pathway links changes in mGluR2 RNA to a stress-related pathway that involves changes in brain-derived neurotrophic factor (BDNF) and a P300 driven change in histone acetylation of the GRM2 gene. 49 Both increased and decreased mGlu2/3 receptor expression has been found in animal models of depression. 50 -52 Studies in postmortem tissue from human depression have found mGluR2/3 expression to be increased, decreased or unchanged. 53 -55 The reason for these discrepancies is unclear.
It is also unclear why expression of mGluR2 should be preferentially affected over mGluR3. mGluR2 and mGluR3 receptors have overlapping but not identical expression patterns. 56,57 Receptor mapping using selective radiolabeled ligands have shown that mGlu2/3 receptors are prominently expressed in cortical and limbic regions. 56,57 In these regions, mGluR2 is prominently localized at perisynaptic sites, where it functions as an autoreceptor or heteroreceptor. 58 mGluR3 is localized at presynaptic and postsynaptic sites, as well as on glial cells. 59 The significance of these differing patterns is unknown although mGluR2 and mGluR3 knockout mice react differently to mGluR2/3 antagonists, 60 indicating a functional significance to their differential patterns of expression or downstream molecular coupling.
The implications of mGluR2 overexpression for the pathophysiology of blast-related brain injury is also unclear. At glutamatergic synapses, glutamate stimulates postsynaptic α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptors. Stimulation of postsynaptic AMPA receptors is linked to increased BDNF and stimulation of mTORC through activation of tropomyosin-related kinase B (TrkB) receptors. 11 Presynaptic mGluR2/3 receptors act as autoreceptors that when stimulated inhibit glutamate release. 11 Thus, by analogy, increased mGluR2 receptor levels would predict that chronic blast injury induces a hypoglutamatergic state. This analogy assumes that pharmacological blockade has the same physiological consequences as reducing receptor overexpression. Support for this argument can be seen in our previous findings with the mGluR2/3 antagonist BCI-83812 and the effects with LY341495 reported here.
Actions at mGluR2/3 receptors are complex, however, in that mGluR2/3 receptor agonists also have antidepressant effects and have shown beneficial effects on markers of acute neuronal injury in rodent TBI models involving lateral fluid percussion and controlled cortical impact injuries. 61 –64 Further study of the neurochemical effects of increased mGluR2 receptor expression after blast exposure will clearly be needed.
A limitation of the current study is the lack of inclusion of female rats. Sex differences in TBI outcomes are well known, 65 with studies in female veterans suggesting that they are more likely to report persisting neurobehavioral symptoms and use more outpatient services than their male counterparts. 66 In experimental animals, sex differences in response to blast has been little studied, although several reports have suggested that blast responses in female rats and mice differ. 67 -69 With the increasing number of female veterans these studies assume a high importance.
Conclusions
The findings in this report strongly suggest that alterations in mGluR2 expression play a key role in mediating the pathophysiological cascades set in motion by repetitive blast. It is additionally significant that blocking this pathway with group II metabotropic antagonists (either before or even after behavioral traits emerge) significantly reduces cognitive and PTSD-like impairment in rats. It is often the case that Veterans seeking care for mTBI-related health concerns first receive clinical attention long after the TBI events which sparked their chronic symptoms. As such, an ongoing “holy grail” of rehabilitative medicine is to find new treatment strategies that work when initiated after substantial delays. The data in this report augur well for the value of further exploring such approaches to treat blast-related mTBI.
Transparency, Rigor, and Reproducibility Summary
The analysis plan was not pre-registered in a public domain but was pre-specified based on similar previous studies. 5,12,15,17,70 Sample sizes were based on previous studies that found that n = 5-6 animals per group were sufficient to obtain statistically significant results in Western blotting 5,15,17,70 and that group sizes of 13 control and 6-7 blast exposed subjects provided sufficient power to see drug effects in blast-exposed rats treated with mGluR2/3 antagonists in behavioral studies. 12
Transportation of rats after blast exposure may induce an added element of stress. Any element of stress, however, has not precluded us from establishing a reproducible model of blast-induced behavioral effects, which model the chronic neurobehavioral syndromes seen in military veterans. 5,8 To control for any effects of environment or transportation, all animals whether assigned to sham or blast conditions are purchased at the same time and housed under similar housing conditions at the WRAIR. Sham exposed animals are treated identically to blast, including receiving anesthesia and placement inside the blast tube, but do not receive a blast exposure. Within 10 days after the last blast or sham exposure animals are transported together within one day from the WRAIR to the Bronx VA where blast and sham are housed under identical conditions. It remains possible, however, that transportation stress could result in an interaction effect that might differentially affect blast versus sham subjects.
Sixty-five rats subjected to blast or sham injury were used in this study. Subjects were singly housed on racks in random order to prevent rack position effects. Investigators performing behavioral testing were blinded to the group status during testing. For each time-point shown in this study, the tissues utilized for Western blot or RNA isolation were from animals that were part of cohorts that were blast/sham exposed the same days and sacrificed at the same time.
Dosing of LY341495 was based on previous studies in rats showing that even single doses administered at 1 mg/kg or 3 mg/kg exert sustained effects on behavior that can last for at least 7 days. 36 –42 No sources of USP pharmaceutical grade LY341495 are available for either humans or animals. Therefore, we used LY341495 from Tocris which has been widely used in rats for IP injection for similar studies 36 –42 without any described toxicity. We noted no toxicity in the current studies. Solutions were prepared fresh the day of each administration, filtered through an 0.45-μm filter to ensure sterility, and stored protected from light until use.
Biochemical data involved two group unpaired designs and comparisons were performed using unpaired t-tests. For behavioral studies following drug treatment, comparisons were performed using one-way ANOVA, repeated-measures ANOVA, or two-tailed unpaired t-tests. When repeated-measures ANOVA was used significance was determined using the Greenhouse-Geisser correction. 23 Due to the three-group design in the behavioral/drug studies, post hoc tests following a significant ANOVA were performed using Fisher's LSD. 24
The authors agree to provide full public access to the paper as required by Department of Veterans Affairs VHA Directive 1200.19. Original data is not currently deposited in any public repository but will be made available upon reasonable request.
Footnotes
Acknowledgments
The views expressed in this article reflect the results of research conducted by the authors and do not necessarily reflect the official policy or position of the Uniformed Services University of the Health Sciences (USUHS), The Henry M. Jackson Foundation for the Advancement of Military Medicine, Inc., the Department of the Navy, or the Department of Defense (DoD). Mention of trade names, commercial products, or organizations does not imply endorsement by the U.S. Government. The study protocol was reviewed and approved by the Walter Reed Army Institute of Research/Naval Medical Research Command Institutional Animal Care and Use Committee in compliance with all applicable federal regulations governing the protection of animals in research. The experiments reported herein were conducted in compliance with the Animal Welfare Act and per the principles set forth in the “Guide for Care and Use of Laboratory Animals,” Institute of Laboratory Animals Resources, National Research Council, National Academy Press, 2011. Some of the authors are military service members or federal/contracted employees of the United States Government. This work was prepared as part of their official duties. Title 17 U.S.C. § 105 provides that “Copyright protection under this title is not available for any work of the United States Government. Title 17 U.S.C. § 101 defines a U.S. Government work as a work prepared by a military service member or employee of the U.S. Government as part of that person's official duties.”
Authors' Contributions
RDG: Conceptualization; Data curation; Formal analysis; Investigation; Project administration; Visualization; Writing—review and editing. MAG: Conceptualization; Data curation; Formal analysis; Investigation; Visualization; Writing—review and editing; GPG: Conceptualization; Data curation; Formal analysis; Investigation; Visualization; Writing—review and editing; GMP: Investigation; DP: Investigation; Visualization. RA: Investigation; Writing—review and editing. UK: Investigation; Writing—review and editing. JS: Investigation. JP: Investigation. PRH: Writing—review and editing. CWZ: Formal analysis; Writing—review and editing. SAE: Methodology; Funding acquisition; Writing—review and editing. DGC: Conceptualization; Funding acquisition; Visualization; Writing—review and editing. GAE: Conceptualization; Data curation; Formal analysis; Funding acquisition; Project administration; Supervision; Visualization; Writing—original draft.
Funding Information
This work was supported by Department of Veterans Affairs, Veterans Health Administration, Rehabilitation Research and Development Service awards 1I21RX003459 (MAGS) 1I01RX002660 (GE), 1I01RX003846 (GE), and Department of Veterans Affairs Office of Research and Development Medical Research Service awards 1I01BX004067 (GE), 1I01BX005882 (GE) and 1I01BX002311 (DC), Department of Defense work unit number 0000B999.0000.000.A1503 (STA) and NIA P30 AG066514 (PRH, CWZ).
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.