
Abstract
Mild traumatic brain injury (mTBI) is often accompanied by neurological and ocular symptoms that involve trigeminal nerve pathways. Laser-induced shock wave (LISW) was applied to the skull of male rats as a model for mTBI, while behavioral and neural recording methods were used to assess trigeminal function. The LISW caused greater eye wiping behavior to ocular instillation of hypertonic saline (Sham = 4.83 ± 0.65 wipes/5 min, LISW = 12.71 ± 1.89 wipes/5 min, p < 0.01) and a marked reduction in the time spent in bright light consistent with enhanced periocular and intraocular hypersensitivity, respectively (Sham = 16.3 ± 5.6 s, LISW = 115.5 ± 27.3 s, p < 0.01). To address the early neural mechanisms of mTBI, single trigeminal brainstem neurons, identified by activation to corneal or dural mechanical stimulation, were recorded in trigeminal subnucleus interpolaris/caudalis (Vi/Vc) and trigeminal subnucleus caudalis/upper cervical cord (Vc/C1) regions. The LISW caused marked sensitization to hypertonic saline and to exposure to bright light in neurons of both regions (p < 0.05). Laser speckle imaging revealed an increase in meningeal arterial blood flow to bright light after LISW (Sham = 4.7 ± 2.0 s, LISW = 469.0 ± 37.9 s, p < 0.001). Local inhibition of synaptic activity at Vi/Vc, but not at Vc/C1, by microinjection of CoCl2, prevented light-evoked increases in meningeal blood flow in LISW-treated rats. By contrast, topical meningeal application of phenylephrine significantly reduced light-evoked responses of Vi/Vc and Vc/C1 neurons. These data suggested that neurons in both regions became sensitized after LISW and were responsive to changes in meningeal blood flow. Neurons at the Vi/Vc transition and at Vc/C1, however, likely serve different roles in mediating the neurovascular and sensory aspects of mTBI.
Introduction
Mild traumatic brain injury (mTBI), often referred to as concussion, is commonly accompanied by headache, cognitive deficits, and vision-related symptoms. 1 The ocular surface also can be affected by mTBI and displays signs seen in patients with dry eye disease. 2,3 Several features of mTBI-induced post-traumatic headache (PTH) and dry eye such as increased sensitivity to bright light and lower cutaneous sensory thresholds to mechanical stimulation in humans 4,5 and in animal models 6 –9 suggest common underlying mechanisms; however, the neurological basis for such mechanisms is not well defined.
The ocular surface and the meninges are innervated mainly by the ophthalmic branch of the trigeminal nerve. 10 Trigeminal sensory nerve fibers that supply the ocular surface 11,12 and the meninges 13 terminate centrally in two spatially discrete regions of the trigeminal brainstem: the trigeminal subnucleus interpolaris/caudalis (Vi/Vc) transition region and more caudally at the subnucleus caudalis/upper cervical spinal cord (Vc/C1) junction region. The neuronal activity marker, c-fos, is highly expressed by neurons at the Vi/Vc and Vc/C1 regions after corneal stimulation, 14,15 dural stimulation, 16,17 and after bright light stimulation. 18
Neural recording studies indicate a significant percentage of Vi/Vc and Vc/C1 neurons are activated by both corneal and dural stimuli 19,20 and suggest these regions are critical for processing convergence of input from craniofacial structures supplied by the ophthalmic branch of the trigeminal nerve. Although neurophysiological evidence suggests that neurons in each group serve different functions related to ocular function in models for dry eye, 21 –23 no previous study has assessed the functional properties of Vi/Vc and Vc/C1 neurons in a model for mTBI.
Photoallodynia, or photophobia, is a common symptom of mTBI, PTH, and dry eye. 5 Neurovascular communication has been proposed as a potential mechanism for mTBI- and PTH-induced photophobia. 24,25 Although light-evoked Vc/C1 neuronal activity has been shown to depend on an increase in autonomic outflow to the eye, 26 the effects of changes in vascular function after mTBI on trigeminal brainstem activity has not been assessed. The present study addressed the issue of vascular function in mTBI-related light sensitivity by quantifying mTBI-induced meningeal blood flow by laser speckle imaging and by trigeminal brainstem recording in male rats. Laser-induced shock waves (LISW) were applied as a closed head model for mTBI 27 and to better approximate the localized impact to the skull that occurs in many concussions.
Methods
Animals
A total of 131 male Sprague–Dawley rats (250–380 g; SLC, Shizuoka, Japan) were used in these experiments. The experimental protocols were approved by the Committee on Research Facilities for Laboratory Animal Science, National Defense Medical College (Japan) and conformed to the established guidelines set by The National Institutes of Health for the care and use of laboratory animals. Rats were maintained under a 12-h light-dark cycle in a temperature-controlled (22–25°C) environment with free access to food and water. Experiments were performed during the periods of 9 AM to 3 PM.
LISW methods
Rats were anesthetized with pentobarbital sodium (50 mg/kg intraperitoneally [ip]), and the parietal regions of the skull were shaved to avoid trapping air in the fur. As described previously, 27 LISW was generated by irradiating a laser target with a 532 nm Q-switched Nd:YAG laser (Brilliant b, Quantel, Lannion, France; pulse width, 6 ns full width at half maximum). The laser target was a 10-mm (diameter), 0.5-mm (thick) black natural rubber disk. A 1.0-mm thick transparent polyethylene sheet was bonded to the top of the target to confine the laser-induced plasma and increase the LISW impulse (Fig. 1). The pulsed laser beam was focused with a planoconvex lens to a 3.0 mm spot on the laser target. The laser fluence was set to 4.0J /cm2 directed to the temporal left side of the head between the corner of the eye and the ear. To ensure acoustic impedance matching, ultrasound conductive gel was used between the laser target and the skin surface. Sham rats also were anesthetized with pentobarbital and prepared surgically for LISW but were not pulsed. All experiments were performed without previous knowledge of treatment (i.e., Sham vs. LISW).
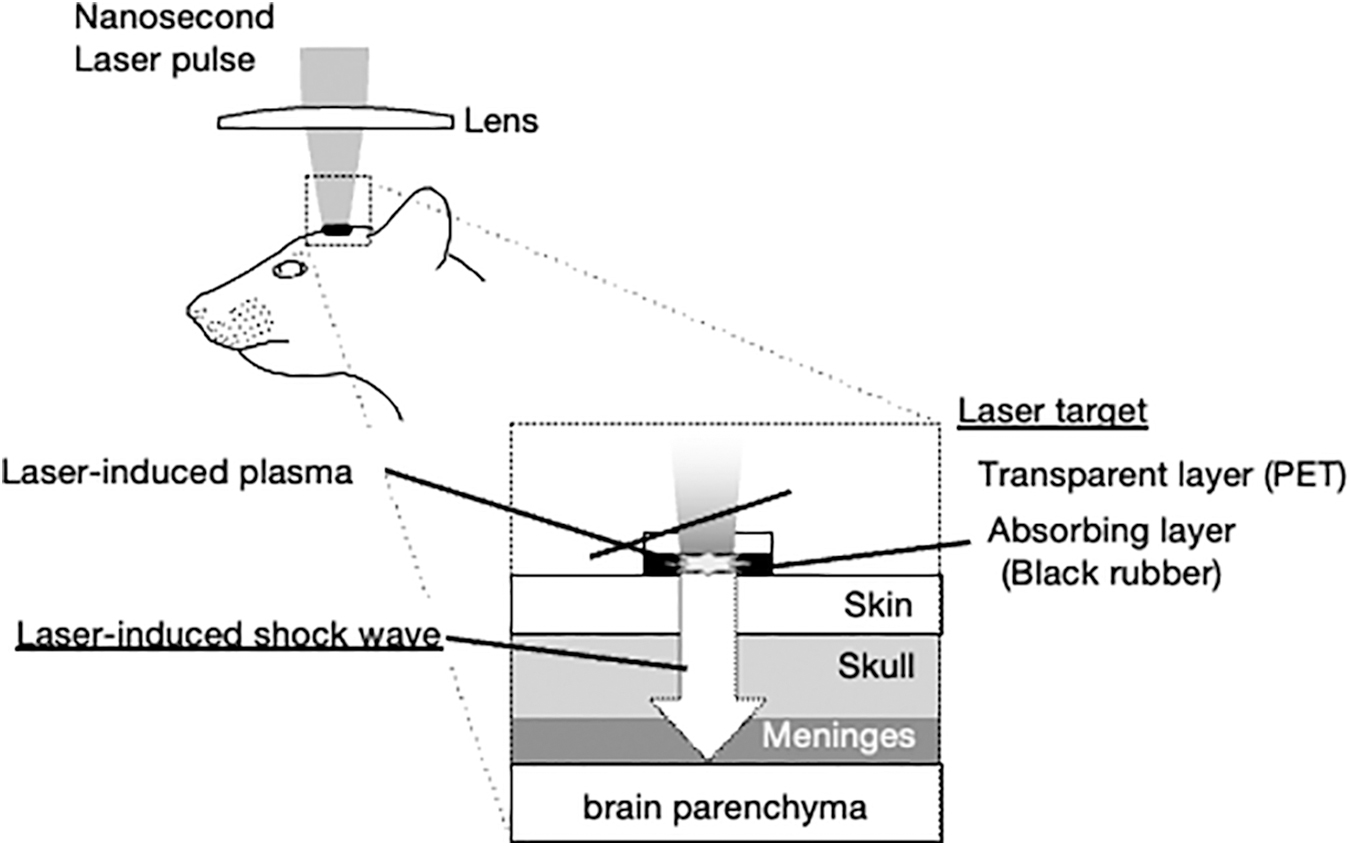
Experimental setup for the generation of laser-induced shock wave (LISW).
Meningeal inflammation
Meningeal plasma extravasation was determined by Evans blue dye to assess the extent of LISW-induced inflammation. Evans blue dye (100 mg/kg/mL) was injected in the right femoral vein, and rats were perfused with 0.9% NaCl after 20 min. After decapitation, the skull was removed, and the head was post-fixed in 4% paraformaldehyde for 1–2h to allow the collection of the parietotemporal meninges. The membrane samples were collected, immersed in formamide (0.1 mL), and incubated overnight at room temperature. Fluorescence intensity was measured by a microplate fluorescence reader (620 nm excitation; iMark™ Microplate Absorbance Reader; Bio-Rad, Hercules, CA), and dye concentrations were based on external standard curves.
Behavioral tests
Eye wipe behavior
The eye wipe test was performed using hypertonic saline as the inducing stimulus. 28,29 The test stimulus consisted of a single drop of saline solution (2.5 M NaCl, 20 μL) applied to the ocular surface from a micropipette at 30 min intervals under ambient light (300 lux). The eye was flushed with normal saline after 10 min after each application of the test solution. The number of eye wipes, defined as purposeful wiping of the face by the forelimb, was counted over 5 min on days 1–10 after LISW.
Light aversion test
Rats were tested in a custom-made light aversion plexiglass chamber (60
Electrophysiology/blood flow recording
Surgical procedures
Rats were initially sedated with pentobarbital sodium (70 mg/kg ip), and catheters were placed in the femoral artery (blood pressure) and jugular vein (paralytic agent). After tracheotomy, animals were respired with oxygen-enriched room air and maintained under isoflurane anesthesia (1.0%–1.5%). Rats received an infusion of the short-acting paralytic agent, gallamine triethiodide (25 mg/kg/h), at the time of neural recording. Expiratory end-tidal CO2 (3.5%–4.5%), mean arterial pressure (100–120 mm Hg), and body temperature (38°C) were monitored continuously and maintained within the normal range.
Rats were placed in a stereotaxic frame, and portions of the upper cervical (C1-2) vertebrae were removed to allow access to the lower brainstem and cervical spinal dorsal horn. A partial craniotomy was cut into the parietal bone to expose the underlying dura mater using a drill and fine forceps to allow topical application of phenylephrine (1 mM, 20 μL) and for monitoring meningeal blood flow.
Electrophysiology procedures
Single neurons were recorded at the ventrolateral trigeminal subnucleus interpolaris/caudalis transition region (Vi/Vc) transition and the superficial laminae (laminae I–II) at the Vc/C1 junction region (see Fig. 4). The Vi/Vc region was approached at a 28-degree angle off vertical, 45 degrees off midline, and 1.5–2.0 mm below the brainstem surface. The Vc/C1 region was approached at a 47-degree angle off vertical, 60 degrees off midline, and within 250 μm of the dorsal brainstem surface. Extracellular unit activity was recorded with tungsten microelectrodes (5 MΩ, Frederick Haer & Co. Inc., Bowdoinham, ME), amplified, discriminated (model WD-2, Bio Research Center Co. Ltd., Nagoya, Japan), stored, and analyzed offline using a PowerLab interface and LabChart software (ADInstruments Pty. Ltd., Bella Vista, NSW, Australia).
Characterization of neurons
A single neuron was recorded from each rat in most experiments and identified by spike sorting methods based on spike amplitude, duration, and waveform shape to confirm adequate isolation. All neurons at the Vi/Vc and Vc/C1 regions were spontaneously active under ambient light conditions (300 lux) and were further identified by an increase in discharge rate to gentle mechanical stroking of the ocular surface.
Chemical stimulation of the ocular surface
A subset of neurons was tested by applying hypertonic saline (NaCl, 0.15–2.5 M, 20 μL) to the ocular surface after establishing a response to mechanical stimulation of the ocular surface. Neural responses were quantified in naive (n = 15) and LISW rats (n = 15) and compared with the response to normal saline solution (22–25°C).
Light stimulation
Light stimulation was delivered from a thermal-neutral fiber optic LED source (KL 1600 LED SCHOTT AG, Mainz, Germany) positioned 3 cm from the left ocular surface under low ambient light conditions. Light stimuli (40 sec duration) were presented in a cumulative intensity design at 20-min intervals (low, moderate, and high intensities; 3000 lux, 10000 lux, and 20,000 lux, respectively).
Effect of phenylephrine on neural activity
In separate experiments, Vi/Vc (n = 3) and Vc/C1 neurons (n = 3) exposed to LISW 7–10 days previously were identified by dual responses to mechanical probing of the ocular surface and to the dura. Phenylephrine (1 mM, 20 μL) was applied topically to the dural surface through the cranial window to determine whether the large afterdischarge typically seen after light stimulation (10,000 lux) was from changes in meningeal blood flow. After topical application, mechanical stimulation was again applied to the dura (1.4g von Frey) to confirm that phenylephrine did not have a generalized effect on nerve ending sensitivity.
Meningeal blood flow procedures
In separate experiments, meningeal blood flow was measured from the branches of the middle meningeal artery in response to moderate intensity light stimulation (total n = 25 rats). Recordings were performed through the cranial window under isoflurane anesthesia (1.5%). Light stimulation (10,000 lux) was presented under low ambient room light (< 100 lux). The region of interest was set over the branches of the middle meningeal artery lying distant from visible cortical blood vessels. 30 Blood flow was imaged with a laser speckle blood flow imaging system (OZ-2 OMEGAWAVE, INC., Tokyo, Japan). The involvement of the Vi/Vc transition and Vc/C1 regions was assessed by microinjection of the synaptic blocker CoCl2 (100 mM, 100 nL, pH 7.2), delivered via a glass micropipette at the coordinates noted above for recording, and ipsilateral to the stimulated eye 10 min before the light stimulus (10,000 lux). Drug controls received an equivalent volume injection of aCSF (pH 7.4, 100 nL). Microinjection sites were confirmed histologically at the end of the experiment.
Data analysis
All statistical analyses were performed using GraphPad software (Prism, v. 9). Evans Blue dye results were compared by analysis of variance (ANOVA). Eye wipe data were compared over time by two-way ANOVA corrected for repeated measures on one factor (days post-LISW), and individual comparisons were made by Tukey test after ANOVA. Neural recording data were analyzed as a response magnitude (Rmag), equivalent to the “area under the curve” and defined as the mean spike count + two times the standard deviation (SD) of the background activity (1 min) for each 1-sec bin for each stimulus period. 31
Total Rmag was assessed by two-way ANOVA corrected for repeated measures on one factor (NaCl concentration), and individual comparisons were made by Tukey test after ANOVA. To assess the effects of phenylephrine on Rmag, the data were log transformed prior to ANOVA to reduce error variance and to better achieve a normal distribution for small sample sizes. Afterdischarges after light stimuli were sampled for 5 min beginning 1 min before vehicle (aCSF) or drug application.
Blood flow was recorded as arbitrary flux units, continuously monitored, stored, and analyzed offline using an analysis software (RIV-V102 and LIV-V321, OMEGAWAVE, INC.). The baseline blood flow was quantified as the area under the curve for a 40-sec epoch sampled immediately before stimulation. The data were normalized to the percentage of basal blood flow sampled for an equivalent 40-sec epoch before light stimulation. The effects of LISW on blood flow were assessed by t test. The effects of CoCl2 microinjections into the Vi/Vc and Vc/C1 regions on meningeal blood flow were assessed by two-way ANOVA corrected for repeated measures on one factor (time after injection). A p-value of <0.05 was considered significant.
Results
General effects of LISW
No visible signs of brain injury were noted over the 10 days after LISW radiation, and bodyweight gain was similar for the sham and LISW rats. Evans blue dye concentration was significantly greater at 2 and 10 days after LISW compared with sham rats (Fig. 2, F2,15 = 8.86, p < 0.01) and confirming significant meningeal inflammation.

Effect of laser-induced shock wave (LISW) on increased plasma extravasation in meningeal samples by Evans blue dye after LISW. Sample sizes: sham, n = 7; 2 days after LISW, n = 5; 10 days after LISW, n = 6. *p < 0.05, **p < 0.01 vs. sham. Color image is available online.
Effect of LISW on eye wipe behavior and light aversion
Eye wiping behavior
Physiological saline (0.15 M) did not evoke eye wiping in naïve or LISW rats (data not shown). By contrast, the eye wipe response to repeated application of 2.5 M NaCl was significantly increased at 2–10 days after LISW when compared with responses in sham rats (F1,11 = 29.1, p < 0.001; Fig. 3A).
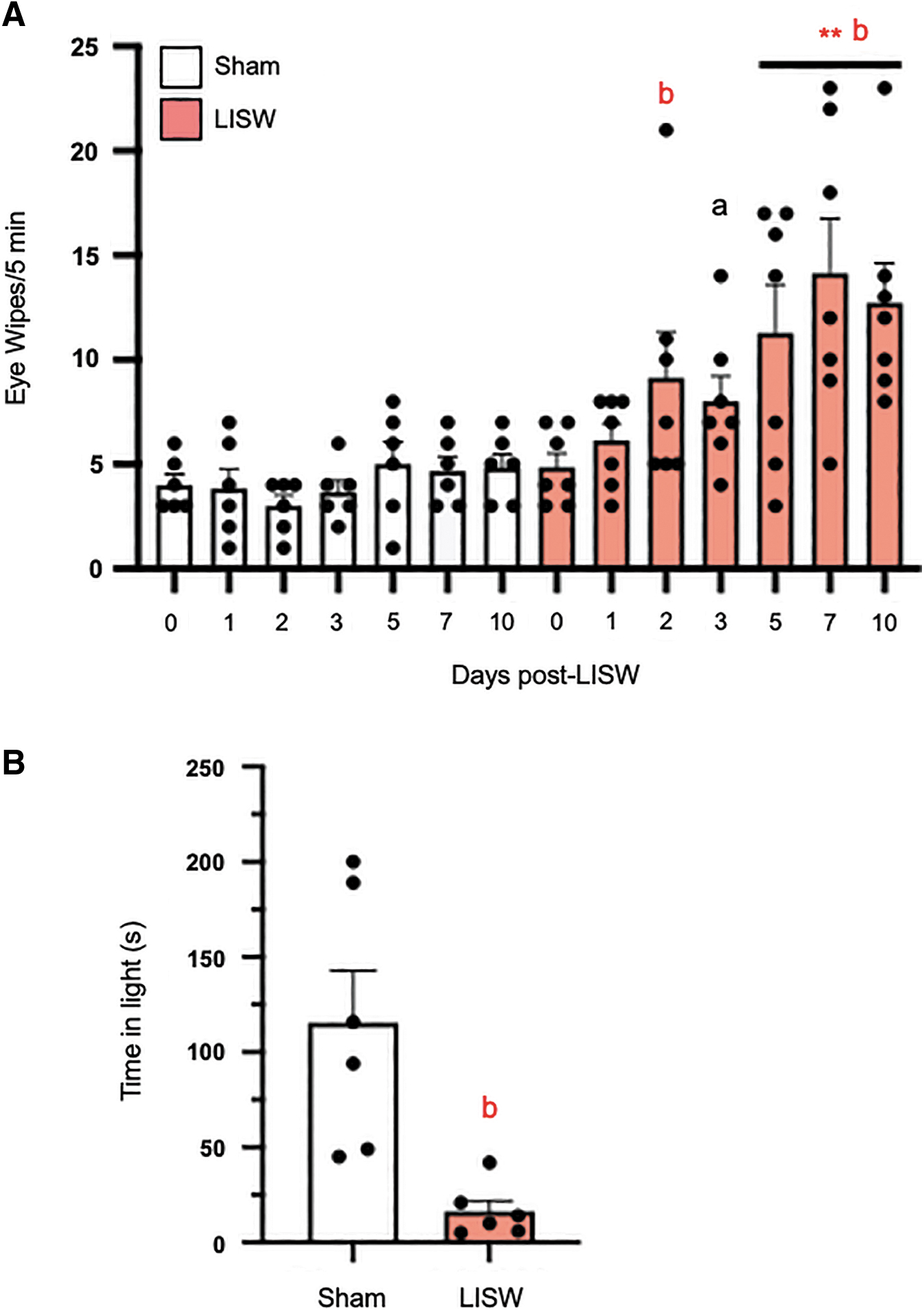
Laser-induced shock wve (LISW) enhanced nocifensive behavior. (
Light aversion
Light sensitivity was assessed by an exploratory assay in which the rats were given the choice of being in the light (3000 lux) or dark compartment (100 lux) of the test chamber. The LISW rats spent 86% less time in the light (16.3 ± 5.6 sec) than the sham rats when tested at 10 days post-LISW (115.5 ± 27.3 sec, t test = 3.55, df = 10, p < 0.005; Fig. 3B).
Effect of LISW on neural activity
Only neurons recorded from the ventrolateral pole of the Vi/Vc transition region or within 250 μm of the dorsal brainstem surface at the Vc/C1 region were included in these analyses (Fig. 4).
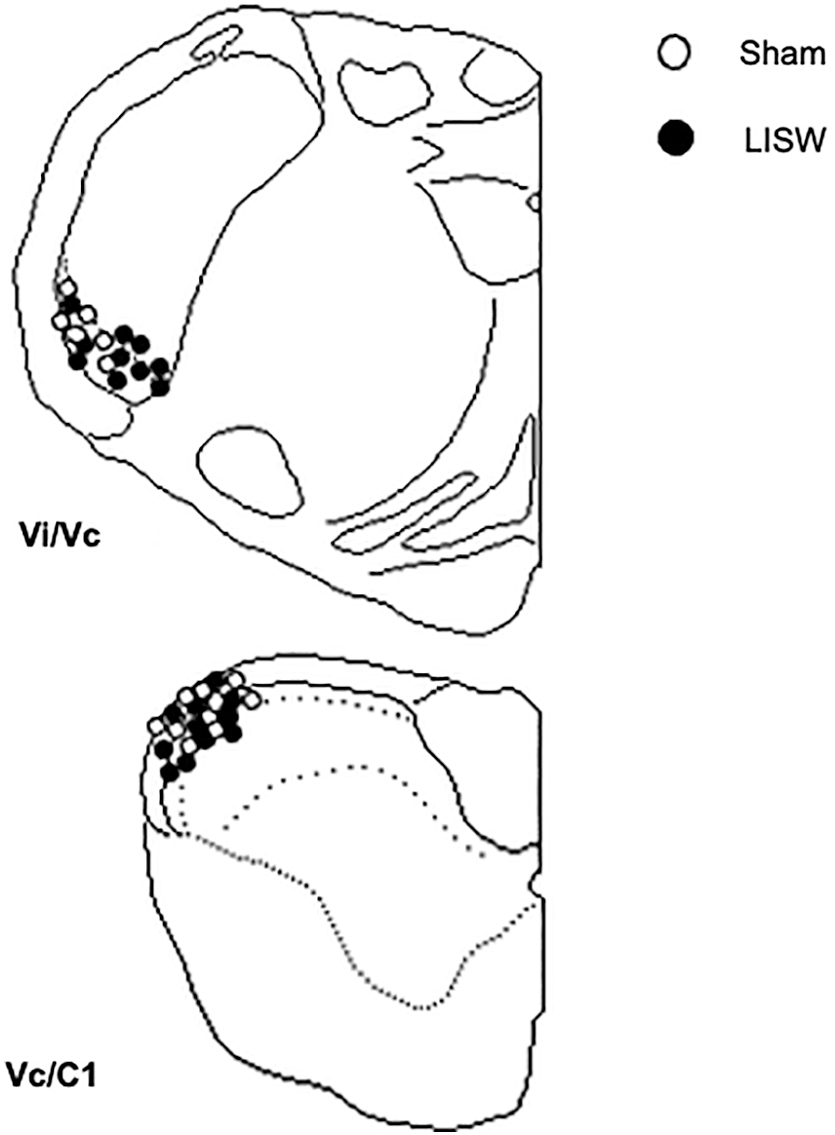
The location of recording sites. Recovered lesion sites for ocular/dura or ocular units recorded from Vi/Vc transition and Vc/C1 junction regions in naïve and laser-induced shock wave rats.
Background activity
The background activity before stimulation for units tested for responses to hypertonic saline or to bright light was analyzed by recording site and found to be similar for neurons at Vi/Vc (sham = 3.1 ± 1.3, n = 15; LISW = 3.5 ± 1.7 spikes/sec, n = 14, p > 0.05) and at the Vc/C1 region (sham = 2.0 ± 0.4, n = 14; LISW = 3.6 ± 0.9 spikes/sec, n = 16, p > 0.05).
Ocular surface stimulation
A total of 30 cornea responsive Vi/Vc (n = 14) and Vc/C1 (n = 15) neurons were recorded from 29 rats and tested for responses to ocular surface instillation of hypertonic saline (0.15, 2.5, 5M NaCl). The examples shown in Figures 5A and 5B are from Vc/C1 units. Hypertonic saline evoked a concentration-dependent increase in all Vc/C1 units as seen in the examples for sham rats (Fig. 5B, upper panel) and for LISW-treated rats (Fig. 5B, lower panel). The responses to hypertonic saline are summarized in Figure 5C for Vi/Vc units from sham and LISW rats (left panel) and for Vc/C1 units (right panel).
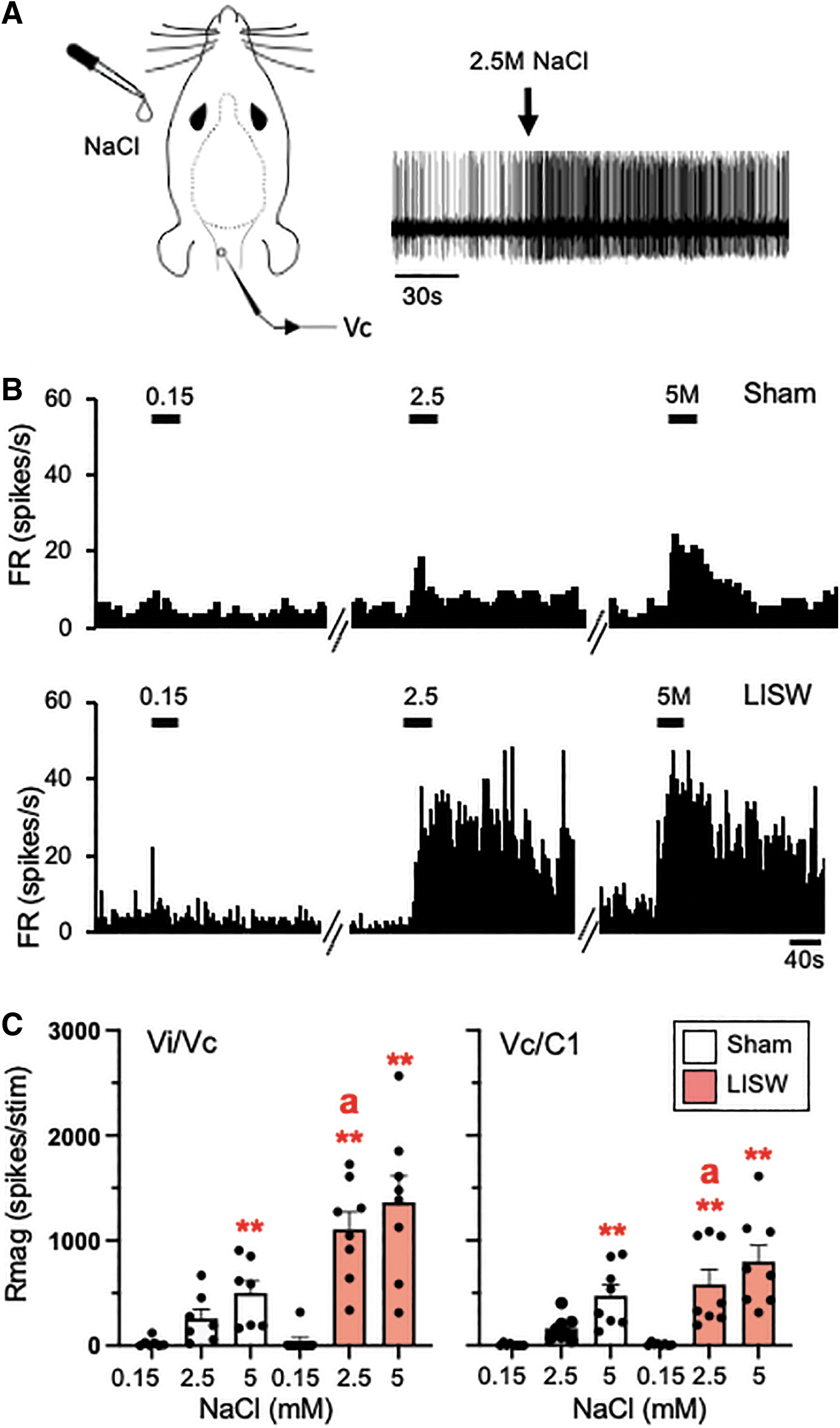
Laser-induced shock wave (LISW) enhanced Vi/Vc and Vc/C1 neural activity evoked by topical instillation of hypertonic saline on the ocular surface. (
When compared with the responses to 0.15 M NaCl, the Rmag responses to hypertonic saline were significantly increased for Vi/Vc units of sham (F2,24 = 25.3, p < 0.001) and LISW-treated rats (F2,24 = 79.1, p < 0.001). Similarly, hypertonic saline evoked significant increases in Rmag for Vc/C1 units of sham (F2,28 = 71.3, p < 0.001) and LISW-treated rats (F2,28 = 70.7, p < 0.001) compared with the responses to 0.15 M NaCl.
Light stimulation
A total of 29 cornea-responsive neurons were recorded from 28 rats at the Vi/Vc (n = 14) and the Vc/C1 region (n = 15) and tested for responses to bright light. The low-intensity light stimulus (3K lux) was sufficient to activate 6 of 14 units from sham rats and 13 of 15 units from LISW rats (Rmag value >10) independent of recording location. The examples shown in Figures 6A and 6B are recorded from Vi/Vc units. Note the delay in light-evoked activity after the 10K lux stimulus, similar to results we reported previously. 26
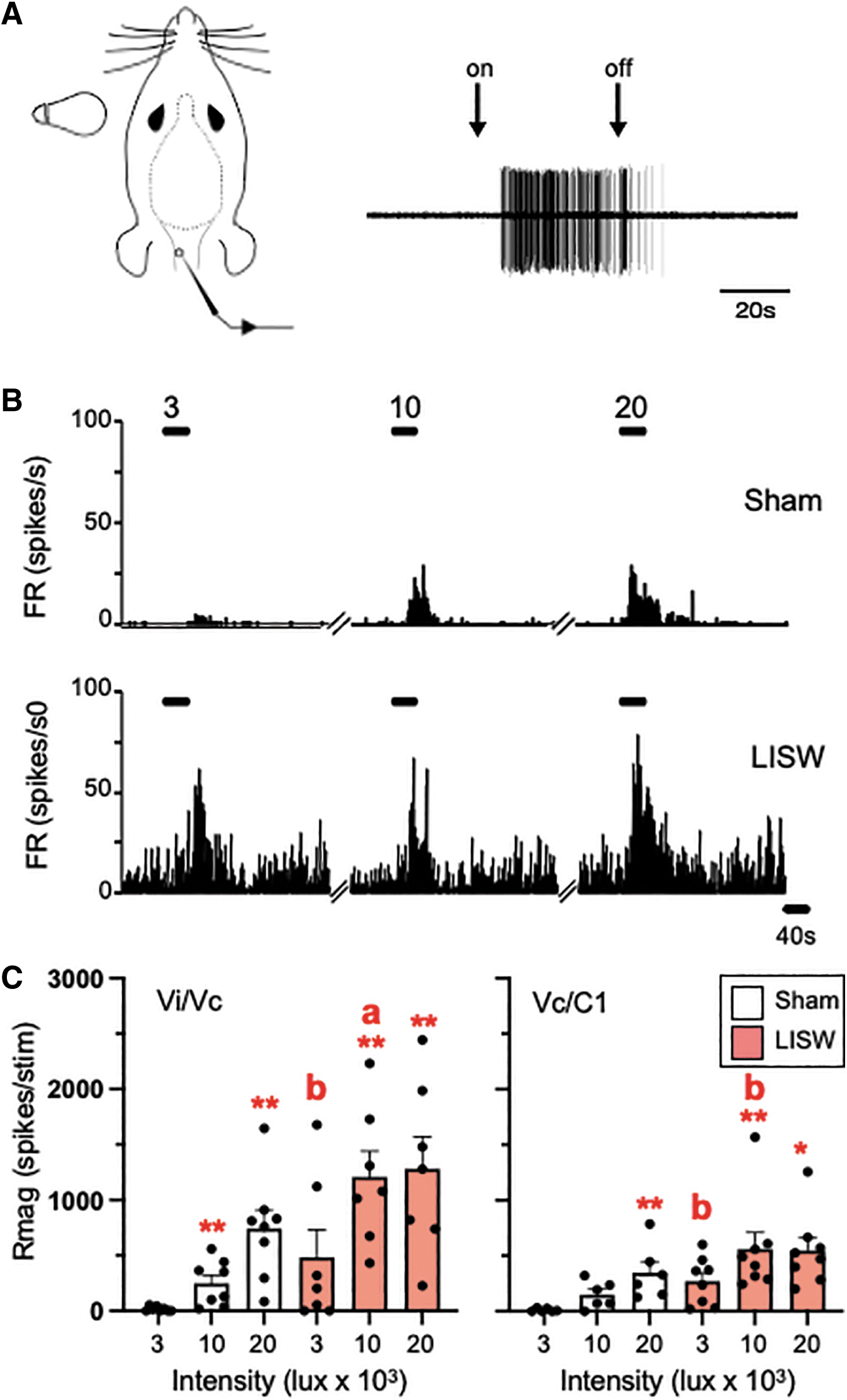
Laser-induced shock wave (LISW) enhanced Vi/Vc and Vc/C1 neural activity evoked by light stimulation of the ocular surface. Light stimuli were presented (3K lux, 10K lux, and 20K lux) as a single 40 sec pulse at 20 min intervals. (
These data are summarized in Figure 6C for Vi/Vc units from sham and LISW rats (left panel) and for Vc/C1 units (right panel). When compared with the responses to low intensity, light evoked significant increases in Rmag to moderate and high intensity light in Vi/Vc units of sham (F2,26 = 19.8, p < 0.001) and LISW-treated rats (F2,26 = 7.49, p < 0.005). Similarly, bright light evoked significant increases in Rmag for Vc/C1 units of sham (F2,24 = 29.2, p < 0.001) and LISW-treated rats (F2,28 = 3.40, p < 0.05) compared with the low intensity stimulus. Note that even the low intensity stimulus was sufficient to enhance the response in Vc/C1 units of LISW-treated rats compared with sham controls (Fig. 6C, right panel, p < 0.01).
Light-evoked changes in meningeal blood flow and trigeminal pathway
As shown by the example in Figure 7A, moderate light intensity (10K lux) evoked a significant increase in meningeal blood flow in LISW rats but not in sham rats (t test = 6.01, df = 23, p < 0.001; Fig. 7B). To determine whether light-induced activity of neurons at the Vi/Vc or Vc/C1 region contributed to the increase in meningeal blood flow, the synaptic blocker CoCl2 (100 mM, 0.2 μL) was microinjected into either region immediately before the light stimulus (10K lux). As shown in Fig. 7C, synaptic blockade at Vi/Vc (F2,32 = 31.9, n = 6, p < 0.001), but not at Vc/C1 (F2,32 = 0.86, n = 6, p > 0.1), prevented the increase in blood flow in LISW-treated rats.
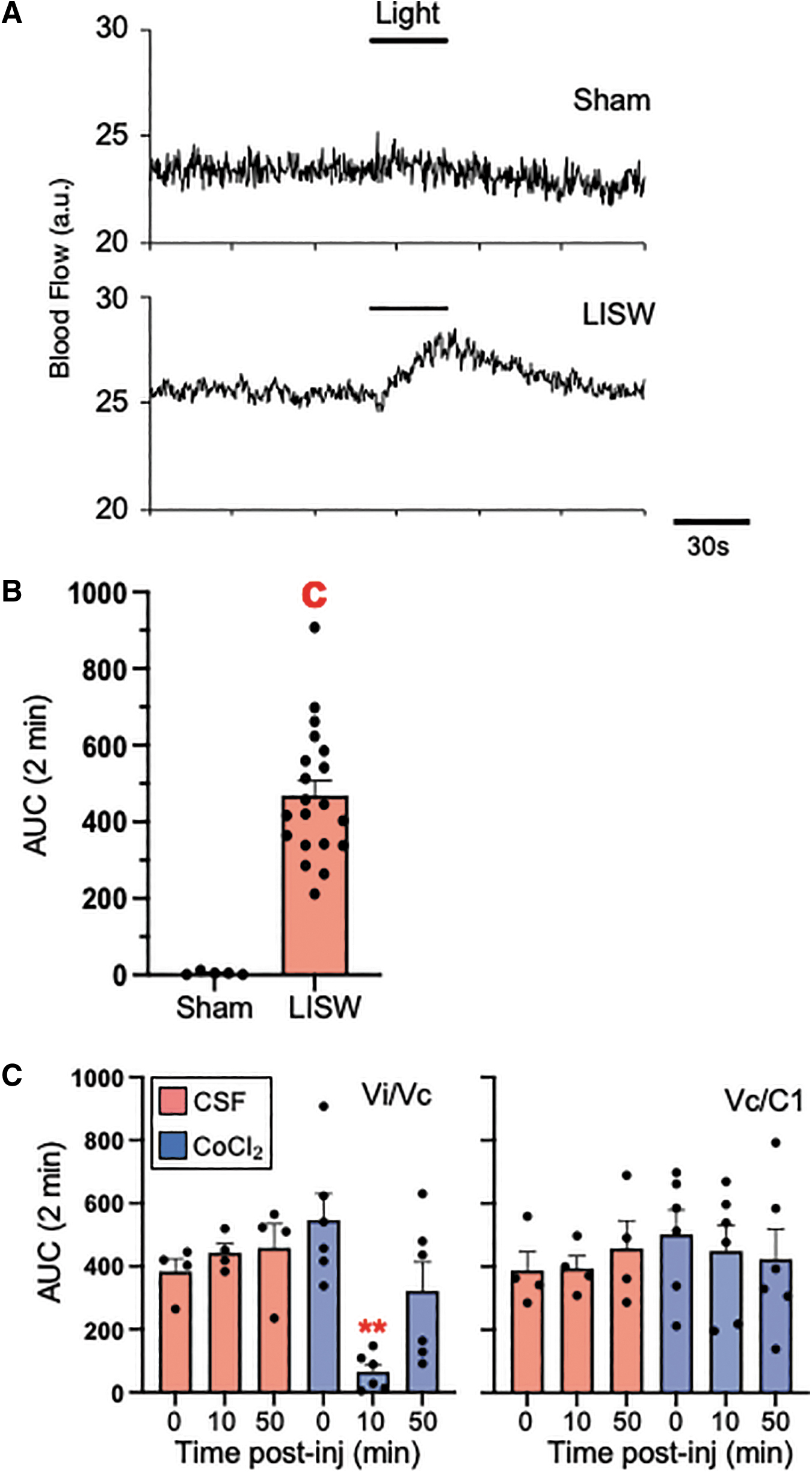
Effect of local synaptic blockade at the Vi/Vc transition or Vc/C1 region on laser-induced shock wave (LISW) enhanced light-evoked increase in meningeal blood flow. (
Injection of CoCl2 had no effect in sham controls regardless of the injection site (p < 0.1). The reduction in meningeal blood after CoCl2 in Vi/Vc of LISW-treated rats was 86.1 ± 5% of control blood flow. Note that the reduction in blood flow returned toward pre-injection values by 30 min suggesting that Vi/Vc neural contributions likely required ongoing activity. In separate animals, microinjection of an equal volume of aCSF into either the Vi/Vc (n = 4) or Vc/C1 (n = 4) region did not alter light-evoked blood flow (F2,32 = <0.6, p > 0.1).
Light-evoked meningeal vasomotor function contributes to Vc neuronal activity
As shown by the example Vi/Vc neuron trace in Fig. 8A, light-induced (20K lux) activity was followed by small, but persistent afterdischarge in LISW-treated rats. To determine whether light-induced increases in meningeal blood flow contributed to the increase in light-evoked firing rate of Vi/Vc or Vc/C1 after LISW treatment, the potent vasoconstrictor, phenylephrine (10 mM, 50 μL) was applied topically to the dural surface immediately before the light stimulus (20K lux).
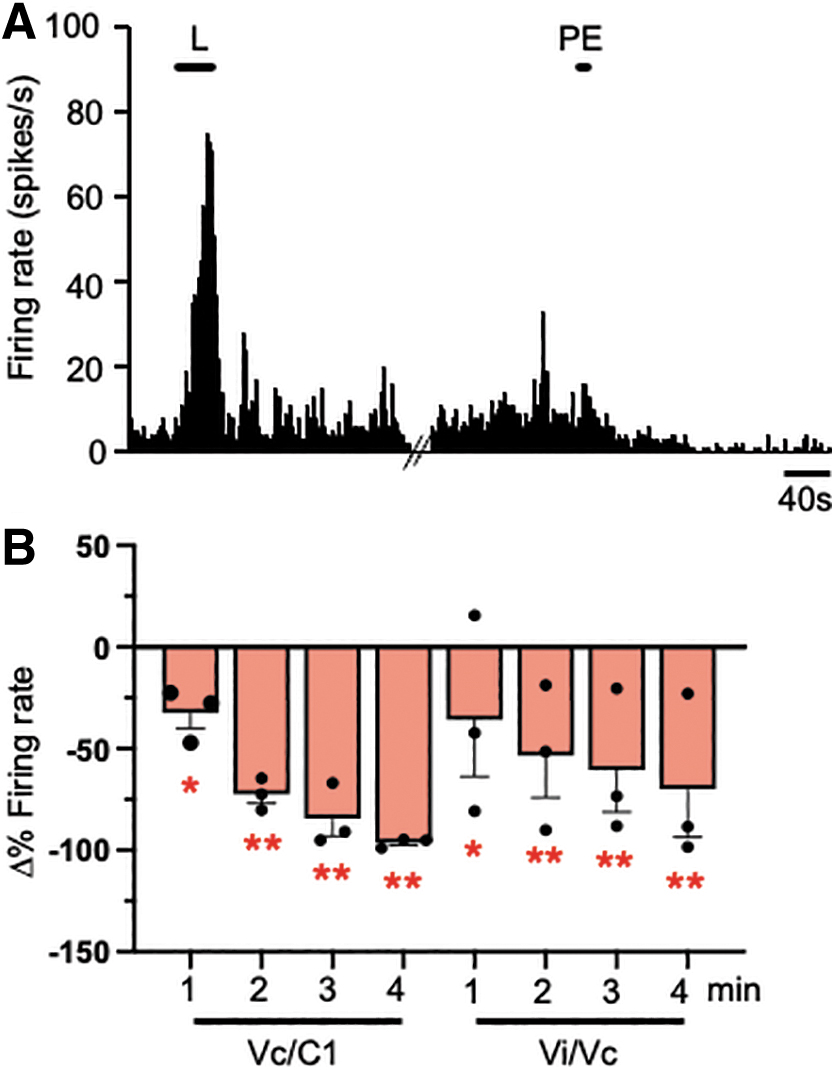
Effect of dural vasoconstriction on light-evoked Vi/Vc and Vc/C1 neuronal activity in laser-induce shock wave (LISW) rats. (
Despite the few numbers of neurons tested (n = 3 per recording site), phenylephrine significantly reduced light-evoked activity in LISW-treated Vi/Vc units (F4,16 = 14.2, p < 0.001) and Vc/C1 units (F4,16 = 4.57, p < 0.025). Phenylephrine application did not affect the neural responses to mechanical stimulation of the dura for any of the units tested (data not shown) suggesting that vascular function was necessary for light-induced activity but did not have a generalized inhibitory effect on the sensitivity of trigeminal nerves that supply the dura.
Discussion
We used a LISW model for mTBI that provided a method to finely control the magnitude and localization of closed head injury by localized shock waves and did not cause skin or bone damage compared with other blunt mechanical TBI models. Although body weight gain was not affected significantly by LISW, it did evoke marked increases in meningeal fluid accumulation, eye wiping behavior, meningeal blood flow, and trigeminal brainstem neural activity induced to ocular surface stimulation. This suggested that LISW was a valid model for mTBI that did not recruit widespread systemic effects.
The main findings of this study revealed an increase in mechanical sensitivity of periocular tissues, light aversion, and sensitization of trigeminal brainstem neurons in a novel closed head model for mTBI. Eye wipe behavior to ocular surface application of hypertonic saline was increased by two days and persisted for at least 10 days after LISW. Light aversion was markedly enhanced at 10 days after LISW.
To better define the neural mechanisms for LISW-induced behaviors, trigeminal brainstem neurons, identified by responses to mechanical stimulation of the ocular surface, were recorded at the ventrolateral pole of the Vi/Vc transition and at the Vc/C1 junction region. Neurons in both regions were sensitized to hypertonic saline and to exposure to bright light at 10 days after LISW. Laser speckle imaging revealed an increase in meningeal blood flow to bright light in LISW-treated rats that was inhibited by synaptic blockade by microinjection of CoCl2 at the Vi/Vc transition region, but not at the Vc/C1 junction. By contrast, application of the potent vasoconstrictor, phenylephrine, to the dural surface inhibited light-evoked responses of neurons in both regions of LISW-treated rats.
The trigeminal brainstem nuclear complex (TBNC) displays several organizational features that have no spinal equivalent. 10 One prominent feature is the uneven distribution of trigeminal nerve terminals that supply ophthalmic structures such as the meninges and the ocular surface in two spatially discrete regions of the TBNC, the ventrolateral pole of the V/Vc transition and the lateral Vc/C1 junction region. 11 –13 This pattern is further supported by evidence that stimulation of the dura, 32 the ocular surface, 14,15 or exposure to bright light 18 induces a significant increase in the expression of the neuronal activity marker c-fos in both regions.
These results suggest that neurons in the Vi/Vc transition and Vc/C1 regions play a significant role in processing convergent sensory input from the ocular surface and the meninges. This is supported by neurophysiological evidence that dural-responsive neurons in the caudal portions of the TBNC also are activated by stimulation of the ocular surface or periorbital tissues 16,19,20,33 Interestingly, in many of those studies, dural-responsive neurons were recorded mainly in deeper laminae at the mid-Vc or Vc/C1 region, whereas expression of Fos protein was largely restricted to neurons in superficial laminae at the Vc/C1 region after corneal or meningeal stimulation.
In the present study, corneal- or dural-responsive neurons were recorded in the superficial laminae (I–III) at the Vc/C1 junction or in the ventrolateral pole of the Vi/Vc transition and matched well the recording sites of earlier studies for corneal-evoked activity. 31,34 We cannot exclude, however, that neurons in deeper laminae of the Vc/C1 region also contribute to control of vascular functions in headache 35 or after mTBI, because we did not assess the responses of neurons from deeper laminae or determine the effects of inhibition of neurons restricted to deep laminae at Vc/C1.
The pathophysiological mechanisms that mediate the primary and secondary effects of mTBI are complex and likely involve intracranial and extracranial inflammation. 36 The LISW caused a significant increase in Evans blue dye in meningeal samples collected 2 and 10 days after irradiation with no visible signs of cortical damage (data not shown). Previous closed head models of mTBI also have reported persistent behavioral deficits after a single bout of cranial impact with no evidence of cortical lesion by conventional imaging; however, use of positron emission tomography (PET) of radiolabeled markers did find elevated levels of markers for neuroinflammation. 37 Similarly, in earlier studies we determined that LISW produced no evidence of tissue contusion or cortical bleeding but did observe marked changes in cerebral vascular function. 27 These results suggest that subtle changes in the brain microenvironment are sufficient to alter nociceptive behavior after mTBI.
Indeed, closed head models for mTBI have reported significant increases in cortical mast cell degranulation 38 and in neurotransmitter levels in the trigeminal brainstem. 8,9 Moreover, microbleeds that occur after mechanical impact in mTBI can induce long-term changes in neuronal appearance with no evidence of neurodegeneration. 39 These results underscore the need to monitor trigeminal neural activity because anatomical correlates alone likely cannot explain the mechanisms that underlie the behavioral changes after mTBI.
Although PTH may be the most debilitating symptom of mTBI, 40 –42 it is often accompanied by other adverse symptoms such as increased mechanical sensitivity of periocular tissues and light aversion. 43 Cutaneous allodynia is a common feature of several classes of headaches 44,45 and has been used as a diagnostic tool to predict headache chronification. 46 Periorbital or facial skin allodynia was increased by 1–2 days after injury in closed head animal models for mTBI 47 –49 and was consistent with the increase by two days in eye wiping behavior that persisted for 7–10 days after LISW. Meningeal inflammation also was increased by two days and remained elevated at 10 days post-LISW. Other closed head models for mTBI found increased degranulation of dural mast cells by three days after concussive impact. 38 Thus, the time course for meningeal inflammation and for periocular allodynia after LISW was similar to that of other closed head models and strengthened the view that LISW is a valid model for mTBI.
Chronic pain requires central neural mechanisms and is driven by a variable degree of peripheral sensory input. 50,51 Meningeal and corneal afferent neurons share several properties such as a high expression of the transient receptor potential vanilloid 1, 52 –54 a receptor molecule that is required for periorbital allodynia after mTBI. 55 Both meningeal 56,57 and corneal afferent neurons 58,59 become sensitized to mechanical stimuli after localized inflammation of their respective receptive field areas. Localized inflammation of the meninges 19,33 or the ocular surface 29,60 sensitize TBNC neurons and are consistent with the present results of enhanced sensitivity of Vi/Vc and Vc/C1 neurons to ocular surface application of hypertonic saline after LISW. Collectively, these results indicate that meningeal and corneal trigeminal pathways share several properties, and their coincident inputs may be required for mTBI-induced hyperalgesia.
Abnormal sensitivity to light, often referred to as photophobia, is a common symptom in many neuro-ophthalmic pain conditions including mTBI 5,24,61,62 Rats exposed to LISW displayed a marked reduction in the time spent in the light versus the dark chamber, while light-evoked responses by neurons at the Vi/Vc transition and at the Vc/C1 region were significantly enhanced at 10 days after LISW. Photophobia develops rapidly after mTBI and only begins to diminish by one week, 63 which supports our rationale for the time course for assessing behavior and neural responses to light after LISW.
Although photophobia is a common symptom for many diverse conditions that may involve multiple mechanisms, 37,62 several features of light sensitivity may be shared across diverse conditions. First, light must be transduced by photoreceptors in the retina or by non-image-forming cells (i.e., intrinsically photosensitive ganglion cells) in the eye. 64 –66 Second, light information must be relayed centrally to converge onto nociceptive neurons at sites along trigeminal pain pathways that project to sensory cortex. 67,68 Third, light information also must be relayed centrally to activate the autonomic nervous system. 69,70
The present study focused on: (i) the response properties of convergent TBNC neurons that received input from ocular mechanical and light stimulation, and (ii) the effects of altered autonomic function on convergent neuron activity and, conversely, the effects of inhibition of regions with convergent neuron activity on meningeal blood flow. Convergence at the brainstem level was confirmed by the finding of increased light-evoked activity by cornea-responsive neurons in the Vi/Vc transition and in the Vc/C1 regions after LISW. Although the magnitude of the light-evoked responses was numerically greater for Vi/Vc neurons, this difference was not statistically significant (Fig. 6C, p < 0.1).
Previously, we reported that cornea-responsive neurons at the Vi/Vc and Vc/C1 regions projected to regions of the posterior thalamus 71 similar to the location of light/nociceptive convergent neurons in the posterior thalamus reported by Noseda and coworkers. 68 Thus, it cannot be excluded that convergence occurs at multiple sites within trigeminal pain pathways to induce photophobia and is supported by clinical 67,72 and experimental studies. 73 Indeed, the sensory thresholds for light and mechanical allodynia are reportedly well correlated in patients with PTH and, conversely, sensory thresholds may predict the frequency of PTH. 43
Autonomic dysfunction is a common feature of mTBI, contributes to symptoms, and is correlated with the extent of brain injury. 74 –76 The role of autonomic function in mediating acute changes in behavior and neural responses to light in mTBI remains uncertain, however. Photosensitive neurons in the eye project directly to several brain areas such as the olivary pretectal nucleus that are critical for control of autonomic function. 69,70 Activation of sympathetic and parasympathetic nerves may contribute to photophobia by altering meningeal blood flow 77 or by releasing neuroinflammatory agents near sensory endings that may alter blood and excite meningeal afferents (Fig. 9). 78 Indeed, meningeal afferent sensory neurons displayed increased activity after local application of vasodilator agents 79 and reductions in activity after application of vasoconstrictor agents. 80

Possible pathways for throbbing pain triggered by bright light after laser-induced shock wave 9LISW). Key and abbreviations: Dotted line, non-visual pathways activated by light stimuli; green line, parasympathetic pathways that induce vasodilation in the eye and inflamed meninges; orange line, trigeminal pathway activated by vasodilation in the eye and meninges; blue line, trigeminal–parasympathetic pathway; black line, ascending sensory pain pathway; PPG, pterygopalatine ganglion; SSN, superior salivary nucleus; Thal., sensory thalamus; TG, trigeminal ganglion; TBNC, caudal portion of trigeminal spinal nucleus. Color image is available online.
Clinical 81 –84 and experimental studies 85 also support a critical role for autonomic involvement in photophobia. Earlier, we reported that light-evoked activity of cornea-responsive neurons at the Vc/C1 region 26 and at the Vi/Vc transition 23 was markedly reduced by intraocular administration of vasoactive agents. In the present study, dural application of phenylephrine greatly reduced the light-evoked responses of cornea-responsive neurons at the Vi/Vc and Vc/C1 regions after LISW treatment, whereas local synaptic blockade of the Vi/Vc transition, but not the Vc/C1 region, by CoCl2 prevented light-evoked increases in meningeal blood flow. The reason for this apparent inconsistency is not certain, although it does support the view that neurons in each region serve different aspects of trigeminal somato-autonomic function. 23,29,34,71 The present results reveal that neurons in the Vi/Vc transition and at the Vc/C1 region likely also play different roles in control of vascular function and symptoms after mTBI (Fig 9).
Transparency, Rigor and Reproducibility Summary
The experimental protocols were approved by the Committee on Research Facilities for Laboratory Animal Science, National Defense Medical College (Japan) and conformed to the established guidelines set by The National Institutes of Health for the care and use of laboratory animals.
Statistical analyses of all data were performed using GraphPad Prism v. 9 software as detailed above. A sample size of 6-8 subjects per group (excluding phenylephrine drug applications) was planned based on an expected effect size of 1.5 calculated to yield 80% power to detect a significant difference using 2-way ANOVA corrected for repeated measures on 1 factor when appropriate. A p-value of <0.05 was considered significant.
For experiments with large differences between groups and little variation in individual data, sample sizes of 3-6 animals per group were used to reduce the number of animals.
A p-value of <0.05 was considered significant.
All 80 shockwave-exposed, mild head trauma model rats were used in the experiment.
Handling of rats was performed by team members who were aware of relevant characteristics of the subjects. Shock wave irradiation was performed by a team member, and shock wave irradiation was performed by a team member, whereas experiments and data analysis were performed partially blinded by AT.
All equipment and analytical reagents used to perform experimental manipulations and measurements are widely available (described in Materials and Methods).
Replication by the study group is ongoing.
Footnotes
Authors' Contributions
A.T, S.S, D.A.B, and Y.M were responsible for study concept and design. A.T, H.O and S.K were responsible for data acquisition and analysis. A.T and D.A.B were responsible for drafting major portions of the text and figures.
Funding Information
This work was supported by JSPS KAKENHI (Grant Number 25462908) and a grant from the National Institutes of Health USA (EY028143) to DAB.
Author Disclosure Statement
No competing financial interests exist.