
Abstract
Children of parents with traumatic brain injury (TBI) are more likely to develop psychiatric disorders. This association is usually attributed to TBI-induced changes in parents' personality and families' social environment. We tested the hypothesis that offspring of young adult male rats with TBI develop neurodevelopmental abnormalities in the absence of direct social contact with sires. Male Sprague-Dawley rats (F0 generation) in the TBI group underwent moderate TBI via a midline fluid percussion injury that involved craniectomy under sevoflurane (SEVO) anesthesia for 40 min on post-natal Day 60 (P60), while F0 rats in the control group were placed in a new cage, one per cage, for the equivalent time duration. A subset of F0 rats was sacrificed on P66 to assess acute changes in hypothalamic-pituitary-adrenal (HPA) axis and inflammation markers. The remaining F0 males were mated with naive females on P90 to generate offspring (F1 generation). The F0 males and F1 males and females were sequentially evaluated in the elevated plus maze, for pre-pulse inhibition of acoustic startle, in the Morris water maze, and for resting and stress levels of serum corticosterone starting on ∼P105 (F0) and ∼P60 (F1), followed by tissue collection for further analyses. Acutely, the F0 TBI males had messenger RNA (mRNA) transcripts altered to support an increased hypothalamic and hippocampal Na+-K+-Cl- (Slc12a2) Cl- importer / K+-2Cl- (Slc12a5) Cl- exporter ratio and decreased hippocampal glucocorticoid receptors (Nr3c1), as well as increased serum levels of corticosterone, interleukin-1β (IL-1β), and biomarkers of activated hippocampal microglia and astrocytes. Long-term, F0 TBI rats exhibited increased corticosterone concentrations at rest and under stress, anxiety-like behavior, impaired sensory-motor gating, and impaired spatial memory. These abnormalities were underpinned by reduced mRNA levels of hypothalamic and hippocampal mineralocorticoid receptors (Nr3c2), hippocampal Nr3c1, and hypothalamic brain-derived neurotrophic factor (Bdnf), as well as elevated serum levels of IL-1β, and biomarkers of activated hippocampal microglia and astrocytes. F1 male offspring of TBI sires exhibited abnormalities in all behavioral tests, while their F1 female counterparts had abnormal pre-pulse inhibition responses only. F1 male offspring of TBI sires also had reduced mRNA levels of hippocampal Nr3c1 and Nr3c2, as well as hypothalamic and hippocampal Bdnf, whereas increases in inflammatory markers were more profound in F1 females. These findings suggest that offspring of sires with a history of a moderate TBI that involved craniectomy under SEVO anesthesia for 40 min, develop sex-dependent neurobehavioral abnormalities in the absence of direct social interaction between the sire and the offspring.
Introduction
Traumatic brain injury (TBI) frequently affects younger individuals through accidents, military action, or sports injuries. 1 –5 The complex pathophysiology initiated by TBI-inducing insults involves lasting dysregulation of stress response systems, persistent inflammation, compromised neuronal activity, and impaired cognitive function. 6 –12 Many patients with TBI also develop long-term, secondary neuropsychiatric symptoms characteristic of chronic traumatic encephalopathy, Alzheimer's disease, and/or complications similar to post-traumatic stress disorder (PTSD). 13 –16
An increasing number of studies, predominantly in animal models, provide experimental evidence that excessive stress, inflammation, endocrine-disrupting chemicals, and other environmental factors affect not only the exposed subjects, but also their future offspring. 17,18 In humans, abnormalities in offspring caused by high levels of parental stress have been especially well studied, in part because of unique circumstances, such as war, when large groups are affected by stressors in relatively compact areas and within a defined period of time. 19 -21 Although the exact mechanisms mediating heritable effects of parental stressful experiences remain to be elucidated, both animal and human studies point to an important role of the hypothalamic-pituitary-adrenal (HPA) axis and the major glucocorticoid stress hormone cortisol (corticosterone in rodents) in mediating the heritable effects of parental stressful experiences. 19 –22
Surprisingly, the possibility of heritable effects of parental TBI remains an unexplored field. Although epidemiological studies indicate that children of parents with TBI are more likely to develop psychiatric disorders, suggesting some risk of transmission from parents to offspring, 23 –26 such transmission is often assumed to result from TBI-induced changes in parental personality or family dynamics. However, observations that abnormalities in offspring of parents with TBI are not only more profound in males, but also depend on whether the father or mother was affected by TBI, 26 in addition to the notion that stress and inflammation are important factors in TBI pathophysiology, may indirectly support the existence of “biological” (as opposed to purely social) intergenerational transmission of TBI effects. Given that worldwide millions of young adults experience TBI 1 –12 and that many will have children after TBI-inducing events, it is important to elucidate how parental TBI can affect future offspring. Depending on whether social or biological or both modes of transmission are involved, different preventive strategies may be needed to alleviate adverse neurocognitive outcomes in offspring.
The respective contributions of biological versus social factors to inherited effects of TBI can be challenging to disentangle in humans, because in humans, determining whether and how intergenerational transmission of parental experiences contribute to the etiology of children's abnormalities is confounded by obvious ethical constraints as well as demographic and environmental factors. This issue can be more readily investigated in male rats, as their contribution to offspring development is largely restricted to spermatozoa containing genomic and epigenomic information. Therefore, in this study we tested the hypothesis that offspring conceived by sires with a history of a moderate TBI develop neurobehavioral abnormalities in the absence of direct social interaction between sire and offspring. At the mechanistic level, we focused on changes in levels of inflammatory markers and HPA axis activity as potential mediators of intergenerational effects of paternal TBI. 17 –22,27 For the latter, we measured systemic levels of corticosterone and transcripts for corticoid receptors, the Na+-K+-Cl- (Slc12a2) Cl- importer, and the K+-2Cl- (Slc12a5) Cl- exporter genes in the brain. Both the Slc12a2/Slc12a5 ratio, by determining the inhibitory-excitatory balance of GABA type A receptor (GABAAR) signaling, and corticoid receptors are involved in controlling HPA axis activity through feedback mechanisms. 28,29
Methods
Animals
All experimental procedures were approved by the University of Florida Institutional Animal Care and Use Committee. The study was conducted and data are reported in accordance with the Animal Research: Reporting of In Vivo Experiments (ARRIVE) guidelines. 30 Sprague–Dawley rats (56 males and 36 females [generation F0]) were purchased from Charles River. The F0 naïve female rats were used as breeders only to generate offspring (generation F1). Rats were housed under controlled illumination (12-h light/12-h dark, lights on at 7:00 am) and temperature (23-24°C) with free access to food and water. Within 24 h of delivery, F1 litters were culled to 12 pups. At 21 days, pups were weaned and housed two per cage for the rest of the study. Experimental data in this study are from 92 male and 36 female rats. Three F0 male rats died immediately after TBI induction and were excluded from all analyses.
Treatment groups
Figure 1 shows an overview of the study design. F0 male rats were randomized into two treatment groups (n = 28/group). Investigators were blinded to group assignments. The P60 F0 male rats in the TBI group were subjected to a moderate TBI via a midline fluid percussion injury (mFPI), 11,27 while the control F0 males (the Control group) were placed in a new cage, one per cage, for a time equivalent to the duration of the TBI induction procedure. Note that data from F0 male control rats and F1 male and female offspring of control sires were reported previously, 27 but data from F0 rats in the TBI group and their F1 male and female offspring (which were tested at the same time as rats in the control groups) have not been published previously. A subset of the F0 TBI and Control rats (n = 10/group) was sacrificed on P66 to collect tissue samples to study acute effects. The remaining F0 males (n = 18/group) were mated with naive females on P90 to generate offspring (generation F1).
Study design.
For the mating process, a male/female pair was placed in the same cage for 72 h. Thereafter, the F0 female was housed individually for the remaining gestation and postpartum rearing periods. A cohort of 18 breeding pairs of F0 male and female rats was used to produce F1 offspring (18 F1 rats/group/sex). A given F1 offspring experimental group included one to two rats/sex from a given F1 litter. At the age of 21 days, F1 pups were weaned and housed in sex-matched pairs for the rest of the study. F1 offspring of TBI and Control sires were not exposed to any treatment and were subjected to animal facility rearing only, except for neurobehavioral evaluations. The analysis of the breeding data does not indicate any difference in fertility of sires with moderate TBI when compared with sires in the Control group. Out of 18 breeding pairs per experimental group, one naïve female rat mated with a male from the TBI group and two naïve female rats mated with males from the Control group were infertile. There was also no statistical difference in the number of offspring sired by rats from the TBI and Control groups.
This study tested the hypothesis that offspring conceived by sires with a history of a moderate TBI develop neurobehavioral abnormalities in the absence of direct social interaction between sire and offspring. It is possible that TBI sires could affect F1 offspring through interactions with otherwise naïve F0 dams during the 72-h mating period, or through stress or other signals (via auditory or olfactory modalities) in the colony room; however, the stress or other signals in the colony room would have been the same for F1 offspring of Control and TBI sires, and thus any such remotely-transmitted signals would have had to have affected offspring of TBI sires selectively. Also, all rats were housed in individually ventilated cages, which each received a separate supply of high-efficiency particulate air (HEPA)-filtered air.
The same F0 sires that were used for mating with naïve females to produce F1 offspring were evaluated sequentially in the elevated plus maze (EPM) on P105, 27,31 pre-pulse inhibition (PPI) of the acoustic startle response on P115, 27,31,32 and Morris water maze (MWM) between P125 and P130. 27,33,34 Ten days after completion of the behavioral tests, stress-induced release of corticosterone was studied after exposing the same F0 males to physical restraint for 30 min, as previously described. 27 The same F1 male and female offspring were sequentially evaluated in the EPM on P60, PPI of the acoustic startle response on P70, and MWM between P90 and P95. Blood samples were collected using the “tail clip” method 30 min after completion of the PPI test to measure stress serum levels of corticosterone, as previously described. 27 Ten days after completing the in vivo studies, the same F0 and F1 rats were anesthetized and sacrificed through decapitation to collect trunk blood and brain tissue samples for further analyses (Fig. 1).
Midline fluid percussion injury model of traumatic brain injury
The FPI device (Model 01-B; Custom Design & Fabrication, Inc., VA) was used to induce moderate TBI via mFPI, as previously described. 27 To provide a uniform sevoflurane (SEVO) anesthesia duration and depth to all experimental animals, anesthesia in all rats was induced with 6% SEVO for 3 min in an anesthesia chamber and then a rat was placed in a stereotaxic frame with a continuous supply of 3% SEVO for 37 min via a nose cone. SEVO was delivered in a mixture with ∼50% O2 in air (1.5 L/min). The rats' rectal temperature during surgery was monitored and maintained at ∼+37.0°C using a homeothermic monitoring system (50-7220F; Harvard Apparatus, St. Laurent, Quebec, Canada). Following a ∼15-mm midline incision in the scalp and fascia removal from the skull, a craniectomy was centrally made over the sagittal suture between bregma and lambda by using a 4.8 mm trephine (Integra Miltex, Princeton, NJ). The dura was kept intact. The female hub of a 20-gauge Luer-Loc needle was placed over the craniectomy and secured first with cyanoacrylate gel and then with methyl-methacrylate dental cement (Henry Schein, Melville, NY). The dura was visually inspected to ensure that it was intact and no debris were present. The inducer was placed over the hub, filled with normal saline, and attached to the male end of the fluid percussion injury device while avoiding air bubbles at the junction. A pressure wave averaging 2.0 ± 0.2 atm was delivered by releasing the pendulum onto the fluid-filled cylinder. The righting reflex time (the time from the initial impact until the rat spontaneously rights itself from a supine position) was 8.97 ± 1.82 min. The injury site was inspected for bleeding, herniation, hematomas, and dura integrity before the wound was sutured.
The elevated plus maze test
The EPM studies were performed using an elevated plus maze apparatus and BIO-EPM 3C video-tracking software (EB Instruments, Pinellas Park, FL, USA) during the light phase of the dark–light cycle, as previously described. 27,31 The maze consists of two opposing open (50 × 10 × 0.5 cm) and two enclosed (50 × 10 × 45 cm) arms elevated 75 cm above the floor, with a 0.5-cm edge on the open arms. Animals were placed in the center of the maze facing an open arm and were allowed to explore the maze for 5 min. The percentage of time spent in the open arms, the percentage of entries into the open arms, and the total distance traveled during the test were compared. The time spent in open arms and number of entries to open arms are calculated and presented as the percentage of the total time spent in the open and closed arms and the total number of entries to either open or closed arms, respectively.
Measurements of PPI of startle
We performed PPI of acoustic startle tests in rats using the SR-Lab startle apparatus (San Diego Instruments, San Diego, CA, USA), as previously described. 27,31,32 Testing occurred during the light phase of the dark–light cycle. The entire test for a given animal lasted 28 min. The animal enclosure (20 cm in length, 9 cm in interior diameter) permitted the animal to turn around in the enclosure. At the beginning of every testing session, each animal was placed in the cylindrical animal enclosure and exposed to a 70-dB white noise background for a 5-min acclimation period. The acclimation period was followed by a test session consisting of five types of trials: a 120-dB 40-msec pulse only; a 120-dB 40-msec pulse preceded by a 20-msec pre-pulse at intensities of 73 dB, 76 dB and 82 dB (i.e., 3 dB, 6 dB, and 12 dB above the background noise of 70 dB); and a no-stimulus trial of background noise. The delay between the onset of the pre-pulse and the onset of the pulse was 100 msec. The trials were presented in pseudorandom order, with variable inter-trial intervals averaging 15 sec. The first four trials and the last three trials consisted of 120-dB pulse-only trials. All five types of trials were presented eight times, each in pseudorandom order after the first four and before the last three pulse-only trials. The percent pre-pulse inhibition for each pre-pulse intensity was calculated using the following formula: % pre-pulse inhibition = 100 × [(pulse alone) – (pre-pulse + pulse)]/pulse alone. Data were collected as VAverage amplitude (the average voltage during the response window).
Morris water maze test
To evaluate spatial learning and memory in rats, we used the MWM behavioral assay, as previously described. 27,33,34 The MWM apparatus consisted of a circular tank (183 cm in diameter, 58 cm in height). The maze was filled with water (23 ± 1°C) made opaque with the addition of nontoxic paint (Blick Art Materials, Highland Park, IL, USA). The pool was surrounded with black curtains on which were affixed large white geometric shapes (extramaze cues). The pool was divided into four equal quadrants, labeled NW, NE, SE and SW (NW—northwest; NE—northeast; SE—southeast; SW—southwest). A 12-cm diameter platform (the escape platform) was submerged 2 cm below the surface of the water at a constant location in one of the quadrants. Data were recorded using a computer-based video tracking system (ANY-maze, Stoelting Co., Wood Dale, IL, USA). Rats received four trials of daily training over 5 consecutive days.
On each trial, rats were placed into the water facing the wall of the maze at one of the four quadrants. The start positions were varied in a pseudorandom manner. Once in the water, rats were allowed to swim until they found the hidden platform or until 60 sec elapsed, at which time the rats were gently guided to the platform. Rats remained on the platform for 15 sec before the next trial. Twenty-four hours later, the spatial memory test was performed. The spatial memory test lasted 60 sec, during which time the escape platform was removed. The rat was placed in the pool in the contralateral quadrant (the entrance quadrant, SE) relative to the original location of the escape platform (the target quadrant, NW). The time spent in each quadrant and the number of times the rat crossed the previous location of the escape platform were recorded.
Stress-induced serum corticosterone levels
To measure stress-induced serum levels of corticosterone, F0 male rats were physically restrained for 30 min. Physical restraint was administered using rodent holders (Kent Scientific Corporation, Torrington, CT, USA). Blood samples (∼300 μL) were collected immediately before physical restraint and 10, 60, and 120 min after the restraint. Blood sampling was done using the “tail clip” method, as previously described. 27 Specifically, the distal 0.5 mm of the tail was removed using a sterile scalpel blade and blood was allowed to drain directly into a microcentrifuge tube. The hemorrhaging was stopped by applying gentle digital pressure to the wound for ∼1 min. To measure stress-induced serum levels of corticosterone in F1 offspring, the tail blood samples were collected 30 min after completion of the PPI of acoustic startle test, using the same technique described above.
Tissue collection
Rats were anesthetized with SEVO and decapitated. The trunk blood samples were collected and centrifuged at 4°C, 1000 g for 15 min, and then kept at -80°C for resting corticosterone and proinflammatory cytokine interleukin-1β (IL-1β) assays. The hypothalamus was isolated by making an anterior cut at the level of the optic chiasm, a posterior coronal section anterior to the mammillary bodies, two sagittal cuts parallel to the lateral ventricles, and a dorsal horizontal cut at the level of the anterior commissure. 23 -25 The hippocampus was isolated from the respective blocks. All tissue samples were placed in vials filled with RNAlater solution (Invitrogen, Carlsbad, CA).
Measurements of serum concentrations of corticosterone and IL-1β
Enzyme-linked immunosorbent assay (ELISA) kits were used to measure serum concentrations of corticosterone (501320; Cayman Chemical Company, Ann Arbor, MI) and IL-1β (BMS630; Invitrogen, Carlsbad, CA) by following the manufacturers' instructions.
Measurement of messenger RNA levels
The messenger RNA (mRNA) levels in the hippocampus and hypothalamus were analyzed via reverse transcription-polymerase chain reaction (qRT-PCR) in a StepOnePlus™ Real-Time PCR System (Applied Biosystems, Foster City, CA), as previously described]. 27 RNA was extracted from the samples using a RNeasy Plus Kit (Qiagen, Valencia, CA), reverse transcribed with a high-capacity cDNA reverse transcription kit (Bio-Rad Laboratories, Hercules, CA), and then analyzed via qRT-PCR. Taqman probes specific for Slc12a2 (Rn00582505_m1), Slc12a5 (Rn00592624_m1), corticotropin-releasing hormone (Crh, Rn01462137_m1), glucocorticoid (Nr3c1, Rn00561369_m1) and mineralocorticoid (Nr3c2, Rn00565562_m1) receptors, and brain-derived neurotrophic factor (Bdnf, Rn02531967_s1) were obtained from Applied Biosystems. Data were normalized to glyceraldehyde-3-phosphate dehydrogenase (Gapdh) mRNA (Rn01775763_g1). Gene expression was calculated using the ΔΔCT method and data are presented as relative fold change from that of control animals.
Immunohistochemistry
Rats were anesthetized with SEVO and transcardially perfused with saline, followed by 4% paraformaldehyde (PFA) in 0.1 M phosphate buffer. The brains were collected and fixed in the 4% PFA solution overnight and then dehydrated in 30% sucrose solution in phosphate-buffered saline (PBS) at 4°C for 2 days. The brains were cut into 40-μm-thick coronal sections using a vibratome (VT1000 S; Leica, Wetzlar, Germany). After blocking with 10% normal goat serum for 1 h at room temperature, the slices were incubated with the primary antibodies rabbit anti-Iba1 (ionized calcium binding adaptor 1, a microglia/macrophage-specific protein marker; 1:500; Wako Pure Chemical Industries, Osaka, Japan) and mouse anti-GFAP (glial fibrillary acidic protein, an astrocyte specific protein marker; 1:500; Invitrogen) in 1% bovine serum albumin at 4°C overnight. After washing with PBS for 10 min × 3 times, the slices were exposed to the secondary antibodies Alexafluor 549 goat anti-rabbit and Alexafluor 488 goat anti-mouse (Invitrogen, Carlsbad, CA) for 1 h at room temperature. After washing out the secondary antibodies, the sections were incubated with 4´,6-diamidine-2´-phenylindole dihydrochloride (DAPI) for nuclear staining.
A fluorescence microscope (BZ-X810E, Keyence, Osaka, Japan) was used to capture the images. Six sections of hippocampus per rat were analyzed using ImageJ software (National Institutes of Health, Bethesda, MD). Three non-overlapping fields of each section in the hippocampal Cornu Ammonis 1 (CA1) area were randomly acquired. The immunoreactivity of a protein was quantified by calculating the percentage area with positive staining relative to the total area of the imaged field. 24,25 All quantitative analyses were performed by an experimenter blinded to the group of each sample.
Statistical analysis
The primary outcomes in this study were the neuroendocrine and behavioral changes in F0 sires and in their F1 offspring. All other outcome measurements were secondary outcomes. Values are reported as mean ± standard error of the mean (SEM). Statistical analyses were carried out on raw data using GraphPad Prism 9 (GraphPad Software, Boston, MA), SigmaPlot 14.0 software (Systat Software, Inc., San Jose, CA) and SPSS v27 (IBM Corp., Armonk, NY). Boxplots were used to identify outliers. No outliers were detected that were not in the plausible range of values for the outcomes; therefore, all data were maintained in analyses. To analyze F0 data for acute Slc12a2, Slc12a5, Slc12a2/Slc12a5 ratio, Crh, Nr3c1, and Nr3c2 mRNA levels, serum corticosterone and IL-1β levels, hippocampal Iba1 and GFAP expression, long term EPM, and number of crossings over the former platform location during the MWM probe test, differences in long term gene expression, resting corticosterone and total corticosterone concentrations during the restraint, IL-1β levels, and hippocampal Iba1 and GFAP expression, t-tests were used.
A two-way repeated measures analysis of variance (ANOVA), with experimental groups and time as the independent variables, was used to analyze changes in serum corticosterone levels before and at three time-points after the restraint. A two-way repeated measures ANOVA was used to analyze the F0 PPI data, with the treatment and pre-pulse intensity as independent variables. A two-way repeated measures ANOVA with experimental groups and days of training as the independent variables was used to analyze latencies to reach the escape platform during MWM training in F0 rats. A two-way repeated measures ANOVA with experimental groups and quadrants as the independent variables was used to analyze time spent in each quadrant during the MWM test in F0 rats. A two-way ANOVA with treatment and sex as the independent variables were used to assess F1 data for changes in EPM, MWM platform location crossing times, changes in gene expression, serum corticosterone levels at rest and after the PPI test, changes in serum IL-1β levels and hippocampal Iba1 and GFAP expression.
For F1 PPI, MWM escape latency and time in each quadrant, linear mixed models for repeated measures were used, with PPI intensity, days of training, and quadrant modeled as repeated measures, respectively. These analyses account for within-subject corrections across repeated measurements. The models also included treatment and sex as main effects, as well as interaction terms. Multiple pairwise comparisons were done with the Holm-Sidak method. A p ≤ 0.05 was considered significant.
Results
Acute effects of TBI in F0 male sires
TBI in young adult male rats led to acute alterations in transcripts of Cl- transporters in the hippocampus and hypothalamus (Fig. 2A, 2B). The F0 male rats from the TBI group had increased Slc12a2 mRNA levels and decreased Slc12a5 mRNA levels (Fig. 2A, 2B) in both hippocampus and hypothalamus (hippocampal Slc12a2 mRNA: t(10) = -2.386; hippocampal Slc12a5 mRNA: t(10) = 2.5; hypothalamic Slc12a2 mRNA: t(10) = -4.706; hypothalamic Slc12a5 mRNA: t(10) = 4.328). The resulting hippocampal and hypothalamic Slc12a2/Slc12a5 mRNA ratios were increased in TBI F0 sires (hippocampal Slc12a2/Slc12a5 mRNA ratio: t(10) = -3.648; hypothalamic Slc12a2/Slc12a5 mRNA ratio: t(10) = -9.588). The F0 TBI rats had lower hippocampal mRNA levels of Nr3c1 (t(10) = 2.626), but not Crh or Nr3c2. There were no changes in the mRNA levels of hypothalamic Crh, Nr3c1 and Nr3c2 (Fig. 2B). The TBI F0 rats had increased serum levels of corticosterone (t(10) = -2.630; Fig. 2C), the proinflammatory cytokine IL-1β (t(10) = -13.446; Fig. 2D) and hippocampal levels of GFAP (t(6) = -9.103; Fig. 2E, 2G) and Iba1 (t(6) = -5.371; Fig. 2F, 2G).

Acute effects of traumatic brain injury (TBI) in F0 male rats.
Long-term effects of TBI in F0 sires
In the EPM test, the F0 TBI rats spent less time in the open arms (t(34) = 3.406; Fig. 3A), and made fewer open arm entries (t(34) = 3.103; Fig. 3B) than controls, but did not differ from their control counterparts in total distance traveled (t(34) = 1.693; Fig. 3C). In addition, a two-way repeated measures ANOVA revealed a significant effect of TBI on PPI of startle in F0 rats (F(1,68) = 5.312; P = 0.027; Fig. 3D). Multiple pairwise comparisons indicated that TBI led to impaired PPI of startle responses in F0 male rats at pre-pulse intensities of 3 dB and 12 dB (Fig. 3D). The MWM test showed no between-subjects effect of TBI on escape latencies across the 5-day training period (F(1,136) = 0.0949, p = 0.760), although there was a significant within-subjects effect of day of training such that latencies decreased across the course of training (F(4,136) = 48.484, p < 0.001; Fig. 3E). During the probe test, analysis of the time spent in each quadrant revealed no main effect of treatment (F(1,102) = 2.457; p = 0.126; Fig. 3F). There was a significant main effect of quadrant (consistent with rats' preference for the target over the other quadrants; F(3,102) = 39.394; p < 0.001; Fig. 3F). Most importantly, there was a significant treatment × quadrant interaction (F(3,102) = 3.866; p = 0.012; Fig. 3F), and post-hoc tests indicated that the F0 rats from the TBI group spent significantly less time in the target quadrant when compared with the Control animals. The TBI rats also made fewer crossings over the escape platform location (t(34) = 3.294; Fig. 3G).
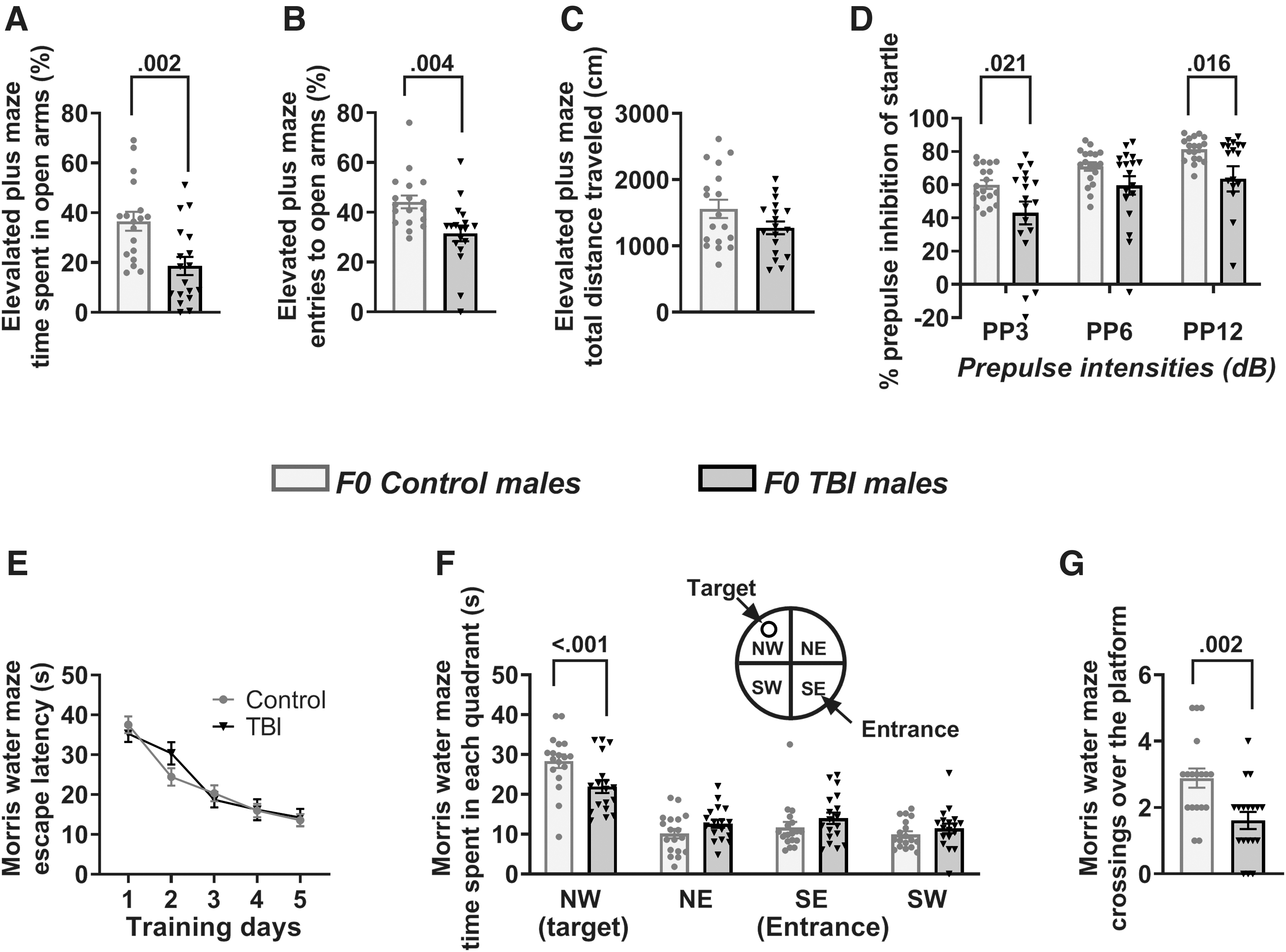
Long-term behavioral effects of traumatic brain injury (TBI) in F0 male rats.
The F0 male rats from the TBI group had decreased Nr3c2 mRNA levels in the hippocampus (t(10) = 5.528; Fig. 4A) and hypothalamus (t(10) = 2.264; Fig. 4B), and decreased Nr3c1 mRNA levels in the hippocampus (t(10) = 3.440; Fig. 4A). The hippocampal and hypothalamic mRNA levels for Crh were similar in the Control and TBI F0 rats (Fig. 4A, 4B). The F0 TBI rats also had reduced hypothalamic mRNA levels of Bdnf (t(9) = 3.403; Fig. 4B). The effect of TBI on resting serum levels of corticosterone (t(13) = -3.856; Fig. 4C), IL-1β (t(14) = -4.295; Fig. 4F), and hippocampal levels of GFAP (t(6) = -4.226; Fig. 4G, 4I) and Iba1 (t(6) = -3.036; Fig. 4H, 4I) could still be detected >2 months after TBI. To assess long-term effects of TBI on systemic levels of corticosterone after exposure to an acute stressor, serum corticosterone was measured in blood samples collected prior to restraint and 10, 60, and 120 min after restraint. F0 TBI rats had significantly higher total corticosterone levels when compared with their control counterparts (measured as AUCg; t(13) = -3.006; Fig. 4E). This increase in corticosterone was due to higher levels of corticosterone at 10 min after the restraint (Fig. 4D).
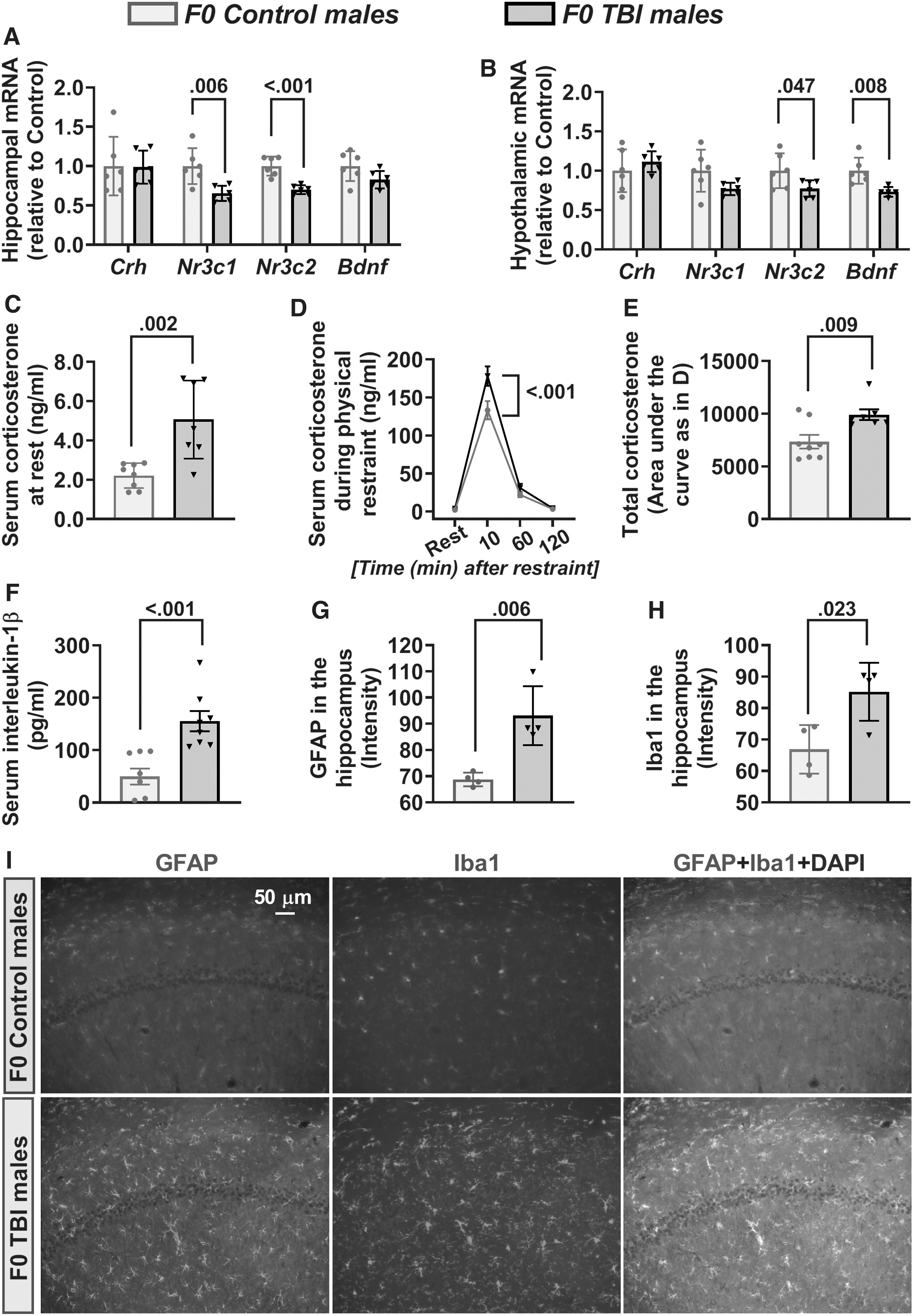
Long-term alterations in hypothalamic-pituitary-adrenal axis activity, inflammation, and brain-derived neurotrophic factor (Bdnf) messenger RNA (mRNA) levels in F0 sires.
Effects of paternal TBI in F1 offspring
In the EPM test, there was a main effect of paternal TBI (F(1,68) = 8.158, p = 0.006) and sex (F(1,68) = 8.693, p = 0.004), but no paternal TBI × sex interaction (F(1,68) = 0.953, p = 0.332), on the percentage of time that F1 offspring spent in the open arms (Fig. 5A). There was also a main effect of paternal TBI (F(1,68) = 10.779, p = 0.002), but no effect of sex (F(1,68) = 3.041, p = 0.086) or paternal TBI × sex interaction (F(1,68) = 2.767, p = 0.101), on the number of entries that F1 rats made into the open arms of the EPM (Fig. 5B). Post hoc pairwise comparisons on both measures showed that F1 male offspring of TBI sires spent less time in the open arms (Fig. 5A) and made fewer entries into the open arms (Fig. 5B) when compared with F1 male offspring of control sires and to F1 females from the TBI group. In contrast to the open arm measures, an analysis of total distance traveled in the EPM revealed a main effect of sex (F(1,68) = 9.903, p = 0.002), but no main effect of paternal TBI (F(1,68) = 1.5, p = 0.225) or paternal TBI × sex interaction (F(1,68) = 0.517, p = 0.474; Fig. 5C).
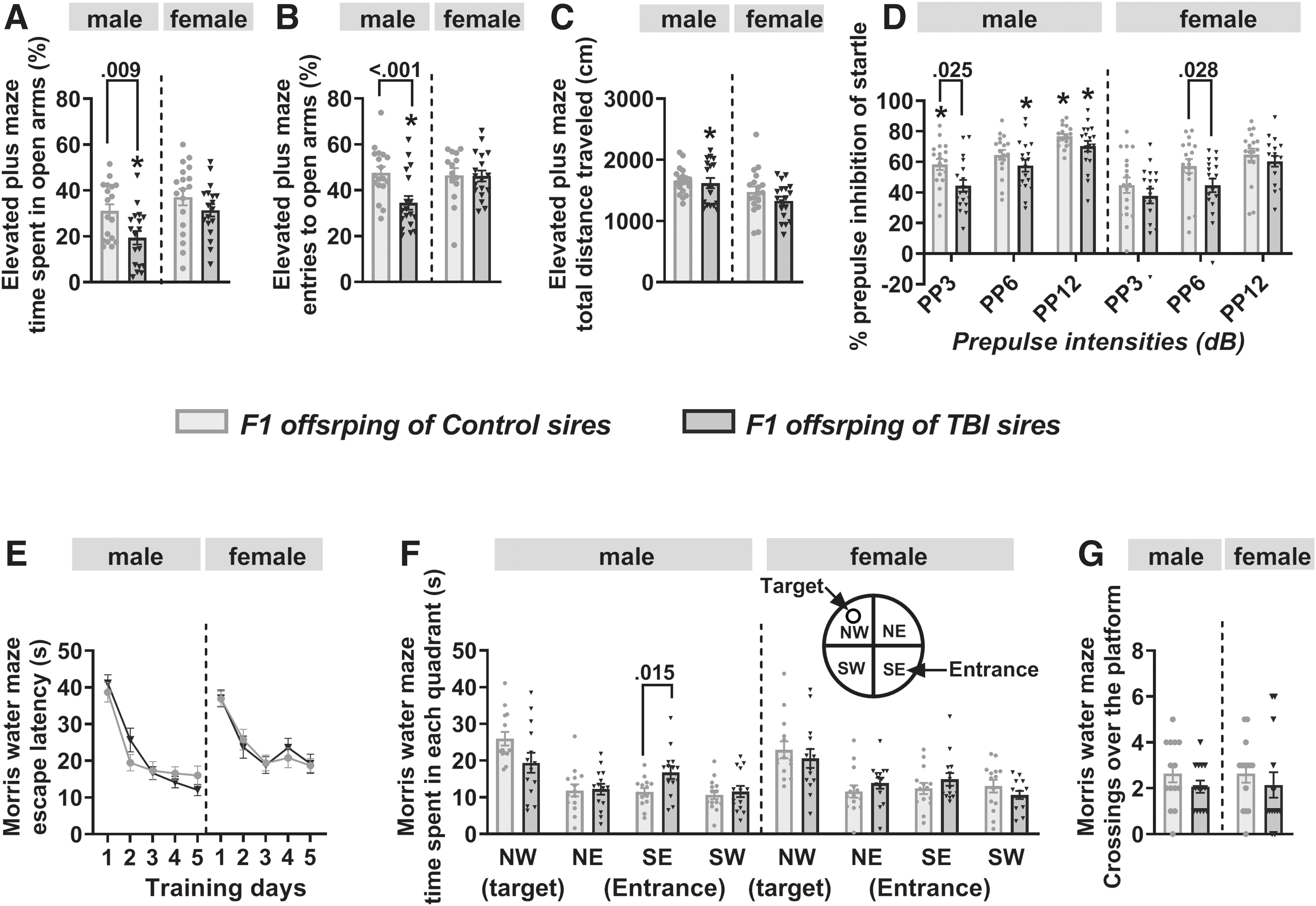
Behavioral effects of paternal traumatic brain injury (TBI) in F1 offspring.
There were overall effects of sex (F(1,204) = 20.719, p < 0.001) and pre-pulse intensity on PPI of startle responses (F(2,204) = 32.857, p < 0.001) and paternal TBI (F(1,204) = 14.302, p < 0.001) in F1 rats, but no paternal TBI × sex (F(1,204) = 0.066, p = 0.798) or pre-pulse intensity × paternal TBI × sex interaction (F(2,204) = 0.57, p = 0.566; Fig. 5D). Multiple pairwise comparisons indicated that when compared with offspring of control sires, F1 males and F1 females of TBI sires exhibited reduced PPI of startle at pre-pulse intensities of 3 dB and 6 dB, respectively (Fig. 5D). Paternal TBI did not affect startle amplitudes in the absence of the pre-pulse in F1 male and F1 female rats.
During training in the MWM, there were significant effects of day (F(4,260) = 60.733, p < 0.001) and sex (F(1,260) = 6.064, p = 0.014; Fig. 5E), but no main effect of paternal TBI (F(1,260) = 0.05, p = 0.824) or interactions between paternal TBI and the other two variables (paternal TBI × day, F(4,260) = 0.385, p = 0.819; paternal TBI × sex, F(1,260) = 0.005, p = 0.942; paternal TBI × day × sex, F(4,260) = 1.166, p = 0.326) on escape latencies across the 5-day training period.
During the probe test, assessment of time spent by F1 male and female offspring in each quadrant of the MWM revealed a main effect of quadrant (F(3,208) = 21.334, p < 0.001) as well as a paternal TBI x quadrant interaction (F(3,208) = 3.407, p = 0.019; Fig. 5F). Subsequent comparisons between groups for each quadrant revealed that, compared with F1 males of control sires, F1 males of TBI sires spent significantly more time in the entrance quadrant and tended to spend less time in the target quadrant, although the reduction in time in the target quadrant was not sufficient to achieve statistical significance (p = 0.053; Fig. 5F). In contrast, assessment of the number of crossings of the platform location revealed no significant effects of paternal TBI (F(1,52) = 1.67, p = 0.202), sex (F(1,52) = 0.007, p = 0.932) or paternal TBI × sex interaction (F(1,52) = 0.007, p = 0.932; Fig. 5G).
Assessment of hippocampal Nr3c1 mRNA revealed a main effect of sex (F(1,20) = 7.423, p = 0.013) and a paternal TBI × sex interaction (F(1,20) = 7.423, p = 0.013), but no main effect of paternal TBI (F(1,20) = 2.177, p = 0.156; Fig. 6A), whereas there was a main effect of paternal TBI (F(1,20) = 5.823, p = 0.026), but no effect of sex (F(1,20) = 0.406, p = 0.531) or paternal TBI × sex interaction (F(1,20) = 0.406, p = 0.531) on hippocampal Nr3c2 mRNA (Fig. 6A). Specifically, F1 male offspring of TBI sires had lower levels of Nr3c1 and Nr3c2 mRNA in the hippocampus when compared with F1 male offspring of Control sires. There was no main effect of paternal TBI, sex, or sex x paternal TBI interaction on hypothalamic Nr3c1 and Nr3c2 transcripts (Fig. 6B); however, there were main effects of sex (hippocampus: F(1,20) = 4.914, p = 0.038; hypothalamus: F(1,20) = 7.971, p = 0.011) and paternal TBI x sex interactions (hippocampus: F(1,20) = 4.914, p = 0.038; hypothalamus: F(1,20) = 7.971, p = 0.011), but no main effect of paternal TBI (hippocampus: F(1,20) = 4.254, p = 0.052; hypothalamus: F(1,20) = 4.239, p = 0.053) on Bdnf mRNA in the hippocampus and hypothalamus of F1 offspring (Fig. 6A, 6B). F1 male offspring of TBI sires had reduced Bdnf mRNA levels when compared with F1 male offspring of Control sires and to the respective F1 females from the TBI group. The latter were not affected by paternal TBI.
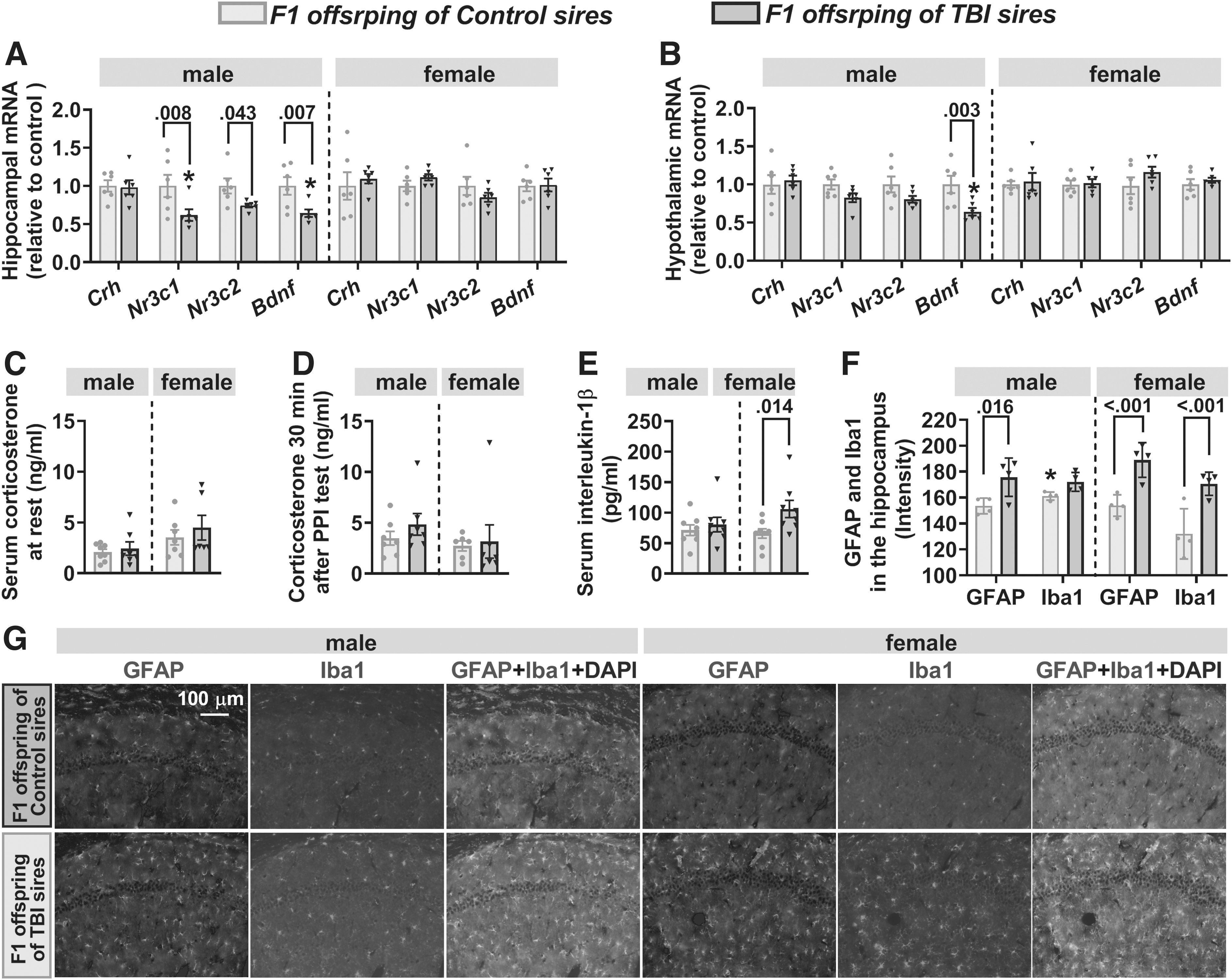
The effects of paternal traumatic brain injury (TBI) on hypothalamic-pituitary-adrenal axis activity, inflammation markers, and the brain-derived neurotrophic factor (Bdnf) messenger RNA (mRNA) levels in adult F1 offspring.
Analysis of resting serum corticosterone levels in F1 rats revealed a significant effect of sex (F(1,24) = 5.748, p = 0.025), but no effect of paternal TBI (F(1,24) = 0.793, p = 0.382) or sex × paternal TBI interaction (F(1,24) = 0.164, p = 0.689; Fig. 6C). In addition (and in contrast to F0 sires, in which TBI animals responded to physical restraint with higher levels of serum corticosterone), serum levels of corticosterone 30 min after the PPI test were not different between F1 offspring of Control and TBI sires within the same sex (Fig. 6D). There was a significant effect of paternal TBI (F(1,28) = 5.133, p = 0.031), but no effect of sex (F(1,28) = 0.852, p = 0.364) or paternal TBI × sex interaction (F(1,28) = 2.024, p = 0.166) on serum levels of IL-1β. Notably, in contrast to the sustained increase in serum IL-1β in F0 TBI sires, F1 males of TBI sires did not differ from F1 males of Control sires, whereas serum levels of IL-1β were increased in F1 female offspring of TBI sires (Fig. 6E). There was a significant effect of paternal TBI (F(1,12) = 26.024, p < 0.001), but no effect of sex (F(1,12) = 1.478, p = 0.247) or paternal TBI × sex interaction (F(1,12) = 1.328, p = 0.272) on hippocampal levels of GFAP (Fig. 6F, 6G). There was significant between-subject effect of paternal TBI (F(1,12) = 18.889, p < 0.001), sex (F(1,12) = 7.225, p = 0.02), and paternal TBI by sex interaction (F(1,12) = 5.89, p = 0.032) on hippocampal levels of Iba1 in F1 offspring (Fig. 6F, 6G). Consistent with the marker of sustained systemic inflammation in F1 females, F1 female offspring of TBI sires had higher hippocampal GFAP and Iba1 levels than F1 females of control sires. In addition, F1 males from TBI sires had higher hippocampal GFAP levels than F1 males from control sires (Fig. 6F, 6G). Levels of hippocampal Iba1 in F1 males were not affected by paternal TBI.
Discussion
The results of this study provide evidence that in young adult male rats, a moderate midline FPI-induced TBI that involved craniectomy under anesthesia with sevoflurane for 40 min leads to neurological abnormalities not only in the exposed sires but also in their future adult offspring, who were subjected to facility rearing only. Importantly, the role of the TBI-affected sires in their offspring's development was limited to conception. As such, the findings of this study provide the first experimental evidence in an animal model that paternal moderate TBI-induced abnormalities can be inherited by offspring via mechanisms that do not involve direct social interactions between the sire and the offspring. The effects of paternal TBI may primarily modulate male-specific mechanisms (processes) in offspring, as F1 female offspring of the TBI-affected sires were less affected in the tests used in this study.
The finding of this rat study of more profound effects of paternal TBI in F1 male offspring is somewhat in line with observations made by the 1987 Finnish Birth Cohort study, which found (after adjusting for covariates) that sons of parents with TBI, especially paternal TBI, exhibited significant abnormalities in terms of any mental disorder, substance use-related disorders, and behavioral and emotional disorders, while in daughters only disorders of psychological development were related to parental TBI. 23 In contrast, substance use among sons in this study was more prevalent when the mother was affected by TBI. 23 It is important to emphasize that in the above-mentioned human study it was not known for how long, or if at all, TBI-affected parents lived with their children, and thus the relative contributions of social versus biological transmission of the observed abnormalities cannot be disentangled. 23
Male offspring developed many, but not all, neurobehavioral abnormalities detected in their TBI-affected fathers. For example, F0 TBI males exhibited acute and persistent dysregulation of the HPA axis, elevated biomarkers of systemic and central inflammation, and long-term behavioral deficiencies. Their F1 male offspring exhibited deficiencies in the same neurobehavioral tests, but their HPA axis activity and biomarkers for systemic inflammation were not different from F1 males fathered by Control sires. The acute stress-like effects (i.e., an increase in serum levels of corticosterone) and complex corticosterone responses to repeated restraint stress were previously reported in young adult male rats that underwent mild or moderate TBI via midline FPI. 11 Endocrinopathies are also increasingly recognized as a serious complication of TBI in human patients. 9,10 We are not aware of animal studies that measured either resting or stress levels of corticosterone in offspring of parents with a history of TBI. Interestingly, similar to findings of this study in rats, the 1987 Finnish Birth Cohort study found that although children of parents with TBI were more likely to develop psychiatric disorders, their reactions to stressful events were not different from reactions to stressful events by children of parents without TBI. 23
Remarkably, qualitatively different intergenerational outcomes were observed when the same TBI model that was tested in this study (paternal midline FPI on P60 after craniectomy under SEVO for 40 min), was combined with repeated prolonged (for 3 h) exposures to SEVO on post-natal days P62, P64 and P66. 27 In this prior study, the male, but not female, offspring of sires subjected to moderate TBI in combination with subsequent repeated exposures to SEVO exhibited heightened levels of inflammatory markers in serum and brain, as well as abnormalities in the HPA axis, including exacerbated corticosterone responses to stress caused by the PPI test. 27 Such qualitatively different intergenerational effects of moderate TBI with 27 and without (current study) repeated prolonged exposure to SEVO are in line with the current understanding of neurotoxic effects of repeated exposure to general anesthetics. Currently there is a wide agreement, supported by clinical and laboratory data, that prolonged repeated exposure to general anesthesia may induce neurocognitive abnormalities in the exposed subjects. 35 -37 In contrast, a single, relatively short (under one hour) exposure to anesthesia is considered safe even in infants. 38 -40 In the current study, all animals were exposed to anesthesia with SEVO for 40 min.
The findings from previous work that the detrimental effects of moderate TBI and subsequent repeated exposure to SEVO interact to induce more profound intergenerational outcomes, 27 suggest that the effects of a moderate TBI and craniectomy under SEVO anesthesia for 40 min may also interact to induce intergenerational neurobehavioral abnormalities. In other words, craniectomy under SEVO anesthesia for 40 min alone may not induce intergenerational neurobehavioral deficits, but may contribute to such deficits when combined with a moderate TBI. Therefore, the use of sham surgery/SEVO for 40 min group as the control, instead of the non-surgery/anesthesia control group as used here, could lead to a misleading conclusion that the observed effects were due to TBI alone, rather than to TBI in combination with surgery (craniectomy) under SEVO anesthesia for 40 min. This issue is difficult to address using the midline FPI model, which requires anesthesia for the surgical component. As such, a more comprehensive understanding of the intergenerational effects of TBI will necessitate the use of different TBI models that do not involve surgery and exposure to general anesthetics. The need for multi-model testing approaches to investigating intergenerational neurobehavioral effects of parental TBI is further supported by the fact that no animal model fully represents the complex pathophysiology of human TBI.
Although the craniectomy and anesthesia that constitute the FPI-induced TBI procedure narrow the applicability of the findings of this study, the findings are still not only novel but also translationally important, as many TBI patients require general anesthesia and surgery to treat injuries occurring at the time of TBI. 41 -43 Importantly, the midline FPI-induced TBI closely models diffuse axonal injury, a pathophysiological hallmark of the diffuse TBI that frequently occurs following injury in young adults during military, athletic, and other activities, and which is frequently accompanied by the need for surgical interventions. 11 Therefore, in future studies it will be important to assess how surgery/anesthesia duration/complexity affect intergenerational neurocognitive outcomes of TBI.
The involvement of inflammation/stress-like initiating mechanisms of the intergenerational effects of TBI and surgery under SEVO anesthesia are supported not only by heightened levels of acute and long-term inflammatory markers and corticosterone responses to stress in F0 rats, but also by changes in hippocampal and hypothalamic mRNA levels of the Slc12a2/Slc12a5 ratio and hippocampal Nr3c1. Both reduced expression of Nr3c1 and resulting impairment of corticoid receptor-mediated negative feedback modulation of the HPA axis activity, 28 and increased Slc12a2/Slc12a5 ratio and resulting impairment of GABAAR-mediated inhibitory control of the HPA axis, 29 may contribute to the stress- and inflammation-like effects of TBI/surgery/SEVO observed in this study. This is also consistent with the roles of an abnormal NKCC1/KCC2 ratio, impaired GABAAR signaling, stress, and inflammation in the pathophysiology of TBI. 7 –11,44 –47
Dysregulated stress response systems and inflammation may be involved in induction of somatic and germ cell effects in the TBI-exposed sires, while neurobehavioral abnormalities in the offspring may be initiated through paternal germ cell-mediated changes in the offspring's physiology, such as changes in levels of corticoid receptors and Bdnf. Dysregulation of BDNF-driven processes is a common component of the pathophysiology of TBI and other neurological disorders, while treatments with BDNF or BDNF mimetics have demonstrated positive outcomes in animal models of such disorders. 48 Abnormally-expressed corticoid receptors have also been associated with anxiety, depression, PTSD, and many other stress-related psychiatric disorders. 49 Further, abnormal regulation of both Gr and Bdnf are found in human offspring of parents with PTSD. 21,50
In this exploratory study, in order to test the hypothesis of biological (i.e., social interaction-independent) heritable effects of parental TBI, we induced TBI in sires, whose sole contribution to their offspring's development can faithfully be traced back to the spermatozoa containing genomic and epigenomic information without further confounding postpartum influences. It will be scientifically and translationally important to extend this work to investigate heritable effects of maternal TBI, for which additional manipulations will be required, such as replacing a biological mother with a foster dam at the time of birth of offspring.
The goal of this initial study was to test the hypothesis that offspring of young adult male rats with TBI develop neurodevelopmental abnormalities in the absence of direct social contact with their sires. It was not the goal of this study to assess intergenerational effects of paternal TBI at a specific age of the sires or their offspring. The sires' age at the time of testing was more advanced because they were involved in TBI procedures and mating. The 10-day delay between the behavioral tests was used to allow sufficient time for recovery from the previous test. Now that we have evidence for long-term intergenerational effects of paternal TBI, many related questions need to be addressed. For example, considering that persistent stress and inflammation are associated with accelerated cognitive decline in later life, these findings suggest that TBI-induced abnormalities may worsen in both generations as life progresses. It will therefore be important to investigate age-dependent dynamics of intergenerational abnormalities induced by parental TBI. In addition, future studies will be needed to determine whether parental TBI lowers offspring's homeostatic reserve in response to stress, inflammation, or their own TBI.
In conclusion, the findings of this study in rats provide evidence that offspring (particularly males) of sires with a history of moderate midline FPI-induced TBI develop neurobehavioral abnormalities in the absence of direct social sire-offspring interaction, supporting a possibility that intergenerational transmission of effects of paternal TBI occurs during conception. The findings further suggest that female offspring of males with moderate TBI are more protected from the adverse effects of paternal TBI, at least during the conditions and tests employed here. Considering evidence from human studies that parental brain injury is associated with adverse psychological/psychiatric outcomes in children, 23 specifically-designed human studies are needed to assess the possibility of direct social interaction-independent transmission of adverse effects of parental TBI to offspring, which may help to develop new therapeutic approaches.
Transparency, Rigor, and Reproducibility Summary
All experimental procedures were approved by the University of Florida Institutional Animal Care and Use Committee. The study was conducted and data are reported in accordance with the ARRIVE guidelines. 30 F0 male rats were randomized into two treatment groups using a random number generator. F0 naïve female rats were used as breeders only to generate offspring (generation F1). Sample size calculations were done, assuming a range of anticipated differences in mean outcomes and standard deviation based on background data and past experience with similar measurements in Sprague-Dawley rats. 27 The primary outcomes of this study were intergenerational neuroendocrine and behavioral effects of paternal traumatic brain injury (TBI); all other outcomes were secondary outcomes. Inclusion and exclusion criteria: the animals were of the same age and a similar weight and have similar baseline behavioral outcomes. Exclusion criteria include immediate post-procedural issues, or any pathophysiological conditions developed later. Animals excluded from the study are reported in the manuscript. A cohort of 18 breeding pairs of F0 rats was used to produce F1 offspring for a given experimental group (18 F1 rats/group/sex). A given F1 offspring experimental group included 1 to 2 rats from a given F0 litter. In all experiments, control and treatment groups were tested using the same experimental conditions (e.g., O2/air mixture in the animal cage during the procedures, time of day). This study investigated both male and female offspring to elucidate sex-specific intergenerational effects of paternal TBI. Data analyses included experimental group (treatment) and sex as the main effects and experimental group (treatment) × sex interactions.
Footnotes
Authors' Contributions
Ling-Sha Ju was responsible for the acquisition, analysis, and discussion of data, and revising the manuscript; Jiepei Zhu was responsible for the traumatic brain injury methodology; Timothy E. Morey, Nikolaus Gravenstein, Christoph N. Seubert, and Barry Setlow were responsible for the conceptualization of the study, data analysis and interpretation, drafting and revising the manuscript; Anatoly E. Martynyuk was responsible for the conception and design of the study, data analysis and interpretation and writing the manuscript.
Funding Information
This study was supported in part by the National Institutes of Health (R56HD102898 and R01HD107722 to A.E.M.), the I. Heermann Anesthesia Foundation (L.-S.J.), and the Jerome H. Modell, M.D., F.A.H.A., Endowed Professorship, Gainesville, Florida (N.G.).
Author Disclosure Statement
T.E.M. owns equity in Xhale, Inc. In addition, the University of Florida owns equity in Xhale, Inc., a faculty start-up company producing alar pulse oximeters for clinical use in humans. N.G. serves as a medical advisor for Teleflex Medical.
For the other authors, no competing financial interests exist.