
Abstract
Traumatic injuries to the spinal cord or the brain have serious medical consequences and lead to long-term disability. The epidemiology, medical complications, and prognosis of isolated spinal cord injury (SCI) and traumatic brain injury (TBI) have been well described. However, there are limited data on patients suffering from concurrent SCI and TBI, even if a large proportion of SCI patients have concomitant TBI. The complications associated with this “dual-diagnosis” such as cognitive or behavioral dysfunction are well known in the rehabilitation setting, but evidence-based and standardized approaches for diagnosis and treatment are lacking. Our goal was to develop and characterize a pre-clinical animal model of concurrent SCI and TBI to help identifying “dual-diagnosis” tools. Female rats received a unilateral contusive SCI at the thoracic level alone (SCI group) or combined with a TBI centered on the contralateral sensorimotor cortex (SCI-TBI group). We first validated that the SCI extent was comparable between SCI-TBI and SCI groups, and that hindlimb function was impaired. We characterized various neurological outcomes, including locomotion, sleep architecture, brain activity during sleep, depressive- and anxiety-like behaviors, and working memory. We report that SCI-TBI and SCI groups show similar impairments in global locomotor function. While wake/sleep amount and distribution and anxiety- and depression-like symptoms were not affected in SCI-TBI and SCI groups in comparison to the control group (laminectomy and craniotomy only), working memory was impaired only in SCI-TBI rats. This pre-clinical model of concomitant SCI and TBI, including more severe variations of it, shows a translational value for the identification of biomarkers to refine the “dual-diagnosis” of neurotrauma in humans.
Introduction
In the United States, approximately 50 individuals per million people are affected by a spinal cord injury (SCI) each year, and in North America about 13,000 individuals per million by a traumatic brain injury (TBI) yearly. 1,2 These injuries are mainly caused by car crashes and falls, sport practice, and violence. 3 –7 Strikingly, up to 70% of SCI individuals also suffer from concomitant TBI, 8 –12 which is undiagnosed in about 60% of cases. 13 This can lead to additional functional deficits and to increased economic burden associated with SCI. 14 A better understanding of the impact of concomitant SCI-TBI is necessary to improve dual-diagnosis and adapt therapies for this patient population.
SCI- and TBI-driven deficits can considerably impact the quality of life. 15,16 Sleep disturbances have been observed in patients suffering from SCI or TBI. 17 –20 In particular, SCI is associated with poor sleep quality, 21,22 sleep-disordered breathing, 22 –24 and periodic leg movements during sleep. 25,26 About half of TBI patients also exhibit wake/sleep disturbances such as insomnia, sleep apneas, hypersomnia, and fatigue. 18,20,27,28 Analyses of the electroencephalographic (EEG) signal recorded during the night, especially during non-rapid eye movement sleep (NREMS) and rapid eye movement sleep (REMS), demonstrated spectral activity modifications in delta (0.8-4 Hz), beta (13-30 Hz), and gamma (30-50 Hz) frequency bands in TBI patients, 29 –31 which also points to alterations in sleep quality. In parallel, emotional distress and cognitive impairments have been observed in both SCI and TBI patients in the acute and chronic phases post-injury. Indeed, 15 to 45% of these patients suffer from anxiety- or depressive-like symptoms, 32 –36 attention deficits, and/or memory impairments. 37 –41 These specific functional deficits arising after SCI or TBI alone could be exacerbated in the context of concomitant SCI-TBI, but the general portrayal of this more complex medical condition remains unclear.
Rodent models of single SCI and TBI have replicated some of these observations in humans. Indeed, decreases in sleep efficiency or in the length of individual bouts of wakefulness/NREMS were observed in rodents submitted to SCI or TBI. 42 –46 Alterations in wake/sleep quality are also supported by higher delta power during wake and NREMS after TBI. 43,47,48 Rodents submitted to either SCI or TBI demonstrate anxiety- and depressive-like behavior, 49,50 and cognitive dysfunctions have been reported in the acute and chronic phases after injuries, including impairments in working and spatial memory. 51 –56 Despite the wide use of SCI or TBI rodent models and their appropriateness in recapitulating features observed in patients, models of concomitant TBI and SCI are lacking. Only one study has evaluated the impact of concomitant SCI and TBI on motor control in rats, demonstrating that motor function after unilateral cervical SCI is hindered by contralateral TBI. 57 With respect to functional deficits related to sleep, cognition and emotional state in the context of concomitant SCI and TBI, there is thus a need to validate a pre-clinical model that will help identifying underlying mechanisms and diagnostic tools to ameliorate the management of this unique patient population.
Here, the impact of TBI on neurological outcomes of SCI was evaluated in a novel model of concomitant neurotrauma. Rats were submitted to a unilateral thoracic spinal cord contusion combined with a brain contusion targeting the contralateral hindlimb sensorimotor cortex, which was followed by weekly assessments of locomotor function, biweekly EEG recordings to evaluate wake/sleep architecture and EEG activity, and emotional state and working memory testing 5-6 weeks post-injuries. We found that the locomotor deficits driven by SCI and the size of the spinal lesion were not exacerbated by concomitant TBI, and that wake/sleep amount and EEG activity, as well as anxiety- and depressive-like symptoms, were unaltered by injuries. However, working memory was found to be impaired only in rats submitted to concomitant SCI and TBI. Our results are discussed with respect to the utility and limitations of the developed model and highlight working memory as a phenotype with potential to distinguish concomitant SCI and TBI from SCI alone.
Methods
Animals
Forty-three Long-Evans female adult rats weighing 220-250 g (Charles River Laboratories, Kingston, NY, USA) were used. Rats were housed individually, and kept in a 12-h light/12-h dark cycle and at 22-25°C, with free access to water and food. Procedures followed the guidelines of the Canadian Council on Animal Care and were approved by the Comité d'éthique de l'expérimentation animale of the CIUSSS-NIM. Twenty rats were excluded due to surgical complications or post-operative complications (two dura matter disruptions during laminectomy or craniotomy, four respiratory arrests, one vertebral dislocation, two lacks of spinal lesion, one hypothermia, one automutilation, two deteriorations of general condition, one death, five losses of electrode montage, one locomotor recovery absence). The final sample comprised 23 animals.
Experimental procedures
The protocol is depicted in Figure 1A. Two weeks after arrival, rats were submitted to a first surgery consisting in EEG and electromyographic (EMG) electrode implantation similar to previously described. 58,59 After a 10-day recovery period, baseline locomotor performance was assessed in an open field, 60,61 and the EEG-EMG signals were recorded for 24 h of baseline/undisturbed conditions (BL). Rats were then randomly assigned to three groups. All rats were submitted to a craniotomy and a laminectomy, followed or not by a spinal/brain injury. The first group did not receive injury (SHAM n = 8), the second received a spinal cord contusion at T8 (SCI n = 9), and the third a concomitant spinal cord and brain injuries (SCI-TBI n = 6). Locomotor performance was reassessed weekly for 4 weeks. EEG-EMG signals were recorded 14 and 28 days after the traumatic/control surgeries (D14 and D28). Anxiety-like behavior, working memory and depressive-like phenotype were evaluated, respectively, at Days 35, 36, and 42 to 46 (D35, D36 and D42-D46), using the open field, Y-maze, and sucrose preference tests. Animals were euthanized at Day 49 (D49), brain/spinal tissues were collected for histological assessment. Weight was measured three times per week from arrival to euthanasia.
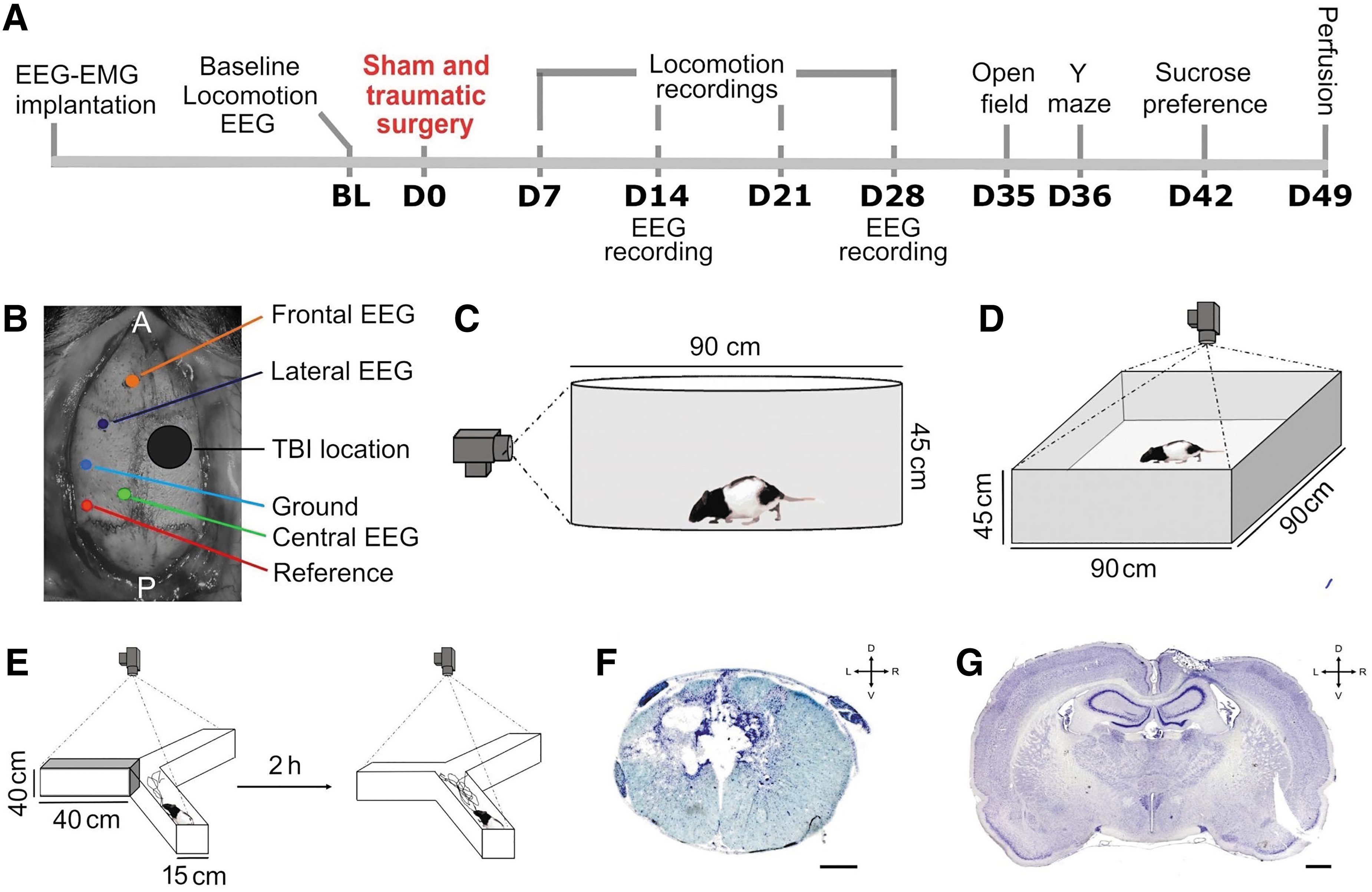
Experimental design and setup.
Surgeries
For electrode implantation, anesthesia was induced with ketamine/xylazine (70/10 mg/kg, intraperitoneally), and was maintained with isoflurane (0.5-1.5%). The head was shaved and disinfected (chlorhexidine gluconate 2% and isopropyl alcohol 70%), and the animal placed in a stereotaxic frame (David Kopf Instrument, CA). Core body temperature was monitored by a rectal thermometer and maintained at 37°C using a heating pad. Three EEG electrodes, a reference, and a ground were implanted on the skull of the left hemisphere after piercing the skull using a 0.7-mm drill tip at the following coordinates: frontal 3.0 mm anterior to bregma, 1.0 mm lateral to midline; central 6.1 mm posterior to bregma, 1.0 mm lateral; lateral 0.7 mm posterior to bregma, 3.0 mm lateral; reference 8.1 mm posterior to bregma, 4.9 mm lateral; ground 4.2 mm posterior to bregma, 4.0 mm lateral (Fig. 1B). Two EMG electrodes were inserted in the neck muscles. Electrodes were cemented to the skull and attached to a connector. Rats received buprenorphine (0.1 mg/kg) upon awakening from anesthesia and the next day. Rats were monitored every day.
Sham or traumatic surgeries took place about 14 days after electrode implantation and 2 days after the start of BL electrophysiological assessment. Anesthesia was achieved with isoflurane (4.0% induction, 0.5-3.0% maintenance). The head and trunk were shaved and disinfected (chlorhexidine gluconate 2% and isopropyl alcohol 70%), and rats were placed in a stereotaxic frame. Body temperature was monitored and maintained at 37°C during surgery. A 4 × 4 mm craniotomy was performed over the right sensorimotor cortex. The window extended between 0.5 to 4.5 mm posterior to bregma and from 0.5 to 4.5 mm lateral to the midline. Laminectomy was then performed as previously described. 62 A midline incision of the skin was made overlaying T6-T10 vertebrae. The skin and superficial muscles were retracted and the paravertebral muscles inserting on the dorsal aspect of the T7-T9 vertebra dissected. A bilateral laminectomy of the T8 vertebra was done to expose the dorsal surface. A moderate lateral contusion of the left side of the spinal cord was performed with a commercial impactor (TBI-0310, Precision Systems and Instrumentation).
The following parameters were used for the spinal impact: speed 4 m/sec, depth 1 mm, dwell-time 150 msec, tip diameter 1.5 mm. A moderate TBI centered 2.5 mm posterior to bregma and 2.5 mm lateral was performed using a 3-mm diameter tip. These coordinates were chosen based on our previous studies showing that the hindlimb representation in rats extends from 0.5 mm to 3 mm posterior to bregma and from 1 to 3 mm lateral to bregma. 60,63 The forelimb representation is anterior to the hindlimb area and ends at around 0.5 mm posterior to bregma. Using these coordinates and tip diameter size, we aimed to spare the forelimb representation and the subcortical circuits to ensure animals could be able to perform the behavioral tests described below. The following parameters were used for the brain impact: speed 4 m/sec, depth 2 mm, and dwell-time 150 msec. In the SCI-TBI group, the spinal and the following brain impacts were performed within 7 min. Rats received buprenorphine (0.1 mg/kg) and lactated Ringer's solution (NaCl 0.9%) upon awakening and the next day. Rats were monitored three times/day during the first week and once/day afterwards. Manual emptying of the bladder was necessary for 3 days post-surgery in three SCI and two SCI-TBI rats.
Locomotor assessment
Locomotion and global motor skills were evaluated using an established locomotor scale recently adapted for evaluating hindlimb locomotor performance. 61 –63 Four days before and 7, 14, 21, and 28 days after traumatic or sham surgeries (D7, D14, D21, and D28), at approximately 1 h after light onset, animals were placed in a circular, transparent acrylic open field arena (90 × 45 cm) for 5 min, and were recorded at 120 Hz with a camera (Basler, acA2000-165uc) for offline analyses (Fig. 1C). Performance was assessed separately for the two hindlimbs. Scores were calculated by assessing excursion amplitude of joints, weight support, fine distal positioning, stepping abilities, and forelimb-hindlimb coordination, for a maximum of 20 points.
EEG-EMG recordings/analyses
Recordings were conducted for 24 h at BL, and at D14 and D28 after traumatic or sham surgeries at a sampling frequency of 1024 Hz using a Lamont amplifier and the software Harmonie (Natus), as previously described. 47 Wake, NREMS, and REMS were identified visually using EEG/EMG traces and attributed to 4-sec epochs. Artifacts were simultaneously identified. The time spent in each vigilance state was averaged for 24-h periods at BL, D14, and D28, and also averaged per hour separately for the 3 days to investigate the 24-h distribution. The number of long bouts of wake (lasting at least 16 min), NREMS, and REMS (lasting at least 60 sec) was calculated for 24-h periods (i.e., BL, D14, D28). Spectral analysis was conducted on the bipolar EEG signal between the frontal and central electrodes for artifact-free epochs using Fast Fourier transform to calculate EEG power density between 0.75 and 20 Hz per 0.25-Hz bin for all states. Two SHAM rats and one SCI rat were excluded from spectral analysis because of insufficient EEG quality. For one SHAM rat, the bipolar signal between the frontal and lateral electrodes was used because the central signal was unusable, while for one SCI rat, the central-lateral bipolar signal was used given an unusable frontal electrode. Spectral profiles for these last two animals were comparable to other animals of the same group. Relative spectral activity was expressed as a percentage of the mean of all 0.25-Hz bin of all three vigilance states during BL, as done previously. 64,65 Low (4.75-6 Hz) and high (6.5-8 Hz) theta activity was computed, respectively, for wake and REMS, and expressed as a percentage of the 24-h BL mean. The time course of NREMS delta (1-4 Hz) activity was also calculated for 12 intervals in the light period and six intervals in the dark period, and expressed as a percentage of the BL mean of intervals nine to 12. 64
Open field
Anxiety-like behavior was evaluated using an open field test. 66,67 Thirty-five days after spinal/brain surgeries, starting 1 h after light onset, rats were placed in the center of a square arena (90 × 90 × 45 cm) without spatial cue (Fig. 1D). The position and movements were recorded for 5 min by a camera (Pelco, 25 frame/sec) positioned on the ceiling. Thigmotaxis (tendency to stay close to walls) is considered an index of anxiety-like behavior in rodents. 68 Two zones were defined by separating the total area in 25 equal squares using Smart 3.0 (PanLab, Harvard Apparatus). The central zone included the nine squares of the middle (54 × 54 cm), and the outer zone all 16 squares adjacent to the walls. 69 The total distance, the time spent in the central zone, and the number of entries in the central zone were analyzed by automatic tracking of the animal using Smart 3.0. The proportion of time spent in the central zone was computed with the formula:

Y-maze
Working memory was evaluated 36 days after brain/spinal surgeries using the Y-maze test (Fig. 1E). 70 –72 During a first session (starting 1 h after light onset), rats were exposed to only two of the three arms (40 × 15 × 40 cm) for 5 min. Spatial cues were placed at the end of each arm. After the first session, rats returned to their home cage for 2 h before a second session. During the second session, rats were exposed to the three arms of the maze for 5 min. Sessions were recorded with a camera (Pelco, 25 frames/sec) positioned on the ceiling. The total distance (in meters) and time spent in each arm were analyzed for the second session using the software Smart 3.0. The time spent in the newly opened arm and in other/previously explored arms (i.e., time per arm averaged for the two previously explored arms) was calculated and reported as a percentage of total time (see formulas below). Rats with preserved working memory usually spend more time exploring the new arm than the previously explored arms.

Sucrose preference
To evaluate a depressive-like behavior indicative of anhedonia, sucrose preference was measured from D42 after spinal/brain surgeries for 5 consecutive days, as described previously. 58,73 –75 Since arrival, animals had access to two bottles containing 150 mL water each to adapt to experimental conditions. During the testing week, animals had access to two bottles for 5 days: one filled with 150 mL water, and one filled with 150 mL of 1% sucrose in water. The position of the bottles was randomized between animals and permuted each day to prevent potential bias related to bottle position. Bottles were weighed each day (2 h after light onset), and the drunk quantity was reported in milliliters. Total sucrose preference over the 5 days and sucrose preference/day were calculated as previously described: 76 –78
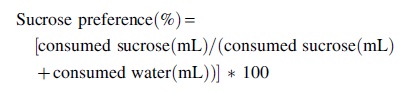
Histology
Seven weeks post-spinal/brain surgeries, rats were anesthetized with ketamine/xylazine (100/10 mg/kg). A transcardial perfusion of 150 mL NaCl 0.9% followed by 250 mL paraformaldehyde (4%) in phosphate buffered saline (PBS; 0.01 M phosphate buffer, 0.0027 M potassium chloride, 0.137 M sodium chloride) was performed. Spinal cords/brains were extracted and post-fixed for 24 h in 4% paraformaldehyde. Tissues were cryoprotected in 30% sucrose, frozen on dry ice, and kept at -80°C. A cryostat (HM525 NX, Thermo Scientific, blade S35 - Feather®) was used to cut 40-μm coronal sections from a spinal block encompassing segments between T6 and T10 and from the brain between the bregma and 4.56 mm posterior to bregma. Every third section was mounted on superfrost plus slides (Fisher Scientific). Spinal sections were stained with cresyl violet (0.5%; Alfa-Aesar, ThermoFisher Scientific) to visualize cell bodies in spinal gray matter and gliosis along the lesion size, and counterstained with Luxol fast blue (0.1%; Sigma-Aldrich) to visualize spinal white matter (Fig. 1F). 60,62 Brain sections were only stained with cresyl violet (0.5%; Fig. 1G). Microscope images were acquired using an Axio Imager 2 (Carl Zeiss Microscopy), and analyzed using Stereoinvestigator (MBF Bioscience). Spinal/brain lesion extent was quantified using the Cavalieri Estimator probe. Each lesion epicenter was drawn manually on templates. Superposition of each lesion profile was conducted with MatLab (MathWorks Inc.) to produce heat maps allowing a general visualization of the extent of lesions (Fig. 2A).

Model validation.
Statistical analyses
An unpaired t-test was used to compare the spinal lesion size between SCI and SCI-TBI groups and paired t-tests were used to compare the time spent in the new arm versus other arms (Y-maze) for each group separately. One-way analyses of variance (ANOVA) were used to compare groups for the time spent in wake/sleep states over the 24-h period, for spectral activity in each 0.25-Hz bin at BL, D14, and D28, for total distance traveled, time in central zone and number of entries in central zone in the open field, for total distance traveled in the Y-maze, and for the 5-day average sucrose preference. One-way repeated measure ANOVA were used to compare days within groups for the 24-h time spent in each state and the number of long state bouts. Two-way repeated measure ANOVA were used with factors Group and Time for weight (weekly averages of Monday, Wednesday, and Friday), with factors Day and Hour for time spent in vigilance states across 24 h, with factors Group and Light/Dark for low and high theta activity, with factors Group and Interval for delta activity, and with factors Group and Day for sucrose preference over 5 days. A three-way repeated measure ANOVA with factors Group, Day, and Hindlimb was used to compare locomotor scores. When appropriate, statistical significance was adjusted for repeated measures with the Greenhouse-Geisser correction and significant effect decomposed using Bonferroni or Fisher post hoc tests. All data are presented as mean ± standard error of the mean, and 0.05 was used as a threshold for statistical significance. Statistical analyses were performed using Statistica (Statsoft) and GraphPad Prism 9.1.2 (GraphPad Software).
Results
Model validation
Histological assessments of the nervous tissue damages were performed for all groups. The craniotomy and laminectomy did not damage the brain and spinal cord, respectively, in SHAM animals. The size of spinal and brain lesions at the epicenter were assessed in SCI and SCI-TBI rats (Fig. 2A). In both groups, the spinal lesion was mainly confined to the left hemicord, with some variability, shared between groups, in damage to the right dorsal column. The mean spinal lesion area at the epicenter did not differ between groups (t = 0.1, p = 0.9). In the SCI-TBI group, the brain lesions were mainly confined to the right hindlimb sensorimotor cortex with some variability between rats. Importantly, none of the rats displayed forelimb deficits that are usually seen in cases of larger TBI. 79
Rats included in this study were weighed three times per week, and weekly averages were compared between groups (Fig. 2B). In general, rats maintained the same weight after electrode implantation and sham surgeries, lost weight after SCI or SCI-TBI, and gain weight from Week 1 post-trauma or sham surgeries until the end of the protocol (Week effect F8,160 = 70.0, p < 0.001). The average weight per week did not differ between groups throughout the experiment (Group by Week interaction F16,160 = 0.7, p = 0.8; Group effect F2,20 = 1.7, p = 0.2), supporting a similar recovery in animals of all groups.
To evaluate the impact of injuries on global motor recovery, hindlimb locomotor function was evaluated once a week for 1 month (Fig. 2C). SHAM rats did not exhibit hindlimb locomotor deficits (maximum score of 20 throughout the experiment). SCI and SCI-TBI groups exhibited hindlimb locomotor deficits from D7 to D28 after injuries. Given that the spinal lesion was mostly localized on the left side of the spinal cord, the left hindlimb showed lower scores (more impairment) than the right hindlimb for both the SCI and SCI-TBI groups (Hindlimb effect F1,12 = 56.3, p < 0.001). In addition, both groups exhibited recovery of motor function between D7 and D28 (Day effect F3,36 = 9.8, p < 0.001). Qualitatively, SCI and SCI-TBI rats initially presented some hindlimb paralysis, showed recovery of bilateral stepping and weight support over the first 2 weeks, and presented prolonged (up to D28) deficits in distal movement and limb coordination. SCI and SCI-TBI groups did not differ in their locomotor scores over the course of the experiment (Group by Day interaction F3,36 = 0.6, p = 0.6; Group effect: F1,12 = 0.9, p = 0.3), and in the left-right hindlimb difference (Group by Hindlimb interaction F1,12 = 0.01, p = 0.9; Group by Hindlimb by Day interaction F3,36 = 0.3, p = 0.8). This indicates that TBI did not exacerbate locomotor deficits related to SCI, which is in line with observations for body weight and size of spinal lesion.
Wake/sleep architecture
Sleep architecture/microarchitecture were assessed using EEG/EMG recordings performed at BL, D14, and D28. The total time spent in wake, NREMS, and REMS was first compared between groups at BL to verify whether groups were similar prior to injury: groups did not differ in time spent awake and NREMS (Group effects F2,20 ≤ 2.0, p ≥ 0.2), but a significant difference was detected for the time spent in REMS (Group effect F2,20 = 4.1, p = 0.03). Given this BL difference, each group was independently analyzed to assess the effect of experimental day for time spent in states and number of long state bouts (each group being its own control). In SHAM rats, the total time spent in wake, NREMS, and REMS did not differ between BL, D14, and D28 (Day effects F2,14 ≤ 0.1, p ≥ 0.9; Fig. 3A). In both groups with traumatic injuries, the total time spent in each state also did not significantly differ between days (SCI Day effects F2,16 ≤ 1.8, p ≥ 0.2; SCI-TBI Day effects F2,10 ≤ 0.5, p ≥ 0.6; Fig. 3A).

Time spent in wake, non-rapid eye movement sleep (NREMS) and rapid eye movement sleep (REMS) and number of long bouts after spinal cord injury (SCI) or concomitant SCI and traumatic brain injury (TBI).
To assess whether traumatic injuries alter time spent in vigilance states only at specific moments, the hourly distribution of wake/sleep states was compared between BL, D14, and D28 within each group (Fig. 3B). The typical light/dark distribution of states (i.e., more time asleep during the light and more time awake during the dark) was observed in all groups (Hour effects F23,115-184 ≥ 7.5, p < 0.01). In SHAM rats, the hourly distribution of wake, NREMS, and REMS was not affected by experimental days as anticipated (Day by Hour interactions F46,322 ≤ 1.8, p ≥ 0.1; Day effects F2,14 ≤ 0.2, p ≥ 0.8). In both groups with traumatic injuries, the hourly distribution of states also did not significantly differ between BL, D14, and D28 (SCI Day by Hour interactions F46,368 ≤ 1.6, p ≥ 0.06; Day effects F2,16 ≤ 1.8, p ≥ 0.2; SCI-TBI Day by Hour interactions F46,230 ≤ 1.1, p ≥ 0.3; Day effects F2,10 ≤ 0.6, p ≥ 0.6). Overall, the time spent in wake, NREMS, and REMS as well as their 24-h hourly distribution were preserved in all groups over the course of the experiment.
Previous work, including ours, has shown that TBI specifically impaired the ability to maintain long wake bouts. 47,80 We therefore investigated whether SCI and SCI-TBI impacted the number of long wake/sleep bouts (Fig. 3C to 3E). In the SHAM group, the total (summed over 24 h) number of long bouts of wakefulness, NREMS, and REMS did not differ between experimental days (Day effects F2,14 ≤ 1.0, p ≥ 0.4). In both groups with traumatic injuries, the total number of long bouts of vigilance states also did not differ between days (SCI Day effects F2,16 ≤ 1.1, p ≥ 0.3; SCI-TBI Day effects F2,10 ≤ 1.3, p ≥ 0.3). These results suggest that, in our model, spinal and brain injuries do not impact the capacity to maintain consolidated episodes of wakefulness and sleep when measured 2 and 4 weeks post-injury.
Wake/sleep EEG activity
EEG spectral analysis provides quantitative metrics informing on the spectral signature and quality of vigilance states. 81 Spectral activity was first calculated between 0.75 and 20 Hz for the full 24 h for each state and compared between groups at BL, D14, and D28 (Fig. 4A). The characteristic spectral signatures of wake, NREMS, and REMS (e.g., predominant low frequency activity during NREMS and theta activity during REMS) were preserved in all groups over experimental days. Spectral activity at BL was generally indistinguishable between groups for all states (Group effects for all 0.25-Hz bins of all states F2,17 ≤ 3.5, p > 0.05, except for the 5.0-Hz bin during wake with F2,17 = 3.6, p < 0.05 and significant difference between SHAM and SCI-TBI p = 0.02). The visual inspection of profiles could indicate that SCI-TBI animals have elevated activity for all frequencies in all states at D14, and that both SCI and SCI-TBI groups have more activity (especially during wake and REMS) at D28. However, significant higher activity in SCI-TBI in comparison to SHAM rats was only found at D14 for 5.25-Hz during REMS (F2,17 = 4.2, p = 0.03, post hoc between SHAM and SCI-TBI p = 0.03), with adjacent bins 4.75 to 5.75 showing only tendencies (0.05 < p < 0.1) for significant differences. Tendencies were also found for 7.0 to 8.25 Hz during NREMS at D14. Therefore, except for REMS 5.25 Hz activity at D14, no significant between-group difference was found for all states (Group effects for all bins of all states: D14 F2,17 ≤ 3.4, p > 0.05; D28 F2,17 ≤ 2.5, p > 0.05).
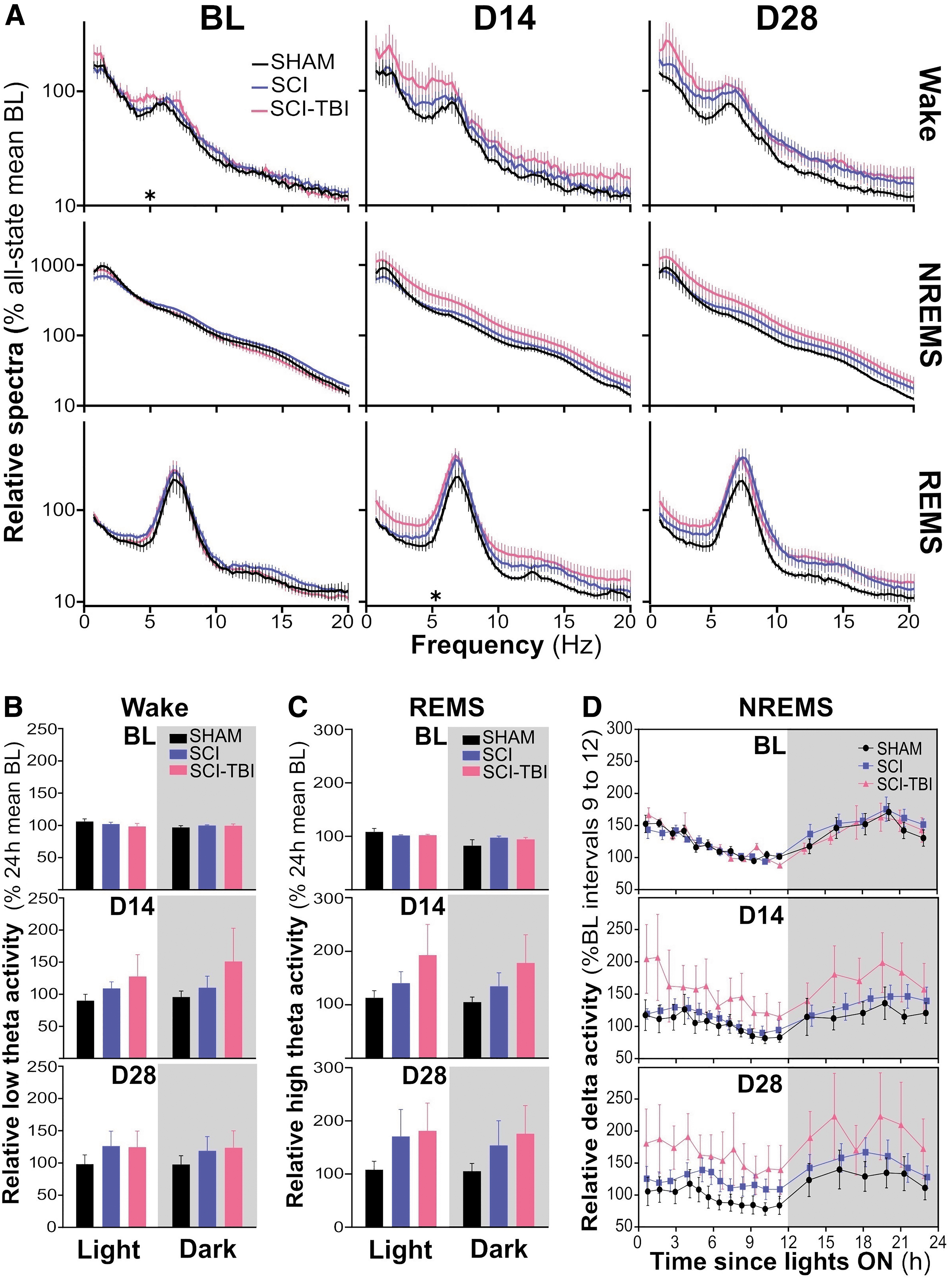
Spectral activity during wake, non-rapid eye movement sleep (NREMS) and rapid eye movement sleep (REMS) after spinal cord injury (SCI) or concomitant SCI and traumatic brain injury (TBI).
Given that theta activity during wake was linked to sleep need and NREMS homeostasis, 82,83 and that REMS theta activity was causally associated with spatial memory, 84 wake and REMS theta activity were analyzed as a function of the light/dark cycle. Low theta (4.75-6 Hz) activity during wake did not differ between groups at BL, which was also the case for both D14 and D28 (Group by Period interactions F2,17 ≤ 0.8, p ≥ 0.5; Group effects F2,17 ≤ 2.3, p ≥ 0.1; Fig. 4B). Similarly, high theta (6.5-8 Hz) activity during REMS did not significantly differ between groups at BL, D14, and D28 (Group by Period interactions F2,17 ≤ 2.0, p ≥ 0.2; Group effects F2,17 ≤ 1.7, p ≥ 0.2; Fig. 4C).
We examined the impact of traumatic injuries on the time course of NREMS delta activity (Fig. 4D), which is considered a marker of the sleep recovery process. 85 –88 The 24-h time course was strikingly similar between SHAM, SCI, and SCI-TBI groups at BL (Group by Interval interaction F34,289 = 0.7, p = 0.9; Group effect F2,17 = 0.1, p = 0.9), showing the typical decrease over the course of the light period and increase during the dark period (Interval effect F17,289 = 16.5, p < 0.001). This pattern was globally preserved at D14 and D28 (Interval effects F17,289 ≥ 7.0, p < 0.001). Although delta activity appeared to be generally increased in SCI-TBI animals in comparison to other groups at D14 and D28, the high variability in the SCI-TBI group prevented from detecting significant differences between groups (Group by Interval interactions F34,289 ≤ 0.9, p ≥ 0.7; Group effects F2,17 ≤ 1.4, p = 0.3). Overall, spectral profiles during wake, NREMS and REMS, wake and REMS theta activity, and the time course of NREMS delta activity were not significantly affected in our model when evaluated 2-4 weeks post-trauma.
Emotional state
Anxiety-like behavior was assessed using the open field at D35. The total distance traveled during the 5 min test did not significantly differ between groups (Group effect F2,20 = 2.6, p = 0.1; Fig. 5A), demonstrating that all groups exhibited similar exploratory activity despite diminished locomotor performance in injured groups (Fig. 2C). An absence of significant difference between groups was also found for the percent time spent in the central (anxiogenic) zone and the number of entries in this zone (Group effects F2.20 ≤ 0.7, p ≥ 0.5; Fig. 5B-C). Depressive-like phenotype was evaluated using the sucrose preference test performed over 5 days (D42 to D46). Sucrose preference averaged over the 5 days did not differ between groups (Group effect F2,20 = 0.7, p = 0.5; Fig. 5D). Sucrose preference calculated separately for each of the five days also did not significantly differ between groups (Group by Day interaction F8,76 = 1.0, p = 0.4; Group effect F2,19 = 0.9, p = 0.4; Fig. 5E). Together, these results demonstrate that neither concomitant SCI-TBI nor SCI induced anxiety- and depressive-like behaviors at 5-6 weeks post-injury.

Anxiety- and depression-like behaviors and working memory after spinal cord injury (SCI) or concomitant SCI and traumatic brain injury (TBI).
Working memory
Working memory was investigated at D36 using the Y-maze. The total distance traveled during the 5-min test session was not different between groups (Group effect F2,19 = 0.7, p = 0.5; Fig. 5F), which confirms capacity to perform the test in groups with traumatic injuries. The time spent in the newly opened arm versus the previously explored arms was used as a memory indicator and compared within each group (Fig. 5G). SHAM and SCI animals spent more time in the new arm compared with other arms (t ≥ 2.7, p ≤ 0.03), indicating that SCI alone did not impair working memory. In contrast, rats with concomitant SCI-TBI did not significantly differ in the time spent exploring the new versus the previously explored arms (t = 0.5, p = 0.6), which points to a memory deficit. Therefore, although working memory was preserved in rats undergoing SCI, it was impaired when SCI was combined with a TBI.
Discussion
Given the high incidence of TBI in the SCI population, 8 –12 the present study aimed to develop a pre-clinical model of concomitant SCI and TBI. We have characterized, for the first time in this model, various neurological outcomes, including locomotion, sleep quantity/quality, depressive- and anxiety-like behaviors, and working memory. We report that concomitant SCI and TBI neither disrupted sleep architecture and spectral activity nor emotional state. However, rats submitted to SCI and TBI expressed a deficit in working memory, which was not present after SCI only. These findings thus support the potential use of working memory evaluations to refine the dual diagnosis toolset applicable to neurotrauma.
Animal models of neurotrauma have been tremendously useful to understand cellular/molecular mechanisms and help identifying biomarkers. 89 –92 The high incidence of co-morbid SCI and TBI justifies the need for relevant pre-clinical models. Only one model of concomitant cervical SCI and TBI was previously developed to evaluate upper limb motor function. 57 We herein developed a model of concomitant thoracic SCI and TBI since thoracic injuries represent 35% of SCI and lead to lower limb deficits that are observed in most SCI individuals. 89 The extent of spinal and brain injuries observed in our model was comparable to those reported in previous studies, 93 –95 and spinal lesion extent was comparable between groups with single and concomitant neurotrauma. Both SCI and SCI-TBI animals gradually recovered locomotion, similar to previous studies of traumatic spinal and brain injuries. 57,79,96 The contralateral (right) TBI, damaging the hindlimb motor cortex representation and given simultaneously to the left-sided thoracic SCI, did not significantly impact global locomotor performance measured with the open field scale. Similar findings using the same locomotor score were reported after unilateral cervical SCI combined to larger contralateral TBI damaging both the forelimb and hindlimb sensorimotor cortices and some subcortical circuits. 57 This fast locomotor recovery (i.e., first week) is likely due to an immediate take-over of function by residual motor pathways, including the ipsilesional motor cortex, 63 brainstem structures, 97,98 and spinal circuits. 99 In accordance with previous studies, our results suggest that, in rats, the hindlimb motor cortex is not necessary for generating automated behaviours, 60,100 such as walking in an open field. However, it may play a compensatory role after SCI, since rats submitted to chronic inactivation of the contralateral hindlimb motor cortex after a spinal cord hemisection fail to properly recover locomotion, exhibiting deficits in initiating flexion during swing and in performing skilled locomotor tasks requiring cortical integration. 63 These deficits, reported during treadmill walking and ladder crossing, were not evaluated in the present study but could emerged with additional motor tests, including kinematic analyses and skilled locomotor tasks. 60,101 A full characterization of the range of locomotor deficits associated with concomitant injuries will be essential together with the assessment of more severe concomitant injuries.
Sleep alterations were reported in animal models of traumatic injuries and clinical populations. A severe SCI achieved via spinal cord transection was found to reduce sleep efficiency and to increase percent time spent awake in rats, especially when considering the light period (corresponding to the rest period in rodents). 42 Similarly, SCI was observed to trigger insomnia in humans, 19 while TBI decreased the ability to maintain arousal/wake in rats, 45 but increased sleep duration in the first week(s) post-injury in humans. 20 Surprisingly, SCI and combined SCI-TBI did not significantly affect sleep duration/architecture in our model, which could be explained by the relatively low severity of both injuries in comparison to previous research (e.g., Esteves and colleagues). 42 In addition, given that previous sleep alterations in rodent models of neurotrauma were found in the first days after injuries (e.g., from days 1 to 12 after severe SCI in rats), 42,47 measurements conducted at Weeks 2 and 4 in the present study might have missed early detectable alterations in sleep. Sleep architecture was indeed found to be unchanged months/year in TBI individuals reporting sleep complaints. 102 Performing sleep evaluation as early as possible after injury is more compatible with the development of diagnostic tools, but represents a challenge in clinical setting. 20 Our model will be useful to test the level of injury severity impacting wake/sleep variables and to define whether acute sleep electrophysiology could help refining dual diagnosis.
Wake/sleep quality assessed using EEG spectral analysis could be more sensitive to identify biomarkers and diagnostic features in comparison to sleep architecture. Alterations in spectral activity during wake and NREMS were reported in animal models of TBI. 43,47,48 For instance, TBI associates with higher delta activity during wake in mice. 47 We here observed that the spectral signature of wakefulness, NREMS, and REMS is not significantly impacted by SCI alone or combined to TBI at Weeks 2 and 4 post-injury. Measurements performed closer in time to the injury would have likely allow to detect EEG activity modifications as found in the second day or first week after TBI in mice. 47,80 Importantly, the relatively modest severity of injuries and the large variability observed in the SCI-TBI group has likely prevented the detection of significant differences. The large variability could be a defining feature of double neurotrauma and is a concern in the development of diagnostic tools in the clinical population (e.g., TBI localizing at different brain regions). As such, the inclusion of additional EEG electrodes, including electrodes targeting the hemisphere ipsilateral to the TBI, might increase the sensitivity to detect alterations in the quality of the wake/sleep EEG. Other metrics related to the quality of the sleep EEG, such as the properties of individual slow waves during NREMS, 103,104 could be used in future studies to investigate their relevance in the development of diagnostic tools and biomarkers.
Anxiety and depressive symptoms are commonly observed in patients after SCI or TBI. 33,35,36,105,106 Our model did not recapitulate these symptoms (i.e., preserved time spent in the open field anxiogenic zone and sucrose preference). The absence of anxiety-like phenotype in the open field is relatively in line with previous SCI studies in rats, 50,107 but not with findings of a difference in a rodent TBI model. 49 In parallel, our finding of preserved sucrose preference differs from the decreased preference reported >5 months after TBI in rats. 108 The ability to detect emotional phenotypes in rodent models could be enhanced by using multiple tests, including the elevated plus maze for anxiety. 107 In the present study, the choice of assessing multiple different phenotypes together with the selection of tests with a high probability of successful completion by animals with impaired locomotion, has prevented an in-depth assessment of the emotional state. In patients, anxiety and depressive symptoms might be mainly driven by extensive changes to the immediate environment (e.g., wheelchair, hospital setting, rehabilitation) and by the stress associated with the economic impact (e.g., job loss, medical care cost). 14,15,109,110 Future animal studies evaluating the impact of environmental changes and stress after neurotrauma could be useful to investigate the mechanisms underlying anxiety- and depressive-like phenotypes.
We found that concomitant SCI and TBI produce a deficit in working memory, whereas SCI alone had no impact. This finding could suggest that a working memory deficit is specifically driven by the concomitant TBI. This interpretation is supported by observations of memory impairments in rodents submitted to TBI only, 51,52,111 but not by findings of working memory deficits after SCI in rats. 53,56 The difference between these last two studies and our study could reside in the use of males versus females, given the reported sex difference in learning strategies in rats. 112 To validate the potential use of working memory as a diagnostic tool that could discriminate concomitant SCI-TBI from SCI alone, additional tests could be performed in males and females at different recovery stages.
One limitation of our study concerns the use of spinal/brain contusion after laminectomy and craniotomy. Although this choice allows to increase reproducibility and to decrease between-animal variability, 90,92 the translational potential could be lower than intact bone configurations more closely resembling injuries in humans. A second limitation concerns the sole use of females, especially given the higher incidence of SCI and TBI in men in comparison to women. 4,113 Female rats were used because of the experimental constraints related to impaired bladder functions in male with low SCI, 114,115 and the ethic issue related to higher mortality rate after low SCI in male rats. Nevertheless, studies in males will be required to develop suitable diagnostic tools and biomarkers for clinical settings. A last limitation of the current research relates to the location of EEG electrodes on the contralesional hemisphere. This montage was chosen given the incompatibility between securing electrode to the skull with cement and applying a TBI with an open skull configuration afterwards.
Conclusions
By assessing sleep phenotypes, emotional state, and cognition after SCI-TBI, our novel model provides new insights regarding potential biomarkers that could differentiate concomitant lesions from a single SCI. We found that working memory was impaired by concomitant injuries and could be considered to develop future diagnostic tools and, eventually, targeted therapeutic strategies for patients suffering from concomitant SCI-TBI. Even if sleep quantity and quality were not particularly disrupted in the current dataset, the role of sleep in the functional recovery from dual injuries to the nervous system could be investigated in more severe variants of the present model, for instance with TBI targeting the cerebral cortex representation of both the forelimb and hindlimb, or in SCI-TBI animals submitted to sleep deprivation. Finally, the proposed model could also be used to test the suitability of novel neuroprosthetic approaches making use of brain stimulation 101,116,117 as a therapeutic treatment of concurrent SCI and TBI.
Transparency, Rigor, and Reproducibility Summary
This exploratory work aimed at developing a novel animal model. Consequently, the study design and analytic plan were not registered and the statistical power was not calculated. Sample size was determined based on our previous studies. Forty-three Long-Evans females were used. Twenty rats were excluded due to surgical complications or post-operative complications (two dura matter disruptions during laminectomy or craniotomy, four respiratory arrests, one vertebral dislocation, two lacks of spinal lesion, one hypothermia, one automutilation, two deteriorations of general condition, one death, five losses of electrode montage, one locomotor recovery absence). The final sample comprised 23 animals that were randomly assigned to one of the following groups: the first group did not receive injury (SHAM n = 8), the second received a spinal cord contusion at T8 (SCI n = 9), and the third a concomitant spinal cord and brain injuries (SCI-TBI n = 6). Analyses were performed by team members who were aware of the characteristics of the subjects and were then re-analyzed blindly by the senior authors. The normal distributions of primary behavioral and histological outcome data were verified using Shapiro-Wilk tests. Data will be made available from the senior authors upon reasonable request. Any additional information required to reanalyze the data reported in this paper is available from the corresponding author.
Footnotes
Acknowledgments
We thank Catherine Henri for her participation in behavioral data acquisition; Dr. Andrew Brown for his help with locomotor scoring; Dr. Marco Bonizzato for sharing the MatLab code to produce heatmaps of brain and spinal cord injuries; and Roger Godbout and Jonathan Brouillette for discussions on the experimental design and analyses.
Authors' Contributions
MR: developed methodology and performed experiments, data analysis and visualization, writing—original draft. JDG: performed electrode implantation, traumatic surgeries, and tissue collection, contributed to data analysis and representation, and writing—original draft. CP performed electrode implantation and traumatic surgeries, and behavioral tests. MM and VM: conceptualization, methodology, validation, resources, data analyses and curation, writing—review and editing, supervision, project administration, funding acquisition.
Funding Information
This work was supported by an operating grant from Recherche CIUSSS-NIM to MM and VM. MR was supported by a fellowship from the Fondation NeuroTrauma Marie-Robert, a prestigious fellowship Tomàs-Reader from the Université de Montréal, and a fellowship for doctoral studies completion within four years from the Université de Montréal. MM was supported by a salary award from Fonds de Recherche du Québec-Santé and VM by the Canada Research Chair in Sleep Molecular Physiology.
Author Disclosure Statement
No competing financial interests exist.