
Abstract
Early life stress (ELS) affects neurogenesis and spatial learning, and increases neuroinflammation after a pediatric mild traumatic brain injury (mTBI). Previous studies have shown that ELS has minimal effects in juveniles but shows age-dependent effects in adults. Hence, we aimed to evaluate the effects of ELS in adult male rats after an mTBI. Maternal separation for 180 min per day (MS180) during the first 21 post-natal (P) days was used as the ELS model. At P110, the rats were subjected to a mild controlled cortical impact injury (2.6 mm) or sham surgery. Spatial learning was evaluated in the Morris water maze (MWM) 14 days after surgery and both microglial activation and neurogenesis were quantified. The results indicate that MS180 + mTBI, but not control (CONT) + mTBI, rats show deficiencies in the acquisition of spatial learning. mTBI led to comparable increases in microglial activation in both the hilus and cortical regions for both groups. However, MS180 + mTBI rats exhibited a greater increase in microglial activation in the ipsilateral CA1 hippocampus subfield compared with CONT + mTBI. Interestingly, for the contralateral CA1 region, this effect was observed exclusively in MS180 + mTBI. ELS and mTBI independently caused a decrease in hippocampal neurogenesis and this effect was not increased further in MS180 + mTBI rats. The findings demonstrate that ELS and mTBI synergistically affect cognitive performance and neuroinflammation, thus supporting the hypothesis that increased inflammation resulting from the combination of ELS and mTBI could underlie the observed effects on learning.
Introduction
Adverse childhood experiences (ACE) are pernicious forms of early life stress (ELS) that affect the development of the central nervous system (CNS) and increase the vulnerability to pathologies later in life. 1 –4 Recent evidence suggests a positive association between ACE and traumatic brain injury (TBI). 5,6 Specifically, individuals reporting high ACE scores have significantly greater chances of suffering an adult TBI than those reporting no ACE. Moreover, a history of stressful events may predispose those with mild TBI (mTBI) to have poor outcomes. 7 Also, vulnerable populations like incarcerated women and the homeless have a positive association between ELS and TBI. 8,9 The importance of early prevention of ACE is undeniable, but there is also a need to focus on the adult population that experienced ELS and have endured its consequences for decades. While pre-clinical models can enhance our understanding of the relationship between ELS and adult TBI, there is currently a scarcity of such models that incorporate these variables and afford an appropriate transability of research.
Maternal separation (MS) in rodents is the most extensively characterized model of ELS. 10 -12 Despite some variations in duration, this model is a chronic stressor that commonly consists of daily separation of newborn animals from their dams for 180 min (3 h, MS180) during the first 2 or 3 weeks of life. As adults, MS180 male rats show an increase in baseline hypothalamic-pituitary-adrenal axis (HPA) activity. 11,13 –17 MS180 also causes a decrease in adult hippocampal neurogenesis 16,18,19 and “primes” microglia to be more responsive to subsequent challenges later in life, leaving a permanent memory of the stressful experience. 20 -22
Previous work from our group has shown that the combination of MS180 and pediatric mTBI impacts cognitive performance in the Morris water maze (MWM), increases microglial activation in the contralateral CA1 and promotes cytokine expression in the hippocampus. 23,24 However, we found that mTBI did not have an additional effect on HPA axis activity in maternally separated adolescents. 24 In addition, MS180 alone did not affect the survival of dentate gyrus newly generated cells, nor did it trigger hippocampal neuroinflammation in juveniles. 23
MS180 has an age-dependent effect on hippocampal neurogenesis and HPA axis reactivity. 16,19 Although we only observed a mild effect of MS180 in adolescents, 23 its impact became more apparent with age. 15 -17,25,26 Moreover, it has been proposed that the heightened inflammatory profile of microglia in aging is associated with “primed” phenotype that could be modulated by stress, 27 and that ELS-induced priming could increase reactivity to a secondary insult such as an immune challenge 28 or CNS injury. 20,29 Taken together, these findings suggest that the vulnerability to TBI induced by ELS could also increase with age. Therefore, the goal of this study was to investigate if ELS affects spatial learning, microglial activation, and neurogenesis following an mTBI in adult rats.
Methods
Animals
Ten time pregnant female Sprague–Dawley rats were obtained from the Neurobiology Institute of the National Autonomous University of Mexico at gestation Day 14 and allowed to acclimatize in the facility 1 week before delivery. Rats were maintained in temperature-controlled rooms (20 ± 2°C, 20 ± 3% humidity) with a 12-h light/dark cycle and ad libitum food and water. Day of birth was considered post-natal Day zero (P0). On P1, litters were cross-fostered, culled to eight pups (4-6 males), and randomly assigned to MS180 or control (CONT) groups. Pups were weaned at P21, and on P110 were randomly assigned to trauma (mTBI at 2.6 mm) or Sham groups using a random number generator. Only males were evaluated; females were reserved for a different study. To avoid litter-dependent effects, cross-fostering and random assignment of each rat to experimental condition was performed. Each group consisted of individuals from five different dams (n = 10 rats per group).
All behavioral tests were performed by an experimenter blinded to conditions and testing was conducted before 13:00 h to avoid the effects of circadian variation. All procedures were approved by the National Research Committee of the Mexican Social Security System (R-2019-785-060) and were carried out in accordance with the Institute of Laboratory Animal Resources Guide for the Care and Use of Laboratory Animals and the official regulations for use and care of laboratory animals of Mexico (NOM-062-ZOO-1999). Every effort was made to reduce the number of rats used and to minimize suffering.
Maternal separation
MS180 was performed as previously described. 15 -17,24 -26,30 Briefly, on P1, MS180 litters were separated from their dam and transferred to a box filled with clean sawdust and placed over a heating pad with controlled temperature (30-32°C). Pups were then relocated to a room independent from the main colony and then returned to the dam after 3 h (180 min) of separation. Separation was performed daily from 09:00 to 12:00 h until weaning at P21. The CONT group remained undisturbed except for routine cage cleaning twice a week.
Controlled cortical injury
Rats weighing 320-490 g were subjected to a controlled cortical injury (CCI) or Sham injury on P110 with some modifications to the procedure previously described. 24,30 -32 Briefly, anesthesia was induced and maintained with inspired concentrations of 5% and 2% isoflurane, respectively, in O2. The rats were secured in a stereotaxic frame and a midline scalp incision was made under aseptic conditions. The skin and fascia were reflected to expose the skull and a craniectomy encompassing bregma and lambda and the sagittal and coronal sutures was made in the right hemisphere with a power drill. The bone flap was removed, the impacting rod (Leica Impact One™ Stereotaxic CCI Instrument) was extended, and the impact tip (5 mm, flat) was centered and lowered through the craniectomy until it touched the dura mater, then the rod was retracted, the impact tip was advanced 2.6 mm and an impact (4 m/sec) produced an mTBI. The incision was sutured and a topical veterinary antimicrobial, anti-inflammatory, and healing cream (Petskrim, Grupo LoVet, Mex) was applied. Body temperature was monitored with a rectal probe and maintained at 37 ± 0.5 °C with a heating blanket. A surgical plane of anesthesia was continued through the entire procedure using a commercial stereotaxic anesthesia mask (Harvard Apparatus), including suturing. Sham-operated rats underwent similar surgical procedures but were not subjected to the impact. The rats were administered penicillin (10,000 U/kg body weight intraperitoneally, PISA, Mex) for 3 days to prevent post-operative infections.
Acute neurological evaluations
Assessment of hind limb reflexive ability commenced immediately after the cessation of anesthesia by gently squeezing the rats' hindpaw every 5 sec and recording the time to elicit a withdrawal response. Righting reflex was determined by the time required to turn from the supine to prone position three consecutive times. 30 –33
Motor performance
The beam-balance task was utilized to evaluate gross motor performance from P110 to P115, as previously described. 31,33 Briefly, rats were trained to balance on an elevated narrow beam (1.5 cm wide) until they reached a 60 s criterion before surgery. The time the rats remained on the beam was recorded on post-operative Days 1-5.
Spatial learning evaluation
Cognitive performance was assessed in a Morris water maze (MWM) task 34 from P124 to P129. The maze consisted of a circular pool (160 cm diameter × 60 cm height) that was filled with water (26°C ± 1°C) to a height of 28 cm. The pool was divided in four quadrants and located in the center of a room with salient visual cues that remained constant throughout the training. The platform was placed 2 cm below the water's surface in the northeast quadrant. For evaluating spatial learning acquisition, each rat was provided four trials per day (4-min inter-trial interval) for 5 consecutive days of training. In each trial, the rat was placed in the maze facing the wall starting from a different quadrant and given a maximum time of 120 sec to find the platform. Rats that were unable to find the platform were manually guided to it and allowed to remain on it for 20 sec before being removed, towel dried, and returned to their cage between trials. The time to reach the platform was recorded in all trials. On P129, a probe trial was performed to assess memory retention and consisted of removing the escape platform and allowing each rat to swim freely for 30 sec. The percentage of time the rats spent in the target quadrant (i.e., previous location of the platform) was calculated. On the same day, but after the probe trial, a visible platform test was performed with the platform elevated 2 cm above the water level to determine the contribution of factors such as visual acuity and sensory-motor performance. 30,32,33,35
Tissue collection
Euthanasia was performed at P131. Rats received an overdose of sodium pentobarbital (intraperitoneally) and were intracardially perfused with 200 mL of saline solution (0.9% NaCl) and 200 mL of 4 % paraformaldehyde (PFA) in 0.1 M phosphate buffer solution (PB). The brains were extracted, placed in 4 % PFA for 24 h, transferred to 20 % and 30 % sucrose in PB until they sank, and then were frozen. Random systematic sampling of 40-μm thick slices along the coronal axis of the hippocampus was performed on a cryostat. The sections were placed in tubes with cryoprotective solution (25 % glycerol, 25 % ethylene glycol, 50 % PB) until processing.
Immunohistochemistry
Immunostainings were performed as previously described. 16,23 Briefly, endogenous peroxidases were blocked in hydrogen peroxide and methanol, and slices were further incubated in primary antibody solution. For neurogenesis assessment, we used a primary antibody for detecting the marker for neuroblasts and immature neurons doublecortin (guinea pig anti-DCX, 1:7000, Millipore Cat. No. AB2253). For microglial detection, we utilized an anti-ionized calcium-binding adapter molecule 1 antibody (rabbit anti-Iba1, 1:3000, Wako Cat No.019-19741). Next, they were incubated in the corresponding biotinylated secondary antibody solution (1:750, anti guinea pig Cat. No BA-700 and anti-rabbit Cat. No. BA-1000, Vector Laboratories) and visualized with an avidin-biotin complex (Elite ABC kit, Vector Laboratories) and a Ni-DAB solution (DAB staining kit, Vector Laboratories), mounted, and cover-slipped.
Microglial morphology analysis
Microglial morphology in the cortical area surrounding the injury (CX), the cornuAmmonis1 (CA1), and hilus hippocampal regions from the ipsilateral and contralateral hemisphere were evaluated. For each rat, 20 cells from four different slices from each area were analyzed (5 cells/4 slices/6 areas). The sections were selected to correspond to the same level in all rats to achieve a uniform analysis. Images were obtained using a light microscope with 200 × magnification. The cell-body area and the covered environment area (CEA) including cytoplasmic elongations were calculated using the ZEISS ZEN lite software (v. 3.7, Carl Zeiss), by a researcher blinded to the experimental conditions as previously described. 36,37 The data were utilized for calculating the ratio of these parameters (CEA/cell-body area). Microglial cells were additionally classified as resident or activated/bushy based on their CEA/cell-body area ratio.
DCX+ cell counting
Rats were euthanized 3 weeks after mTBI or Sham surgery so that the double cortin-positive (DCX+) cells included in the quantifications were those generated during the first few days after CCI. Total number estimations were performed as previously described. 15,16,23 For DCX+ cell number estimations, a series of sagittal sections (240-μm intervals) were randomly selected for immunohistochemistry and all DCX+ positive cells along the entire hippocampus dentate gyrus (DG) were counted by a researcher blinded to the experimental conditions. Total cell numbers were calculated according to the Cavalieri principle. Both the contralateral and ipsilateral hemispheres were evaluated.
Statistical analysis
The results are expressed as the mean ± standard error of the mean (S.E.M.). The acute neurological assessments, probe trial, microglial activation, and neurogenesis values were analyzed by two-way analyses of variance (ANOVAs) with ELS and injury as factors. The escape latencies were evaluated by a three-way repeated measures ANOVA with time (repeated factor), stress, and injury as factors. When the ANOVA revealed a significant effect, Tukey´s post hoc tests were utilized to determine specific group differences. The results were considered significant when p ≤ 0.05.
For estimating the CEA/cell-body area ratio threshold, microglial cells were classified as resident, or activated/bushy based on their morphology 38,39 by an experienced researcher blinded to the condition. The CEA/cell-body area ratio from 473 resident cells, 363 intermediate, and 713 bushy/activated cells were utilized to perform a simple descriptive analysis and was calculated as previously described. 36,37 The median ratio for resident cells was 63.6 (99.5% CI: 28.0-248.6) and for bushy/activated cells was 9.5 (99.5% CI: 3.8-26.7), thus allowing us to establish that a ratio smaller than 23 allows detection of bushy cells with a 99.5% confidence. This threshold was utilized to calculate the percentage of activated cells per subject.
For correlation analysis and multi-variate models, entire data sets for each variable (i.e., all rats) were tested with the Shapiro-Wilk test for normal distribution. Microglial activation and neurogenesis entire data sets (n = 32) failed the normality test, and therefore a Spearman's correlation analysis was estimated using GraphPad Prism 7.0 (GraphPad software, USA). A heat-map was obtained for correlation coefficients and p values. Further multi-variate linear regressions (MLR) and linear mixed models (LMM) were performed using the Statistical Package for the Social Sciences (SPSS) to confirm the associations. For LMM, we entered the treatments, inflammation, and neurogenesis markers as fixed effects, while the day of training and the individual were considered as a random effect. The model with the best goodness of fit was selected using the smaller Conditional Akaike Information Criterion (cAIC).
Results
MS180 did not affect body weight on the day of surgery (CONT: 390.9 ± 6.7 g, MS180: 401 ± 6.1 g, p > 0.05). Mild TBI rats showed an increase in right and left hindlimb withdrawal reflex after a brief paw pinch (p = 0.001 and p = 0.005, respectively) as well as in righting reflex after the cessation of anesthesia (p = 0.04) when compared with Sham controls. Two-way ANOVAs for the four groups indicated a significant effect of mTBI on the time required for right (F[1,35] = 11.7, p = 0.001) and left (F[1,35] = 8.14, p = 0.004) hindlimb withdrawal reflex after a brief paw pinch, and for the righting reflex after the cessation of anesthesia (F[1,35] = 4.3, p = 0.044). No effects of stress or interaction were observed
Multiple comparisons revealed that there were no differences among the mTBI groups in hindlimb withdrawal reflex (right paw reflex: CONT + mTBI: 196.9 ± 14.0 sec, MS180 + mTBI: 182.0 ± 24.1 sec, p> 0.05; left paw reflex: CONT mTBI = 209.3 ± 16.0 sec, MS180 mTBI: 191.7 ± 25.3 sec, p > 0.05), or in righting reflex (CONT + mTBI: 255.6 ± 18.7 sec, MS180 + mTBI: 249.8 ± 17.2 sec, p > 0.05) after the cessation of anesthesia. The lack of significant differences in the neurological indices suggests that both CONT and MS180 groups experienced an equivalent level of injury and anesthesia.
No differences were observed in reflexes for Sham rats (right paw reflex: CONT: 139.7 ± 9.1 sec and MS180: 119.1 ± 17.2 sec, p > 0.05; left paw reflex: CONT: 154.4 ± 19.2 sec and MS180 = 125.7 ± 17.9 sec, p > 0.05; and righting reflex: CONT: 201.9 ± 31.3 sec and MS180: 189.3 ± 39.0 sec, p > 0.05). One MS180 + Sham rat presented a post-surgical infection and was excluded from the study. There were no additional health issues with the rats as they all gained weight over time and reached the visible platform in under 120 sec, which is indicative of normal visual acuity.
Gross motor assessment: beam-balance
One rat was excluded from the beam balance analysis due to it being too heavy and unable to sustain its own weight in the beam during the training. A three-way repeated measures ANOVA revealed a significant effect of injury (F[1,34] = 19.8, p < 0.0001) and day (F[4,136] = 3.71, p = 0.008), as well as injury × time interaction (F[4,136] = 5.1, p = 0.003), but not of stress or other interactions (Fig. 1B). The post hoc analysis revealed that both CONT and MS180 mTBI groups were significantly impaired relative to Shams (p < 0.001). TBI groups showed a significant motor impairment that improved over the course of the 5 days of testing (p = 0.02). Post hoc analysis showed that although no main differences were observed between CONT and MS180 injured groups (p > 0.05), only CONT + mTBI rats significantly decreased their time on the beam on Day 1 of training when compared with Sham (Fig. 1B).
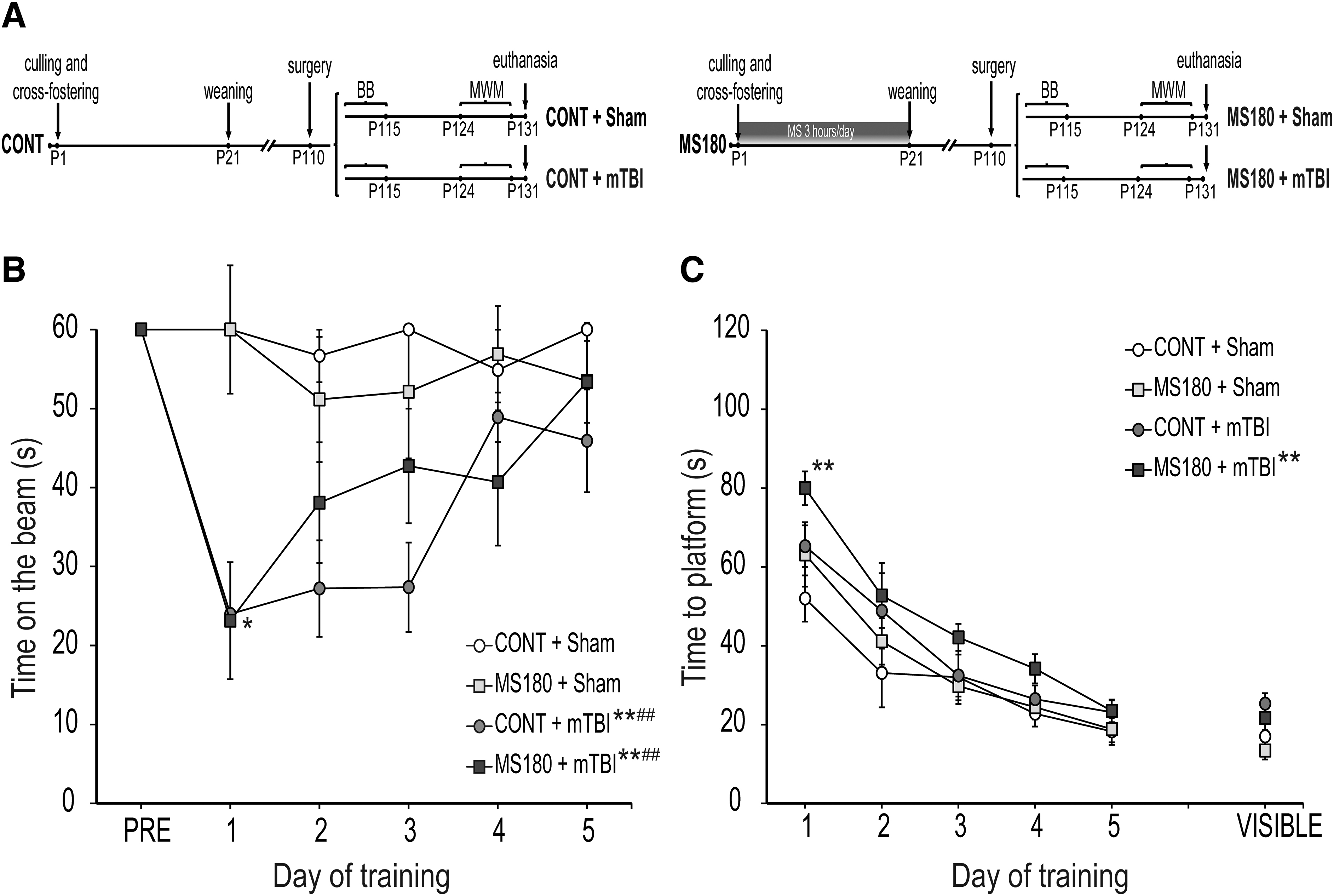
Mild traumatic brain injury (mTBI) affected motor performance regardless of early life condition; however, early life stress (ELS) and mTBI synergistically affected cognitive performance in the Morris water maze (MWM).
Cognitive performance: acquisition of spatial learning and probe trial
A repeated measures three-way ANOVA for the acquisition phase of the MWM (Fig. 1C) indicated a significant main effect of injury (F[1,35] = 8.5, p = 0.006) and training day (F[4,140] = 47.3, p < 0.0001), but not of stress or interactions (p > 0.05). Multiple comparisons for the main treatment effect revealed that only MS180 + mTBI rats exhibited cognitive deficits as evidenced by significantly longer time to locate the escape platform compared with the Shams (p = 0.009). Additionally, MS180 rats showed a significantly longer time to locate the escape platform at Day 1 compared with the Shams (p < 0.01), thus suggesting that MS180 and mTBI synergistically affect spatial learning in this task (Fig. 1C). There were no differences between CONT and maternally separated Sham rats, or between CONT + Sham and CONT + mTBI rats in the acquisition phase of the MWM (p > 0.05).
On P132, the platform was removed, and the percent time spent in the target quadrant was calculated. There was no effect of treatments on memory retrieval (CONT + Sham: 24.5 ± 4.6 %, MS180 + Sham: 25.1 ± 6.7 %, CONT + mTBI: 21.4 ± 4.3 %, MS180 + mTBI: 21.4 ± 5.6 %, p > 0.05). On the same day, a visible platform assay was performed to determine visual accuracy (Fig. 1B) and swimming speed. There were no effects of treatments in any of these parameters (p > 0.05).
Microglial activation: CEA/cell body ratio and percent of activated cells
We quantified the covered environment area (CEA)/cell body ratio and estimated the proportion of Iba1+ cells that presented an activated phenotype as markers of inflammation. Two-way ANOVA showed a significant effect of TBI on the CEA/cell body ratio (Table 1) of the ipsilateral cortex (F[1,28] = 34.8, p < 0.0001), hippocampal CA1 (F[1,28] = 36.9, p <0.0001), and hilus subfields (F[1,28] = 37.8, p < 0.0001), but no effect of stress or interaction (p > 0.05). Multiple comparisons revealed that all TBI groups significantly decreased the CEA/cell body ratio of Iba1+ cells when compared with CONT + Sham (p <0.01) and MS180 + Sham (p <0.05). No significant effects were observed for the CEA/cell body ratio of Iba1+ cells in the contralateral hemisphere (Table 1; p > 0.05)
Microglial Morphological Analysis: CEA/Cell Body Ratio of Iba1+ Cells
Mean ± standard error of the mean, analysis of variance * p < 0.05, ** p < 0.01 and *** p < 0.001 vs. CONT + Sham; # p < 0.05, ## p < 0.01, and ### p < 0.001 vs. MS180 + Sham; n = 8.
mTBI, mild traumatic brain injury.
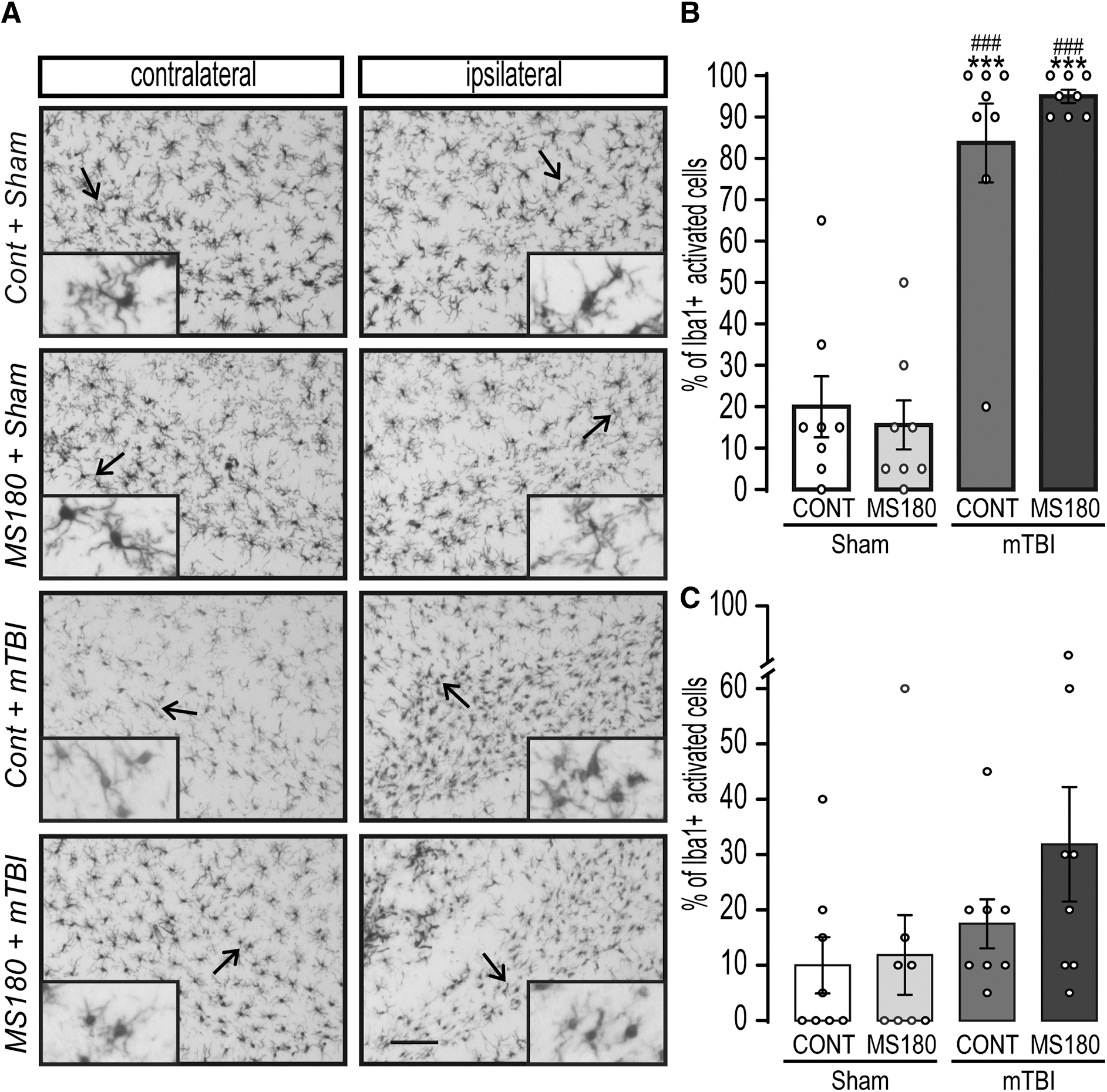
A two-way ANOVA for the proportion of Iba1+ cells that presented an activated phenotype showed a significant effect of injury in the ipsilateral CA1 subfield (F[1,28] = 58.64, p < 0.0001; Fig. 2B) and stress × mTBI interaction (F[1,28] = 6.5, p < 0.016). Multiple comparisons revealed that both CONT + mTBI and MS180 + mTBI rats showed a significant increase in the percentage of activated Iba1+ cells in the CA1, when compared with CONT + Sham rats (p < 0.0001), and this effect was significantly higher in MS180 + mTBI rats (p = 0.023). A significant effect of stress was observed in the contralateral CA1 subfield (F[1,28] = 6.7, p = 0.014; Fig. 2C), and multiple comparisons indicated that only MS180 + TBI rats showed an increase in the proportion of bushy and ameboid Iba1+ cells compared with shams (p = 0.022). Moreover, MS180 + TBI rats were significantly different from CONT + TBI (p = 0.007).

Early life stress and mild traumatic brain injury (mTBI) synergistically increased neuroinflammation in the ipsilateral CA1 hippocampus subfield. Control (CONT) and maternally separated (180 min/day, MS180) rats were subjected to either a mild controlled cortical impact (2.6 mm, mTBI), or sham surgery on post-natal Day (P) 110. Microglial morphology was evaluated on post-injury Day 21.
Two-way ANOVAs showed a significant effect of TBI on the ipsilateral hilus (F[1,28] = 111.3, p < 0.0001; Fig. 3) and ipsilateral cortex (F[1,28] = 71.5, p < 0.0001; Fig. 4), but no effect of stress or interaction (p > 0.05). Multiple comparisons revealed that both CONT + mTBI and MS180 + mTBI rats showed a significant increase in the percentage of bushy and ameboid Iba1+ cells when compared with CONT + Sham rats (p < 0.001). No differences were observed in the proportion of activated microglial cells contralaterally (p > 0.05).
Mild traumatic brain injury (mTBI) increased neuroinflammation in the ipsilateral hilus.

Mild traumatic brain injury (mTBI) increased neuroinflammation in the ipsilateral cortex adjacent to the injury.
Neurogenesis: estimations of DCX+ cell numbers
Adult neurogenesis was evaluated by estimating the number of DCX+ cells in the dentate gyrus of the hippocampus (Fig. 5). A two-way ANOVA for the ipsilateral hemisphere (Fig. 5B) indicated there was a significant effect of injury (F[1,28] = 6.7, p = 0.014) and stress × mTBI interaction (F[1,28] = 4.2, p = 0.046). The Tukey's post hoc test revealed that MS180 + Sham, CONT + TBI, and MS180 + TBI groups showed a similar decrease in the number of DCX+ cells (p < 0.05). In the contralateral side (Fig. 5C), a significant effect of stress × mTBI interaction was observed (F[1,28] = 5.5, p = 0.025), however no further differences were observed in the post hoc test.

Early life stress caused a decrease in hippocampal neurogenesis and this effect was not exacerbated by mTBI.
Correlation analysis and linear models
A Spearman's correlation analysis was performed to determine if neurogenesis or increased neuroinflammation across the different brain structures showed a significant association with our outcome variables (Supplementary Fig. S1). As expected, the analysis indicated a positive correlation between the percentage of Iba1+ cells with amoeboid or bushy morphology in the three brain structures of the ipsilateral hemisphere and acute neurological parameters (p < 0.05).
The percentage of activated Iba1+ cells in the ipsilateral CA1 region was negatively correlated with time spent balancing on the beam on the first day (p < 0.01). Also, a positive correlation was found between time to platform during all MWM acquisition days, more pronounced initially (p < 0.01) compared with the fourth and fifth days (p < 0.05). Conversely, microglial activation in the contralateral CA1 area correlated positively with time to platform during Days 2-5 (p < 0.05). The ipsilateral hilus region showed a negative correlation with time on the beam during Days 1-3, and a positive correlation with time to platform during training. The association was stronger on Day 1 (p < 0.01) than the following days (p < 0.05). For the contralateral hilus, microglial activation showed a negative link with the first beam balance day and a positive one with platform time during MWM Days 1 and 2 (p < 0.05). Interestingly, microglial activation in the ipsilateral cortex was positively linked during the initial three training days and shifted to the contralateral cortex on Days 4 and 5 (p < 0.05).
In contrast to microglial activation, the number of DCX+ cells in the ipsilateral hemisphere positively correlated with the time on the beam on the first day (p < 0.05) and was negatively linked to the second day of MWM training (p < 0.05). Conversely, DCX+ cell count in the contralateral hemisphere only correlated positively with the second and third days of beam balance (p < 0.05).
Type III ANOVA of fixed effects in LMM indicated a significant effect of TBI (F[1,155.5] = 41.12, p <0.001), but not of stress (p > 0.05) on the time on the beam in the beam balance task. A tendency for a significant effect was observed for the TBI × Stress interaction (F[1,155.5] = 41.12, p < 0.051). The ANOVA for the fixed effects in the LMM for the MWM showed a significant effect of TBI (F[1,80.5] = 5.51, p = 0.021), but not of stress or interaction (p > 0.05). The individual fixed effects estimations are presented in the Supplementary Table S1. MLRs (Supplementary Table S1) indicated that TBI significantly predicted for microglial activation in the ipsilateral CA1 (adjusted R2 : 0.595, p < 0.001), hilus (adjusted R2 : 0.783, p < 0.001) and cortex (adjusted R2 : 0.696, p <0.001); while in the contralateral side a significant effect was observed only for the CA1 region (adjusted R2 : 0.110, p = 0.036).
We further assessed the relationship between behavior, plasticity, and inflammation by adjusting a series of LMM including DCX+ cell number and Iba1+ cell activation as covariates (Supplementary Data S1). For the beam balance test, type III ANOVA of fixed effects from the best fitted model suggested a significant effect of TBI (F[1,153.46] = 31.78, p <0.001) and the interaction between TBI and the proportion of activated microglial cells in the ipsilateral CA1(F[1,153.46] = 4.13, p = 0.018). The individual fixed effects estimations are presented in the Supplementary Table S2. While for the MWM we observed that the model with the best goodness of fit (Supplementary Data S1) included a significant interaction between TBI and the proportion of bushy and activated Iba1+ cells in the ipsilateral CA1 (F[2,52.69] = 6.25, p = 0.004), the interaction between TBI and the proportion of bushy and activated Iba1+ cells in the contralateral CA1 (F [2,52.69] = 9.09, p < 0.001), and the triple interaction involving TBI, proportion of bushy and activated Iba1+ cells in the ipsilateral and contralateral CA1 (F [1,52.69] = 8.04, p = 0.006). Also, the model included a significant interaction between the ipsilateral and contralateral CA1 regions (F [1,52.69] = 11.17, p = 0.002). Additionally, we observed that the interaction between stress and TBI did not demonstrate a significant effect (F [3,52.69] = 1.9, p > 0.05); however, it did contribute to reduce the AIC coefficient and improving model fitness (Supplementary Data S1). The individual fixed effects estimations are presented in the Supplementary Table S2.
Discussion
Adverse childhood experiences (ACE) are forms of chronic childhood stress that increase the vulnerability to subsequent stressors later in life, 40,41 and are associated with a higher risk of experiencing an mTBI. 5 Moreover, it is known that a history of stressful events may predispose persons with mTBI to have poor outcomes. 7 In this study, we evaluated for the first time the relationship between ELS and mTBI using a well-validated CCI injury model, as well as their relationship with hippocampal neuroinflammation and neurogenesis. The data show that ELS and mTBI synergistically affect the acquisition of spatial learning evaluated in the MWM. However, this synergistic effect was only observed when analyzing the proportion of activated microglia in the CA1 hippocampus subfield but not when analyzing hippocampal neurogenesis and microglial activation in the cortical area adjacent to the lesion and the hilus.
The MWM is a task that has been widely used to assess spatial learning and memory in rodents, and the deficits observed in this task are considered a strong correlate to the memory impairments observed in humans post-TBI. 31,34,42,43 In the present study, we used an experimental strategy specifically designed to determine if there is a synergistic effect of both factors on cognitive performance evaluated in the MWM. On one hand, we chose to use an mTBI model (2.6 mm/5 mm tip/no angle) as it is not severe enough to mask the potential additive effect of stress and avoids the ceiling effect. Consistent with observations in humans, 6,44 -46 CONT + mTBI rats showed spontaneous recovery, as mTBI only caused motor impairments during the first three days post-injury but did not have any deleterious effects on spatial learning during the chronic phase. However, in agreement with our prior results in juveniles, 23 the subgroup previously exposed to ELS did show deficits in the acquisition of spatial learning. It is important to note that, unlike what is observed in pediatric TBI, we did not observe any effect on the probe trial in this study, which confirms the idea that the developing brain is more vulnerable to the combined effect of ELS and brain injury than the adult brain.
We chose to perform MS during the first 3 weeks of life (P1-21) to increase the deleterious effects of stress on hippocampal development. This is because the second and third weeks of life in rodents are a critically vulnerable time-window during which the hippocampus undergoes a delicate process of dendritic refinement and synaptic pruning that makes it highly sensitive to environmental influences. 47 –49 Further, this methodological design allowed us to directly compare the results with our previous data obtained in pediatrics and thus more finely determine the effect of age.
The long-term effects of ELS on hippocampal-dependent memory across different ELS rodent models are controversial, 50 as some groups report deleterious effects on the MWM 49 and others do not report any effect. 52 Previous work shows that the timing of MS differentially affects MWM performance in adults. 53 MS180 during the first week of life does not affect cognitive performance but does cause alterations when MS180 occurs during the third week of life (P14-P21). In the present study, we separated pups from P1 to 21 and did not find any effect of MS180 on spatial learning, which confirms the hypothesis that the timing and duration of the stressor can affect behavior differentially. Consistent with this idea, Sanchez and colleagues studied the relationship between ELS and TBI using a fluid percussion brain injury model; however, unlike our results, no synergistic effect of ELS with TBI was observed in the learning phase of the MWM. 52 Nevertheless, it is important to note that in that study the authors did not observe the increase in baseline corticosterone levels that is normally associated with MS form P1-14, 15 –17 and only observed an elevation when evaluating rats 90 min after subjecting them to immobilization stress. 54
Microglial cell activation increased in the cortex adjacent to the injury, as well as the ipsilateral hippocampus (CA1 and hilus) in both control and stressed rats. Importantly, MS increased both the ipsilateral and contralateral CA1 microglial activation after mTBI. This microglial activation pattern coincides with that reported in juvenile rats. 23 Also, in congruence with previous results, under basal conditions, no differences were observed between CONT and MS180 rats, and increases in Iba1+ immunoreactivity were only observed after a secondary challenge/injury. 28,37,55 Here, we utilized Iba1 immunostaining for the evaluations of microglial activation, instead of employing molecular methodologies such as cytokine quantification. Accumulating evidence shows that “primed” microglial metabolic and transcriptomic profiles are associated with cytoskeletal remodeling, which supports both pro-inflammatory and anti-inflammatory roles in both physiological and pathological conditions. 55 –58
Moreover, cell number and morphology assessed in Iba1 immunostaining, but not protein expression, can serve as a sensitive marker for detecting microglial activation, as it efficiently correlates with activation markers such as CD11b. 59 In this study, we evaluated microglial cell activation based on phenotypical changes in Iba1+ cells. To achieve this, we conducted a detailed characterization of cell morphology, allowing us to establish a 99% confidence threshold to distinguish cells with an activated phenotype (bushy and ameboid). Further studies should be done to confirm if increased cytokine activity is also present in MS180 + mTBI adults.
The Spearman analysis indicated a positive correlation between time to platform and microglial activation in the ipsilateral CA1 region, which is more pronounced during the initial three days of the test. Additionally, the contralateral CA1 region showed an association during Days 2-5. These associations were further confirmed through an LMM, where we observed that the best-fitting model includes the interaction between the lesion and the proportion of bushy and ameboid Iba1+ cells in the CA1 regions of both hemispheres. These interactions suggest that the impact of TBI on spatial learning might be modulated by microglial activation in both hemispheres. Notably, although the interaction between mTBI and stress did not exhibit a significant effect in the model, removing these variables reduced the goodness of fit. This implies that these variables are essential for capturing the complexity of the effects observed within a biological context. Taken together, these findings suggest that the effect of mTBI on microglial activation is not constant, but rather influenced by stress, particularly in the contralateral CA1 region, supporting the hypothesis that the increased inflammation resulting from the combination of mTBI and stress could underlie the observed effects on learning. The differential microglial activation within brain areas observed here is supported by studies showing a variance in immunophenotypes between cortical and hippocampal regions in adult mice. 60
Clinical studies have demonstrated that under normal conditions, brain injury symptoms often subside within days or weeks after the injury; however, a subgroup of patients exhibit persistent cognitive deficits. 44,45 It has been proposed that this heightened vulnerability could be attributed to prior exposure to ELS. 6 In line with this, our data revealed that under control conditions, the rats effectively recover from motor deficits caused by mTBI and do not exhibit cognitive impairments. Nevertheless, MS180 negatively impacts spatial learning after an mTBI. Previous reports utilizing a focal brain injury model showed that, ELS caused an increase in microglial activation in remote areas of the brain that was consistent with a primed phenotype. 20 In congruence, we observed that mTBI increases microglial activation in the ipsilateral CA1 region of CONT rats, but not in the contralateral region; conversely, in MS180 rats, we observed enhanced microglial activation in both hemispheres.
The LMMs indicated that mTBI and stress interact with the differential inflammatory response between the injured and non-injured sides to explain the alterations observed in the MWM. Collectively, our findings lead us to hypothesize that under control conditions, TBI leads to heightened neuroinflammation on the ipsilateral side; however, as the contralateral side remains intact, it could compensate for the deficits caused by the injury and enable recovery. Conversely, MS triggers microglial priming, rendering the brain more reactive and causing abnormal microglial activation in both the ipsilateral and contralateral sides. Consequently, the contralateral side is no longer capable of compensating for the injured hemisphere, resulting in cognitive deficiencies. While LMMs provide valuable insights, they also have limitations. LMMs involve numerous parameters, including fixed effects, random effects, and various interactions that can complicate the determination of precise causal relationships between variables. Further studies are needed to confirm our hypothesis. Nevertheless, our hypothesis finds support in previous research that demonstrated that contralateral hippocampal function is inhibited after cerebral ischemic injury; however, environmental enrichment (EE) can promote plasticity in the contralateral hippocampus to promote cognitive function. 59
Microglial activation is associated with neuroinflammation, decreased neurogenesis, and abnormal neuronal circuits. 62 Resting microglia support neuronal activity by controlling synaptic pruning, neuronal excitability, debris removal, and by providing trophic support. 62,63 Moreover, previous studies have shown that cognitive improvement induced by EE after a moderate TBI correlates with a decrease in microglial activation in the hippocampus CA subfield. 64 This connection between microglial activation and cognition relates to alterations in synaptic plasticity by upregulation of genes associated with inflammation and long-term potentiation. 65
In contrast to our previous report in juveniles, 23 we observed that ELS decreases the number of DCX+ cells that survive in the hippocampus, and this effect is not exacerbated by mTBI, which does not support our hypothesis. Previous works have shown that MS decreases hippocampal neurogenesis in adult males. 16,19,37,66 In the present study, we observed a main effect of ELS on the number of DCX+ cells contralaterally, but not ipsilaterally; and the previously reported differences in the ipsilateral hemisphere could only be confirmed by performing a simple comparison equivalent to that of previous studies. 16 Moreover, this effect was not further increased by mTBI, thus suggesting that ELS affects the hippocampus neurogenic niche down to a lower threshold that cannot be surpassed. Additionally, the lack of an additive effect on neurogenesis indicates that microglial effects are not associated with neuroblast apoptosis or decreased proliferation.
Our study is not without limitations, and one of them is that we did not include female rats in our evaluations at this stage. This decision was based on the controversial nature of the effects of MS on female rats, as several studies have indicated that MS has no effects on female offspring. 67 –70 Specifically, female rats are resilient to the behavioral effects of MS and exhibit stress-induced neurogenesis. 71 This sexual dimorphism deserves attention by itself; therefore, our study aimed to conduct an initial characterization of the synergistic effects of MS and mTBI using only male subjects, as the effects of MS in this sex have been consistently reported. Further studies are necessary to determine if female rats also demonstrate resilience to the synergistic effect of MS + TBI, similar to their resistance to MS alone.
In summary, our findings suggest that although all mTBI rats gain motor recovery during the post-injury phase, ELS and mTBI synergistically affect cognitive performance in the chronic phase. We theorize that ELS causes a chronic activation of microglial cells after an mTBI that is specific to the CA1 hippocampus subfield and that is only evidenced after mild injuries. This effect could account for the worse outcomes observed after mTBI in humans with a history of ELS.
Transparency, Rigor, and Reproducibility Summary
The study and analysis plan were registered prior to beginning data collection with the National Research Committee of the Mexican Social Security System (R-2019-785-060). 1,2,3 A sample size of 10 rats per group was planned for behavior, and eight rats for histology were estimated using the finite population formula and based on previous studies. 1,2,3 The progeny of 10 time pregnant rats were cross-fostered, culled to eight pups (4-6 males), and randomly assigned to groups. 4,5 Forty male rats were randomly assigned to trauma versus sham groups using a random number generator and were blinded. 4,5,6 One rat presented a post-surgical infection and was excluded from the study. 4 All behavioral analyses and histological materials were performed by investigators blinded to group and trauma status. 6 All rats were tested in the animal facility under standard light and temperature conditions between 9:00 AM and 1:00 PM in a fed state. 7 Histological analyses were performed in eight batches, with one sample from each group randomly assigned to each batch. 8 Specificity of antibodies used for histology was verified by suppliers and confirmed with a negative control. 8 Normal distribution of data was confirmed using the Shapiro-Wilk test. 9 The sample sizes and degrees of freedom reflect the number of independent measurements (rats). 9 Non-independence of measurements was addressed using repeated measures ANOVAs and linear mixed models. 9 Data will be made available on request. 13 The authors agree to provide the full content of the manuscript on request by contacting the corresponding author. 15
Data Availability
Data will be made available on request.
Footnotes
Authors' Contributions
Ana Fernanda Salinas-García: Investigation, Data Curation, Visualization, Writing - Original Draft. Angélica Roque: Investigation, Data Curation, Visualization. Jonathan Zamudio-Flores: Investigation, Data Curation. Esperanza Meléndez-Herrera: Supervision, Conceptualization, Writing—Review and Editing. Anthony E. Kline: Methodology, Writing - Review and Editing. Naima Lajud: Project administration, Supervision, Formal analysis, Resources, Writing—Review and Editing, Funding acquisition.
Funding Information
This work was supported by a grant from Coordinación de Investigación en Salud- Fundación IMSS (NL). AFSG and JZF are graduate students in the Programa de MaestríaenCienciasenEcologíaIntegrativa—Universidad Michoacana de San Nicolás de Hidalgo and were supported by CONACYT (CVU 805864 and 1187161).
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Figure S1
Supplementary Table S1
Supplementary Table S2
Supplementary Data S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.