
Abstract
This study utilized the Noldus PhenoTyper Home Cage Monitoring system (HCM) to assess the behavioral and cognitive changes of experimental closed-head mild traumatic brain injury (mTBI). Seventy-nine adult male Institute of Cancer Research (ICR) mice were subjected to either a sham procedure or closed-head mTBI using the weight-drop model. Seven days post-injury, separate cohorts of mice underwent either a non-cognitive or a cognitive home cage assessment, a treadmill fatigue test, or the Open Field Test. mTBI significantly influenced habituation behavior and circadian wheel-running activity. Notably, mTBI mice exhibited an increased frequency of visits to the running wheel, but each visit was shorter than those of controls. No significant differences between the groups in discrimination or reversal learning performance were observed. However, during the reversal learning stage, mTBI mice performed similarly to their initial discrimination learning levels, suggesting an abnormally faster rate of reversal learning. Home cage monitoring is a valuable tool for studying the subtle effects of mTBI, complementing traditional assays. The automated evaluation of habituation to novel stimuli (e.g., novel environment) could serve as a potentially sensitive tool for assessing mTBI-associated behavioral deficits.
Introduction
Mild traumatic brain injury (mTBI) comprises ∼90% of all brain injury cases. 1 Despite the prevalence of mTBI among patients with brain trauma, its diagnosis remains challenging, even with imaging methods such as computed tomography (CT) or magnetic resonance imaging (MRI). mTBI patients may experience a broad range of short- and long-term behavioral impairments. These impairments include cognitive impairments (e.g., memory loss, attention deficit 2 ), emotional difficulties (e.g., anxiety and depression 3 ), motor impairments (e.g., balance problems, lack of coordination 4 ), and circadian rhythm disruption (e.g., alterations in sleep stages, poor sleep continuity 5,6 ). To better understand these impairments and find viable treatments, various animal models are used in studies investigating the consequences of mTBI, with rodent models being the predominant choice (see the review by Shultz and coworkers 7 ).
The weight-drop model is one of the most widely used closed-head mTBI models. 8 This model replicates key facets of mTBI, such as cognitive and motor impairments, and is a relevant tool for studying this injury. 7 In previous work, from our laboratory and others, we have thoroughly studied behavioral alteration in the weight-drop model. For example, cognitive evaluations of mTBI mice revealed impairments in spatial and non-spatial learning and recognition memory. 8 –10 Additionally, assessments of psychiatric phenotypes in mTBI mice revealed behaviors consistent with depression 11 and anxiety. 12 In contrast, motor assessments demonstrated intact motor performance and no balance impairments in mTBI mice, possibly as a result of compensatory mechanisms. 13 Nevertheless, the above-described results were obtained through short-term standard evaluation tests. Although these tests offer valuable insights and serve as a reference point for novel discoveries, they also have inherent limitations. 14 Particularly, they often require assessing mice under stressful conditions (e.g., unfamiliar arenas, human handling, and shock-induced learning) and for brief periods (e.g., a few minutes). As these assays provide only a snapshot of behavior at a specific point in time, they may not accurately capture the full spectrum of natural behaviors or the progression and recovery of symptoms over time.
However, these limitations can be overcome using automated sensor-based monitoring methods that track mouse behavior within their home cage environment. 15,16 Home cage monitoring (HCM) systems, a complementary approach to behavioral phenotyping techniques, have gained popularity in recent years because of their ability to record multiple parameters across the circadian cycle. 17 Various technical solutions are currently being tested and used for HCM, including video tracking, infrared light beams, floor sensors, radio frequency identification system (RFID) technology, and combinations of these technologies. 15 A particular advantage of these approaches is that they facilitate the quantification of behavior at multiple time scales, such as habituation within a single test session and fluctuation over time. 18 Further, they enable the measurement of mouse activity during active phases (feeding, exploration) and inactive phases (sleeping, resting) of the circadian cycle and bouts of activity within these phases. In the context of mTBI, the comprehensive “round-the-clock” data provided by HCM systems can offer valuable insights into the animal's overall well-being. 19,20
Despite the potential of HCM systems, little research has been done on home cage behavior in the mTBI mouse model, in particular the closed-head model. 16 However, the studies that have been conducted have reported several interesting phenotypes across a range of research methodologies.
For example, Pinkowski and coworkers 21 assessed the impact of repeated mTBIs on general activity and the light/dark cycle and found that injured mice exhibited reduced home cage activity. Similarly, Vu and coworkers 22 examined the effects of two prevalent mTBI models (concussion and blast injuries). They found that both injury types resulted in acute but transient hypoactivity, as evidenced by reduced home cage wheel running, eating, and drinking levels. A contrary finding was reported by Chen and coworkers, 23 who found that the distance traveled by injured mice was similar to that of control mice. However, injured mice displayed impaired discrimination learning and slowed cognitive flexibility. Likewise, in a study by Siedhoff and coworkers, 24 blast mTBI and control mice showed similar distance traveled and phase transition response. Nevertheless, the authors found abnormal inter-session habituation, as evidenced by shorter shelter visits during the third active phase, suggesting anxiety-like behavior. In previous work from our laboratory, anxiety was measured using an HCM system with a spotlight test, providing novel findings. mTBI mice have difficulty recovering from light stressors, exhibiting residual avoidance behavior, as indicated by a reduction in cage exploration after exposure compared with before and during exposure. 25
In summary, HCM offers the benefit of continuous, long-term monitoring of mouse behavior in a stress-free and natural environment. This way, subtle behavioral changes can be identified and mapped, discovering previously unknown phenotypes. Prior HCM-based mTBI studies demonstrated cognitive and behavioral deficits following injury across different injury models and time frames. However, the literature also indicates the need for more research in closed-head mouse models. This study used the Noldus PhenoTyper HCM to assess the behavioral and cognitive outcomes of an experimental closed-head mTBI.
Methods
Animals and experimental timeline
Seventy-nine Institute for Cancer Research 26 (ICR) male mice, 6–8 weeks of age, were obtained from Envigo RMS Israel, and randomly assigned to either the mTBI or control groups (controls n = 40, mTBI n = 39). The group sizes are described under each assay in this section. Separate cohorts of mice were used for the home cage assays and non-home cage (treadmill fatigue test, open field test) assays. Additionally, to ensure the reliability and validity of the findings, separate cohorts of mice were employed for the cognitive and non-cognitive home cage assays, considering the potential impact of wheel running on motor and cognitive abilities. 27 The experimental timeline is shown in Figure 1. The housing and experimental room were kept in a reversed light cycle conditions (8:00–20:00 dark; 20:00–8:00 light). All procedures were approved by the Tel-Aviv Faculty of Medicine Ethics Committee (01-21-059) and were conducted according to the guidelines for animal experimentation of the National Institutes of Health (DHEW publication 85-23, revised, 1995).

Experimental timeline. Separate cohorts of mice were used for the different assays, including home cage and non-home cage (treadmill fatigue test, open field test) assays.
Experimental mTBI model
A weight-drop concussive brain injury device consists of a metal tube with an internal diameter of 13 mm and a length of 80 cm. Before injury induction, each mouse was anesthetized using isoflurane inhalation and was placed immediately below the device on top of a sponge maintaining its head position. At the beginning of the tube, a 30-g weight was released, and free-fell along the tube length. It struck the right lobe of the mouse's brain between the eyes and ears. Control mice underwent a similar anesthesia (isoflurane) procedure without head trauma. After recovering from anesthesia, all mice returned to their respective cages. 28
Behavioral assays
Separate groups of mice were used for behavioral assays. Testing began 1 week following mTBI induction, except for the Open Field (OF) Test, which was conducted 20 days post-injury.
Treadmill Fatigue Test
The Treadmill Fatigue Test was conducted using the Panlab 5-lane treadmill system (Panlab, Harvard Apparatus, Barcelona, Spain), as previously described. 29 In each test stage, grid stimulation intensity was set to 0.6 mA. The treadmill and grid were cleaned with Virusolve after each trial.
Treadmill acclimatization (48 h)
On the 1st day, mice (n = 10 in each group) were individually placed into separate lanes and allowed to explore the treadmill freely when the treadmill was not activated. After 3 min, the speed increased gradually (every 60 sec increased by 2 cm\sec) to a walking rate of 10 cm/sec for 10 min. After 10 min, the treadmill was turned off, and the mice were allowed a 3-min exploration of the non-active treadmill before being removed and returned to their cage.
On the 2nd day, mice were individually placed on the treadmill with a 10 cm/sec starting speed. The speed was increased (every 60 sec, increased by 1 cm\sec) up to 15 cm/sec for an overall time of 15 min. After the treadmill was turned off, mice were allowed a 3-min exploration of the non-active treadmill before being removed and returned to their cage. There was one resting day between the acclimatization stage and the test.
Fatigue Test (24 h)
In this test, fatigue behavior was defined as 5 consecutive sec in the “fatigue zone.” The fatigue zone is the grid and the region of one mouse's body length at the end of the treadmill lanes. Mice were placed individually into separate lanes of the treadmill. The test started with the speed set at 15 cm/sec. The test began at a speed of 15 cm/sec, increased by 5 cm/sec during the 1st min, then by 4 cm/sec each min until the 6th min. The speed remained constant at 24 cm/sec from the 6th to the 30th min. From the 30th min onwards, it increased by 8 cm/sec every 15 min. When a mouse remained in the fatigue zone for more than 5 continuous sec, it was removed from the treadmill.
Outcome measures
Run time, running velocity at the end of the test, and grid stimulation frequency were recorded and analyzed.
OF Test
The OF Test systematically evaluates novel environment exploration and overall locomotor activity. 30 Additionally, it serves as an initial screening method for detecting rodent anxiety-like behavior. 13 Mice (mTBI n = 9 and control n = 10) were placed individually at the center of a plexiglass arena designed for open-field testing (60 × 60 × 20 cm), and their behavior was recorded over 5 min.
Outcome measures
We extracted the average activity within the arena and the frequency of in-zone visits.
PhenoTyper Home Cage System
PhenoTyper Home Cage System (PhenoTyper model 3000, Noldus Information Technology, The Netherlands) was used for video-based observation. The cages are made of transparent Perspex walls with an opaque Perspex floor covered with cellulose bedding. Each cage (L = 30 × W = 30 × H = 35 cm) consists of a top unit with built-in hardware for infrared-sensitive video tracking, ensuring constant and even illumination. An infrared filter placed in front of the camera prevents room illumination interference. This method allows continuous behavioral recordings in both dark and light periods. EthoVision 15XT was used for video tracking (15 samples/sec) and trial control software. A total of 8 PhenoTyper cages were used simultaneously, with video tracks recorded by two computers (four PhenoTypers per PC running video tracking and trial control software).
Experimental setup
Separate cohorts of mice were assigned to either the PhenoTyper cages general well-being method for habituation and circadian behavior assessment or the CognitionWall method for operant learning assessment. All recordings began at 8:00, 1:00 h in system time, at the start of the active dark phase. In all experimental setups, each cage was equipped with bedding, a drinking bottle, and a shelter (height: 10 cm, diameter: 9 cm; infrared transparent material). As part of the habituation assessment, the cage had a feeding station and running wheel (diameter: 15 cm, W: 7 cm; PhenoWheel, Noldus, Wageningen, Netherlands). For the operant learning assessment, a reward food pallet dispenser (H = 25 cm, W = 17 cm, diameter holes = 3.3 cm) replaced the feeding station, and an operant wall (CognitionWall, Noldus Information Technology, Wageningen, The Netherlands) replaced the running wheel.
Habituation and circadian behavior (PhenoTyper)
Control and mTBI mice (n = 8 in each group) underwent a 72-h assessment of short-term (intra-session) habituation, long-term (inter-session) habituation, and overall circadian behavior.
Data were recorded and evaluated at both the temporal and spatial levels to better understand home cage behavior dynamics.
Spatial organization
The home cage arena was virtually divided into four zones for activity recording: running wheel, shelter, food, and water zones.
Temporal organization
Behavioral patterns were analyzed within and across days, phases, and hours. For this purpose, three temporal segments were examined: short-term habituation (within day 1, across hours 1–3), long-term habituation (across days 1–3, across light-dark phases), and overall circadian behavior (within day 3, across light-dark subphases).
Based on previous research, 31 –33 data were segmented into two 12-h periods: an active phase from 01:00 to 12:00 and an inactive phase from 13:00 to 24:00 (system time). To gain a deeper understanding of wheel-running behavior on the third home cage day, the light and dark phases were further segmented into 2-h subphases. The active subphases are A1 (dark onset; 1:00–3:00), A2 (mid-dark; 4:00–6:00), A3 (late dark; 7:00–9:00), and A4 (anticipation for light onset; 10:00–12:00). The inactive subphases are IN1 (light onset; 13:00–15:00), IN2 (mid-light; 16:00–18:00), IN3 (late light; 19:00–21:00), and IN4 (anticipation for dark onset; 22:00–24:00).
Outcome measures
Based on previously published methods, 24,34,35 we extracted data on total activity, total distance traveled (cm), total wheel revolutions, frequency of visits to each zone, and duration of time spent in each zone (sec). We computed the bout length (sec), estimated running speed (cm/sec), and habituation change from these raw measures.
Briefly, the bout length, or the duration of visits to the wheel zone, was calculated by dividing the duration spent in the wheel zone by the frequency of visits to the wheel zone when the frequency was greater than one. The estimated running speed was determined by dividing the running distance by the duration spent in the wheel zone, provided that the wheel revolutions were greater than or equal to one. It is important to note that running speed is an estimation rather than an exact measure. However, it contributes to a more holistic understanding of wheel-running characteristics. The habituation change index was separately calculated for the dark and light phases across days 1–3. This index was computed using the following formula: habituation change index = [(value mTBI - value control) / (value mTBI + value control)]. Positive values indicate increased behavior in the mTBI group relative to controls.
Discrimination learning (DL) and reversal learning (RL) (PhenoTyper)
The CognitionWall operant learning test was administered on 12 control and 12 mTBI mice. Following exclusions resulting from technical issues, the final analysis of DL and RL performance comprised eight controls and seven mTBI mice. As part of the Fixed Ratio 5 schedule of reinforcement, mice learned to receive a food pellet reward (14 mg Dustless Precision Pellets; Bio-Serve) for every fifth entry to the designated CognitionWall entrance. The paradigm did not involve food deprivation or a restriction on the number of food pellets mice could earn. In cases in which mice received <100 food pellets, additional food pallets were provided to ensure that a total of 100 pallets were delivered. Water was available ad libitum throughout the protocol. The learning protocol consisted of three stages, namely habituation, DL, and RL.
Habituation (24 h)
A 1-day habituation period preceded the DL and RL stages. The mice were initially trained to approach the cognition wall by receiving one food pellet per entering any of the three holes. Their habituation behavior during the initial 3 h was analyzed.
DL (48 h)
In this stage, mice learned to earn food by entering the left CognitionWall hole. Subsequently, mice were returned to their original non-automated cages for 1 week.
RL (48 h)
Seven days following the initial DL, mice were returned to the PhenoTyper cages. In this stage, the correct hole was switched, and mice learned to earn food by entering the right CognitionWall hole. The time taken to achieve the RL criteria indicates cognitive flexibility.
Outcome measures
For the habituation stage, we extracted data on total activity, total distance traveled (cm), and the time spent in shelter and CognitionWall zones (sec). For the DL and RL phases, learning was assessed using a criterion of 80% correct hole entries and a chance level of performance set at 33%. We used a moving window approach to determine the number of entries required to achieve the learning criterion. This calculation considers the ratio of correct to incorrect entries within the last 30 entries rather than the total number of entries or the time required to reach the criterion.
Statistical analyses
RStudio was used for data analysis. We used linear mixed effects (LME) models to study short-term habituation and wheel-running behavior across different subphases. In these models, we treated the experimental group and subphase as fixed effects and the animal identifier as random effect to account for repeated measurements. The LME models were fitted using the R lmer4 and lmerTest packages. 36 The emmeans 37 R package was used for multiple comparisons with Sidak's correction when significant group-by-time interaction effect was found. One-sample t tests with Holm correction were used to study long-term habituation, and dependent and independent sample t tests were used to study fatigue-like behavior, behavior in the open field, and DL and RL. All statistical tests were two-tailed, with p < 0.05 considered statistically significant.
Results
This study included a home cage assessment of habituation and overall circadian behavior, a treadmill fatigue test, OF test, and a home cage CognitionWall test. The experimental timeline can be found in Figure 1 in the Methods section. To analyze habituation and overall circadian behavior, we adopted a three-step hierarchical approach. First, we evaluated short-term habituation within the initial hours in the home cage (Short-term habituation section). Second, we examined long-term habituation profiles over days (Long-term habituation section). Lastly, we examined wheel usage patterns on the third home cage day with a more detailed intra-day resolution (Qualitative aspects of wheel running and Descriptive characteristics of wheel running bouts sections). These analyses enabled us to gain a comprehensive understanding of the temporal structure of behavior in mTBI mice. In addition to these analyses, we also assessed fatigue-like behavior (Treadmill Fatigue Test section) and OF behavior (OF Test section). Further, we investigated DL and RL within the home cage (DL and RL section), which included a habituation stage.
Short-term habituation
The first 3 h in the home cage, or short-term habituation, was assessed through a series of LME models for each dependent measure. The analysis revealed a significant main effect of group on distance moved (cm walked), indicating that mTBI mice exhibited hyperactivity by covering a greater distance than controls during this period (F[1, 43.67] = 4.78, p = 0.034). There was no significant group-by-time interaction effect (F[1, 30] = 2.04, p = 0.163), suggesting that this hyperactivity was not restricted to a specific time point, such as solely the 1st h. No significant differences were detected in other measures (all, p > 0.05). Detailed results are provided in Table S1. Overall, our findings indicate that the novelty associated with the initial introduction to the home cage triggers robust locomotor and exploratory responses in mTBI mice.
Long-term habituation
To examine changes in behavior over time in the home cage, a series of one-sample t tests were conducted using the habituation change index compared against a change index of zero. The analysis revealed significant differences between mTBI and control mice in running wheel behavior, sheltering, and cage floor activity lasting even three days after being introduced to the home cage. These differences were primarily evident throughout the active phase (01:00–12:00 system time) but were also observed during the inactive phase (13:00–24:00 system time). See Figure 2 and Table S2.

Differential habituation across days and light–dark phases in mild traumatic brain injury (mTBI) relative to control mice (n = 16). The habituation change index, shown for wheel running, sheltering behavior, and cage floor activity, is calculated as (mTBI values - control values) / (mTBI values + control values). Lower panel: active phase (1:00–12:00, system time); Top panel: inactive phase (13:00–24:00, system time). The dashed horizontal line at 0 represents no habituation effect, with higher values indicating an increase in behavior for mTBI mice relative to controls. Values are shown as mean ± standard error (SE). ***p < 0.001, **p < 0.01, *p < 0.05.
Day 1
During the first active phase, mTBI mice displayed significantly heightened activity levels compared with control mice (p = 0.047), moving more distance across the home cage floor (p < 0.001), and spending less time sheltering (p = 0.012). Additionally, mTBI mice displayed increased engagement with the running wheel, characterized by a greater number of wheel revolutions (p = 0.024), more frequent visits to the wheel zone (p < 0.001), and extended time spent within the wheel zone (p < 0.001). No differences were found in the inactive phase (all, p > 0.05).
Day 2
During the second active phase, mTBI mice visited the wheel zone more frequently (p < 0.001) and spent more time there (p = 0.007) than control mice. Additionally, the distance covered on the cage floor remained significantly greater (p < 0.001), and the duration of sheltering remained shorter (p = 0.002). Moreover, in the second inactive phase, mTBI mice continued to exhibit reduced sheltering duration compared with control mice (p = 0.008).
Day 3
Even as long as 3 days after being introduced to the home cage, mTBI mice continued to show significantly higher activity levels (p < 0.001), greater distance moved (p = 0.015), and more frequent wheel zone visits (p < 0.001) than control mice during the active phase. No differences were found in the inactive phase (all, p > 0.05). In summary, mTBI mice demonstrated persistent heightened activity levels in terms of total activity, cage-floor activity, and running wheel behavior, suggesting potential impairments in their habituation processes.
Qualitative aspects of wheel running
An in-depth analysis of wheel running behavior on the 3rd day in the home cage was conducted to profile the organization of circadian behavioral activity (e.g., anticipation and response to phase shifts). The analysis revealed several interesting behavioral phenotypes of circadian running wheel activity. There was a group-by-subphase interaction effect on the frequency of wheel zone visits (F[1, 14.02] = 2.37, p = 0.022), indicating that the effect of mTBI on the frequency of wheel zone visits varies across different subphases (see Fig. 3). To gain further insight into the group-by-subphase interaction, we conducted further between-groups comparisons and subsequent analysis within each group.
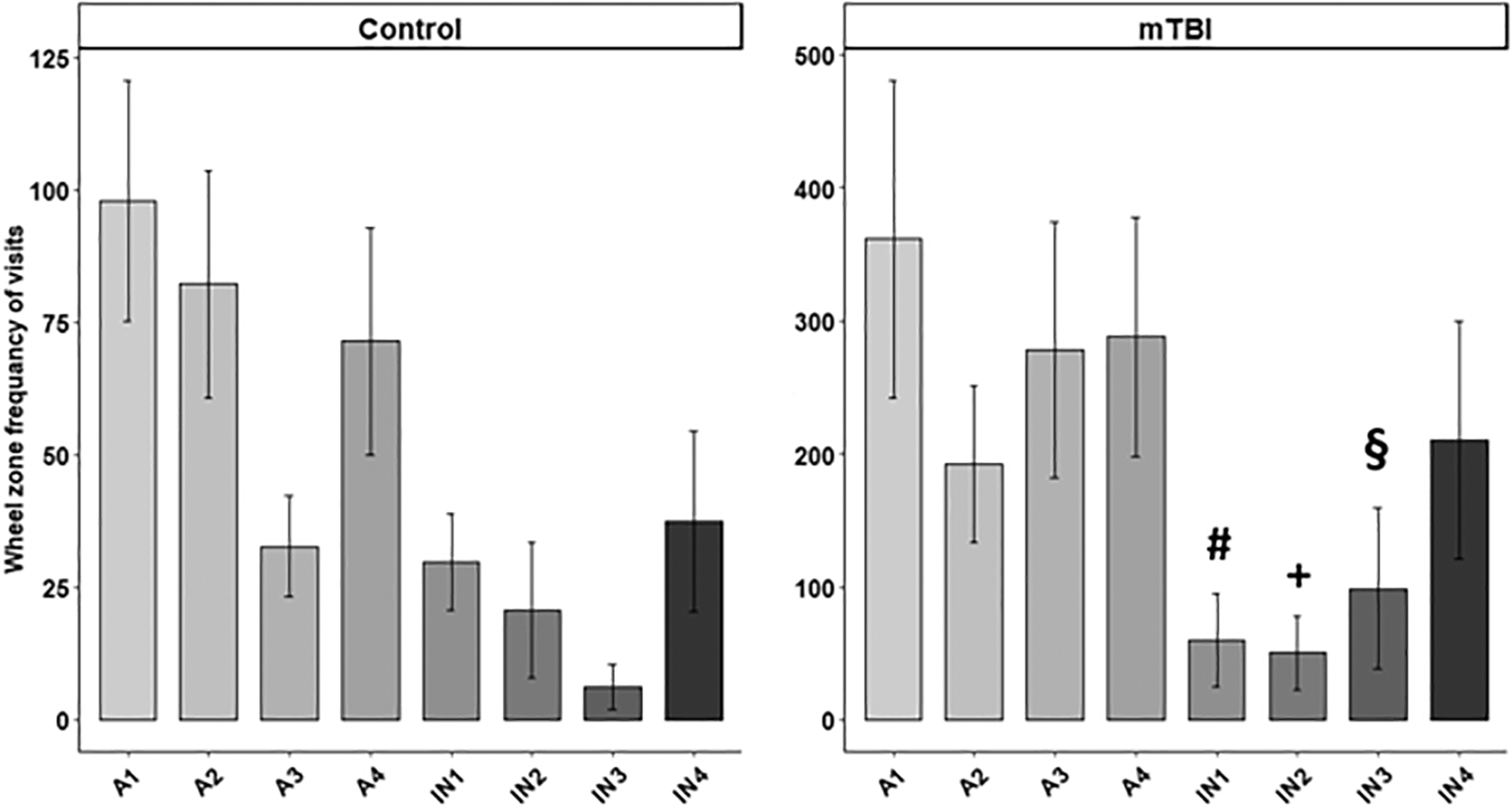
Wheel-running behavior on the 3rd home cage day in mild traumatic brain injury (mTBI) and control mice (n = 16). In mTBI mice, the frequency of visits to the running wheel zone markedly decreased with the onset of the inactive light phase, as evidenced by a significant group-by-subphase interaction effect. No Significant differences were found within the control group. Within the mTBI group, symbols denote specific differences between light–dark subphases: # denotes differences between IN1 and A1 (p < 0.001), IN1 and A3 (p = 0.009), and IN1 and A4 (p = 0.005). + denotes differences between IN2 and A4 (p = 0.003), IN2 and A3 (p = 0.005), and IN2 and A1 (p < 0.001). § denotes a difference between IN3 and A1 (p = 0.002). Values are shown as mean ± standard error (SE)
Between-group comparisons
Significant differences were found in wheel zone visits between mTBI mice and controls during the active subphases (A1–A4), with mTBI mice visiting the wheel zone significantly more frequently than controls (p values ranging from 0.020 to 0.050). However, during the inactive subphases (IN1–IN4), no significant differences were detected (all, p > 0.05).
Within-group comparisons
Analysis within each group revealed significant subphase-dependent patterns only among mTBI mice (p values ranging from 0.001 to 0.002). These patterns were characterized by a pronounced decrease in wheel zone visits in response to the onset of the inactive phase (IN1), suggesting an altered response to phase transitions in injured mice.
Descriptive characteristics of wheel running bouts
To further characterize the circadian behavioral phenotypes of mTBI mice on day 3, we focused on two aspects: the duration of wheel zone visits and the estimated wheel running speed. Because of technical constraints, direct measurements of running speeds were not possible. Therefore, we used an estimated metric to evaluate speed trends over time.
Duration of wheel bouts (sec)
On average, mTBI mice had shorter visits, ranging from 2.88 to 13.37 sec, whereas control mice had longer visits, ranging from 4.57 to 32.70 sec. However, this trend did not reach statistical significance (F[1, 201.4] = 1.02, p = 0.419). The lack of statistical significance in the relationship between injury and visit duration may imply that the relationship is complex and non-linear or not consistently influenced by the subphase. Indeed, there were variations between the two groups in the timing of when mice were most engaged with the wheel. Control mice exhibited their most extended engagement periods during the mid-late stages of the dark active phase (subphase A2: mean [M] = 32.70, standard deviation [SD] = 57.45; subphase A3: M = 30.14, SD = 49.35). On the other hand, mTBI mice exhibited their most extended engagement periods at the onset of the dark active phase and near its end (subphase A1: M = 13.02, SD = 12.28; subphase A3: M = 13.37, SD = 20.84) (see Fig. 4A).

Descriptive characteristics of the duration of visits to the wheel zone and the estimated running speed in each light–dark subphase on the 3rd day of wheel use in mild traumatic brain injury (mTBI) and control mice (n = 16). On the X-axis are
Running speed (cm/sec)
Control mice exhibited peak running intensity at the offset of the inactive light phase and the onset of the active dark phase (subphase IN4: M = 48.61, SD = 27.53; subphase A1: M = 40.32, SD = 18.05). On the other hand, mTBI mice exhibited peak running intensity during the late inactive light phase and at the offset of the inactive light phase (subphase IN3: M = 46.46, SD = 8.48; subphase IN4: M = 38.77, SD = 12.06) (see Fig. 4B).
When evaluating qualitative characteristics of wheel-running taken together, mTBI mice exhibited distinct behavioral patterns compared with control mice. Specifically, mTBI mice visited the wheel zone more frequently but for shorter durations, and their peak running speed occurred at different times of the day. These differences in the temporal profile of wheel engagement and intensity of wheel-running suggest potential alterations in the circadian behavior of mTBI mice. The findings highlight the complex nature of mTBI, particularly the anticipation and response to phase transitions and the overall timing of wheel running. Consequently, a further assessment was conducted to determine whether injury-related fatigue or diminished physical capabilities may have contributed to these changes.
Treadmill Fatigue Test
The result of the independent sample t tests revealed no significant differences between mTBI mice and controls in run times (t[1, 18] = 0.623, p = 0.766), running velocity at the end of the test (t[1, 18] = 0.049, p = 0.361), and stimulation frequency (t[1, 18] = 1.142, p = 0.863). mTBI mice showed treadmill performance similar to that of control mice; they reached equal speeds, finished the test at comparable times, and needed no additional motivation to continue running. Therefore, it is likely that the altered circadian patterns in mTBI mice are a result of factors other than their physical capabilities.
OF Test
The result of the independent sample t tests revealed no significant differences between the control and mTBI groups in average activity within the arena (t[1, 17] = 2.02, p = 0.059) and the frequency of in-zone visits (t[1, 17] = 1.44, p = 0.168).
DL and RL
The final test measured cognitive flexibility, which is the ability to extinguish previous learning and acquire a new response.
Habituation
Prior to the DL and RL stages, the mice underwent a 1-day habituation to the cage. Their habituation behavior during the initial first hours was analyzed using a series of LME models for each dependent measure. Detailed results are provided in Table S1.
DL
The analysis revealed a significant group-by-time interaction effect on total activity levels during the first 3 h in the cage (F[1, 46] = 6.76, p = 0.013). Despite no significant between-group differences (p values ranging from 0.687 to 0.963), within-group comparisons revealed significant differences in activity levels from the 1st to the 3rd h for both control and mTBI groups (p < 0.001), indicating a decrease in activity levels from the 1st to the 3rd h in the home cage. Notably, despite similar patterns of change over time, the magnitude of this decrease was slightly more pronounced in the control group (-39.53%) than in the mTBI group (-31.72%). No significant differences were found in measures other than activity (all, p > 0.05).
RL
The analysis revealed significant main effect of group on total activity (F[1,47.5] = 6.57, p = 0.014), independent of the time of day. A week following the DL stage and 18 days post-injury, upon their reintroduction to the home cage for the RL stage, mTBI mice were 23.06% more active than controls. No significant differences were found in other measures (all, p > 0.05).
Collectively, these findings corroborate the results previously discussed in short-term habituation section, demonstrating that mTBI mice exhibit restless behavior in response to environmental changes.
Entries to learning criteria
The number of entries required to achieve the DL and RL learning criteria (80% threshold) was first compared between the control and mTBI groups.
DL
Independent t test indicated no statistically significant difference between the two groups in discrimination ability (t[1, 13] = -1.00, p = 0.335). The mean number of entries to reach the DL learning criteria was 187.75 for the control group and 239.71 for the mTBI group.
RL
One week after the initial learning, no statistically significant difference was found between the two groups in reversal ability (t[1, 11] = 1.31 = -1.00, p = 0.216). The mean number of entries to reach the RL learning criteria was 444.50 for the control group and 329.71 for the mTBI group.
We subsequently compared the number of entries required to reach the RL learning criteria versus those required to reach the DL learning criteria separately within each group.
In control mice, the number of entries required to meet the RL learning criteria was significantly higher than that for the DL criteria (mean difference = 253.83, t[1, 5] = 3.80, p = 0.013). In contrast, for mTBI mice, the number of entries needed to reach the RL criteria was not significantly different from those needed to reach the DL criteria (mean difference = 83.60, t[1, 4] = 1.17, p = 0.305), suggesting that injury may have caused overly flexible learning (see Fig. 5A).

Discrimination and reversal learning in mild traumatic brain injury (mTBI) and control mice (n = 15).
Chance level performance
After confirming that the learning criterion was met, a one-sample t test against a chance level of 33% was conducted to determine whether random factors might have influenced the performance of mice in either group.
DL
The discrimination performance of control mice significantly exceeded chance level (mean performance = 77.3%, t[1, 7] = 13.1, p < 0.001). Similarly, the mTBI group also exceeded chance level (mean performance = 78.4%, t[1, 6] = 13.1, p < 0.001).
RL
The reversal performance of control mice did not differ significantly from chance (mean performance = 51.4%; t [7] = 1.90, p = 0.099). In contrast, the performance of the mTBI group was significantly above chance (mean performance = 69.0%; t[1, 6] = 9.57, p < 0.001), further highlighting a possible association between cognitive instability (specifically hyper-flexibility) and our mTBI model (see Fig. 5B).
Key findings
Overall, the results of our investigation of the mTBI closed-head mouse model using the HCM system indicate that injured mice exhibited altered habituation patterns (e.g., novelty-induced behavioral reactivity), hyperactivity, disrupted circadian rhythms, and an overly flexible learning process.
These findings not only underscore the multifaceted effects of mTBI but also demonstrate the promise of home cage assessments in detecting them.
Discussion
This study utilized an automated HCM system to assess behavioral and cognitive changes in a weight-drop closed-head mTBI mouse model. Our findings indicate that mTBI affects not only the overall pattern of running wheel activity but also the way it is performed. Specifically, we found notable behavioral differences between mTBI and control mice, including altered adaptivity and habituation to the novel home cage environment, disrupted circadian rhythm, hyperactivity, and abnormally faster RL.
Short-term and long-term habituation
We first assessed intrasession short-term habituation to better understand temporal dynamics (i.e., the initial response to the novel home cage environment). In the first 3 h within the home cage, mTBI mice exhibited restless exploration, indicating a heightened response to novel surroundings. This was evidenced by greater distance traveled and a slightly diverging pattern of total activity compared with control mice. Gradually, this initial hyper-exploration activity was accompanied by non-exploration motor activity such as wheel running. Moreover, at 18 days post-injury, mTBI mice continued to demonstrate higher activity levels than controls when returned to the home cages, which represented a semi-novel environment, for the reversal stage of the operant learning task,. On the other hand, the OF test, conducted at 20 days post-injury, did not reveal notable differences between the mTBI and control mice in activity levels or the frequency of in-zone visits.
The observation that an abrupt transition to a novel and potentially stressful environment leads to increased exploration rather than behaviors such as sheltering is consistent with earlier home cage research from our laboratory of abnormal stress responses in mTBI mice. In the study by Tseitlin and coworkers, 25 whereas short exposure to novelty (e.g., 15 min exposure to an unfamiliar open field) elicited no stress response, the presence of a stressor at the home cage (e.g., a spot of light) or an extended assessment duration (e.g., >30 min at the marble burring) elicited a stress reaction. One explanation for this distinction between stressors could be the human presence effect. Mice may hide signs of poor health from the human handler, whom they may perceive as a potential predator. 38 As a result, a stress reaction in a short standard test may elicit exploration behavior driven by the stress elicited by human presence, whereas more extended tests allow mice to focus on the environment stressor alone (e.g., novelty) and present a more focused response to the environment. In our study, the use of the HCM eliminates the human factor, allowing us to assess the effect of the novel home cage alone. Further, our assessment of long-term inter-session habituation profiles revealed that, even after 3 days in the home cage, mTBI mice continued to show higher activity levels than controls (e.g., greater distances traveled and more frequent visits to the wheel zone). These heightened activity patterns were particularly evident during the active dark phase.
Evidence suggests that short-term and long-term habituation have distinct neuroanatomical and neurofunctional correlates. Broadly, short-term habituation is considered a measure of short-term memory, whereas long-term habituation is considered a measure of long-term memory. 39,40 In light of the central role of the hippocampus in long-term habituation, 41 it is not surprising that in the present study, mice subjected to right temporal lobe mTBI failed to exhibit typical habituation responses (i.e., gradual reduction in response to repeated stimulation). In the context of HCM assessment, long-term habituation may be a valuable paradigm for evaluating hippocampal-dependent memory.
Qualitative aspects of wheel running
For the assessment of circadian-related behavior (e.g., anticipation and response to phase shifts), 3rd-day wheel activity was analyzed. Only data obtained on the 3rd day was used, as it is a time point when complete habituation is expected. 32 In line with the findings of Namdar and coworkers, 13 as the inactive light phase commenced, mTBI mice showed a sharp decline in wheel zone visits compared with their own activity levels during the dark phase. Although a somewhat similar pattern was observed in control mice, it was much less pronounced. Interestingly, the phase shift did not have a clear differential effect on the number of wheel revolutions of mTBI mice and controls. As this pattern was not accompanied by any motor dysfunction, as demonstrated in our treadmill test and past motor assessment studies, 13 we assume that this difference in wheel use pattern is not motor related. In addition to the increased frequency of visits to the wheel zone, there was an evident trend toward shorter visit durations in mTBI mice.
The number and duration of activity bouts are often used as indicators of fragmented activity rhythm. 42 In this context, our observations suggest a potential circadian disruption in mTBI mice. It is also possible that this pattern is related to attentional impairment. Phenotypes such as short activity bouts with a high frequency of bouts and brief inter-bout intervals observed in mTBI mice mirror symptoms of human attention-deficit/hyperactivity disorder (ADHD). 43 Earlier research has indicated that similar hyperactivity and altered response habituation, as seen in our study, are caused by disruptions in the dopaminergic system. 44 Specifically, the authors reported a sustained increase in wheel-running behavior, which continued to be evident even after 3 days of testing, along with a diminished response habituation to novelty. 44 This interpretation aligns with our findings on RL, pointing to abnormal attention-shifting abilities in mTBI mice. Therefore, it could be beneficial for future studies to focus on examining the function of the dopaminergic system in our mouse model.
DL and RL
Last, we assessed cognitive flexibility through DL and RL tasks in the Phenotyper home cage. Contrary to past evidence from a CognitionWall assessment, 23 this study found intact DL in mTBI mice. Our results, however, align with studies that used other operant tests outside the home cage environment, 45,46 indicating that discrimination performance is not entirely dependent on the testing environment. Instead, it is likely that the sensitivity of different tests varies across injury models, mouse strains, post-injury time points, and task types. Further research is required to examine the individual and combined effects of these factors, both inside and outside the home cage environment.
One week after the DL, we conducted an RL task, which requires a flexible adjustment of behavior when reward contingencies are reversed. 47 Although displaying similar learning abilities as controls, mTBI mice exhibited abnormally fast reversal learning, where they quickly shifted their attention to the previously non-reinforced alternative, treating it as if it were a novel stimulus. Interestingly, a similar finding, although in the opposite direction, was reported by Chen and coworkers, 23 who found blast mTBI mice have hypo-flexible learning.
Limitations and future directions
Behavioral outputs provide a valuable window into brain function, and our research is centered on analyzing these outputs. Our detailed longitudinal cognitive-behavioral evaluation of ICR mice subjected to closed-head mTBI has provided valuable insight into injury effects. However, it should be noted that different mouse strains can have different genetic characteristics, which can result in unique behavioral profiles. In particular, inbred mouse strains can vary in their habituation rates, with some habituating faster than others. 32,41 In the context of HCM, it is clear that spontaneous home cage behaviors are complex and strongly influenced by genetics, especially regarding certain behaviors occurring over specific time scales like light–dark phase transitions. 32 As a result, our findings from the ICR mouse strain may not apply to other strains. More home cage studies on mTBI outcomes are needed in other mouse strains. Second, the current research is based solely on male mice, leaving a gap in our understanding of mTBI outcomes in female mice. Given the unique hormonal profile 48 and potential sexual dimorphism in brain function, 49 female mice may respond differently to mTBI. To further our understanding of mTBI consequences, we intend to include female mice in our future studies.
Conclusion
In this study, we used an HCM system to investigate the behavioral consequences of closed-head mTBI. We found that mTBI mice exhibited abnormal habituation, hyperactivity, and circadian rhythm disruptions. Also, our mTBI model appears to selectively alter the ability to learn novel responses or extinguish old ones while leaving DL relatively intact. These results underscore the value of HCM systems as a complementary tool to traditional assays in studying mTBI, as they allow comprehensive and longitudinal assessment of behavioral changes in a more naturalistic environment. Moreover, our research highlights the importance of integrating both home cage and conventional assays for a comprehensive understanding of mTBI outcomes. By advancing our understanding of mTBI, HCM systems pave the way for improved treatments and interventions, ultimately improving the quality of life for mTBI patients.
Transparency, Rigor, and Reproducibility Summary
The sample size consisted of 79 ICR male mice; 6–8 weeks old. Mice were randomly assigned to either a weight-drop closed-head mTBI group (n = 39) or a control group (n = 40). To ensure the validity of our findings, we used separate cohorts for the different behavioral assays, considering the potential impact of wheel running on cognitive and motor abilities. Testing began a week following mTBI induction, except for the OF test, conducted 20 days post-injury. The housing and experimental room were kept under reversed light cycle conditions. Statistical analyses were conducted using RStudio and included linear mixed-effects models, one-sample t-tests, and dependent and independent two-sample t tests. All tests were two-tailed with a statistical significance level set at p < 0.05.
Footnotes
Authors' Contributions
B.R.-H.: writing – original draft (lead), formal analysis (lead); L.T.: conceptualization (supporting), writing – original draft (supporting), writing – review and editing (supporting); L.B.: conceptualization (lead), review and editing (equal), methodology (lead); C.G.P.: conceptualization (equal), review and editing (lead), methodology (equal).
Funding Information
This research was funded by the Ari and Regine Aprijaskis Fund, grant number 347300-00; the Dr. Miriam and Sheldon G. Adelson Center for the Biology of Addictive Diseases, grant number 601133461; and the Sylvan Adams Sports Institute, grant number 0601133671.
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.