
Abstract
Traumatic brain injury (TBI) persists as a substantial clinical dilemma, largely because of the absence of effective treatments. This challenge is exacerbated by the hindered clearance of intracranial metabolic byproducts and the continual accrual of deleterious proteins. The glymphatic system (GS) and meningeal lymphatic vessels (MLVs), key elements of the intracranial lymphatic network, play critical roles in the clearance of harmful substances. Cannabidiol (CBD) has shown promise in reducing metabolite overload and bolstering cognitive performance in various neurodegenerative diseases. The precise mechanisms attributing to its beneficial effects in TBI scenarios, however, are yet to be distinctly understood. Utilizing a fluid percussion injury paradigm, our research adopted a multifaceted approach, encompassing behavioral testing, immunofluorescence and immunohistochemical analyses, laser speckle imaging, western blot techniques, and bilateral cervical efferent lymphatic ligation. This methodology aimed to discern the influence of CBD on both neurological outcomes and intracranial lymphatic clearance in a murine TBI model. We observed that CBD administration notably ameliorated motor, memory, and cognitive functions, concurrently with a significant reduction in the concentration of phosphorylated tau protein and amyloid-β. In addition, CBD expedited the turnover and elimination of intracranial tracers, increased cerebral blood flow, and enhanced the efficacy of fluorescent tracer migration from MLVs to deep cervical lymph nodes (dCLNs). Remarkably, CBD treatment also induced a reversion in aquaporin-4 (AQP-4) polarization and curtailed neuroinflammatory indices. A pivotal discovery was that the surgical interruption of efferent lymphatic conduits in the neck nullified CBD's positive contributions to intracranial waste disposal and cognitive improvement, yet the anti-neuroinflammatory actions remained unaffected. These insights suggest that CBD may enhance intracranial metabolite clearance, potentially via the regulation of the intracranial lymphatic system, thereby offering neurofunctional prognostic improvement in TBI models. Our findings underscore the potential therapeutic applicability of CBD in TBI interventions, necessitating further comprehensive investigations and clinical validations to substantiate these initial conclusions.
Introduction
Traumatic brain injury (TBI)—a serious global health issue—is infamous for its prevalence and severe outcomes, including rising disability and mortality rates. 1 Studies have reported that moderate to severe TBI elevates the risk of neurodegenerative diseases, such as Alzheimer (AD) and Parkinson disease (PD). The brain experiences profound pathological alterations post-TBI, ranging from intracranial lymphatic system disruptions and blood–brain barrier compromise to neuronal damage and intense inflammation, all converging to promote abnormal protein accumulation. 1,2 This complex pathology is characterized by extracellular plaques triggered by β-amyloid (Aβ) and neurofibrillary tangles because of hyperphosphorylated tau protein, with ensuing inflammation exacerbating the situation. There are no effective treatments to improve TBI outcomes, highlighting the dire need for novel therapeutic targets and neuroprotective strategies.
Cannabidiol (CBD)—derived from the Cannabis sativa plant—has garnered accolades in the scientific community, primarily attributable to its putative neuroprotective virtues, ostensibly orchestrated through intricate liaisons with the endocannabinoid system. 3,4 The endocannabinoid system, a paramount architect in the labyrinth of neurodegeneration, exerts a profound influence over an array of neurological constellations, such as AD, PD, and multiple sclerosis (MS). 5 –7
In the contemporary scientific tableau, CBD emerges as a therapeutic harbinger in the battle against TBI, with an abundance of investigative discourses converging on its neuroprotective prowess, articulated principally through the attenuation of oxidative duress and inflammatory cascades. Pre-clinical archetypes venturing into the empirical domains offer an irrefutable testament to CBD's potential in bestowing neuroprotection, orchestrated through a symphony of regulatory mechanisms encompassing oxidative stress amelioration, autophagic modulation, apoptotic control, and the quelling of neuroinflammatory pathways. 1,8,9
Recent forays into AD rodent paradigms have yielded illuminating insights that further support the effect of CBD in curtailing the insidious advancement of amyloid plaque agglomerations, a hallmark of neurodegenerative pathology. 10 This burgeoning scholarly narrative not only augments our collective understanding but also heralds an era of therapeutic innovation, potentially culminating in the development of effective interventions for neurological conditions once deemed inexplicable.
The glymphatic system (GS) is a complex perivascular conduit that is essential for maintaining cerebral homeostasis by regulating the flux of cerebrospinal fluid (CSF) and interstitial fluid (ISF) within the neuronal milieu. 11,12 This architecturally complex network, demarcated by the perivascular astrocytic end-feet, is essential for maintaining the metabolic equilibrium of the central nervous system (CNS), as it supports cerebral fluid hemodynamics and solute clearance. 13 GS's operational efficacy, a cornerstone in sustaining aqueous stasis and proteostatic clearance within the CNS, is modulated by determinants, including intracranial pressure dynamics, arterial pulsatility, and the intricacies of circadian biology.
Integral to the optimal functioning of this system is Aquaporin-4 (AQP-4)—a protein localized primarily on the polarized foot processes of astrocytes and plays an essential role in the exchange process of CSF and ISF. 11,14 –16 GS dysfunction, observed in conditions like TBI, AD, MS, and PD, may stem from the dislocation of AQP-4, diminished cerebral blood flow (CBF), and increased neuroinflammatory buildup. 17 –19
Further, research on AQP-4 knockout mice has uncovered that meningeal lymphatic vessels (MLVs), which are essential for clearing intracranial metabolites and facilitating a fluid exchange between the GS and cervical lymph nodes (CLNs), 12,20 exhibit impaired functionality. This is evident from the diminished drainage of intracranial tracers to the CLNs. 21 Studies have suggested a close relationship between TBI and the intracranial lymphatic drainage system. 12,22,23
Based on preliminary evidence indicating the anti-inflammatory properties of CBD and its potential role in promoting cerebral lymphatic circulation, we hypothesize that CBD treatment may contribute to the amelioration of cognitive and motor deficits observed after TBI. This therapeutic effect is proposed to stem from the improved function of the entire intracranial lymphatic drainage system, which includes the GS, thereby facilitating the removal of neurotoxic substances and reducing neuroinflammation post-injury.
Methods
Animals
Pathogen-free male C57BL/6J mice aged 8–10 weeks and weighing 22–25 grams were obtained from Beijing Vital River Laboratory Animal Technology Co., Ltd., Beijing, China (license number SCXK [Jing] 2021-0006). The mice were housed at a stable temperature of 20 ± 2°C with 45% humidity, a 12h light/dark cycle, and free access to food and water. In brief, the mice were initially anesthetized with a 4% isoflurane inhalation and subsequently maintained in an anesthetic state with 1.5–2% isoflurane. After this, they received a subcutaneous injection of 5 mg/kg carprofen (Cayman Chemical, Ann Harbor, MI) for analgesia and were securely positioned in a stereotaxic frame. 24 Throughout the procedure, a mixture of 20% oxygen and 80% air was used, allowing for spontaneous breathing in the mice during the entire course of the study. All procedures were conducted strictly with the ARRIVE guidelines and were approved by the Animal Care and Use Committee of the General Hospital of Tianjin Medical University, China.
Fluid percussion injury (FPI) model
We adopted a FPI model based on established methodologies. 25 In this procedure, mice initially anesthetized were subjected to a craniotomy of 3 mm diameter, located 2 mm posterior to the bregma and 1.5 mm lateral to the sagittal suture. After this, a fluid percussion device (Custom Design & Fabrication, Richmond, VA), containing sterile saline, was utilized to administer a controlled fluid pressure pulse directly to the intact dura mater through the cranial opening. This pressure was meticulously maintained at 1.9 ± 0.2 atmospheres. Next, the scalp incision was closed with sutures, and the mice were placed on a heated pad at 37°C to maintain normal temperature until awakening. Except for the strike program, the Sham group underwent the same procedure.
Drug administration and experimental protocols
CBD (10 mg, PHL85705, Sigma Systems) was dissolved in HS 15 with 5% ethanol and administered intraperitoneally 1h post-TBI at a dosage of 5 mg/kg (in a volume of 6 mL/kg). 26,27 We will designate 1h, 24h, and 48h post-injury as the intervention time points, and the third day after TBI will be the time point for assessing the GS, MLVs, and neuronal apoptosis. All experimental protocols and configurations are detailed in Fig. 1. The mice were randomized across eight distinct experimental groups. Notably, the Sham+CBD group was instrumental in discerning any potential adverse effects because of CBD administration.

Schematic diagram of various experimental protocols and setups used in this study. CBD, cannabidiol; mNSS, modified neurological severity score; MWM, Morris water maze; NOR, Novel object recognition; MLVs, meningeal lymphatic vessels; TBI, traumatic brain injury, dCLNs, deep cervical lymph nodes; Aβ, β-amyloid.
Modified neurological severity score (mNSS)
The mNSS is a comprehensive battery that encompasses a range of tests to evaluate motor function, sensory acuity, reflexive activities, and balance. 28 These assessments were conducted at specific time points after the injury: 1, 3, 5, 7, and 14 days (denoted as D1, D3, D5, D7, and D14, respectively). The mNSS yields a cumulative score on a scale from 0 to 18, with higher scores indicating more severe neurological impairments. This scoring system provides a standardized measure to gauge the extent of functional deficits arising from TBI and monitor their progression or amelioration over time.
Rotarod test
Assessment of motor coordination and balance in mice was performed using a rotarod apparatus (Model YLS-4C, Beijing). This evaluation was carried out at designated post-TBI time points: D1, D3, D5, D7, and D14, respectively, by the protocol delineated in a previous study. 29 Before the trial, all mice were trained on the rotarod (5–10 RPM) for 300 sec. On each testing day, the rod was conducted at a uniform accelerating speed from 5–40 RPM for 300 sec, repeated three times. The latency to fall of each mouse was recorded and averaged.
Novel object recognition (NOR) test
The NOR test protocol was adapted from established methodologies.
30
Briefly, the tests were performed in a 40
Morris water maze (MWM) test
Assessment of spatial learning and memory in mice post-TBI was conducted using the MWM test, as outlined in previous research. 29 Briefly, the experiment consisted of a training phase of five consecutive days and a testing phase of one day. For training on the 15–19th days, the mice were trained to seek a hidden platform within the 90 sec and stay on the platform for 15 sec. If the platform was not found within 90 sec, the mouse was gently guided to reach the platform and remained there for 15 sec, and the recording latency was 90 sec.
During the training phase, the mice were trained from the first to the fourth quadrant four times a day. The test was conducted on the 20th day, with the platform removed, and the mouse was placed on the opposite side of the platform quadrant. We used video tracking software (EthoVision XT 13, Noldus Information Technology, Wageningen, Netherlands) to record and analyze the latency to the platform, platform crossing times, and swimming traces.
Intracisternal injections
Intracisternal injections followed established protocols. 22 A syringe pump was used to deliver 10 μL of rhodamine B isothiocyanate-dextran (RITC-Dextran, 70 kD, R9379, Sigma) diluted to 2.5% in artificial CSF at 1 μL/min. Brain tissue and dCLNs were retrieved 30 min post-injection. In parallel, 2 μL of Beads 505/515 (Invitrogen, cat# F8813) were infused at 0.5 μL/min. Dorsal meninges and dCLNs were collected after 30 min. These specimens were fixed overnight at 4°C in 4% paraformaldehyde (PFA). The brain tissues were then coronally sectioned at 100 μm using a cryostat (CM1950, Leica Biosystems), stained with 4',6-diamidino-2-phenylindole (DAPI) and examined under an inverted fluorescence microscope (IX73, Olympus). For meningeal samples, after skin and muscle removal, the skull was excised and fixed in 4% PFA at 4°C overnight. The meninges were carefully extracted and placed in phosphate-buffered saline (PBS) in preparation for immunofluorescence staining.
Intraparenchymal injections
Following established protocols, intraparenchymal injections with a 0.5 μL, 2.5% 70 kD RITC-Dextran tracer were infused into the brain at coordinates 2.0 mm posterior, 1.5 mm lateral, and 2 mm ventral to the bregma over 5 min. After injection, the needle was allowed to stay at the injection site for 5 min and then withdrawn slowly to prevent the solution from leaking. One hour post-injection, brain tissues were retrieved and processed similarly to intracisternal injections.
Immunofluorescence staining
Brain tissues were initially excised and preserved in 4% PFA at 4°C overnight, followed by sequential dehydration through 15% and 30% sucrose solutions. The tissues were then embedded in an optimal cutting temperature compound (Sakura Finetek USA, Torrance, CA) and sectioned into 8-μm coronal slices using a cryostat. Brain membranes and sections were washed in PBS and then blocked for 1.5h at room temperature in a solution comprising 3% bovine serum albumin, 0.2% Triton X-100, and 0.05% Tween 20 in PBS. They were then incubated with primary antibodies overnight at 4°C and secondary antibodies at room temperature for an additional 2h. The primary antibodies included rabbit anti-LYVE-1 (1:200, Abcam, ab14917), rabbit anti-AQP-4 (1:500, Cell Signaling Technology, 59678), goat anti-CD31 (1:200, R&D Systems, AF3628), rabbit anti-glial fibrillary acidic protein (GFAP, 1:500, Abcam, ab7260), and goat anti-IBA-1 (1:500, Abcam, ab5076). After staining, the sections were examined under an inverted fluorescence microscope.
Hematoxylin and eosin (H&E) and Nissl staining
The brain tissues of mice were initially preserved in a 4% PFA solution, then sequentially dehydrated in varying ethanol concentrations, and finally embedded in paraffin. The tissues were then sectioned into 8-mm-thick slices and prepared for histological examination using H&E (G1120, Solarbio) and Nissl (G1432, Solarbio) staining techniques, following previously established methodologies. 29 Microscopic analysis of the stained sections was conducted using a standard light microscope (IX73, Olympus Corporation).
Immunohistochemical staining
Using previously established protocols, we used immunohistochemistry (IHC) to identify Aβ. The sections were incubated in 0.3% hydrogen peroxide for 30 min, followed by antigen unmasking using a heated citrate buffer. Subsequently, the sections were treated with 1% bovine serum for 30 min to diminish non-specific interactions. The sections were then incubated overnight at 4°C with a primary antibody specifically against Aβ (rabbit anti-Aβ, 1:400, GB111197; Servicebio, Wuhan, China). A secondary biotinylated goat anti-rabbit antibody (GK500705, GeneTech) was applied for 1h at room temperature. The sections were observed under a conventional light microscope. For quantitative analysis, the mean optical density (MOD) of Aβ 31 was evaluated using Image-Pro Plus 6.0 software (Media Cybernetics, Inc., Rockville, MD).
Laser speckle imaging
The CBF was quantified using the PeriCam PSI System (Perimed AB, Sweden) based on laser speckle imaging methodologies. Anesthetized mice underwent a midline skin incision to expose the cranium, facilitating the speckle contrast-based CBF assessment through the intact skull. After the measurement, aseptic techniques were used for wound closure. The perfusion data were analyzed using PIMsoft 1.2 software. Data captured from defined regions of interest (ROIs)—comprising the entire cerebrum, ipsilateral, and contralateral regions—were synchronized at specific intervals: baseline (pre-TBI) and on days 3 and 7 post-TBI.
Western blotting
Western blotting was used to quantify phosphorylated tau (p-tau) protein levels in the affected brain hemispheres, following methodologies delineated in prior research. After euthanasia and perfusion with cold PBS, tissue samples were collected on day 3 post-TBI. Lysates were prepared using a radio-immunoprecipitation assay buffer that included protease and phosphatase inhibitors (P1260, Solarbio, Beijing, China), and protein concentrations were ascertained using the BCA (bicinchoninic acid) protein assay kit (ThermoFisher Scientific, Waltham, MA). Protein samples (10 μL) were resolved via 10% SDS-PAGE under uniform voltage before being transferred to polyvinylidene difluoride (PVDF) membranes using a wet electrotransfer apparatus (Bio-Rad).
The membranes were then incubated overnight at 4°C with specific primary antibodies: rabbit anti-phospho-tau (Thr205) (1:2000, Abclonal, AP0168), rabbit anti-phospho-tau (Ser404) (1:2000, Abclonal, AP0170), and rabbit anti-phospho-tau (Ser396) (1:1000, Cell Signaling Technology, 9632s). After primary antibody exposure, the membranes were thoroughly washed and incubated with horseradish peroxidase-conjugated secondary antibodies. Density was quantified using FIJI (ImageJ) software, with relative protein expression levels normalized against β-actin levels.
Bilateral cervical efferent lymphatic ligation
Before placement under a surgical microscope, the mice were first anesthetized, and their necks were meticulously disinfected during the surgical procedure. A 2-cm skin incision was executed along the cervical midline, followed by blunt dissection of the cervical fascia. The trachea and the sternocleidomastoid muscle were revealed by delicately elevating glandular tissue to facilitate the identification and isolation of lymphatic vessels or nodes within the deep cervical region. Utmost care was taken to ligate all profound efferent lymphatic vessels in the cervical area using sterile 11-0 nylon sutures. Subsequently, the glandular tissue was repositioned, and a symmetrical procedure was performed to ligate the lymphatic vessels on the contralateral side. This meticulous methodology ensures the precision and replicability vital for scientific inquiry in an academic context.
Cytokine array detection
Proteins were extracted from brain tissues on day 3 post-TBI using established protocols for Western blotting and then analyzed using a cytokine array (ARY006; R&D Systems, Minneapolis, MN). Each membrane of the protein detection chip was incubated overnight at 4°C with 2000 μg of protein (in a 5:1 mixture). The membranes were then washed with 1 × washing buffer and incubated for 30 min on a shaker with streptomycin-horseradish peroxidase conjugate. After additional washing, the membranes were exposed to ECL luminescent solution for 1 min and imaged using the ChemiDoc imaging system (Bio-Rad, Hercules, CA) for detailed analysis.
Statistical analysis
This study's sample size adheres to quantities documented in prior studies. 14 –16 Analytical procedures were conducted by researchers blinded to the specific experimental allocations. Results are articulated as mean ± standard deviation (SD). All datasets were subjected to the Shapiro-Wilk normality test, yielding p values greater than 0.1000. Statistical evaluations were executed through one-way analysis of variance (ANOVA) accompanied by the Tukey post hoc test for comparisons among multiple groups. Statistical significance was acknowledged when p values were less than 0.05. All computations and graphical depictions were facilitated using GraphPad Prism software (version 9.0).
Results
CBD Mitigates impairments in motor and cognitive functions in mice post-TBI
We investigated the effect of CBD on motor and cognitive deficits post-TBI in mice using a TBI model. In the mNSS, the TBI mice receiving vehicle treatment (TBI+Vehicle) exhibited notably deteriorated scores compared with the CBD-treated cohort (TBI+CBD) at all examined intervals (Fig. 2a). Rotarod assessments revealed a pronounced decline in motor coordination and balance in the TBI+Vehicle mice Fig. 2b). Moreover, in the MWM trials, the escape latency of the TBI+Vehicle group significantly exceeded that of the control group, a disparity mitigated by CBD administration from day 19 post-TBI. Notably, the swim speeds across all groups remained consistent (Fig. 2c-d).

Effect of cannabidiol (CBD) treatment on the neurological function of mice with traumatic brain injury (TBI). (
On day 20 post-TBI, platform removal for probe trials revealed a significant reduction in platform crossings in the TBI+Vehicle group compared with the TBI+CBD group. In addition, the TBI+Vehicle mice dedicated less time to the target quadrant than the control group, an adverse effect attenuated by CBD (Fig. 2e–g). During NOR assessments, the discrimination index was markedly lower in the TBI+Vehicle group compared with the Sham group; however, this discrepancy significantly narrowed in the TBI+CBD mice. Notably, no differences in motor skills and cognition emerged between the Sham and Sham+CBD groups, underlining CBD's specificity for TBI-induced deficits (Fig. 2h-i). These findings highlight CBD's potential to alleviate TBI-induced motor and cognitive impairments, warranting further exploration into its mechanisms and therapeutic prospects.
CBD reduces metabolite deposition and inflammatory responses in mice with acute TBI
Studies have indicated an exacerbated accumulation of protein metabolites in post-TBI, including p-tau and Aβ. To corroborate these findings, we examined p-tau expression through Western blotting and Aβ deposition using H&E staining in the cerebral cortex of acute TBI mice on day 3 post-TBI. Our data revealed a substantial surge in cortical p-tau (Thr205), p-tau (Ser396), p-tau (Ser404), and Aβ levels in the untreated group compared with the Sham and Sham+CBD groups. This escalation was significantly mitigated after CBD intervention. Notably, there were no significant variances in p-tau/Aβ concentrations between the Sham and Sham+CBD cohorts (Fig. 3a,b, d,e).

Effect of cannabidiol (CBD) application on metabolite deposition, cellular degeneration, and neuroinflammation in mice with traumatic brain injury. (
In our histological and fluorescent analyses, the selected sites for immunohistochemistry or fluorescence were strategically focused around the injured cortical hemisphere of the mouse brain, ensuring the relevance and precision of our observations (Fig. 3c). We evaluated CBD's neuroprotective efficacy post-TBI using Nissl staining to quantify neuronal death within the cerebral cortex. The CBD substantially enhanced neuronal preservation on day 3 post-TBI versus the untreated group. H&E staining was used to investigate cerebral cortical morphology. The CBD administration significantly reduced cortical tissue degeneration, cerebral damage, and traumatic intraparenchymal hemorrhaging induced on day 3 post-TBI. Neither the Sham nor Sham+CBD groups exhibited any discernible cortical histological impairments (Fig. 3d,f).
Concurrent studies have suggested that microglial lysosomal vesicles contribute to post-TBI neural inflammation. Our analyses confirmed a pronounced expansion in the IBA-1 positive domain within the mouse cortex in the acute TBI phase, indicative of augmented microglial activation, which was substantially curtailed in the CBD-treated subjects. In addition, assessments of activated GFAP-positive astrocytes in the cortex mirrored the microglial trends observed between the untreated and CBD-treated groups. Conversely, no marked disparity in glial activation was discerned between the Sham and Sham+CBD groups Fig. 3g–i). These findings highlight CBD's capacity to reduce metabolite deposition and temper inflammatory responses in an acute TBI context, signaling its potential therapeutic utility.
CBD improves the solute drainage function of the GS and AQP-4 polarization and enhanced CBF in acute TBI mice
We used an FPI model and administered RITC-Dextran into the cisterna magna of mice on day 3 post-TBI to ascertain the effect of CBD on GS's CSF penetration into the brain parenchyma. After a 30-min circulation period, we observed a significantly enhanced tracer influx into the brain parenchyma in the CBD-treated group compared with the untreated cohorts. Notably, there was no substantial discrepancy in the fluorescence area ratio of brain sections between the Sham and Sham+CBD groups. Further, we evaluated the ISF clearance rate from the brain.
Analyses conducted 1h post-cortical injection revealed a markedly lower proportion of tracer fluorescence within the brain parenchyma in the CBD-treated group than the TBI+Vehicle group. Again, no significant difference in fluorescence area ratio was detected between the Sham and Sham+CBD groups (Fig. 4). These results indicate that CBD significantly enhances lymphoid clearance after TBI, while its effect on glymphatic clearance in the normal, uninjured brain is insignificant.
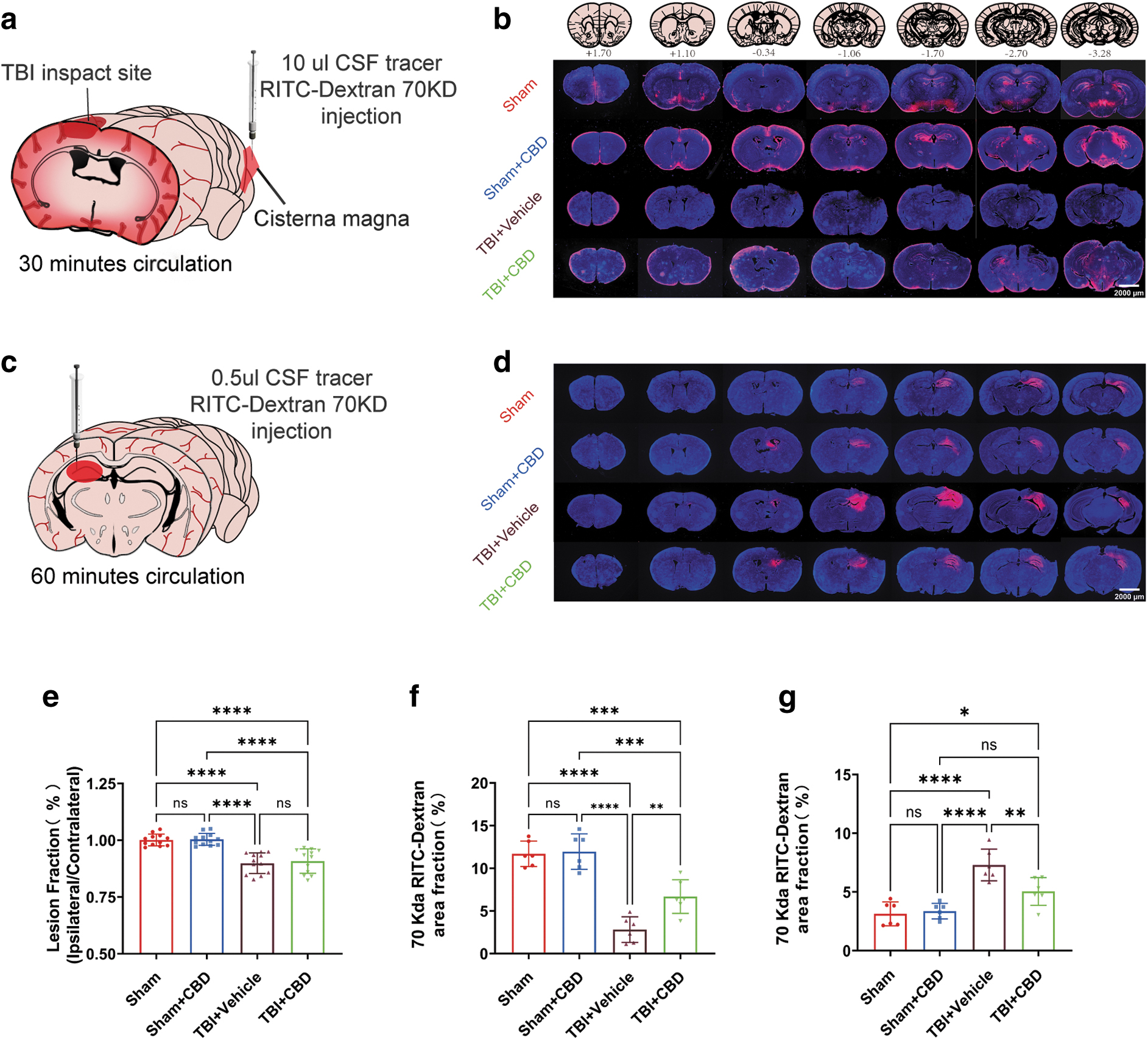
Effect of cannabidiol (CBD) treatment on glymphatic system (GS) function in traumatic brain injury (TBI) mice. (
The functionality of GS is contingent on the paravascular polarization of AQP-4 water channels and the dynamics of CBF, both vital for CSF recirculation and ISF solute clearance. We initially examined the perivascular polarization of AQP-4 using immunofluorescence, costained for CD31 and AQP-4. Consistent with prior findings, there was a pronounced diminution in AQP-4 polarization on day 3 post-TBI, which was effectively ameliorated with CBD treatment. Conversely, AQP-4 polarization in the Sham group remained unaltered by the CBD administration (Fig. 5a,b).

Effect of cannabidiol (CBD) application on brain Aquaporin-4 (AQP-4) polarization and blood flow in mice with traumatic brain injury (TBI). (
We subsequently monitored ipsilateral, contralateral, and global brain CBF on days 3 and 7 post-TBI. CBF demonstrated a temporal increment post-TBI. There was a conspicuous reduction in CBF in the TBI+Vehicle group, however, particularly noticeable globally and in the injured hemisphere on day 3, compared with the TBI+CBD group. No statistical differences in CBF were discerned between the Sham and Sham+CBD cohorts (Fig. 5c–e). These findings suggest CBD's potential in bolstering glymphatic solute clearance, AQP-4 polarization, and CBF post-acute TBI, offering its therapeutic promise.
CBD improves the drainage of MLVs in acute TBI mice
Recent studies have identified MLVs in the dura mater that communicate with dCLNs, playing a pivotal role in the clearance of large molecules from the brain parenchyma. To assess the MLVs' drainage efficiency post-TBI, we injected a tracer into the cisterna magna and allowed a 30-min circulation period. Subsequently, dorsal MLVs and dCLNs sections were procured for analysis. The TBI+Vehicle group exhibited a significantly diminished presence of Beads within the MLVs compared with the Sham group. Conversely, the TBI+CBD group had a markedly elevated proportion of Beads in the MLVs compared with the TBI+Vehicle group. There was also a pronounced decrement in Beads or fluorescent tracer enrichment within the dCLNs post-TBI.
The TBI+CBD group displayed a significantly higher drainage ratio of Beads or fluorescent tracers to the dCLNs than the TBI+Vehicle group, however. Notably, no alteration in the proportion of the fluorescence area was observed in dorsal MLVs and dCLNs between the Sham and Sham+CBD groups (Fig. 6). These findings highlight the potential of CBD in facilitating the drainage function of MLVs post-acute TBI, underscoring its therapeutic viability.

Effect of cannabidiol (CBD) treatment on meningeal lymphatic vessels (MLVs) function in mice with traumatic brain injury (TBI). A representative image of the dorsal meningeal stained for LYVE-1 dye (red color) shows the uptake of Beads in cerebrospinal fluid (CSF) by dorsal MLVs. (
CBD improves cognitive function in TBI mice by regulating intracranial lymphatic drainage
The present study sought to delineate the principal mechanism underlying the neuroprotective effect of CBD in TBI mice by obstructing intracranial lymphatic drainage through bilateral cervical afferent lymphatic vessel ligation. The results of the study demonstrated that administration of CBD led to a reduction in inflammatory mediators in both the ligation+TBI+CBD and non-ligation+TBI+CBD groups, compared with their respective vehicle-treated groups. Intriguingly, a decrease in inflammation was also observed in the ligation+TBI+Vehicle, suggesting that the ligation procedure itself may confer an anti-inflammatory effect in the context of TBI (Fig. 7a–f). These outcomes indicate that CBD's anti-inflammatory properties are exerted independently of the lymphatic drainage state.

Ligation of lymphatic vessels blocks the recovery effect of cannabidiol (CBD) on the intracerebral lymphatic system. A cartoon diagram illustrating the process of lymphatic vessel ligation (
Our investigation began with evaluating the proportion of fluorescent tracers administered into the brain parenchyma to gauge intracranial lymphatic drainage efficiency. The results demonstrated a discernible increase in the fluorescence ratio in the ligation+TBI+CBD, ligation+TBI+Vehicle, and non-ligation+TBI+Vehicle groups compared with the non-ligation+TBI+CBD groups (Fig. 7g,h). Subsequent immunohistochemical analysis was conducted to quantify Aβ deposition in the cortex. In parallel with the fluorescent tracer data, Aβ accumulation in the cortical region of the other groups was significantly more pronounced than that in the non-ligation+TBI+CBD group (Fig. 7i-j).
We used the MWM and NOR assays to corroborate the hypothesis that CBD ameliorates cognitive deficits by modulating the intracranial lymphatic drainage system. The results demonstrated that only the non-ligation+TBI+CBD group exhibited substantial mitigation of deficits in both short-term and spatial memory functionalities (Fig. 7k–q). These findings demonstrate CBD's potential as a modulator of intracranial lymphatic drainage, highlighting its therapeutic promise in ameliorating cognitive impairments post-TBI.
Discussion
CBD—a distinguished non-psychoactive extract of Cannabis sativa—is renowned for its therapeutic efficacy in mitigating diverse CNS maladies, encompassing ischemia, AD, PD, epilepsy, and schizophrenia. 5,32 –36 Our findings align with prevailing research, demonstrating pronounced improvements in motor skills, cognition, and memory in murine TBI models after CBD treatment. Although current scientific inquiries predominantly concentrate on CBD's neuroprotection via immunomodulatory pathways amid neuroinflammation, investigations into its effects on intracranial lymphatic regulation are scant.
Our initial observations (unpublished data) revealed an uptick in macrophage responsiveness and vascular permeability in the vicinity of MLVs in CBD-administered Parkinson models, accentuating CBD's critical role in orchestrating intracranial lymphatic responses during neurological perturbations. Consequently, our study investigated the effects of CBD on the structural integrity and functional efficacy of lymphatic conduits and meningeal lymphatics post-acute TBI in mice, thereby enriching the discourse in CBD-mediated neurotherapeutics.
Based on our findings, we hypothesize that CBD enhances intracranial lymphatic drainage through several mechanisms: (1) re-establishing the polarized state of AQP-4 adjacent to cerebral blood vessels, (2) restoring CBF, and (3) attenuating neuroinflammation. This study innovatively investigated CBD's neuroprotective effects post-TBI in mice, focusing on brain lymphatic drainage modulation. Our findings reveal that CBD enhances cerebral cortex CSF-ISF exchange, reverses AQP-4 depolarization, restores CBF, and boosts meningeal lymphatic drainage, thereby improving cognitive functions beyond merely regulating neuroinflammation.
The GS—mirroring the CNS's lymphatic structure—manages fluid movement and waste removal via astrocytic AQP-4 channels. 37 The GS function, essential for neurological health, depends on glial cell and CBF interaction, with dysfunction contributing significantly to conditions such as TBI and AD through poor clearance of deleterious proteins. 37 –42 Our findings indicate that CBD treatment enhances CSF penetration and interstitial waste clearance in the brain's lymphatic system in acute TBI models. We posit that CBD's therapeutic application could alleviate neurological impairments post-TBI by bolstering glymphatic functionality and reducing toxic metabolite accumulation, thereby presenting new avenues for neurotrauma treatment and prognosis.
To delve profoundly into the effects of CBD on GS, we further examined the CBD-induced molecular phenotypic alterations within the lymphatic network. AQP-4 is predominantly expressed at the termini of astrocytic foot processes surrounding capillaries, subpial spaces, and ventricular walls. 43,44 It is integral to the pathophysiology of diverse neurological disorders, including cerebral edema, 37,44,45 ischemia, 46 AD, 45,47 and PD. 48 Recent studies have demonstrated AQP-4's crucial role in brain homeostasis, facilitating Aβ clearance, synaptic function, and solute transport through mechanisms such as lymphatic clearance, transcytosis, and neuroglial degradation. 47
Studies have indicated that wild-type mice have higher CSF influx than AQP-4 knockout variants, with the latter exhibiting significantly reduced tracer transport. 49 In addition, AQP-4-deficient mice demonstrate anomalies in brain structure and water molecule transport, 50 signifying AQP-4's essential role in CSF and lymphatic solute transport. Reduced perivascular AQP-4 localization in the cortical gray matter of Alzheimer subjects suggests an association with increased Aβ and neurofibrillary pathology and cognitive decline pre-dementia. 18
In our study, CBD treatment facilitated increased tracer influx in neural parenchyma in acute TBI mice, notably reversing the AQP-4 polarization observed on day 3 post-TBI and thus promoting vascular tracer exchange. Prior studies have linked reactive astrocyte proliferation in acute TBI mice to alterations in AQP-4 expression and polarization. 21 AQP-4's expression and subcellular placement are influenced by various signals, including CKII-catalyzed Ser276 phosphorylation, calmodulin (CaM), and protein kinase A (PKA), which mediate changes via intracellular calcium oscillations. 51
Crucial for intracellular and extracellular water equilibrium, AQP-4's absence in the blood–brain barrier markedly diminishes cerebral cellular swelling. AQP-4 in astrocytes regulates water permeability, internal and external water balance, and intracellular calcium dynamics. 52 The CBD may promote intracranial lymphatic drainage by inducing the polarization of AQP-4 on the end feet of astrocytes, a crucial step in the regulation of water homeostasis and the facilitation of waste clearance in the brain. The potential modulation of AQP-4 by CBD could be attributed to its ability to reduce the proliferation of reactive astrocytes or to its role in stabilizing the localization of AQP-4 within the perivascular astrocytic end-feet, potentially through the selective regulation of AQP-4 subtypes. Elucidating this mechanism requires further detailed investigation.
Meningeal lymphatic vessels—functional lymphatic systems within the meninges—ordinarily facilitate the intricate circulation and exchange of soluble substances between CSF and ISF. 53,54 Dysfunction in these vessels leads to an accumulation of metabolic waste, exacerbating neuroinflammation and promoting the release of pro-inflammatory cytokines in the brain. 54,55 The CBD exhibits extensive anti-inflammatory properties; in vitro studies have demonstrated the anti-inflammatory properties of CBD comparable with dexamethasone. 56
Our findings indicate that CBD significantly diminishes intracranial inflammatory cytokines and improves cognitive outcomes post-TBI, corroborating earlier studies. Given the importance of dorsally located meningeal lymphatic vessels in CSF clearance, we focused on dorsal MLVs. These vessels are essential conduits for large molecules to reach directly from the CSF to the CLNs. Previous work has demonstrated that disrupting these pathways through surgical or photodynamic means exacerbates neural damage and increases death in virally infected mice. 57,58 Our bilateral cervical afferent lymphatic vessel ligation led to a surge in intracranial inflammatory cytokines in healthy mice, demonstrating that impaired intracranial drainage pathways amplify neural harm and neuroinflammation.
Recent findings highlight impaired MLVs drainage in models of closed TBI, subarachnoid hemorrhage, and cranial hypertension, restricting substance transport to dCLNs. 58 Our findings reveal that CBD boosts fluorescent tracer drainage in post-ligation MLVs. Notably, cognitive function remained unimproved despite CBD suppressing inflammatory cytokine expression in TBI mice with ligated cervical afferent lymphatic vessels. This dissociation suggests that the anti-inflammatory effects of CBD are independent of intracranial lymphatic drainage. Our findings indicate that CBD's modulation of intracranial lymphatic drainage, as opposed to simple regulation of inflammatory cytokines, is responsible for restoring cognitive function in TBI mice. We propose that CBD may disrupt inflammatory cytokine interactions, reducing toxic metabolite release.
It is well established that physical compression of brain tissue adversely affects intracranial lymphatic circulation. Studies have indicated that an escalation in intracranial pressure results in cerebral vascular obstruction, attenuating vascular pulsations. 39 Further, the augmented TBI-induced intracranial pressure also disrupts the lymphatic drainage functions of the cervical lymph nodes and the dorsal meningeal lymphatics, thereby enhancing lymphatic flow toward the sacral lymph nodes. 22,59 The CBD can promote angiogenesis, thereby restoring CBF diminished by TBI. Adequate CBF is imperative for maintaining normal arterial pulsations, providing indirect corroboration for our hypothesis. Our findings confirm that the TBI+CBD group exhibited a significant increase in cortical blood perfusion on day 3 post-TBI.
The brain's lymphatic drainage system—comprising perivascular pathways based on the basement membrane, comprehensive brain-like lymphatic routes, and CSF drainage routes—functions by expelling large molecules and inflammatory mediators from CSF into CLNs, thereby modulating inflammatory responses. 55,60 Studies have indicated that early brain injury and delayed cerebral ischemia after aneurysmal subarachnoid hemorrhage manifest through the dysfunction of blood circulation, CSF circulation, and lymphatic-like systems, subsequently precipitating vasospasm, hematoma neurotoxicity, and a cerebral inflammatory storm. 61
In the context of AD, MLVs dysfunction, leading to Aβ accumulation in brain tissues and vessels, precedes cerebral amyloid angiopathy and related neuroinflammatory conditions. 19,62 This information highlights the critical link between cerebral drainage and neurodegenerative diseases. Our data affirm this link, demonstrating that CBD treatment post-acute TBI in mice improved cerebral drainage, reduced tau/p-tau and Aβ levels, and enhanced short-term and spatial memory.
We must acknowledge, however, the limitations present in our experiment. Specifically, the rationale behind the dosing and timing of CBD administration remains unclear, necessitating further exploration to understand the therapeutic efficacy of CBD. In addition, the impact of brain tissue loss on parenchymal clearance calculations, particularly in the context of potential leakage into the extra-axial space, is not thoroughly addressed. The researchers recognize that the intracranial lymphatic system's complexity and the interaction of multiple mechanisms demand a more exhaustive investigation. Future research aims to fill these gaps, deepening our understanding of CBD's role in improving this system after TBI.
Conclusion
The CBD enhances the function of intracranial lymphatic drainage, diminishes the accumulation of pathogenic p-tau and Aβ proteins, and ameliorates neurological outcomes in mice afflicted with TBI. These findings suggest that CBD holds promise as a therapeutic strategy for a global population of patients with TBI.
Transparency, Rigor, and Reproducibility Summary
Experimental data, representing groups of 5-10 mice, is conveyed as a mean ± standard deviation (SD). The Shapiro-Wilk test was employed to assess the normality of the data distribution. For comparisons involving multiple groups, a one-way analysis of variance (ANOVA) was conducted, supplemented by the Tukey post hoc examination. A p value below 0.05 was established as statistically significant. A cohort of 334 male mice participated in this investigation. Varied researchers bore the mantle of administering treatments or orchestrating behavioral assessments. Regrettably, nine mice succumbed post-FPI, preceding any intervention; four were precluded owing to technical discrepancies during cervical lymphatic duct ligation, and five lacked the vigor for behavioral evaluations. Comprehensive data sets were procured from 316 mice, aged between 8 to 10 weeks. Post undergoing experimental FPI, cervical lymphatic duct ligation, or sham procedures, these specimens were allocated to experimental cohorts through a digital numeric generator.
Footnotes
Acknowledgments
Authors' Contributions
RJ, CG, and SD conceived and designed the study and analyzed the data. MN, HZ, and JF developed the methodology. SD, ZS, ML, CL, YC, WJ, and JY conducted the experiments. SD and HW analyzed and interpreted the data. SD wrote the manuscript. YQ and CG provided technical support. RJ and CG reviewed and revised the manuscript and supervised the study. All authors have read and approved the manuscript.
Availability of Data and Materials
The datasets used and analyzed during the current study are available from the corresponding author on reasonable request.
Funding Informaton
This work was supported by the National Natural Science Foundation of China (Nos. 82271394).
Author Disclosure Statement
All experiments were approved by the Animal Care and Use Committee of Tianjin Medical University General Hospital, China, and were conducted following the ARRIVE guidelines.
No competing financial interests exist.