
Abstract
In the past decade, signature clinical neuropathology of blast-induced traumatic brain injury has been under intense debate, but interface astroglial scarring (IAS) seems to be convincing. In this study, we examined whether IAS could be replicated in the rat brain exposed to a laser-induced shock wave(s) (LISW[s]), a tool that can produce a pure shock wave (primary mechanism) without dynamic pressure (tertiary mechanism). Under certain conditions, we observed astroglial scarring in the subpial glial plate (SGP), gray-white matter junctions (GM-WM), ventricular wall (VW), and regions surrounding cortical blood vessels, accurately reproducing clinical IAS. We also observed shock wave impulse-dependent meningeal damage (dural microhemorrhage) in vivo by transcranial near-infrared (NIR) reflectance imaging. Importantly, there were significant correlations between the degree of dural microhemorrhage and the extent of astroglial scarring more than 7 days post-exposure, suggesting an association of meningeal damage with astroglial scarring. The results demonstrated that the primary mechanism alone caused the IAS and meningeal damage, both of which are attributable to acoustic impedance mismatching at multi-layered tissue boundaries. The time course of glial fibrillary acidic protein (GFAP) immunoreactivity depended not only on the LISW conditions but also on the regions. In the SGP, significant increases in GFAP immunoreactivity were observed at 3 days post-exposure, whereas in the GM-WM and VW, GFAP immunoreactivity was not significantly increased before 28 days post-exposure, suggesting different pathological mechanisms. With the high-impulse single exposure or the multiple exposure (low impulse), fibrotic reaction or fibrotic scar formation was observed, in addition to astroglial scarring, in the cortical surface region. Although there are some limitations, this seems to be the first report on the shock-wave-induced IAS rodent model. The model may be useful to explore potential therapeutic approaches for IAS.
Introduction
Despite extensive studies on blast-induced traumatic brain injury (bTBI), its signature neuropathology is still under intense debate. In 2016, Shively and colleagues reported astroglial scarring in the post-mortem brains of U.S. military personnel who had been exposed to blasts. 1 Distinct lesions were observed in the subpial glial plates (SGPs), circumference of cortical blood vessels, gray-white matter junctions (GM-WM), and structures lining the ventricles. These regions correspond to anatomical tissue boundaries, and the pathology was thus termed interface astroglial scarring (IAS). As another neuropathology for bTBI, Goldstein and associates reported chronic traumatic encephalopathy (CTE), which is characterized by accumulated phosphorylated tau protein, 2 but Priemer and co-workers reported that CTE did not frequently appear in blast-exposed military personnel who sustained chronic neuropsychiatric sequelae. 3 Recently, Benjamini and colleagues proposed multi-dimensional magnetic resonance imaging for non-invasive diagnosis of IAS. 4
An incoming shock wave or pressure wave is reflected at tissue boundaries with different acoustic impedances (the product of sound velocity and density of the tissue), by which a positive pressure can be converted to a negative pressure for the free-end reflection (reflection at the interface with a smaller acoustic impedance). In general, soft tissue is more susceptible to a tensile force than to a compressive force, and the tissue can therefore be more easily altered with a negative pressure. 5 In addition, a negative pressure can cause cavitation, which is another speculated injury mechanism for bTBI. 6 –8 Such pressure-reflection-related phenomena could be associated with IAS. Miller and colleagues showed by computer simulation that during blast exposure, physical distortion/strain generated in conjunction with cavitation was maximal at anatomical interfaces in the brain. 9
On the basis of issues described above, we assumed that IAS was a signature neuropathology for bTBI, and reliable animal models are therefore needed for shock-wave-induced IAS in the brain. Schwerin and associates examined IAS in the ferret brain exposed to a blast(s). 10 Because ferrets have a gyrencephalic brain, the model would be useful to investigate astroglial scarring in gyri and sulci. Zhou and co-workers reported astroglial scarring in the mouse brain exposed to a moderate intensity shock wave, but it was only around the ventricles and blood vessels. 11 There seems to be no report on the clinically relevant rodent model for shock-wave-induced IAS.
Although shock tubes have been widely used in animal studies of bTBI, we have been using laser-induced shock waves (LISWs) to investigate the mechanisms of shock-wave-induced brain injury. 12 –15 A blast wave consists of a shock wave (static pressure) and a blast wind (dynamic pressure). 16,17 Primary blast injury is defined to be caused directly from the shock-wave overpressure, 18 –22 whereas a wind or dynamic pressure causes displacement of the body with subsequent impact or crush, which is the tertiary blast injury. 18 –23 Because mild to moderate bTBI is believed to be associated with the primary mechanism, many researchers have been paying attention to excluding the effects of the tertiary mechanism in their animal studies for bTBI. Because LISW is not accompanied by dynamic pressure and its energy is spatially well confined, the effects of pure shock-wave exposure (primary mechanism) solely on the brain can be examined with an LISW. 12 –15,23 –25
In this study, we examined whether IAS could be replicated in the rat brain exposed to an LISW(s) and revealed its spatio-temporal characteristics based on immunohistochemistry. In our previous experiments using an LISW, we observed that shock-wave exposure often caused microhemorrhage specifically in the dura of the meninges. 12 This is also attributable to the effect of pressure wave reflections at tissue boundaries in the meninges that have a multi-layer structure with considerably different acoustic impedances: the dura attached to the skull, arachnoid mater, cerebrospinal fluid (CSF) in the subarachnoid space, and pia mater. It should also be noted that dural vessels have no blood–brain barrier (BBB) and are fenestrated, 26 and they would therefore be inherently permeable, thus easily causing extravasation by shock-wave exposure. In addition, various immunocompetent cells and fibroblasts are present in the meninges, 26 and they can infiltrate into the brain tissue due to the meningeal damage, causing inflammatory reactions.
On the basis of these facts, we assumed that meningeal damage was associated with shock-wave-induced IAS. To confirm this, we analyzed the correlation between the degree of dural microhemorrhage, which was non-invasively detected by near-infrared (NIR) reflectance imaging, and the extent of the expression of glial fibrillary acidic protein (GFAP), which is a marker of astrocyte reactivity and has been used for detection of IAS. 1,4,10,11 Behaviors of fibroblast and microglia were also analyzed to examine their association with IAS.
Methods
All requests for animals and procedures intended to be used in this study were approved by the Ethics Committee of Animal Care and Experimentation, National Defense Medical College, Japan (19035, 20006).
Animals
Male Sprague-Dawley rats 9–10 weeks of age were obtained from Japan SLC, Inc. and were housed in standard laboratory cages on a 12:12-h light/dark cycle with free access to food and water after arrival. Rats at 10–11 weeks of age weighing 300–375 g (n = 135) were randomly assigned to undergo one of the following two experiments, Experiment 1 or 2. Experiment 1 consisted of LISW exposure and NIR reflectance imaging + histological observation to characterize meningeal damage at 30–60 min post-exposure (Group 1, n = 8). Experiment 2 involved LISW exposure and NIR reflectance imaging + immunohistochemistry to examine meningeal damage and astrocyte reactivity at 1 day, 3 days, 7 days, 14 days, and 28 days post-exposure (hereafter described as day 1, day 2, etc.; Group 2, n = 127). Table 1 summarizes the number of animals used in each group including the sham control groups.
Number of Rats in Each Group in This Study
NIR, near-infrared; LISW, laser-induced shock wave.
Generation and characteristics of LISWs
The method used for generating an LISW in this study was described previously. 12 Briefly, a laser target, which was a light-absorbing material (0.5-mm-thick natural black rubber disk) adhered with an optically transparent material (1.0-mm-thick polyethylene terephthalate sheet), was placed on the tissue. For the interface between the rubber bottom surface and the tissue upper surface, ultrasound gel was applied for acoustic impedance matching. A Q-switched Nd:YAG laser (pulse width, 6-ns full width at half-maximum [FWHM]; Brilliant b, Quantel, France) was used to generate an LISW with a diameter of 4 mm as in the previous study. The laser pulse was focused onto the laser target, a laser-indued plasma was formed at the surface of the rubber sheet, and the plasma expansion was accompanied by generation of a shock wave. In this scheme, there is no direct interaction between the plasma and the tissue, and the tissue therefore interacts only with a shock wave. Pressure waveforms of the LISWs used in this study are shown in Figure 1A.

Characteristics of LISWs and site of transcranial LISW exposure for a rat.
In our previous study, we observed microhemorrhage in the meninges of the rat brain exposed to a single LISW, whereas there was no hemorrhage in the cortex in the laser fluence range of 1.0–1.5 J/cm2. 12 In this study, we used fluences in this range (1.0, 1.25, and 1.5 J/cm2) for single exposure, the corresponding impulses of LISWs being ∼14.7, ∼17.2, and ∼19.4 Pa・s, respectively. For multiple exposure (three shots), the fluence was fixed to 1.0 J/cm2.
The duration of LISW used in this study was as short as ∼1 μs (Fig. 1A), whereas the optimum shock-wave duration to be applied to the rat brain would be a few tens of microseconds, considering the anatomical differences between the rat brain and the human brain, 27 as noted in the Discussion section. In this study, we assumed that the shock-wave impulse was one of the most important parameters to determine brain injury. We used impulses within the range that had been shown to cause mild bTBI in rats by experiments using a shock tube. 28 A schematic arrangement for the LISW application is shown in Figure 1B. The model was based on the focal shock-wave exposure. Although real bTBI in humans is caused by the whole brain or even systemic exposure to a shock wave, the use of a focal shock wave enables analysis of the mechanism and neuropathology of bTBI through an inductive approach.
Transcranial NIR diffuse reflectance imaging for meningeal damage detection
To non-invasively detect meningeal damage after LISW exposure in vivo, transcranial NIR diffuse reflectance imaging was performed for the rat brain before and after LISW exposure (single exposure experiments only). In the NIR spectral region, light efficiently penetrates through the skull and reaches the cortex, and the diffusely reflected light can be detected outside the brain. With hemorrhage in the meninges, the NIR light is absorbed by the blood, and the intensity of the reflected light is therefore decreased. Thus, meningeal hemorrhage can be non-invasively detected by NIR reflectance imaging.
Rats were anesthetized with isoflurane inhalation (5% for induction; 1.5–2.0% for maintenance) and placed in a stereotactic frame. The head of each rat was shaved, the scalp was incised at the midline, and the left parietal bone was exposed. The exposed parietal region was illuminated with NIR light from a halogen lamp (HL 100E, HOYA-Schott Co., Japan) via a fiber optic guide equipped with a bandpass filter (800 ± 40 nm). The reflected light was imaged with a CCD (10 bit, 639 × 480 pixels, 1/60 s/frame; OZ-2, Omegawave, Inc., Japan) using software (LRI-V101, Omegawave, Inc.). Baseline images were first acquired, and then a laser target was placed on the parietal bone to generate an LISW. Figure 1B shows a schematic arrangement for LISW application. Immediately after LISW exposure, the laser target was removed and NIR reflectance imaging was performed for ∼10 min. After imaging, the scalp was sutured, and the rat was returned to the home cage. For each acquired NIR image, the light intensity profile across the center of the exposure region was obtained to assess percent decrease in the reflectance intensity from the baseline (ImageJ Fiji software; National Institutes of Health [NIH], USA). During the experiments under anesthesia, the body temperature of the rat was kept constant at 37.0 ± 0.5°C with a temperature-controlled body mat.
Histological observation of meningeal damage
To correlate meningeal damage detected by NIR reflectance imaging with the results of histological assessment, hematoxylin and eosin (H&E) staining was performed for the decalcified rat brain with the skull after imaging. Rats were deeply anesthetized with pentobarbital sodium (50 mg/kg) and transcardially perfused with normal saline containing heparin at a concentration of 1.0 U/mL followed by 10% neutral buffered formalin solution. After perfusion fixation, rats were decapitated, and the skulls with their contents were post-fixed with the same fixative solution for 24 h, defatted with 70% ethanol for 72 h, and decalcified in 0.5 M ethylenediaminetetraacetic acid (EDTA) for 6 days. Coronal sections of the decalcified samples were cut and embedded in paraffin, from which 4-μm-thick sections were obtained at the center of the exposure region and stained with H&E. The specimens were observed under a light microscope (Axiovert 200; Carl Zeiss Microscopy GmbH, Germany), and images were acquired with a CCD (Axiocam 506 color; Carl Zeiss Microscopy GmbH).
Immunohistochemistry
We assessed the time course of astrocyte reactivity based on immunohistochemical staining with a GFAP antibody for the rat brains at day 1, day 3, day 7, day 14, and day 28. To explore the associations of meningeal damage and microglial activation with astrocyte reactivity, we also conducted immunohistochemical staining with a fibronectin (FN) antibody and ionized calcium-binding adapter molecule 1 (Iba1) antibody, respectively. At each time-point, rats were deeply anesthetized with pentobarbital sodium (50 mg/kg) and transcardially perfused with normal saline containing heparin at a concentration of 1.0 U/mL followed by 4% paraformaldehyde phosphate buffer solution.
Extracted brains were post-fixed overnight in 4% paraformaldehyde. A 4-mm-thick coronal section was cut from each extracted brain at the site of LISW application, cryoprotected through a graded series of sucrose solution from 5% to 20%, embedded in optimal cutting temperature (OCT) mounting compound, and then frozen on dry ice. Coronal specimens 10-μm thick were obtained from an area between the center of the LISW exposure region and 1 mm anterior to the exposure center using a cryostat and adhered onto adhesive glass slides. Specimens were permeabilized in phosphate-buffered saline (PBS) containing 0.05% Tween 20 (Sigma-Aldrich Co. LLC, USA) for 10 min, blocked in PBS containing 5% Blocking One (Nacalai Tesque, Inc., Japan) and 0.05% Tween 20 (PBS-Tween 20-Blocking One) for 45 min at room temperature, and then incubated with primary antibodies diluted in PBS-Tween 20-Blocking One overnight at 4°C in a humidified chamber. Specimens were washed in three changes of PBS and incubated with the relevant secondary antibody diluted in PBS-Tween 20-Blocking One for 40 min at 37°C in a humidified chamber.
Primary antibodies were as follows: mouse anti-GFAP-Cy3 conjugated (1:800; Sigma-Aldrich Co.), rabbit anti-fibronectin (1:200; Abcam PLC, UK), and rabbit anti-Iba1 (1:500; Fujifilm Wako Chemicals Corp., Japan). A secondary antibody used was AlexaFluor 488-conjugated goat anti-rabbit IgG (1:800; Invitrogen Corp., USA). Specimens were washed in three changes of PBS. Coverslips were mounted using DAPI (4',6-diamidino-2-phenylindole) Fluoromount-G (Southern Biotechnology, Inc., USA) and viewed under a fluorescent/light microscope (BZ-X800; Keyence Corp., Japan).
For high-resolution observations of reactive astrocytes and microglia, stained brain specimens that were mounted using ProLong Gold Antifade Mountant with DAPI (Invitrogen Corp., USA) were imaged with a Quorum WaveFX laser-scanning confocal microscope (Quorum Technologies, Inc., Canada) at 400 × magnification, and images were captured with a CCD (EM-CCD C9100-24B; Hamamatsu Photonics, K.K., Japan). Finally, captured images were stitched together using a module from MetaMorph software (Molecular Devices, Ltd., USA), and the areas of interest were then cropped to show representative cellular morphological changes.
Image analysis
We analyzed temporal and regional characteristics of astrocyte reactivity and astroglial scarring in the rat brains exposed to an LISW(s). For given regions of interest (ROIs) on the representative coronal sections obtained from four to six animals in each group, we measured the fractional area of GFAP immunoreactivity (ImageJ Fiji software; NIH, USA) at day 1, day 3, day 7, day 14, and day 28. We set ROIs in three sites on each section: SGP, GM-WM, and structures lining the ventricles (ventricular wall, VW).
The ROI for the SGP was a 0.8-mm2 region just below the site of LISW exposure: 4.0 mm in width by 0.2 mm in depth along the cortical surface. For GM-WM, we drew a line on the boundary between the gray matter and white matter and then outlined upper and lower outer contours of an ROI at 0.2 mm above and below the boundary, respectively; the ROI was located just below the LISW exposure site. Because the length of the drawn boundary line varied depending on the specimen, the area of the ROI varied in the range of 1.0–1.6 mm2: horizontally, 2.5–4.0 mm along the gray-white matter boundary; vertically, 0.4 mm across the boundary. For the VW, an ROI was set above the lateral ventricle (LV) under the LISW exposure site, for which a lower outer contour of the ROI was determined on the upper edge of the LV and then an upper outer contour was outlined at 0.2 mm from the lower contour. The length of the upper edge of the LV also depended on the specimen and the area of the ROI ranged from 0.1 to 0.5 mm2: horizontally, 0.5–2.5 mm along the upper edge of the LV; vertically, 0.2 mm above the edge. To measure the fractional area of GFAP immunoreactivity on each fluorescence image, the areas of pixels with fluorescence intensities greater than or equal to those of cortical normal astrocytes were determined. For the areas, the mean fractional area of GFAP immunoreactivity was measured in each ROI.
Statistical analysis
Methods for statistical analysis of data obtained in this study were selected on the basis of the data uniformity of variance. Correlations between the mean fractional area of GFAP immunoreactivity and the percent decrease in NIR reflectance were analyzed by Pearson's correlation coefficients. Time-dependent changes in the mean fractional area of GFAP immunoreactivity were assessed by a two-way analysis of variance (ANOVA) with Sidak's multiple comparison tests for the SGP, GM-WM, and VW. Statistical analyses were conducted using GraphPad Prism software (Prism 6 for Windows; GraphPad Software, Inc., USA). Differences were considered statistically significant when p < 0.05. Data are represented as means ± standard deviation.
Results
Meningeal damage
Figures 2A–D show representative transcranial NIR reflectance images of the brains of a sham control rat and rats ∼10 min after single LISW exposure at different impulses. Dashed line circles in Figures 2B–D indicate the sites of LISW application, within which darker regions are seen, indicating meningeal hemorrhage. The average percent decreases in reflectance intensity from the baseline were ∼4% at the low impulse (14.7 Pa・s), ∼10% at the middle impulse (17.2 Pa・s), and ∼15% at the high impulse (19.4 Pa・s).

Transcranial near-infrared reflectance images of the rat ipsilateral (left) hemisphere after single LISW exposure
Figures 2E–H show the corresponding H&E-stained histopathological images of the skull and brain with meninges, where the dura is attached to the skull, and arachnoid and pia maters exist on the surface of the cortex. A large space between the skull (with dura) and the brain (with arachnoid and pia maters) was inevitably formed by shrinkage of the brain tissue due to the fixation and decalcification processes. In the specimen of sham control, the upper arachnoid layer and pial vessels were partially beneath the dura under the skull. At the low impulse (Fig. 2F), a very thin layer of red blood cells (RBCs) was observed between the skull and dura. At the middle impulse (Fig. 2G), the layer of RBCs was thickened, and RBCs had infiltrated within the dura. In both cases, no RBCs were seen in the cortex. At the high impulse (Fig. 2H), RBCs were observed both in the dura and the arachnoid mater, but hemorrhage was not evident in the cortex. The average total thicknesses of the RCB layers in the dura and/or arachnoid mater were ∼24 mm, ∼60 mm, and ∼104 mm at the respective impulses. There was an almost linear correlation between the reflectance decrease and hemorrhage thickness, indicating that transcranial NIR reflectance imaging is useful for in vivo, quasi-quantitative characterization of meningeal damage.
Astrocyte reactivity and fibrotic reaction
Figure 3 shows representative images of GFAP and FN immunohistochemical staining of the brains of (A) a sham control rat and (B–D) rats 7 days after single-LISW exposure at different impulses and (E) multiple-LISW exposure at the low impulse (ipsilateral hemisphere). An impulse-dependent increase in the GFAP immunoreactivity was observed. At the low impulse (Fig. 3B), there were slight increases in the GFAP immunoreactivity just below the pia mater and in the white matter. FN immunoreactivity in the arachnoid mater and pial vessels was also slightly increased.
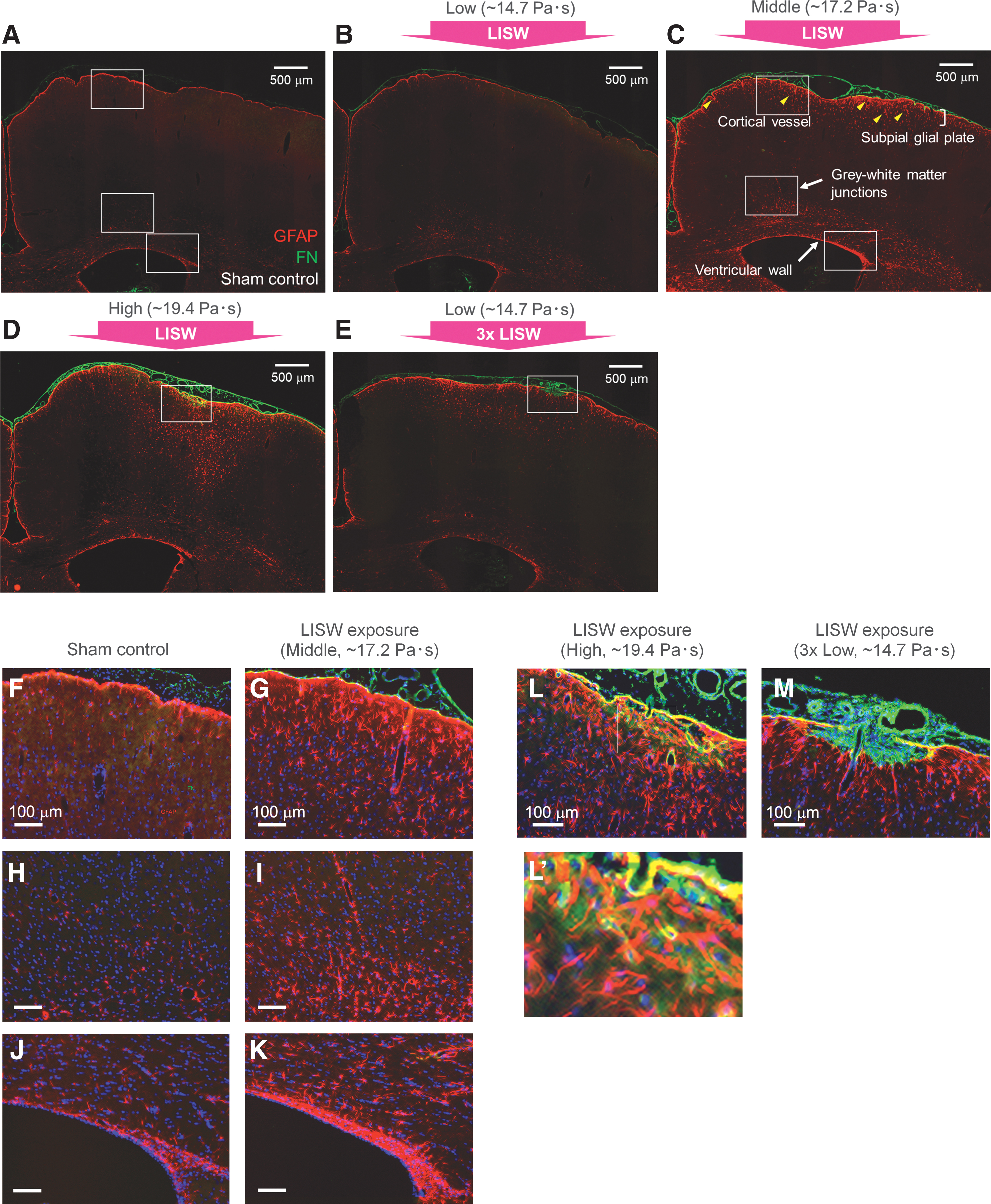
Representative images of immunohistochemical staining for GFAP and FN in the rat brains at 7 days post-exposure.
At the middle impulse (Fig. 3C), strong GFAP immunoreactivity was seen in the SGP, GM-WM, structures lining the ventricles (VW), and regions surrounding cortical blood vessels. Importantly, these regions are consistent with those of clinical IAS. In this condition, FN immunoreactivity was also strong between the arachnoid mater and the pia mater, and pial vessels were dilated and the subarachnoid space was extended around the exposure center. Figures 3F,H,J and 3G,I,K show magnified images of the portions of the SGP, GM-WM, and VW in Figure 3A (sham control) and Figure 3C (middle impulse), respectively. In Figure 3G,I,K many reactive astrocytes exhibit enlarged soma and extended foot processes.
At the high impulse (Fig. 3D), reactive astrocytes with strong GFAP immunoreactivity were more deeply distributed in the cortex. FN immunoreactivity was further enhanced between the arachnoid mater and the pia mater and it was also seen within the cortical surface region. Figures 3L and 3L’ show magnified images of the cortical surface region in Fig. 3D. GFAP immunoreactivity exhibits dense, hypertrophic reactive astrocytes with heterogeneously elongated processes. Enhanced FN immunoreactivity was seen on the pia mater and luminal structures in the subsurface cortex, where colocalization with GFAP immunoreactivity was seen, indicating secretion of FN in the astrocytes. There were also lattice-like structures of FN immunoreactivity around the reactive astrocytes just underneath the cortical surface (Fig. 3L’). These indicate fibrotic reactions involving activations of meningeal fibroblasts and astrocytes and their contributions to the deposition of the extracellular matrix (ECM) including FN.
With multiple exposure at the low impulse (Fig. 3E), relatively strong GFAP immunoreactivity was seen in the SGP and around the cortical blood vessels, which were more distinct than those with single exposure at the same impulse. Strong FN immunoreactivity was observed not only in the subarachnoid space but also in the cortical surface region; Figure 3M shows a magnified image corresponding to these regions. The pia mater/glia limitans is disrupted, and FN-positive cells have locally proliferated between the arachnoid mater and the subsurface cortex, indicating fibrotic scarring. The border between the fibrotic scar and the astroglial scar is clear, to which reactive astrocytes extend their elongated processes. FN immunoreactivity is partially co-localized with GFAP immunoreactivity at the border and the cortical surface.
Time courses of immunoreactivity of glial fibrillary acidic protein
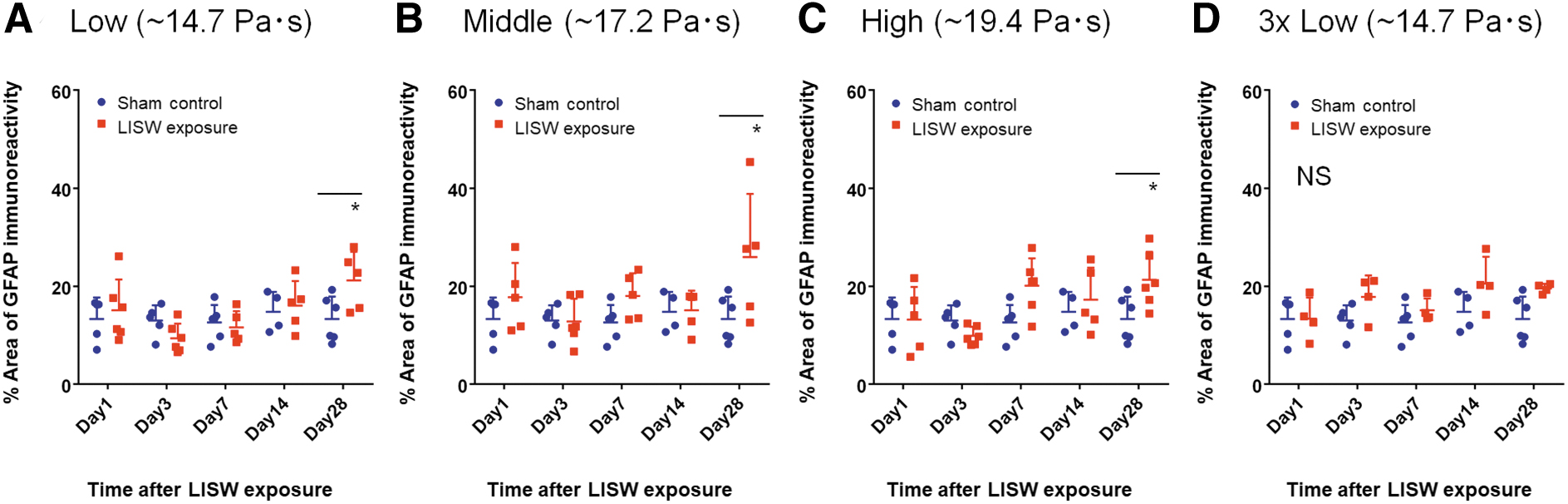
Figures 4–6 show time courses of mean fractional areas of GFAP immunoreactivity in the SGP, GM-WM, and VW, respectively, under different LISW exposure conditions. In the SGP with single exposure at the low and middle impulses, GFAP immunoreactivity was increased until 3 days post-exposure (day 3), but thereafter it was decreased (Fig. 4A,B). Compared with the respective sham control, there were significant differences in GFAP immunoreactivities at day 3 and day 7 at the low impulse and at day 3, day 7, and day 14 at the middle impulse. At the high impulse, GFAP immunoreactivity was increased until day 14, and it was still significantly higher at day 28 compared with the sham control. With multiple exposure at the low impulse, GFAP immunoreactivity continued to increase until day 7 (Fig. 4D), indicating that multiple exposure enhances astrocyte reactivity when compared with single exposure at the same impulse.

Time courses of mean fractional areas of GFAP immunoreactivity in the subpial glial plate.
Time courses of GFAP immunoreactivity for the GM-WM and VW were very different from that for the SGP. In the GM-WM with single exposure, GFAP immunoreactivity was not significantly increased for up to day 14 at any impulse, but it became significantly higher at day 28 at the low, middle, and high impulses, compared with the respective sham control (Fig. 5B,C). With multiple exposure (low impulse), no significant increase in GFAP immunoreactivity was observed at any post-exposure days. Results from the VW were almost the same as those for GM-WM. In the VW with single exposure, GFAP immunoreactivity was not significantly increased up to day 14 but significantly increased at day 28 at any impulse (Fig. 6A–C). With multiple exposure (low impulse), no significant increase in GFAP immunoreactivity was observed at any post-exposure days.

Time courses of mean fractional areas of GFAP immunoreactivity in the gray-white matter junction.
Time courses of mean fractional areas of GFAP immunoreactivity in the ventricular wall. (A–C) Single LISW exposure at low (A), middle (B), and high (C) impulses.
Correlation between meningeal damage and astroglial scarring
Figure 7 shows the results of correlation analysis for the meningeal damage (percent decrease in NIR reflectance intensity) and the increase in astrocyte reactivity (mean fractional area of GFAP immunoreactivity) in (A) the SGP (day 7), (B) GM-WM (day 28), and (C) VW (day 28) with single exposure at all the impulses including the sham control. There were statistically significant correlations between the NIR reflectance decrease and the GFAP immunoreactivity in all regions, that is, in the SGP with Spearman's correlation coefficient of r = 0.8159 (p < 0.0001), GM-WM (r = 0.5539, p = 0.0075), and VW (r = 0.5088, p = 0.0156), indicating very strong, strong, and moderate correlations, respectively. These results suggest that shock-wave-induced microhemorrhage in the dura is associated with astroglial scarring in the SGP, GM-WM, and VW.
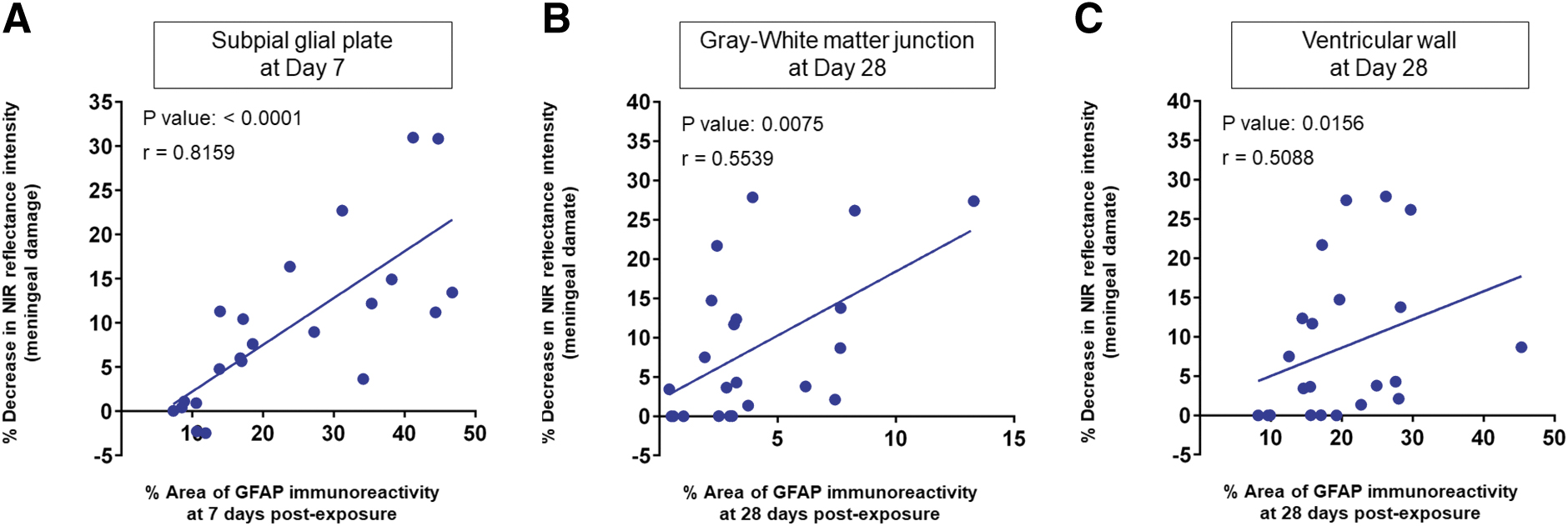
Correlation between meningeal damage (percent decrease in NIR reflectance intensity) and astroglial scarring (mean fractional area of GFAP immunoreactivity) in the SPG
Microglial activation
Figure 8 shows time-course confocal images of GFAP (red) and microglia (green) in (A) the SGP/cortex and (B) GM-WM of the sham control brain and the brains with single exposure at the middle impulse. Microglial cells in the sham control showed ramified morphology with their fine processes, small cell soma, and distal arborization. In the SGP with exposure, activated microglia showing retracted thick processes, enlarged cell soma, and decreased arborization were observed from day 1. The number of activated microglia was maximized at day 3, but thereafter it was decreased until day 28. In the GM-WM with exposure, there were few activated microglia at day 1, and the number of activated microglia was increased at day 3. After that, the number of activated microglia was decreased, but it was increased again at day 28. The different time courses of microglial activation between the SGP and GM-WM would be associated with the different time-dependent changes in GFAP immunoreactivity (Fig. 4 vs. Fig. 5), as discussed below.
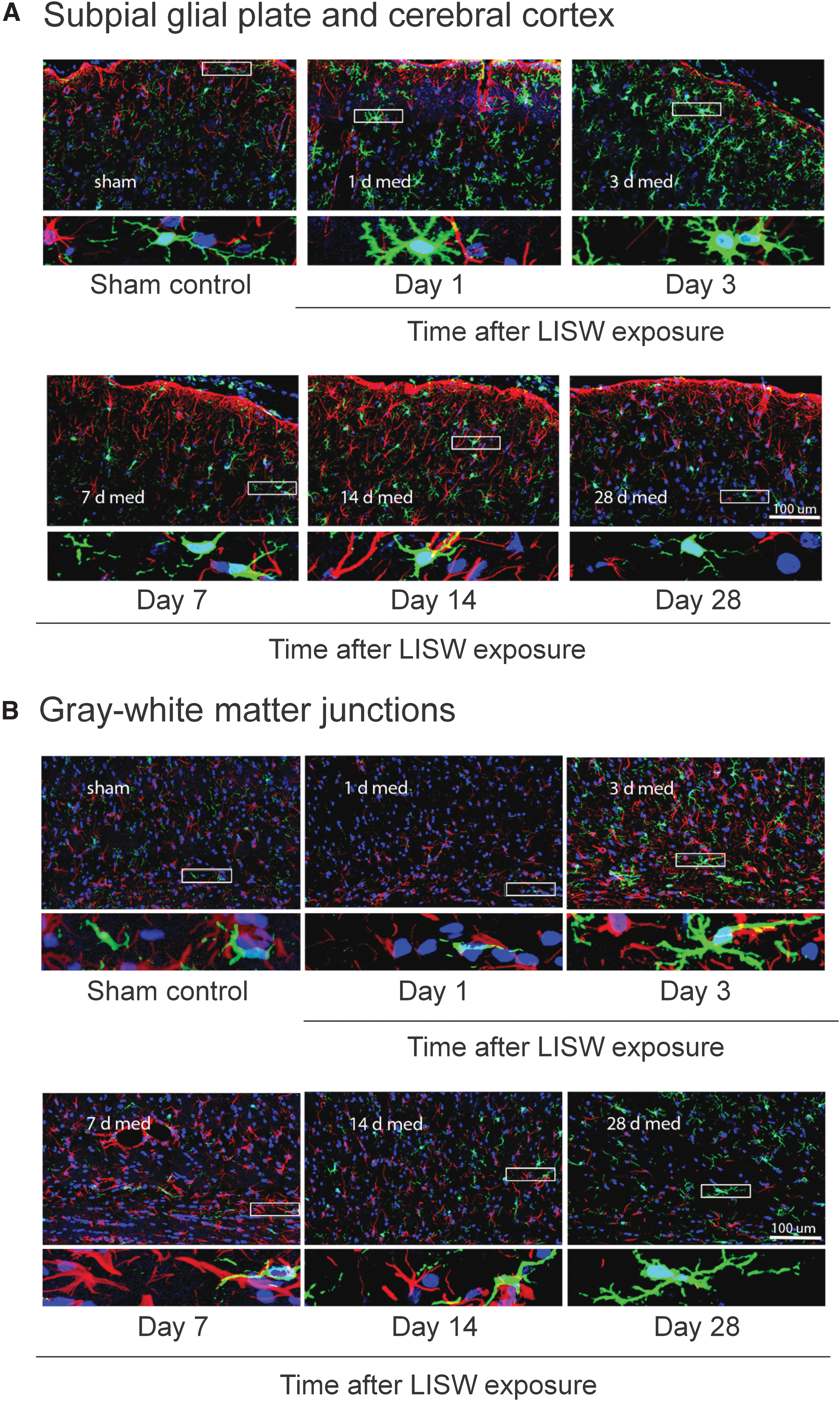
Representative confocal microscopic images for microglial activation in the subpial glial plate and cerebral cortex
Discussion
In this study, we observed increases in GFAP immunoreactivity in the SGP, GM-WM, VW, and regions surrounding cortical blood vessels in the rat brain exposed to an LISW(s), accurately replicating clinical IAS in rodents. Because an LISW is not accompanied by dynamic pressure, the results demonstrate that the primary mechanism alone causes IAS. The regions with increased GFAP immunoreactivity correspond to anatomical tissue boundaries, where pressure-wave reflections due to acoustic impedance mismatching can cause tissue alteration. Importantly, there were significant correlations between the degree of meningeal damage (dural microhemorrhage) and the extent of GFAP immunoreactivity, suggesting an association of meningeal damage with astroglial scarring. At the high impulse (∼19.4 Pa
The dural microhemorrhage observed in this study would be a characteristic phenomenon in shock-wave-induced brain injury. Because the dura mater and arachnoid mater are located between the skull (hard tissue) and CSF (liquid), the layers are considered to be inherently susceptible to shock-wave exposure due to the complex reflections of an incident pressure wave. Dural hemorrhage causes leakage of blood proteins and hematogenous cells, by which immunocompetent cells residing in the dura can be activated, producing inflammatory cytokines and chemokines. 29 These proteins can cross the arachnoid membrane due to its loosened tight junction caused by the shock-wave exposure. The pia mater would also be affected by shock-wave reflections, increasing its permeability. Accordingly, those proteins may also infiltrate into the cortex.
Blood proteins including thrombin and fibrinogen, 30,31 as well as various cytokines and chemokines, 32,33 can trigger inflammatory reactions, resulting in astrocyte reactivity in the cortical subsurface region. When the arachnoid mater and pia mater are mechanically disrupted, as observed in the case of the multiple exposure, hematogenous cells and immune cells, such as T cells and macrophages, can also directly infiltrate into the cortex, causing a more intense inflammatory response in the brain parenchyma. Meningeal fibroblasts could also infiltrate into the brain parenchyma due to the meningeal damage, leading to fibrotic scarring (Fig. 3M). 34 In this case, an astroglial scar was formed around the fibrotic scar to enclose the region with glial limitans. 29 Although it is not clear how meningeal bleeding due to mild TBI affects inflammatory regulations in the central nervous system (CNS), 35 our observations seem to provide some insight into the pathophysiology related to meningeal barrier disruption.
In the SGP, the time course of GFAP immunoreactivity was greatly dependent on the extent of meningeal damage. When meningeal damage was limited to the dural layers (low and middle impulses), average GFAP immunoreactivity peaked at day 3 (Fig. 4A,B), whereas when all of the meningeal layers were damaged (high impulse), it continued to increase until day 14 and was still significantly higher at day 28. Although the roles and characteristics of reactive astrocytes in the CNS are not fully understood, there is a concept that classifies reactive astrocytes into non-proliferative and proliferative subtypes. 36
Non-proliferative reactivity is directly initiated by the triggers and can resolve over time if acute triggers recede. 36 In this study, the reactive astrocytes that maximized at day 3 might be associated with the non-proliferative reactivity. On the other hand, proliferative reactivity is defined as reactivity that occurs in response to tissue damage and newly proliferating astrocytes form borders to separate and isolate damaged, inflamed, and fibrotic tissue from adjacent viable neural tissue (permanently separate damaged areas). 36 In this study, the high-impulse single exposure resulted in damage of all layers of the meninges with hemorrhage both in the dural and pial vessels (Fig. 2H), which could cause infiltrations of high concentrations of blood proteins and inflammatory cytokines into the cortical surface region and further enhance the inflammatory reactions. The reactive astrocytes that peaked at day 14 with the whole meningeal damage would be associated with proliferative reactivity.
With the high-impulse exposure, fibrotic reaction was distinct, in addition to astroglial scarring; FN immunoreactivity was co-localized with GFAP immunoreactivity in the SGP, indicating secretion and deposition of FN in the astrocytes (Fig. 3L). Such FN deposition in the reactive astrocytes can result from both the extravasated FN and synthesis and accumulation of astrocyte-derived FN through activation by the cytokine tumor necrosis factor-a (TNF-a). 37 This could promote a pro-inflammatory phenotype of astrocytes, exacerbating neuroinflammation by establishing an inflammatory loop. 37
It should also be noted that there was an FN-positive lattice-like structure around the dense reactive astrocytes (Fig. 3L’), which was not merged with GFAP positivity. The structure may indicate aberrant FN expression in the neural interstitial matrix. The neural interstitial matrix comprises a dense network of chondroitin sulphate proteoglycan (CSPG) and other ECM components and normally includes a small amount of FN. 38 Reactive astrocytes are known to be the principal source of CSPG in scar formation following traumatic CNS injuries, and the deposition of ECM components in the interstitial matrix is stimulated by the growth factors and inflammatory cytokines. 38 Although the exact roles and mechanisms of such FN deposition are still not known, persistent deposition of the ECM could be a part of the inflammatory reactions associated with reactive astrocytes. The low-impulse multiple exposure caused disruption of the pia mater, leading to fibrotic scar formation (Fig. 3M). Fibrotic scar is known to be formed by proliferating meningeal and perivascular fibroblasts, secreting ECM components such as type IV collagen, FN, and laminin. 29 The lesion scar would be a major barrier to neuronal regeneration and thus result in neuronal dysfunction.
The time courses of GFAP immunoreactivity for the GM-WM and VW considerably differed from that in the SGP, suggesting a different mechanism for astrocyte reactivity. The extent of meningeal damage was strongly correlated with GFAP immunoreactivity in the GM-WM and was moderately correlated with that in the VW at day 28, and astrocyte reactivity in these regions seemed, therefore, to be also associated with meningeal damage. In the GM-WM, GFAP immunoreactivity was increased at day 14 and day 28, and correspondingly, activated microglia were increased. This suggests a delayed onset of inflammation when compared with that in the SGP. Importantly, the white matter is thought to be one of the main routes for interstitial solute movements in cerebral waste clearance 39 ; the GM-WM can be the channel for interstitial fluid and CSF from the cortical parenchyma and the subarachnoid space through the perivascular space. Thus, the damage to this region may impair the waste clearance function, leading to increased exposure to various substances in the brain. The time course of GFAP immunoreactivity in the VW seemed to be similar to that in the GM-WM. Periventricular white matter (VW) may also be affected by waste clearance dysfunction.
The impulse is believed to be one of the most important parameters to determine shock-wave-induced tissue injury; the impulses of LISWs used in this study were within the range of actual improvised explosive device (IED)-related shock waves. 28 However, the durations of LISWs were ∼1 μs (Fig. 1D), which are much shorter than those of actual IED-related shock waves. 40 Thus, future experiments using an LISW with the optimum duration are needed to recheck the results obtained in this study. As Needham and colleagues pointed out, because rodents' brains are roughly 10 times smaller in dimension than human brains, the appropriate duration for rodent experiments would be 10 times shorter than the duration of real blast shock waves to make the shock-wave transit time within the skull the same. 27 Courtney and associates reported typical positive durations of representative blast sources as follows 41 : 0.01–0.2 ms for underwater blast, 42 0.25–0.7 ms for IED/mine, 40 and 1–10 ms for larger, conventional high explosives. 43 Thus, the appropriate shock-wave duration would be in the range of 25–70 μs to replicate IED shock-wave-caused phenomena in the rat brain. We have recently developed a method to extend the LISW duration to ∼50 μs. We plan to confirm the reproducibility of our results with longer duration LISWs in the next study.
From the anatomical point of view, we should also note that the sulci and gyri are important interfaces potentially associated with IAS, but rodents lack them. Actually, in the ferret model described above, intense GFAP immunoreactivity was observed in the fundus of sulci, 10 but for humans, there are still few data on the predilection for the sites of astroglial scarring in the sulci and gyri, although the SGP is involved in them. Further study is needed to understand astroglial scarring in the gyrencephalic brain. It should be noted, however, that important four interfaces for IAS are involved in our model.
Another crucial future challenge is to investigate the detailed mechanisms of shock-wave-induced meningeal damage based on computer simulation, for which a multi-layer meningeal model for a finite element method is now under development. Another limitation of the current study includes the fact that only GFAP was used as a marker of glial scarring. We plan to use markers associated with scar-forming cellular components and molecular triggers that lead to scar formation in future work. 44 From a therapeutic point of view, it would also be important to assess the roles of astrocyte reactivity and astroglial scarring, which could be both negative and positive, in our bTBI model. We plan to clarify this point using a method such as spatial transcriptomics for future translational research.
Conclusions
In this study, we successfully replicated IAS, which has been reported as a signature clinical neuropathology of bTBI, in the rat brain exposed to an LISW. This demonstrates that the primary mechanism alone caused IAS. Our in vivo transcranial NIR reflectance imaging showed a significant correlation between the degree of meningeal damage (dural microhemorrhage) and the extent of expression of GFAP. Both the meningeal damage and IAS are thought to result from pressure-wave reflections due to the acoustic impedance mismatching at anatomical tissue boundaries. Our model would be useful for replicating shock-wave-related IAS and examining its therapeutic strategy.
Transparency, Rigor, and Reproducibility Summary
Neither the study design nor analysis plan was formally pre-registered. Subjects were randomly divided to the experimental Group 1 or Group 2. Experimental manipulations for the animals, that is, LISW exposure, transcranial NIR diffuse reflectance imaging, and transcardial perfusion fixations, were performed between the hours of 08:00 and 18:00. Sectioning and staining of the brain tissues, as well as all the relevant analyses were performed by investigators blinded to experimental groups. Sample sizes per group for each outcome measure were planned on the basis of previous studies in which similar sample sizes were shown to be sufficient for detecting pathophysiological changes due to shock-wave exposure. Slides for staining were allocated across batches to ensure consistency of staining across different experimental conditions, that is, days after shock-wave exposure, and all slides were imaged within 1 day after staining.
In sectioning brain tissue, cryostat malfunction occurred for ∼30% of samples during the experiments, and thus additional sample collection was required. When we used different lots of primary or secondary antibodies, the reproducibility of staining was checked using standard slides obtained from the same rat brain samples with and without LISW exposure. The selection of methods for statistical analysis was based on the assumption of data uniformity of variance; when this assumption was breached, non-parametric tests were used in place of parametric tests. The authors agree to provide the full content of the manuscript upon request by contacting the corresponding authors.
Footnotes
Authors' Contributions
Conceptualization: SK, SSa; methodology: SK, ST, YK, IN, SM, SSa; validation: SK, AK, YM; formal analysis: SK; investigation: SK, AK, YM, GH, SSe, ST, YW; data curation: SK, AK, YM, YH, YW; writing–original draft: SK, SSa; writing–review and editing: SK, IN, SM, YW, SSa; visualization: SK, YW; supervision: SSa; project administration: SK, SSa; funding acquisition: SSa.
Funding Information
This work was supported by the Defense Medicine Basic Research Program under the Ministry of Defense.
Author Disclosure Statement
No competing financial interests exist.