
Abstract
Motorized cycling (MC) is utilized as an alternative to traditional exercise in individuals who are unable to perform voluntary movements post-spinal cord injury. Although rodent models of MC often show more positive outcomes when compared with clinical studies, the cause of this difference is unknown. We postulate that biomechanical differences between rats and humans may contribute to this discrepancy. To begin to test this theory, we examined pedal reaction forces and electromyography (EMG) of hindlimb muscles as a function of cycle phase and cadence in a rat model of MC. We found that higher cadences (≥30 RPM) increased EMG and force, with higher forces observed in animals with contusion injuries as compared with transections. To further investigate the forces, we developed a technique to separate rhythmic (developed with the motion of the pedals) from nonrhythmic forces. Rhythmic forces resulted from induced eccentric muscle contractions that increased (amplitude and prevalence) at higher cadences, whereas nonrhythmic forces showed the opposite pattern. Our results suggest that muscle activity during MC in rats depends on the stretch reflex, which, in turn, depends on the rate of muscle lengthening that is modulated by cadence. Additionally, we provide a framework for understanding MC that may help translate results from rat models to clinical use in the future.
Introduction
Exercise and motorized cycling (MC) are used to target central nervous system plasticity and/or cardiovascular health post-spinal cord injury (SCI). Despite the widespread use of MC, clinical studies have shown very limited beneficial effects on spasticity 1,2 and arterial peripheral circulation. 3,4 Conversely, rat models of MC have demonstrated effects on H-reflex habituation, 5 wind-up of stretch reflexes, 6 electrophysiological properties of motoneurons, 7 and cardiovascular properties such as stroke/end diastolic volume, 8 among other benefits. These contradicting results suggest a fundamental difference between the animal model and clinical practice.
During MC, the feet/paws are attached to the cycle pedals, and an electric motor turns the pedals at cadences that generally range from 30 to 50 or 25 to 60 RPM for rat 8,9 and human studies, 10,11 respectively. The combination of leg length, crank length (distance from pedal to crank shaft), and cycle cadence combines to determine the range of motion and rate of change in the limb joint angles and thus are sources of variability in the limb responses and the magnitude of muscle activation that occurs during MC. Although often termed “passive,” MC was shown to elicit rhythmic electromyography (EMG) activity in the soleus of a rat after a T10 transection that corresponded with the flexion of the ankle. 12 This pattern suggests that spinally mediated muscle activation during MC is caused by stretch reflexes below the level of the lesion. Because muscle spindles are responsive to length and rate of lengthening, 13 differences in biomechanics due to the crank length/leg length ratio and/or cadence during MC could cause varying magnitudes and patterns of muscle lengthening and thus activation between species or even individuals. This theory suggests that greater muscle activation could be achieved by altering parameters such as range of motion (crank length/leg length ratio) or cadence; however, any modifications must be considered in the context of SCI.
Spasticity is a common issue in the SCI population that could be induced during MC due to dependence on movement velocity and limb position. 14,15 Although limb and pedal position during MC is set to not overextend the limb or overstretch the muscles, increases in cadence naturally increase the rate of muscle lengthening. Furthermore, maladaptive afferent input from hindlimb stretching has been shown to induce eccentric muscle contractions 16 and reduce locomotor function in a rat model of SCI, 17 even when applied in a phasic pattern. 18 The reduction in locomotor function was associated with an increased density of calcitonin gene-related peptide positive (CGRP+) afferents in the dorsal horn, 17 suggesting that maladaptive plasticity may play a role. While exercise has been shown to reduce the density of CGRP+ afferents in the dorsal horn, 19 this is thought to be because of task-specific training. Muscle contractions during MC would not only be eccentric but would also be opposite to those seen in overground locomotion or active cycling. Furthermore, activity-dependent plasticity can be increased by MC training via a modification of phosphatase and tensin homolog and mechanistic target of rapamycin signaling 20 ; however, whether these changes are adaptive or maladaptive is unknown. It is possible that the effects of afferent information associated with MC-induced eccentric contractions may have a negative effect on locomotion, like manual muscle stretching has, as spinal learning is dependent on afferent input that is coordinated with limb position. 21
Another challenge in translating animal SCI research is the injury model itself. Most studies employ injuries that target a specific spinal level and injury severity. A complete transection is often employed to reduce variability and to avoid the impact of spared axons/tracts that occurs with contusion models. However, clinical SCI is highly variable in terms of level and injury severity, with the majority of patients presenting with incomplete neurological injury. 22 The rat model of MC typically utilizes a complete transection, which potentially further limits its clinical relevance/translation.
In conclusion, the underlying causes of improvements seen from MC are poorly understood, and the apparent differences between animal and human studies are unclear. To better understand the contributions of stretch reflex activity in a clinically relevant model, it was the goal of this study to first characterize the effects of MC at different cadences and with two different crank lengths on hindlimb EMG responses and cycling load (pedal reaction forces). Second, we sought to determine if daily MC influences EMG responses, pedal reaction forces, and locomotor/sensory function over time in both a contusion and transection rat model of SCI. We hypothesized that EMG and cycling load would correspond to the rate of muscle lengthening and would be modulated by both cycling cadence and crank length. Furthermore, given that previous MC studies found no significant impact on locomotor function, 23 we hypothesized that daily training would reduce nociceptive hypersensitivity while having no effect on locomotor function.
Materials and Methods
Experimental design
This goal of this study was to describe the biomechanics of a rat model of MC and how these mechanics affect the magnitude and timing of EMG activity and force production during cycling. We furthermore sought to understand how daily cycle training affects locomotor and sensory function and if these changes are reflected in the spinal cord. Cycling began 1 week post-SCI and consisted of both training and evaluation cycling paradigms. Training was performed Tuesday to Thursday (30 min/day at 45 RPM), whereas evaluations were performed Monday and Friday (8–9 min). Other outcome measures were performed throughout the week before cycling began. Animals in the transection group were euthanized at 4 weeks postinjury, whereas animals in the contusion group were euthanized 5 weeks postinjury. This difference was due to time constraints, as animals had to be euthanized ∼1 h after their final cycling session for histological outcome measures. A timeline of the experimental procedures and pedal design can be found in Figure 1, where Figure 1A shows the timeline of experimental events while Figure 1B details the weekly training and assessments that began on week 1 postinjury.
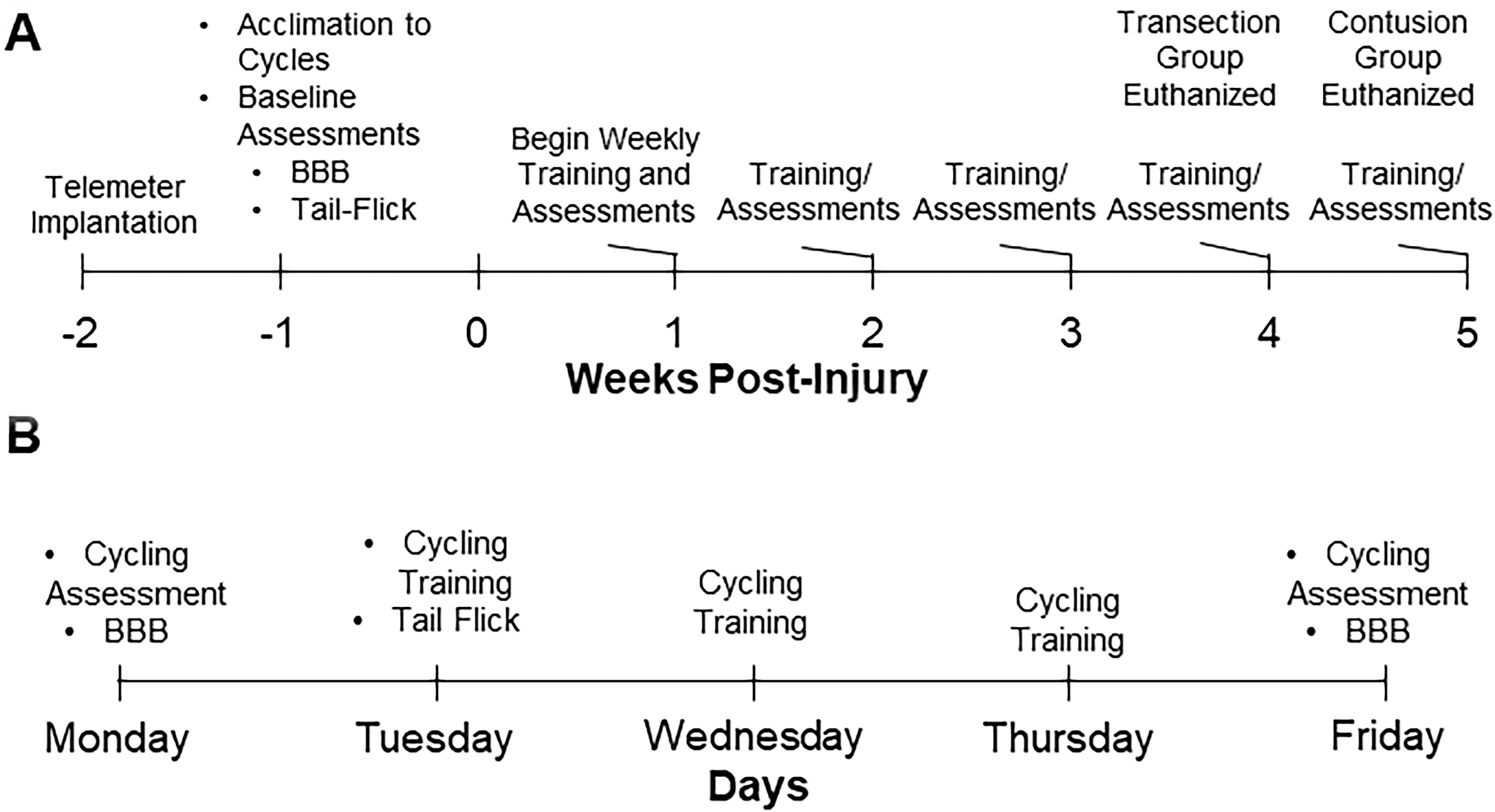
Timeline of experimental procedures. The overall experimental design is shown in
Animals, EMG transmitter implantation, and SCI
Twenty-three adult female Sprague–Dawley rats (225 ± 16 g, Envigo) were used for this experiment. All procedures involving animals were approved by the Institutional Animal Care and Use Committee at the University of Louisville. Animals were acclimated in their home cages and then handled and exposed to all testing devices used in the study for 2 weeks before evaluations. Handling was continued throughout the experiment on days when the animals did not receive cycling. Following acclimation, animals were divided into four groups: contusion/cycling (n = 9), contusion/no cycling (n = 4), transection/cycling (n = 6), and transection/no cycling (n = 4). Of the contusion/cycling group, four animals were randomly selected to be implanted with EMG transmitters 3 weeks before SCI. Animals were anesthetized for all surgeries using a ketamine (50 mg/kg)/xylazine (0.024 mg/kg) cocktail to provide a surgical level of anesthesia as confirmed by the absence of paw withdrawal reflexes in response to a strong toe pinch. The upper back of each animal was shaved and treated to produce an aseptic incision area. A small skin incision was made between the shoulder blades to allow placement of the transmitter (HD-X02, DSI). The transmitter wires were tunneled subcutaneously to the left hindlimb by separating the skin from the muscle layer using blunt dissection. A small incision was made over the thigh of the animal, allowing the electrodes to be inserted into the main belly of the vastus lateralis (VL) and biceps femoris (BF) muscles. Sutures were used to hold the electrodes in place. The incisions were closed using sutures, and animals were allowed to recover, receiving our standard postoperative care. Three weeks after this surgery, the animals received either a moderate–severe spinal cord contusion injury (25 g/cm, NYU Mascis Impactor) or complete transection at the T10 level as previously described. 24 The animals were allowed to recover for 7 days before cycling.
Kinematic recordings, locomotor assessments, and pedal forces
Locomotor function was assessed using the Basso, Beattie, Bresnahan (BBB) locomotor scale. 25 Baseline measurements were taken preinjury (pre- and posttransmitter implant) and then biweekly on Monday and Friday.
Hindlimb kinematics were recorded using two sagittal-oriented cameras (Basler AG) at 100 Hz in a custom LabView program. Animals were marked using 3 mm OptiTrack hemispherical reflective markers (Planar Systems) placed on the skin over the iliac crest (anterior rim of the pelvis), hip (head of the greater trochanter), ankle (lateral malleolus), and toe (fifth metatarsophalangeal joint). Videos were digitized and converted into three dimensional (3D) coordinates using the MaxTraq software package (Innovision Systems Inc). It should be noted that the knee was not marked since excessive movement of the skin relative to the joint results in inaccurate information. 26 To account for this, the femur and tibia of each rat were dissected and measured postmortem. These measurements were used to scale custom models of each rat 27 in the OpenSim platform. 28 Inverse kinematics were calculated using the measured marker locations, which were then analyzed using custom MATLAB scripts (MathWorks). Cycling kinematics were recorded during the evaluation cycling protocol, which consists of 20 cycles at 6 cadences (5, 10, 15, 30, 45, and 60 RPM for 120 cycles total) applied in a random order without breaks in between. Pedal reaction forces were collected using a custom-designed pedal that incorporated a triaxial multidirectional force sensor (Nano17 SI-12-0.12, ATI Industrial Automation). Three hemispheres forming a right angle were added to the outside of the pedal, allowing it to be tracked along with other kinematic markers. It should be noted that the modified pedal and shorter crank were 3D printed using polylactic acid filament (Hatchbox 3D) at 30% density.
Single-limb hindlimb cycling kinematics were collected in synchrony with EMG (described below) and forces (1000 Hz) using a custom LabView script weekly on Monday and Friday. After applying markers as described, the hindpaws of the rat were attached to the pedals using 3M transpore surgical tape with the pedals oriented 180° out of sync, and the rat was secured in place by a strap with Velcro.
EMG recordings
EMGs were recorded at 1000 Hz using LabChart 8 (ADInstruments) during cycling evaluations and exported for further analysis to MATLAB. Although the transmitter communicates to the receiver at a frequency of 455 kHz to reduce signal contamination, testing was performed before implantation to insure no contamination from external sources such as the force sensor or cycle motor. This, combined with isolation of the transmitter and electrodes within the animal, ensured a high-quality EMG signal with a signal-to-noise ratio >100. Kinematic recordings were upsampled to 1000 Hz and synchronized to the EMG recordings, which were then used to divide EMG into gait cycles for further analysis. Cycles were divided into flexion and extension based on toe trajectories, where extension was defined as minimum to maximum while flexion was defined as maximum to minimum.
Data processing
Forces
Forces were processed before analysis using a custom MATLAB script. Briefly, a second-order zero-lag lowpass filter with a cutoff frequency of 6 Hz was applied to the raw data. The data were then separated into individual cadences, and the fundamental frequency (f0) was identified at each cadence. A notch filter was applied at f0 and then the subsequent two harmonics (2*f0 and 3*f0). The resultant force was used as the spastic force, whereas the nonspastic force was calculated as the difference between the original and spastic forces.
Nonspastic forces were divided into flexion and extension phases, and the normalized area under the curve (AUC) was used for analysis (total AUC/cycle length). This process was performed on the cycling rats under isoflurane preinjury; these normalized values (greatest normalized AUC from each cycling cadence) were subtracted from awake cycling to approximate the effects of limb inertia and gravity. Spastic forces were also divided into flexion and extension; however, peak values were used for analysis. Peaks >20% body weight were detected, and trials containing a spasm were identified as trials that contained one or more peaks. Heatmaps were created by defining the onset and termination of each spasm (individual spasms were defined as a gap >1 s between peaks) and representing activity as either “on” or “off.”
EMG
After collection, the raw EMG signal was high-pass filtered at 50 Hz to remove movement artifacts and then rectified and separated into flexion and extension phases for analysis. Root mean square values were calculated for each cycle to allow analysis of EMG magnitude. This technique was limited to analysis between cadences in individual animals at discrete timepoints due to nonphysiologically relevant variations in EMG magnitude. As obtaining maximum voluntary activations in rats is not possible, EMG was instead normalized using the Teager–Kaiser energy (TKE) operator combined with morphological operators as described by Sinha 29 and Yang et al. 30 Briefly, this method detects and classifies bursts of EMG activity as “on,” with periods of inactivity classified as “off.” The resultant signal is therefore classified on a scale of 0–1, where 0 represents no muscle activity and 1 represents muscle activity throughout the duration of the cycle.
Sensory assessments
A tail-flick assay was employed to assess thermal sensitivity using a radiant heat tail-flick device (Columbus Instruments). A rat was placed on the platform and gently restrained, whereas the base of the tail was placed above the photocell for thermal stimulation. The intensity of the heat source was set to 10 out of 25, which is calibrated to elicit a tail-flick latency of ∼3 s in neurologically intact rats and a maximum duration of 10 s for each trial to avoid tissue damage. For each rat, tail-flick latency was measured three times to the nearest 0.1 s with at least 2 min between measurements. A baseline assessment was performed preinjury, and subsequent tests were performed weekly on Tuesday.
Euthanasia and tissue histology
Animals were sacrificed 1–2 h after the last cycling evaluation using a ketamine (50 mg/kg)/xylazine (0.024 mg/kg) cocktail and transcardially perfused with 4% PFA. Spinal cords were dissected and postfixed in 4% paraformaldehyde (PFA) for 2 h and then transferred to 30% sucrose for cryoprotection. The injury epicenter was separated from the contusion animals, whereas the L1–L5 segments were separated from the rest of the spinal cord of all animals under a dissecting microscope and blocked in optimal cutting temperature (OCT) compound. The injury epicenter of the transected rats was determined completely by visual inspection. For the injury epicenters, transverse sections were cut at 50 μm on a cryostat.
Transverse sections were cut at 20 μm on a cryostat for immunohistochemical analysis into six blocks. Immunohistochemistry was performed using calcitonin gene-related peptide (CGRP) and protein c-Fos (c-Fos) antibodies as previously described. 31 Briefly, sections of L3 spinal cord were warmed and washed with PBS and 0.3% PBS-Triton (PBST) and then blocked for 1 h with PBST, 5% bovine serum albumin (BSA), and 10% normal donkey serum (NDS). After blocking, slides were washed and incubated overnight with CGRP primary antibody (guinea pig polyclonal anti-CGRP, 1:1000, 20R-CP007, lot #P17101902, Fitzgerald), 5% BSA, and 5% NDS at 4°C. Following incubation, sections were washed and incubated with a secondary antibody (Alexa Fluor 594-conjugated Donkey anti-guinea pig, 1:200, 706–585–148, Lot #129041, Jackson ImmunoResearch Laboratories) for 1 h. For c-Fos, sections of L2–L4 spinal cord were warmed and washed with PBS and PBST. Sections were then incubated in antibody diluent reagent (Invitrogen # 00-3218, Lot1966331A) with c-Fos primary antibody (mouse monoclonal anti-c-Fos, 1:1000, cat#ab208942, Abcam) overnight at 4°C. The next day, sections were washed and a secondary antibody (donkey anti-mouse FITC, 1:250, cat#715-096-151, Jackson ImmunoResearch Laboratories) was combined with antibody diluent reagent for 1 h at room temperature. Following incubation, slides were washed, and a secondary amplification process was performed using AlexaFluor 488 (mouse anti-FITC, 1:500, cat#200-542-037, Jackson ImmunoResearch Laboratories) in antibody diluent reagent for 1 h at room temperature. Sections were then washed with 4′,-6-diamidino-2-phenylindole (DAPI; 1:1000) in PBST for 5 min. After a set of three washes, Sudan Black B was dissolved (1 mg/mL) in 70% EtOH and applied for 15 min at room temperature. Slides were coverslipped using fluoromount after a final set of washes.
Image analysis
Images of each section were acquired using a Nikon Ti2 microscope (Nikon Corporation) using a 20× objective for all histological analyses (resolution 0.32 μm/pixel). An area encompassing each section was defined, and images were stitched such that the entire section was represented in one image. Image analysis of c-Fos and CGRP was performed as previously described. 31 Briefly, CGRP area was quantified in the dorsal horns, excluding the dorsal root entry zones, which were removed manually from each image. Analysis of images was conducted using MATLAB (MathWorks). For c-Fos analysis, each c-Fos positive nucleus was manually identified and marked based on overlap between c-Fos and DAPI using Nikon Elements software. To generate contour plots and heatmaps, previously marked neurons were exported from Nikon Elements. A custom-made MATLAB program was then developed to reconstruct and normalize the position of labeled neurons across sections. A reference axis was created for each image with the origin centered on the central canal, the y-axis parallel to the spinal cord midline, and the x-axis orthogonal to the y-axis. Contour/scatter plots were constructed using R. Distribution contours were created by calculating the 2D kernel density (using the kde2d function in the MASS library) and then connecting points of equal density values between 30% and 100% of the estimated density range in increments of 10%.
Statistical analyses
Data were analyzed using SPSS (IBM SPSS Statistics for Windows, Versions 26/27, IBM) except for the linear regressions shown in Figure 4B and D, which were created using Prism, Version 9 (GraphPad Software). Distributions were checked for normality; if data were normal, a repeated-measures analysis of variance (RMANOVA) was used for analysis, with strict post hoc t tests (Bonferroni t test for multiple comparisons). In instances where data were not normal (c-Fos), a Mann–Whitney U test was used.
Results
Muscle activity during MC
An example trial is illustrated in Figure 2, where rectified EMG data for each cadence are shown in Figure 2A, and EMG normalized using the TKE operator is shown in Figure 2B. EMG analyses of the BF and VL are shown in Figures 3A and 3B, respectively. Percent activation was evaluated using TKE for BF and VL at each week to examine the effect of phase (flexion vs. extension) and cadence (low vs. high) using RMANOVA. In the BF responses, significant effects of cadence were found at weeks two (F 1,8 = 8.1, p < 0.05), three (F 1,7 = 9.1, p < 0.05), and five (F 1,12 = 16.0, p < 0.01), although no effects were found for phase. Similarly, significant effects of cadence were found in the VL at weeks two (F 1,9 = 10.9, p < 0.01), three (F 1,11 = 12.3, p < 0.01), and five (F 1,11 = 28.7, p < 0.001), and significant effects of phase were found at weeks two (F 1,9 = 7.0, p < 0.05), three (F 1,11 = 7.0, p < 0.01), four (F 1,5 = 8.0, p < 0.05), and five (F 1,9 = 9.4, p < 0.05). Although the sample size is small (n = 3), the data suggest that activation of both muscles occurs more on average for both the BF and VL at higher cadences, with more differences seen during extension, when the flexors are being lengthened. However, it appears as though the VL is preferentially activated during flexion, although an increased sample is warranted before definitive conclusions can be drawn.
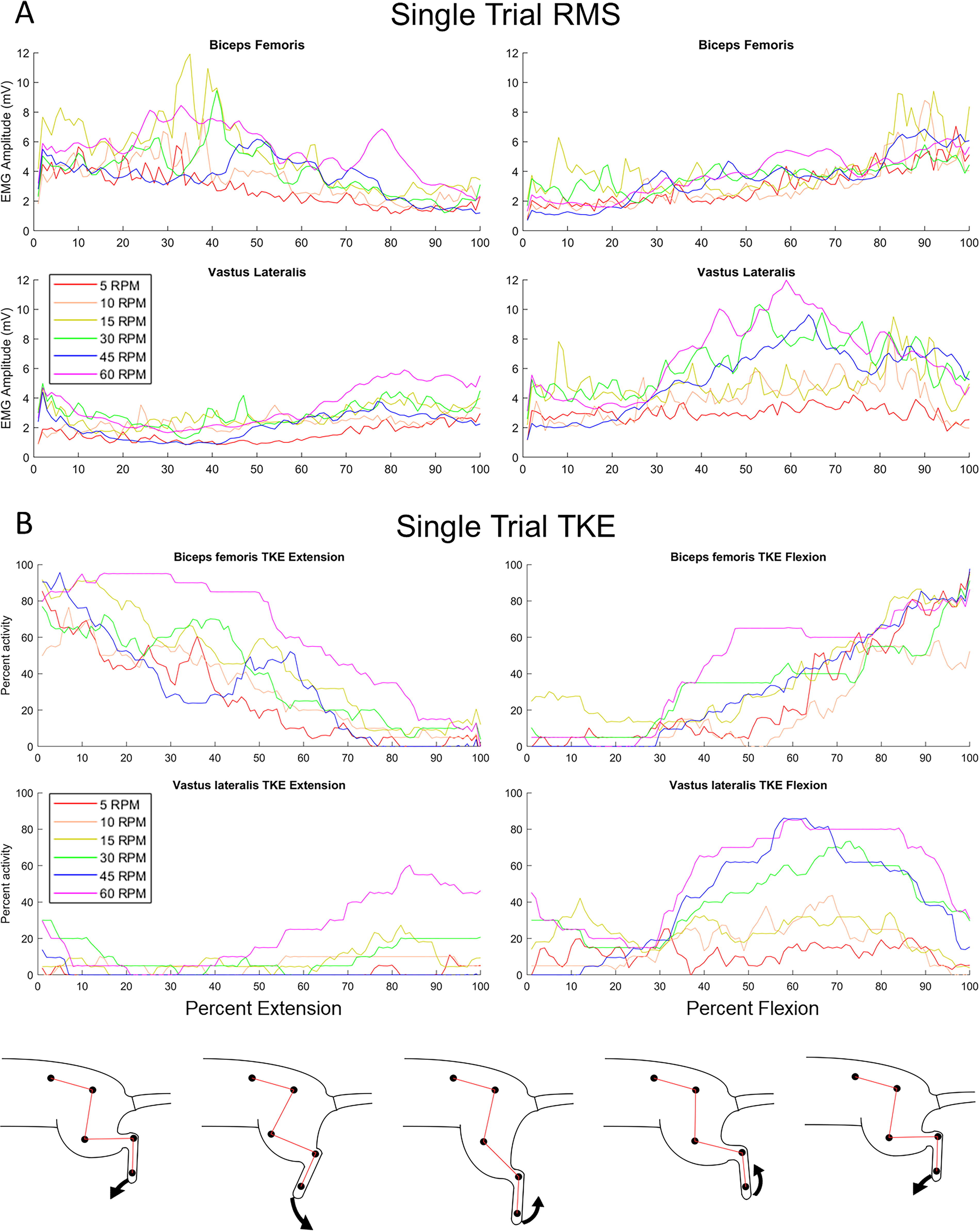
Example trial of EMG analysis.
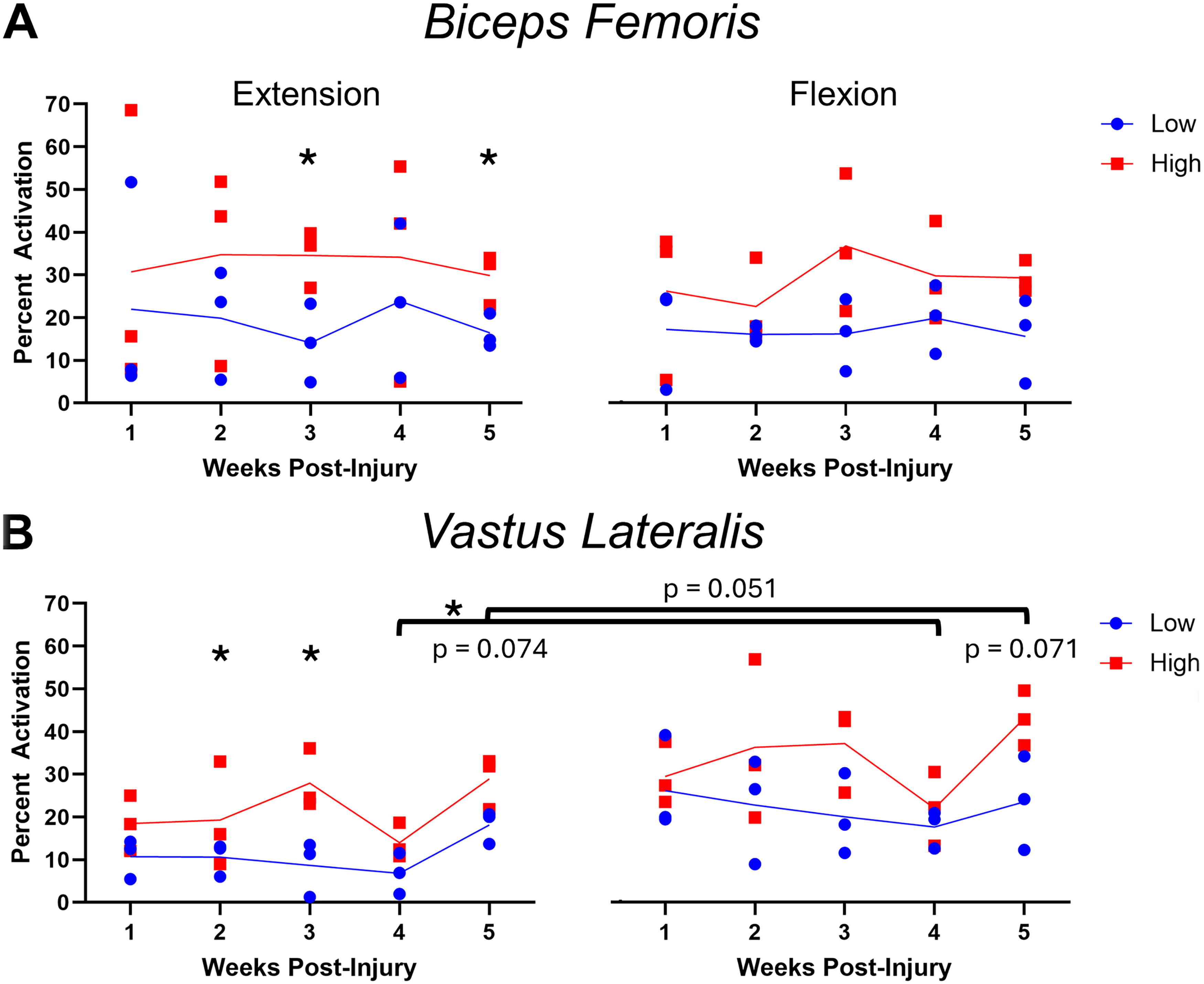
Comparison of EMG responses in the BF and VL during MC using percent activation.
Spastic and nonspastic forces during MC
An example of the relationship between force (spastic and nonspastic) and EMG is shown in Figure 4 for high and low cadences. Figure 4A and C contains example force traces along with EMG activity in the VL and BF obtained during the same cycling session, with the flexion phase indicated by the gray box. During high-cadence cycling, bursts of EMG in the VL are seen during flexion, which result in mild nonspastic force. EMG increases until a spasm is seen; however, the muscle activity still appears to be primarily during flexion. This is reflected in the force trace, which contains both a spastic and nonspastic component. In contrast, the spasms during low-cadence cycling (Fig. 4C, arrows) begin around the point of peak extension and are characterized by coactivation of the BF and VL with high frequency (5–8 Hz) spikes in the EMG. Scatterplots between EMG and force are shown for this trial in Figure 4B and D, where forces are divided into nonspastic and spastic in the left and right columns, respectively. A relationship between the nonspastic force and EMG is seen for the VL at high cadences (R 2 = 0.41) but at no other conditions. In contrast, a relationship between spastic forces and EMG is seen not only for the BF at high cadences (R 2 = 0.52) but also for the VL and BF at low cadences (R 2 = 0.33 and 0.51, respectively). In summary, higher cadences appear to elicit patterns of alternating EMG activity between knee flexors and extensors that correspond with nonspastic forces that are not present during low-cadence cycling. However, it appears as though leg extension elicited spasticity in our model during both low and high cadences, as seen by the positive relationship between spastic forces and EMG in the BF (knee flexor), which is stretched during extension.
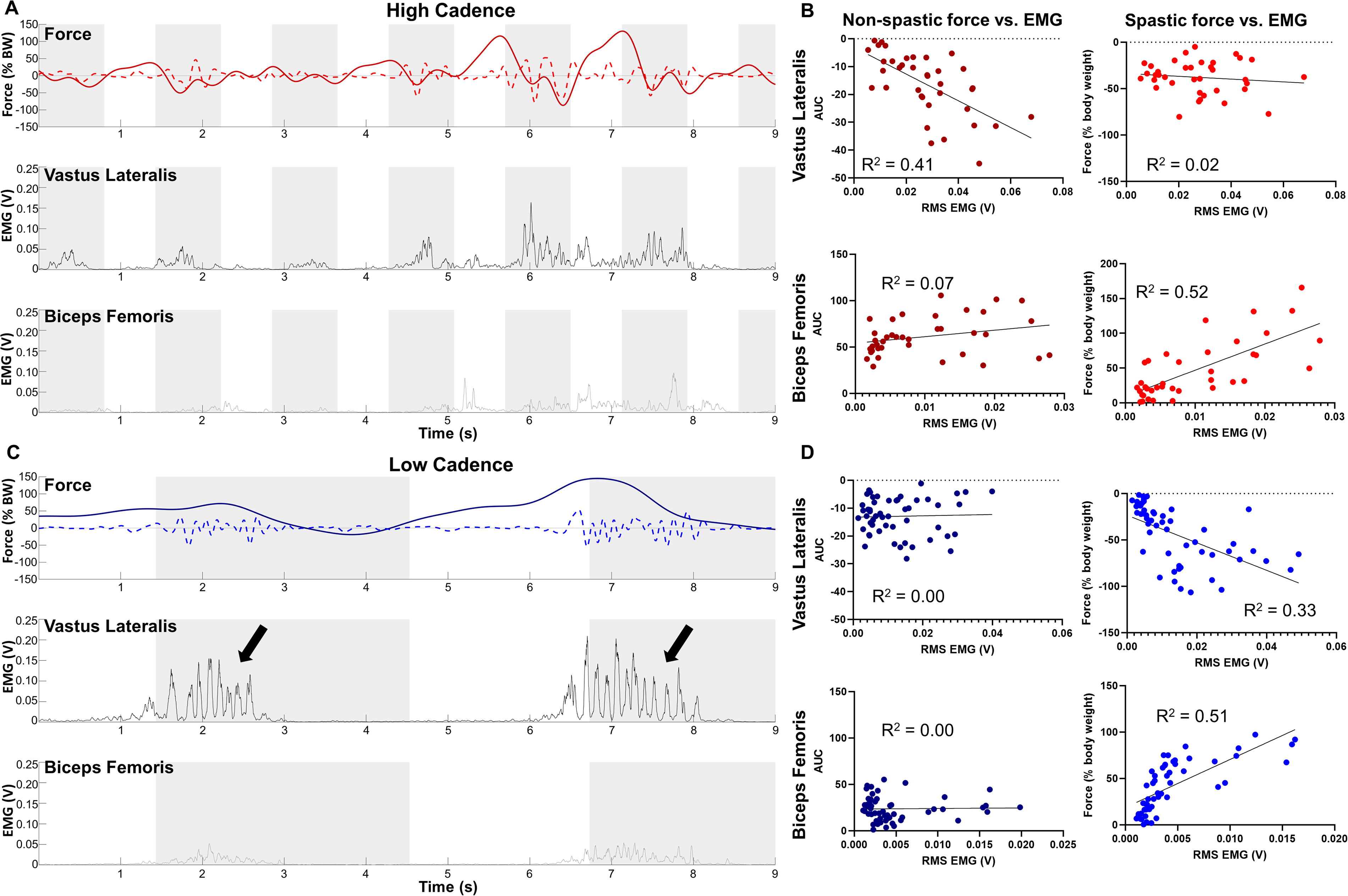
Example relationship between rhythmic force and EMG.
A quantification of the average nonspastic forces is shown in Figure 5. During the flexion phase, RMANOVA of rhythmic forces revealed an effect of GROUP (F 2,130 = 6.5, p < 0.005), CADENCE (F 1,115 = 12.3, p < 0.001), and WEEK (F 4,56 = 22.8, p < 0.001) but not DAY (Monday vs. Friday). Similarly, an effect of GROUP (F 2,67 = 5.6, p < 0.001), CADENCE (F 1,61 = 9.0, p < 0.01), and WEEK (F 4,40 = 33.9, p < 0.001) but not DAY was found during extension. Thus, overall positive and negative pedal reaction forces due to nonspastic responses increased over time postinjury, and this change was more pronounced for contused animals and more variable for contused animals with indwelling transmitters.
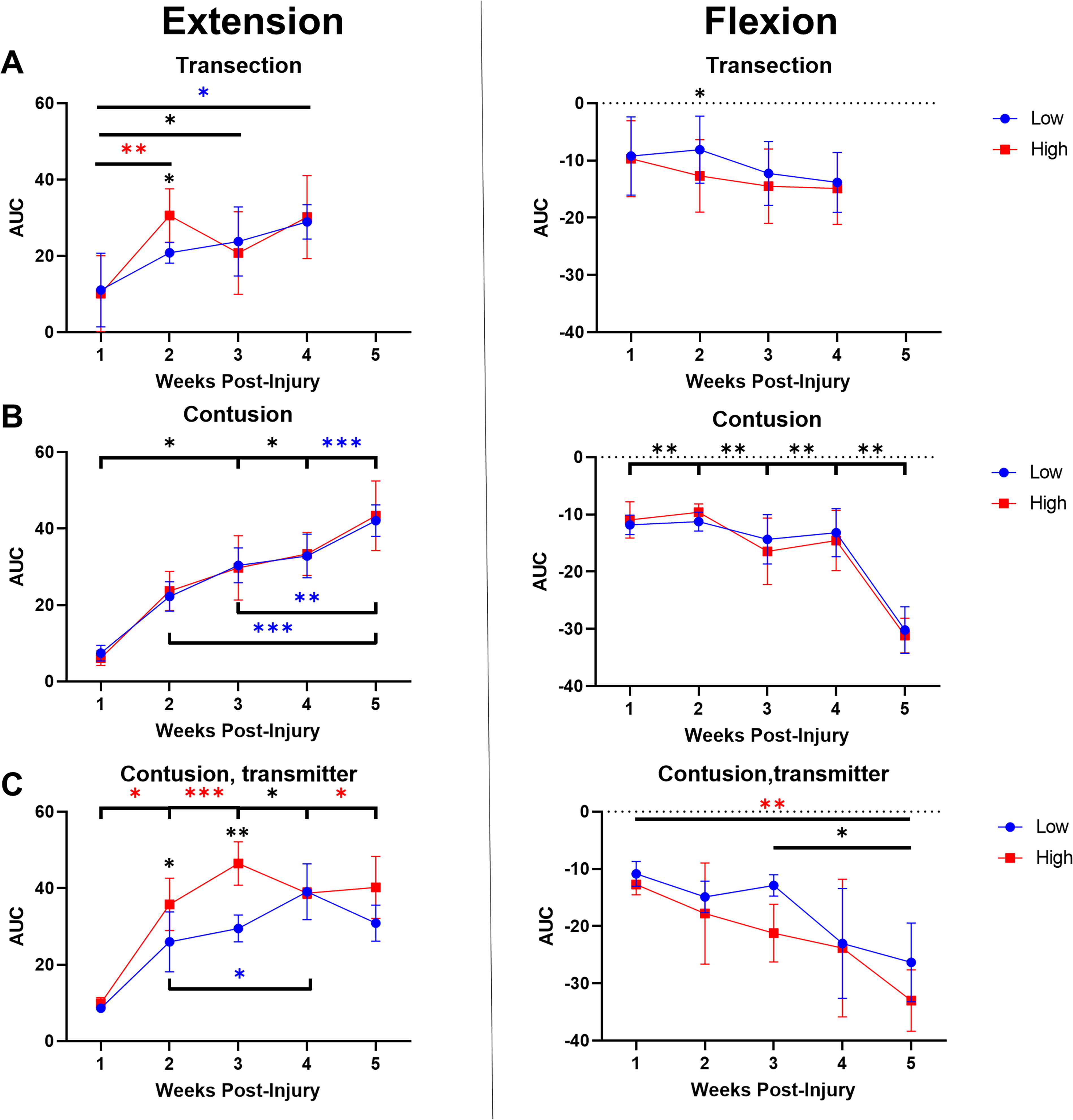
Average nonspastic forces (normalized AUC) for cycling groups.
Spastic forces are represented as heatmaps over the course of a gait cycle in Figure 6, where Figure 6A represents the percentages of trials with a spasm present, while Figure 6B represents the number of spasms per minute. A quantification of spasticity occurrence is shown in Figure 7, where cadences were divided into low and high groups. RMANOVA revealed significant effects of group (F 2,62 = 83.9, p < 0.001), cadence (F 1,97 = 121.8, p < 0.001), and week (F 4,38 = 46.8, p < 0.001) but not day. Several significant interactions also occurred: group-cadence (F 2,77 = 6.5, p < 0.01), group-week (F 7,43 = 2.7, p < 0.05), group-day (F 2,95 = 3.8, p < 0.05), week-day (F 3,39 = 5.1, p < 0.05), and group-cadence-week (F 7,48 = 3.3, p < 0.01). All groups showed an effect of cadence, as trials at low cadences were significantly more likely to contain a spastic event. The spasticity mirrors the locomotor recovery in all groups, particularly at week 1, where animals with EMG transmitters had more trials with spasms than transection or contusion cycling groups. Overall, the contusion and contusion/transmitter groups exhibited more spastic activity than the transection group, although spasticity was present in all groups. While more low-cadence cycling trials contained spastic events, it should be noted that high-cadence cycling contained more spastic events when normalized to time.
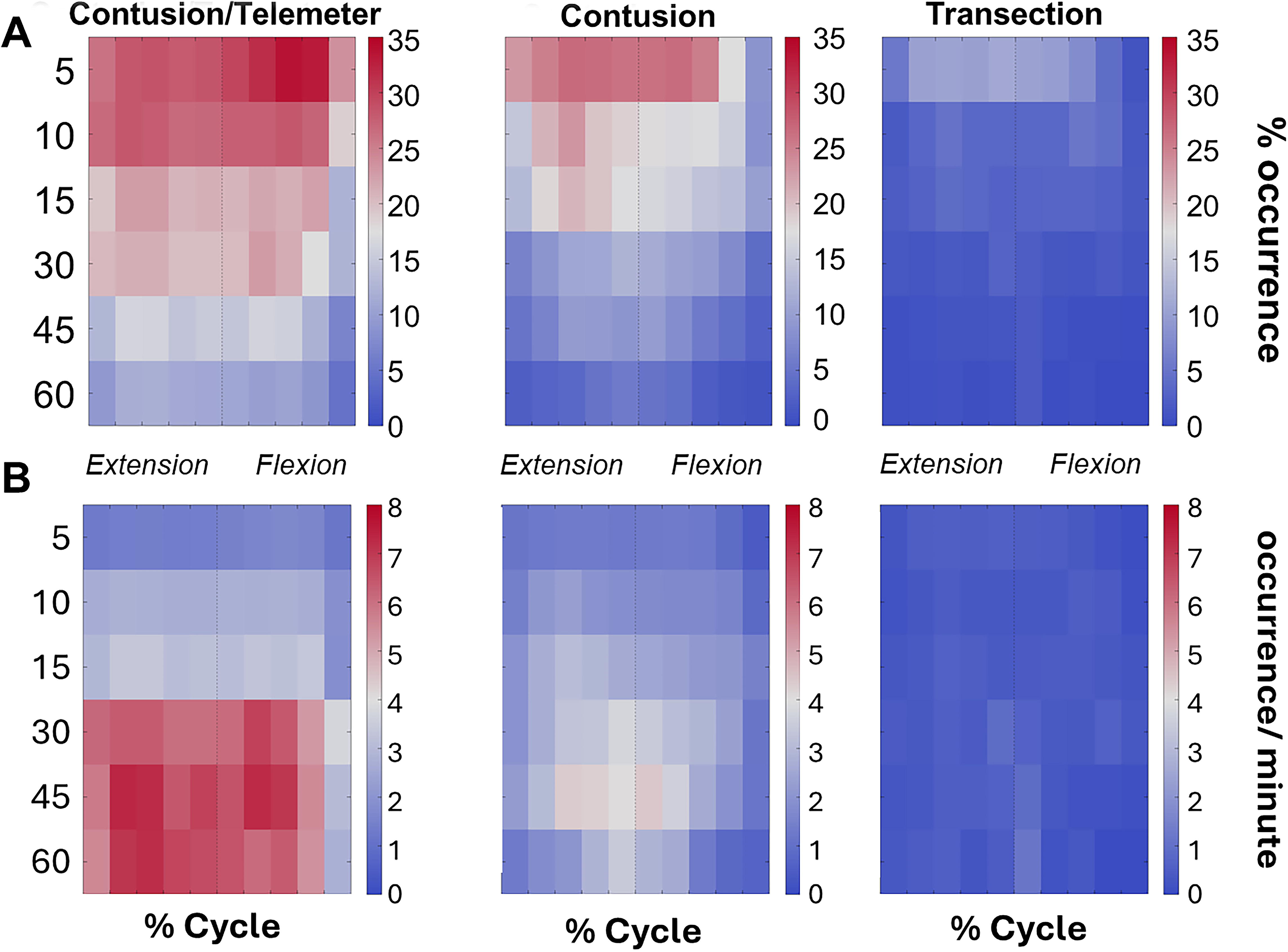
Nonrhythmic forces over the course of each cycle, pooled and represented as a heatmap.
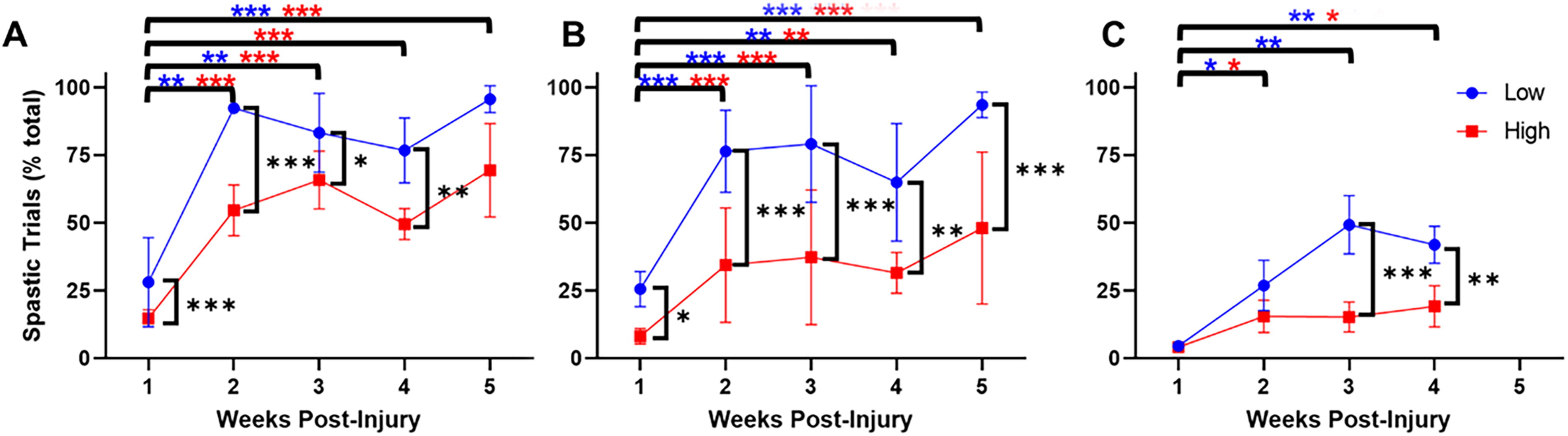
Quantification of trials with spastic forces present. Results are shown for the
Histological outcomes
The distribution of c-Fos positive neurons did not meet the criteria for a normal distribution; therefore, a Mann–Whitney U test was performed for the c-Fos data. As this analysis includes comparisons between five groups at three spinal levels, results are shown in Table 1 for ease of viewing. Significant differences between groups are represented in Figure 8 graphs at individual levels, whereas heatmaps representing the distribution of c-Fos+ neurons from L2-L4 are shown in Figure 8 (top images). The results of this analysis indicate that cycling upregulated c-Fos expression in the lumbar spinal cord in both contusion and transection groups, although slightly more neurons were found in contusion animals that received cycling. The expression of c-Fos was mostly constrained to the dorsal horn laminae I–V, with detectable numbers present in the most dorsal and medial intermediate gray.
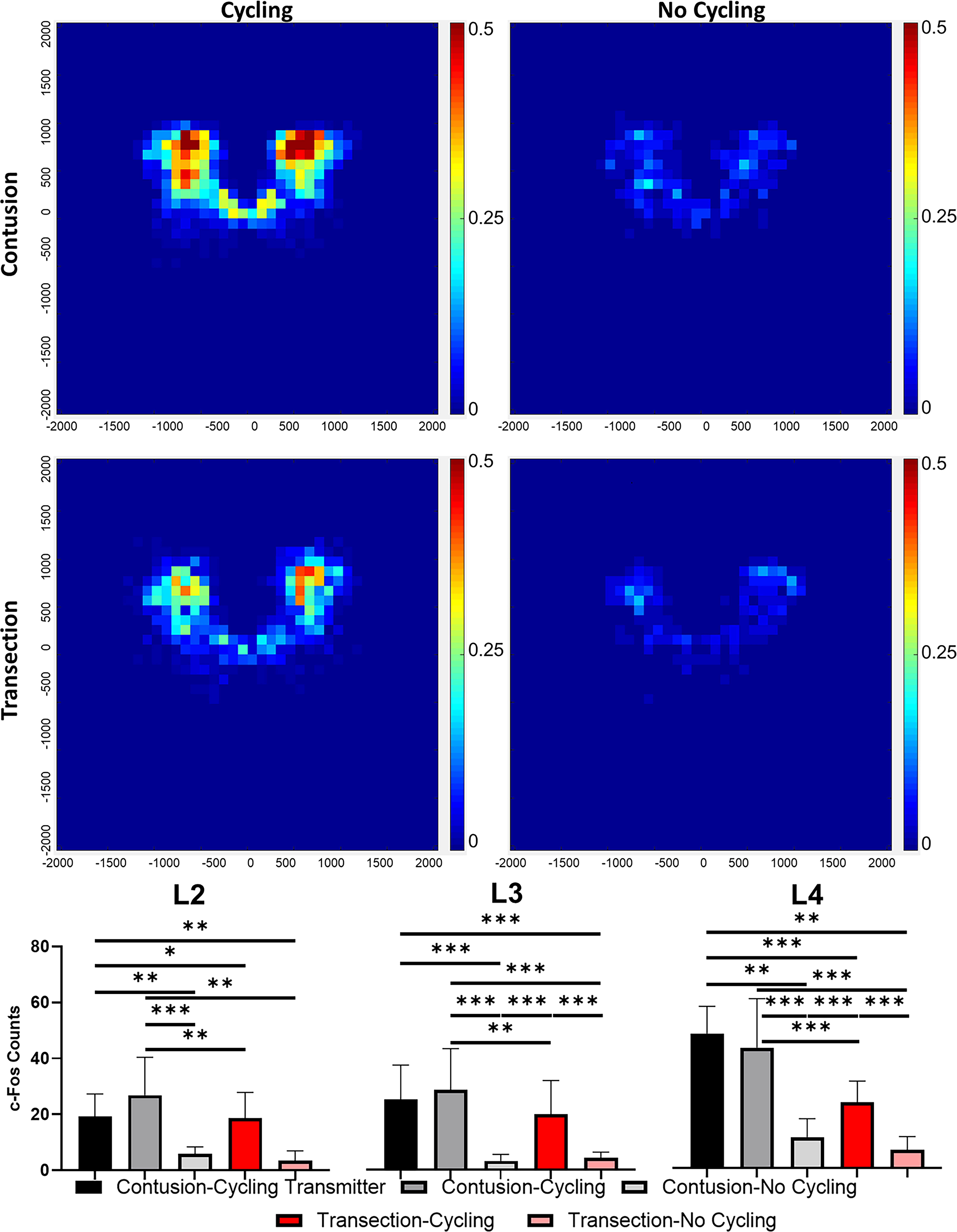
c-Fos expression in the lumbar spinal cord. Top images show heatmap visualizations of c-Fos distribution, L2-L4 pooled. No significant differences were found between contusion/cycling groups, which were pooled for visualization. Bottom graphs show Quantifications of neuron counts divided into spinal level, where each bar represents an average number of c-Fos positive neurons from 5 sections at each level. Data are displayed as group mean ± SD. All data were analyzed using a Mann–Whitney U test, *p < 0.05, **p < 0.01, ***p < 0.001. SD, standard deviation.
Mann–Whitney U Values and Their Associated p-Values
CC, contusion-cycling; CCT, contusion-cycling transmitter; CNC, contusion-no cycling; TC, transection-cycling; TNC, transection-no cycling.
Locomotor function following MC training
Figure 9 shows BBB scores acquired over the course of the cycling evaluation. A repeated-measures ANOVA revealed an effect of GROUP (F 4,117 = 80.2, p < 0.001), DAY (F 9,27 = 26.8, p < 0.001), and in the GROUP × DAY interaction (F 32,33 = 2.0, p < 0.05). Bonferroni post hoc t tests revealed that most of the differences were between contusion and transection groups, as expected. However, the CT-TR group consistently had higher BBB scores than all other groups, which were significantly higher than the CT-CY group at week 1M/F and week 5F. It is worthwhile noting that none of the groups achieved any kind of stepping but were restricted to sweeping and/or movements about 1 or 2 hindlimb joints without plantar paw placement.
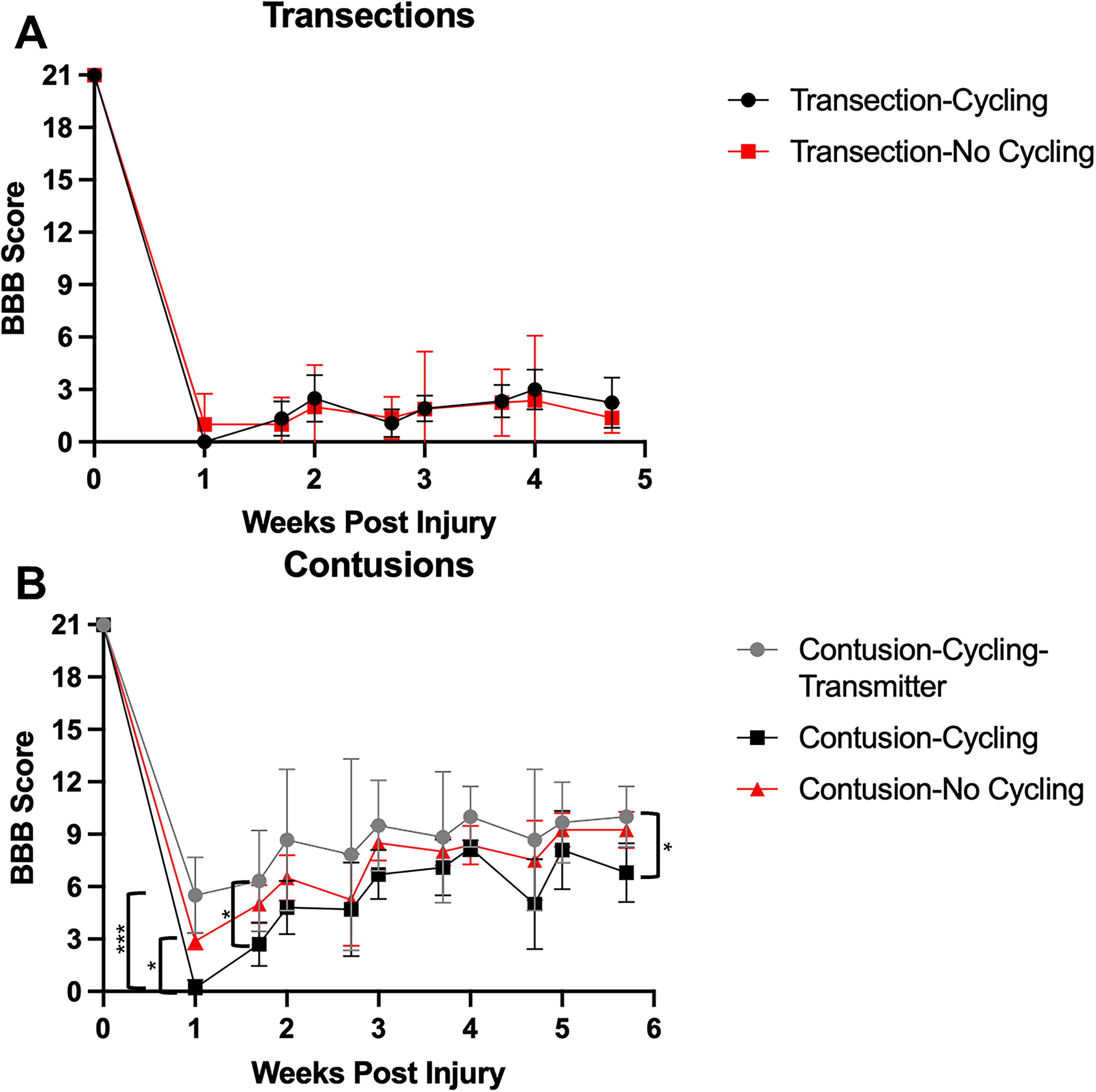
BBB scores during cycling training.
Sensory function following MC training
Sensory function (thermal hyperalgesia) evaluated by tail-flick latency is shown in Figure 10. A repeated-measures ANOVA revealed an effect of GROUP (F 4,89 = 7.7, p < 0.001), DAY (F 5,27 = 3.5, p < 0.05), and in the GROUP × DAY interaction (F 18,34 = 1.9, p < 0.05). Bonferroni post hoc t tests showed the cycling. Additionally, post hoc testing showed that response latency decreases at 2, 3, and 4 weeks postinjury in the TX-NC group compared with baseline. Thus, both contusion and transection groups showed a latency decrease postinjury, suggesting nociceptor hypersensitivity that was not measurably influenced by cycling. Interestingly, the contusion cycling transmitter group did not show hypersensitivity.
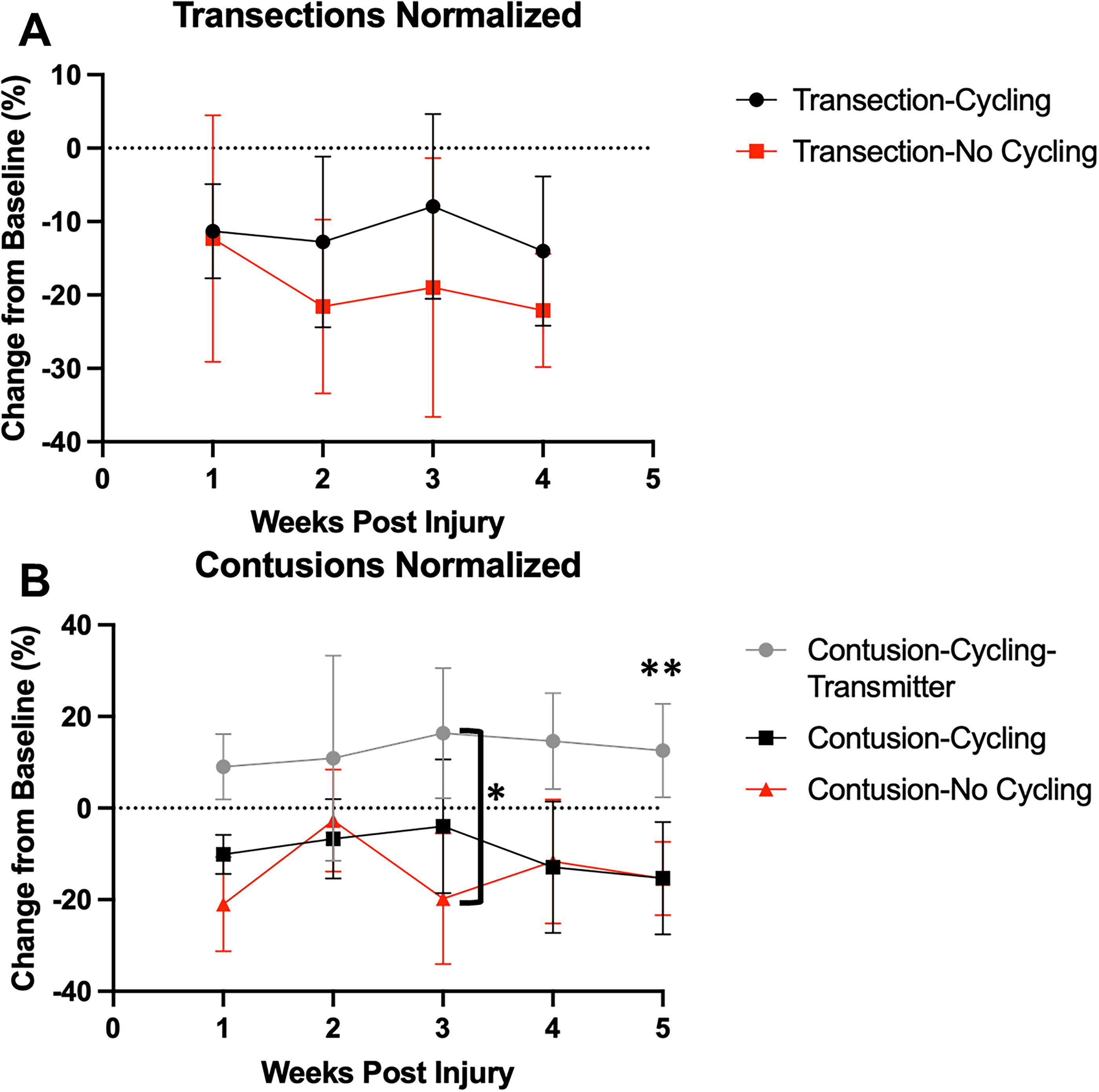
Tail-flick latency during cycling training, normalized to baseline.
Discussion
Although muscle activity has been recorded in a rat model of MC previously, the relationship between pedal reaction forces and cadence in both a transection and contusion model of SCI in rats has not yet been investigated. In addition, in the current study, we employed a technique to separate kicking/nonrhythmic forces from rhythmic forces developed with the motion of the pedals. Using this technique, we found that higher cadences (≥30 RPM) significantly increase both rhythmic pedal forces and the magnitude/duration of EMG activity, whereas lower cadences (≤15 RPM) increase the likelihood of inducing nonrhythmic forces, which contain hallmarks of spasticity—specifically clonic frequency components in the 5–8 Hz range 32 —in EMG and force recordings. Although pedal forces were higher in contusion animals, both rhythmic and nonrhythmic forces were present in transected animals. We propose that pedal reaction forces and muscle activity may play a significant role in outcome measures following MC training in rats.
Muscle activity during MC
SCI causes a disruption of supraspinal pathways projecting onto locomotor centers, which causes locomotor control to disproportionately rely on sensory feedback. 33 –35 Although the movements are not volitional, MC provides sensory information to the spinal cord (via sources such as cutaneous/ligament mechanoreceptors, muscle spindles, and Golgi tendon organs), which can lead to reflexive muscle activation. Evidence from our study suggests that muscle activity can be induced during MC in rats and is modulated strongly by the cycling cadence. Overall, the analysis of individual trials demonstrated that higher cadences were more likely to induce longer burst durations and stronger contractions. Of the trials where EMG was higher during low cadences, the majority of these (6/7) were during extension, where a higher proportion of nonrhythmic forces are seen at lower cadences. Interestingly, muscle activity in BF and VL is increased at higher cadences in both flexion and extension, although the response to flexion in the vastus is slightly higher than in the biceps. These results suggest that although the monosynaptic stretch reflex likely plays a role, it is not the only factor contributing to cycle frequency-dependent EMG activity, a concept supported by the increase in c-Fos+ nuclei throughout the dorsal horn and into the intermediate gray matter.
It is important to note that this study primarily investigated the activity of knee flexors/extensors. MC results in a complex movement involving the hip, knee, and ankle joints. Hip extension is known to play a role in activation of central pattern generators 36 and stepping during treadmill walking after T12 transection, 37 implying a key role of the hip in the control of spinal locomotor circuitry, which could be expected to continue or be even augmented after a SCI. In humans with SCI, hip oscillations have been shown to drive leg reflex activity in a velocity-dependent manner. 38 These multisegmental responses in human resemble a kicking motion we observed in our rats that was primarily induced during extension in addition to pulling or pushing to resist the motion of the pedal. Although rats are capable of hip extension, their resting position is far more flexed than humans due to their quadrupedal nature and limb posture at stance, 39 suggesting that extension of the hip may play a more pronounced role in rat MC as compared with humans.
Forces during MC
Interpretation of pedal forces in MC presents a unique challenge because the movements are induced by an external moment provided by the motor. Like clinical studies, we record a resting baseline to approximate passive joint moments. 40 However, small kinematic changes in the rats (i.e., postural, within the constraint of the sling and harness) lead to different force profiles, making it difficult to justify a simple subtraction of baseline from collected data. Our approach of computationally separating rhythmic from nonrhythmic forces offers a solution to this problem and additionally extends their interpretation in the context of reflex activation. In particular, the stretch reflex contains tonic and phasic components, defined as increased muscle tone and hyperreflexia/clonus, respectively 41,42 These components can also be thought of as static and dynamic, 43 characterized as a response to a sustained stretch of the muscle and a sudden, rapid stretch of the muscle, respectively. 44 As rhythmic forces are developed in resistance to the motion of the pedal while nonrhythmic forces are the result of quick, hyperexcitable, and high-frequency movements, a possible interpretation of this work is that they provide a representation of the tonic and phasic components of this reflex.
While high variability resulted in few differences at discrete timepoints, an overall view of the data showed significant differences between low and high cadences, with transected animals having generally lower rhythmic forces than contusion animals. Nonrhythmic forces also showed a difference between cadences; however, it appears that lower cadences increase the occurrence of these forces. Although initially puzzling, it is important to consider that the stretch reflex stimuli are being applied repetitively during cycling. Stretch reflexes are subject to depression following repetitive activation, 42 suggesting that more frequent stimulation at higher cadences could lead to different levels and types of modulation. Heatmaps show an increase in time-normalized forces above 30 RPM in all groups (Fig. 5D) despite a higher per-trial frequency at lower cadences, particularly 5 RPM (Fig. 5B). Interestingly, it has been demonstrated that monosynaptic reflex pathway depression is slightly present at a frequency of 0.1 Hz (6/min) but not 0.05 Hz (3/min), 45 which may explain why 5 RPM cycling elicits a proportionately higher number of trials containing nonrhythmic forces.
Cycle training effects on locomotor and sensory ability
Although sensory information has been shown to affect spinal learning and locomotor output, the effects of passive cycling on locomotion have not been described, as most previous studies used a transection model of SCI. Here, we performed MC using a moderately severe contusion model for the first time and described the effect of MC on locomotion. BBB testing revealed that cycle training had essentially no effect on locomotor performance, although noncycled rats had a significantly higher BBB score at week 5/Friday compared with cycle-trained rats (nontransmitter). A sawtooth pattern emerges starting at week 4 in the contusion (nontransmitter) animals, where the BBB of cycled animals is lower on Friday following 4 days of cycling. This pattern is reminiscent of locomotor deficits caused by daily hindlimb stretching, 18,46 although the effect is much more robust following stretching. However, it has been demonstrated that animals without functional nociceptors have a milder decrease in BBB scores, 17 with magnitudes comparable to the mild deficits reported in this study after cycling. This study also reported an increase in c-Fos-positive neurons in the lumbar spinal cord for stretched rats, which we also observed in the cycling group. Interestingly, the c-Fos expression after stretching in the nociceptor-depleted animals was mostly constrained to the dorsal horn, which was also true for our cycled rats. Taken together, these results suggest the responses we see during cycling are likely not nociceptive in nature.
Finally, it should be noted that animals with transmitter implants were analyzed separately from the other contusion. Although this group had marginally higher BBB scores, we were most surprised to find the injury appeared to have no effect on tail-flick latency, even showing a mild increase post-SCI. We speculate that the indwelling transmitter resulted in a system-wide inflammatory response that potentially altered the immune activation to the subsequent spinal trauma. A peripheral injury or surgical intervention leads to a systemic inflammatory response that can skew the balance of pro- and anti-inflammatory cytokines and alter immune cell function. 47 This effect has been primarily studied in regard to secondary complications post-trauma such as sepsis, 48 and the effect of a prior surgical intervention on SCI is not well characterized. However, it is well established that SCI triggers an inflammatory immune response, which results in secondary damage, 49 and there is evidence to suggest that the immune response may play a role in neuropathic pain. 50 Therefore, it is possible that our surgical intervention prior to experimental SCI had an unintended effect on the injury itself by changing the systemic immune response in this group.
Functional significance
Although MC may bring about improvements in rats post-SCI such as increased myofiber cross-sectional area 12 or improved cardiovascular function, 8 the effectiveness and clinical translatability of these results remain unclear. It has been demonstrated that physiological responses are only elicited in human cycling when paired with muscle activity and not during passive movements. 4,51 Here, we demonstrate that EMG can be elicited in response to MC in a rat and that rhythmic and nonrhythmic forces are modulated in a cadence-dependent manner in both transection and contusion models of SCI. It may be difficult to directly translate these results as humans display reduced spinal excitability post-SCI compared with rodents as proprioceptive interneurons depend more on descending input. 52 However, various technologies currently exist that raise excitability in the spinal cord 53,54 that could be paired with MC in patients that lack the strength for traditional locomotor training. Critically ill but spinally intact patients have shown increased peripheral muscle strength in response to MC. 55 About 20% of these patients showed signs of active muscle contractions during cycling, suggesting this may be an effective therapy if spinal excitability can be raised. Finally, observations from this study may be able to translate into active cycling post-SCI. Although functional electrical stimulation (FES) cycling may provide benefits similar to rodent MC such as increased muscle mass 56 and cardiovascular benefits, 57 it quickly subjects patients to fatigue. Many/most of the muscle contractions in rats are eccentric due to the activation of the stretch reflex. Although intense eccentric exercise has been shown to cause muscle damage, 58 mild eccentric exercise can increase muscle strength while putting less stress on the cardiovascular system. 59 Therefore, a milder FES paradigm paired with MC to resist the motion of the pedals by inducing consistent and modest eccentric muscle contractions may be an alternative to traditional FES cycling in some patients.
Transparency, Rigor, and Reproducibility Summary
The goal of this study was to characterize EMG and pedal force in a rat model of SCI and determine the effects of daily training on locomotor and sensory outcomes. As a mix of injury models was used, uneven group sizes were chosen based on expected variability in each injury model, and a larger group was used in the contusion/training group to account for additional variability due to training. Although the telemetry group was intended to be included in the contusion/training group, post hoc analysis revealed group differences, and therefore, they were analyzed separately. One animal in the contusion/cycling group with a telemetry implant was removed from analysis due to high locomotor performance (BBB of 20 at week 2), which precluded it from participating in MC; otherwise, all animals completed the study. Outcome measures were assessed by individuals blinded to group conditions. Additionally, strict post hoc t tests (Bonferroni t test for multiple comparisons) were used.
Footnotes
Acknowledgments
The authors would like to thank the core staff of the Kentucky Spinal Cord Injury Research Center for their assistance with surgical procedures (Christine Armstrong) and microscopy (Jason Beare) and the faculty and staff of the Comparative Medicine Research Unit at the University of Louisville for their assistance with animal care.
Authors’ Contributions
Conceptualization: G.J.R.S. and D.S.K.M.; methodology: G.J.R.S.; software: G.J.R.S. and T.C.; validation: G.J.R.S.; formal analysis: D.A.B. and G.J.R.S.; investigation: G.J.R.S. and T.C.; resources: A.S.-S.; data curation: G.J.R.S. and T.C.; writing—original draft: G.J.R.S.; writing—review and editing: G.J.R.S. and D.S.K.M.; visualization: G.J.R.S.; supervision: D.S.K.M.; project administration: G.J.R.S. and A.S.-S.; funding acquisition: D.S.K.M.
Author Disclosure Statement
The authors have no competing interest to disclose.
Funding Information
This work was funded by a grant from the Craig H. Neilsen Foundation to D.S.K.M.