
Abstract
A major portion of spinal cord injury (SCI) cases occur in the cervical region, where essential components of the respiratory neural circuitry are located. Phrenic motor neurons (PhMNs) housed at cervical spinal cord level C3–C5 directly innervate the diaphragm, and SCI-induced damage to these cells severely impairs respiratory function. In this study, we tested a biomaterial-based approach aimed at preserving this critical phrenic motor circuitry after cervical SCI by locally delivering hepatocyte growth factor (HGF). HGF is a potent mitogen that promotes survival, proliferation, migration, repair, and regeneration of a number of different cell and tissue types in response to injury. We developed a hydrogel-based HGF delivery system that can be injected into the intrathecal space for local delivery of high levels of HGF without damaging the spinal cord. Implantation of HGF hydrogel after unilateral C5 contusion-type SCI in rats preserved diaphragm function, as assessed by in vivo recordings of both compound muscle action potentials and inspiratory electromyography amplitudes. HGF hydrogel also preserved PhMN innervation of the diaphragm, as assessed by both retrograde PhMN tracing and detailed neuromuscular junction morphological analysis. Furthermore, HGF hydrogel significantly decreased lesion size and degeneration of cervical motor neuron cell bodies, as well as reduced levels surrounding the injury site of scar-associated chondroitin sulfate proteoglycan molecules that limit axon growth capacity. Our findings demonstrate that local biomaterial-based delivery of HGF hydrogel to injured cervical spinal cord is an effective strategy for preserving respiratory circuitry and diaphragm function.
Introduction
Traumatic spinal cord injury (SCI) occurs when an external force, such as a sudden physical impact or vertebral compression, damages the spinal cord. 1,2 Injury severity depends on a number of factors, including the anatomical level of the spinal cord affected and the degree of damage incurred. Injuries to the cervical spinal cord represent the majority of all human SCI cases 3,4 and, unfortunately, tend to result in the most severe outcomes. 5 Given that a major portion of ascending and descending circuits pass through the cervical spinal cord, it is not surprising that damage here results in a wide range of sensorimotor and autonomic deficits, including impairments in respiratory function. 5 –7 Pulmonary compromise is a leading cause of morbidity and mortality after SCI 8,9 and is characterized by reductions in lung and chest wall compliance, decreased vital capacity, and an impaired cough reflex. 9,10 As a result, patients are at increased risk of developing secondary respiratory complications such as atelectasis and pneumonia, which may themselves become life-threatening. Furthermore, many become reliant on mechanical ventilation, decreasing life expectancy and overall quality of life, 10 –12 highlighting the importance of managing respiratory dysfunction.
SCI pathophysiology is complex but can be broadly divided into primary and secondary injury processes. The primary injury occurs at the moment of trauma and involves mechanical disruption of the spinal cord tissue. 2 The secondary injury immediately follows and exacerbates and enlarges the initial injury over a period of hours to months. 2 Secondary damage is propagated through both physical and biochemical events that lead to inflammation, excitotoxicity, oxidative stress, apoptosis, changes in vascular permeability, and scar formation. 13,14 These processes inflict damage to critical neural circuits that control the muscles of respiration after cervical SCI. 9,15 Paresis or paralysis of the diaphragm, the principal muscle of inspiration, is a major contributor to respiratory impairment. 7 The diaphragm is innervated by phrenic motor neurons (PhMNs) spanning cervical levels 3–5 (C3–C5) in the ventral horn of the spinal cord. 16,17 Contraction of the diaphragm is driven by PhMN activation, which is, in turn, activated by descending excitatory input from a nucleus of pre-motor bulbospinal neurons originating from the rostral part of the ventral respiratory group (rVRG) in the medulla. 16 –20 Cervical SCI damages this important rVRG-PhMN-diaphragm circuit and impairs respiratory function through two main mechanisms: (1) loss of PhMNs, which results in diaphragm neuromuscular junction (NMJ) denervation; and (2) loss of monosynaptic and/or polysynaptic excitatory input from severed descending rVRG axons, which effectively silences spared PhMNs. Therefore, therapies that target either of these processes have the potential to improve respiratory function after SCI.
Hepatocyte growth factor (HGF) has emerged as a promising therapeutic approach for the treatment of SCI. HGF is a potent mitogen that promotes survival, proliferation, migration, repair, and regeneration of many different cell and tissue types in response to injury. 21,22 The diverse actions of HGF are mediated through the binding and activation of its only known receptor, c-MET. 23 –25 c-MET activation initiates downstream signaling pathways (e.g., Akt, Ras, and STAT3) that exert a variety of biological effects. 26,27 In several organs (including the liver, lungs, and kidneys), rapid increases in HGF–c-MET activity are observed within 1 day of injury, which were found to be mediated by large increases in HGF derived from both local and distant (endocrine) sources. 28 –32 In contrast, the spinal cord mounts a significantly reduced response to injury. Although increases in HGF levels do occur, they are severely delayed, taking about 14 days to peak. 33 On the contrary, sustained increases in c-MET expression occur within the first 24 h, 33 suggesting that endogenous upregulation of HGF in response to SCI is inadequate and that providing an exogenous source of HGF in the early stages after injury may enhance tissue repair.
HGF has shown promising effects in both SCI and amyotrophic lateral sclerosis (ALS), offering potential therapeutic paths for these conditions. In SCI, HGF administration has demonstrated regenerative and neuroprotective effects. It has been shown to promote neuronal survival, increase angiogenesis, reduce inflammation, and improve functional recovery in several preclinical models. 33 –35 HGF has also been found to inhibit scar formation, 36 promoting a more conducive environment for neural regeneration and repair. Similarly, in ALS, HGF exhibits neuroprotective properties by preventing motor neuron death, reducing oxidative stress, protecting against excitotoxicity, and slowing disease progression. 37 –39 Importantly, however, the specific effects of HGF on respiratory circuitry have yet to be evaluated despite the critical importance of this circuitry in these disease conditions. Furthermore, although treatments using HGF have progressed to clinical trials for both SCI 40 and ALS, 41,42 administration routes for optimal drug dosing and efficacy have yet to be determined. Therefore, the central objectives of this study are to assess whether HGF has neuroprotective and/or regenerative effects on critically important respiratory circuitry and to optimize its delivery to the injured spinal cord.
Hydrogels, three-dimensional networks of hydrophilic polymers, have gained considerable interest as drug delivery vehicles due to their unique properties. These materials possess high water content, biocompatibility, 43 tunable mechanical properties, 44,45 and the ability to encapsulate and deliver bioactive molecules, 46 making them an attractive vehicle to administer HGF. Furthermore, hydrogel-based delivery systems permit the local delivery of biologics directly to the injured spinal cord, bypassing the blood–spinal cord barrier and avoiding the deleterious side effects associated with systemic delivery. 47,48 Here, we developed a novel biomaterial-based method to safely and locally deliver HGF to the injured cervical spinal cord to preserve respiratory function. We demonstrate that HGF hydrogel exerts a neuroprotective effect on respiratory circuitry after cervical SCI, preserving diaphragm innervation and function, decreasing motor neuron degeneration, and reducing the levels of inhibitory scar-associated chondroitin sulfate proteoglycan (CSPG) molecules.
Materials and Methods
Animals
Female Sprague–Dawley rats (250–300 g; Taconic Biosciences) were housed under conditions controlled for appropriate humidity, temperature, and light, and rats were given proper access to water and food ad libitum. Experiments were conducted in compliance with the Animal Research: Reporting of In Vivo Experiments guidelines and the National Institutes of Health’s guide for the care and use of laboratory animals, with approval by the Thomas Jefferson University Institutional Animal Care and Use Committee (IACUC) and the Drexel University IACUC. Rats were randomized into three groups: laminectomy-only, contusion plus blank hydrogel, and contusion plus HGF hydrogel.
Unilateral cervical contusion
The model of unilateral mid-cervical hemi-contusion SCI in the rat produces robust loss of PhMNs in the cervical spinal cord, diaphragm denervation, and altered physiological activity of the diaphragm. 49,50 Rats were anesthetized with an intraperitoneal injection of a cocktail of ketamine (100 mg/kg), xylazine (5 mg/kg), and acepromazine (2 mg/kg), and anesthesia was monitored by orbital and toe pinch reflexes. A hemi-laminectomy was performed on the right side of the spinal cord to expose C4/5 spinal cord following an incision of the cervical dorsal skin and muscles. A unilateral contusion was conducted with a 1.5 mm tip using the Infinite Horizon spinal impactor (Precision Systems and Instrumentation) at a force of 395 kDynes. The rationale for the location of this injury is that it targets damage to a significant portion of the PhMN pool, allowing us to assess effects on neuroprotection and consequent functional innervation of the diaphragm. Importantly, we performed both functional electromyography (EMG) and morphological NMJ innervation analyses separately at various subregions of the hemi-diaphragm, which allowed us to correlate the location and degree of damage/protection in the spinal cord with outcome measures at the corresponding portion of the muscle. As a control for the contusion injury, we chose to use laminectomy-only and not a fully spinal intact condition with no surgery. As the laminectomy procedure can impact diaphragm motor output, this is not a completely uninjured control condition.
HGF hydrogel preparation and implantation
Hydrogels were prepared and implanted as previously described. 51 A solution containing 4 mg/mL dextran sulfate (DS; Sigma-Aldrich) and 0.8 mg/mL HGF (PeproTech) was prepared in deionized water. A separate solution containing 4 mg/mL chitosan (CH) was prepared in 0.6% (v/v) acetic acid. Equal volumes (10 μL) of each solution were mixed together and allowed to self-assemble into DS-HGF-CH complexes. Next, the mixture was centrifuged at 10,000 RPM for 10 min to remove the supernatant. DS-HGF-CH complexes were then loaded into 20 μL of a 1.5% (w/v) agarose hydrogel for implantation as follows. Immediately following contusion SCI, 20 μL of either HGF hydrogel or blank (control) hydrogel was injected intrathecally at the injury site. To protect the injury site, an additional 100 μL of blank hydrogel was applied epidurally.
Characterization of HGF release
HGF released from the hydrogel was measured as follows. Rhodamine was conjugated to HGF using the Pierce Rhodamine Protein Labeling Kit (Thermo Fisher Scientific) according to the manufacturer’s instructions. Next, equal amounts of rhodamine-HGF and HGF were used to recreate the drug delivery system in vitro. This agarose hydrogel loaded with DS-HGF-CH complexes was incubated in Hank’s Balanced Salt Solution pH 7.4 at 37°C over a period of 23 days. The release medium was changed every 24 h, and the amount of HGF released was quantified via fluorescence readout (excitation 552 nm, emission 575 nm).
Diaphragm compound muscle action potentials
We used isoflurane (2.0–2.5% diluted in oxygen; Piramal Healthcare) to anesthetize rats for compound muscle action potential (CMAP) recording. Stimulating needle electrodes (positive and negative) spaced 0.5 cm apart were inserted transcutaneously close to the passage of the phrenic nerve either ipsilateral or contralateral to the injury. 52,53 A ground needle electrode was placed subcutaneously into the tail, and a reference electrode was inserted subcutaneously into the abdominal region. A surface strip recording electrode was placed along the costal margin of the diaphragm. External stimulation (0.5 ms duration; 6 mV amplitude) with 5 sec intervals was presented, and 10–20 CMAP recordings were obtained. ADI Powerlab 8/30 stimulator and BioAMP amplifier (ADInstruments) were used for recordings, and Scope 3.5.6 (ADInstruments) was used to analyze CMAP amplitude from baseline to peak. CMAPs were measured in each animal at both 14 and 28 days following SCI. We did not perform CMAP recording on laminectomy-only rats, as we present the CMAP amplitudes from the within-animal contralateral hemi-diaphragm of all the SCI subjects.
EMG recordings
EMG recordings were obtained during a terminal procedure. Animals were anesthetized with isoflurane (2.0–2.5% oxygen). The hemi-diaphragm was exposed via laparotomy ipsilateral to the injury. By inserting bipolar electrodes (3 mm apart) into the dorsal, medial, or ventral subregions of the hemi-diaphragm muscle, inspiratory EMG bursting was recorded over 2 min in each animal to quantify peak amplitude, burst frequency, and burst duration. 54 –56 The EMG signal was amplified and filtered through a band-pass filter (50–3000 Hz) using LabChart 7 software (ADInstruments).
Retrograde labeling of PhMNs
To identify PhMNs in the cervical ventral horn, 20 μL of cholera toxin subunit B (CTB; 2.5 mg/mL; Life Technologies), a retrograde neuronal tracer, was delivered into the intrapleural space 3 days prior to sacrifice. Rats were anesthetized with isoflurane (2.0–2.5% in oxygen), and a Hamilton syringe was inserted 3 mm through the intercostal space between the sixth and seventh ribs (intersection of the anterior axillary line and xiphoid process). CTB was delivered into the intrapleural space ipsilateral to cause a contusion. Rats were removed from isoflurane and allowed to recover in home cages. Using this retrograde CTB labeling, PhMN somata and proximal dendrites in the ventral horn of the cervical spinal cord can be visualized. 49,54 We quantified CTB-labeled PhMN cell bodies in every 20 μm-thick transverse section, and we present these data as total counted CTB+ PhMNs across 1 mm-long rostral–caudal spans of cervical spinal cord.
Diaphragm and spinal cord dissection
Animals were euthanized with an overdose of ketamine (300 mg/kg), xylazine (15 mg/kg), and acepromazine (6 mg/kg) 8 weeks following SCI. A fresh diaphragm was first removed, followed immediately by perfusion. The hemi-diaphragm was exposed laterally along the rib cage and excised using spring scissors (Fine Science Tools), then stretched flat, and pinned down to Sylgard medium (Fisher Scientific), followed by washing with phosphate-buffered saline (PBS). Four percent paraformaldehyde (Electron Microscopy Sciences) was then used to fix the diaphragm for 20 min, followed by a PBS wash. After the removal of superficial fascia from the diaphragm surface, muscles were processed for NMJ labeling. Animals were perfused with a 0.9% saline solution. Spinal cord and brain were dissected and post-fixed in 4% paraformaldehyde overnight at 4°C, washed with a 0.1 M phosphate buffer solution for 24 h, and cryoprotected with 30% sucrose for 3 days. The spinal cord was embedded in tissue freezing medium and sectioned in transverse or sagittal orientation at 20 μm thickness.
Assessment of lesion volume and ventral horn motor neurons
To quantify lesion size and overall lesion volume, transverse spinal cord sections from C3–C5 spaced 160 μm apart in the rostral–caudal axis were stained for Nissl bodies and myelin using cresyl violet and eriochrome cyanine, respectively. Images were acquired with a Zeiss Imager M2 upright microscope. Using ImageJ software, the lesion area per section was measured, and these values were used to quantify the total lesion volume. 57 A lesion in the spinal cord was defined as areas including both lost tissue with cystic cavity and surrounding damaged tissue in which the normal anatomical structure of the spinal cord was lost. The number of motor neuron cell bodies in these sections was quantified manually in a blinded manner. Motor neurons with a clearly identifiable nucleolus and a cell soma greater than 200 μm2 were counted. 57 We present these motor neuron quantification data both as motor neurons per section and as total counted motor neurons across a 10 mm rostral–caudal extent of the cervical spinal cord.
NMJ analysis
Immediately after diaphragm dissection, connective tissue was removed to ensure optimum antibody penetration. The muscle was stretched and pinned down to Sylgard medium (Fisher Scientific). To label motor axons and their terminals, muscles were incubated with SMI-312R (1:1000; Covance) and SV2-s (1:10; Developmental Studies Hybridoma Bank, University of Iowa), respectively, followed by incubation with FITC anti-mouse IgG secondary (1:100; Jackson ImmunoResearch). In addition, post-synaptic nicotinic acetylcholine receptors were labeled with rhodamine-conjugated alpha-bungarotoxin (1:400; Life Technologies). Finally, the diaphragm was coverslipped with Vectashield medium (Vector Laboratories). Diaphragm imaging was performed with a FluoView FV1000 confocal microscope (Olympus), and images were analyzed in a blinded manner for the total number of intact, completely denervated, and partially denervated NMJs. 58 –61
Immunohistochemistry
Spinal cord sections were air-dried for 30 min and washed three times (5 min each) with PBS. Samples were incubated in blocking solution (5% normal horse serum, 1% Triton X-100, and 0.05% tween 20 diluted in PBS) for 1 h at room temperature. Sections were labeled for 48 h at 4°C with goat-anti-CTB (1:10,000; List Biological Laboratories) or mouse-anti-CS56 (1:250; Sigma-Aldrich) in blocking solution. Sections were then washed three times with PBS (5 min/wash) and incubated with Alexa Fluor 594 donkey-anti-mouse (1:200; Life Technologies) or Alexa Fluor 488 donkey-anti-goat (1:200; Jackson ImmunoResearch) in blocking solution for 2 h at room temperature. After PBS washing (3 washes, 5 min each), sections were coverslipped. For CTB, images were acquired with a Zeiss Imager M2 microscope. For CS56, images were acquired on a Nikon Eclipse TE2000-U microscope. CTB-labeled PhMNs in C3–C5 ventral horn were counted in 100 μm spaced transverse sections in a blinded manner. 49,54 Somal size of individual CTB-labeled PhMNs was determined using ImageJ software. ImageJ was used to quantify the area of tissue staining positive for CS56 from multiple transverse sections along a 5–6 mm rostro-caudal expanse centered about the lesion epicenter. We performed CS56 staining and imaging for all tissue sections at the same time, and we subtracted the same level of background for all images when performing the quantification of this CS56 staining.
Statistical analysis
Results were expressed as means ± standard error of the mean (SEM). An unpaired t-test was used to analyze comparisons between two groups, and an analysis of variance (one-way analysis of variance [ANOVA]) followed by a post hoc test (Bonferroni’s method) was used for multiple comparisons between three or more groups. Prism 10 (GraphPad Software) was used to calculate statistics. p-Value < 0.05 was used as a threshold for statistical significance.
Results
Extended release of HGF from the hydrogel construct
In this study, we used self-assembling DS-HGF-CH complexes incorporated into an agarose hydrogel to immobilize them at the injury site for achieving HGF release. To first validate the construct’s capacity for HGF release and to quantify both the amount and the duration of this release, we performed an in vitro release assay. Relatively high amounts of HGF are released during the first day (1.2 ± 0.04 μg), followed by a sustained period of reduced release ranging from 587.8 ± 9.1 to 2.8 ± 0.5 ng/day over the following 22 days (Fig. 1; n = 3).
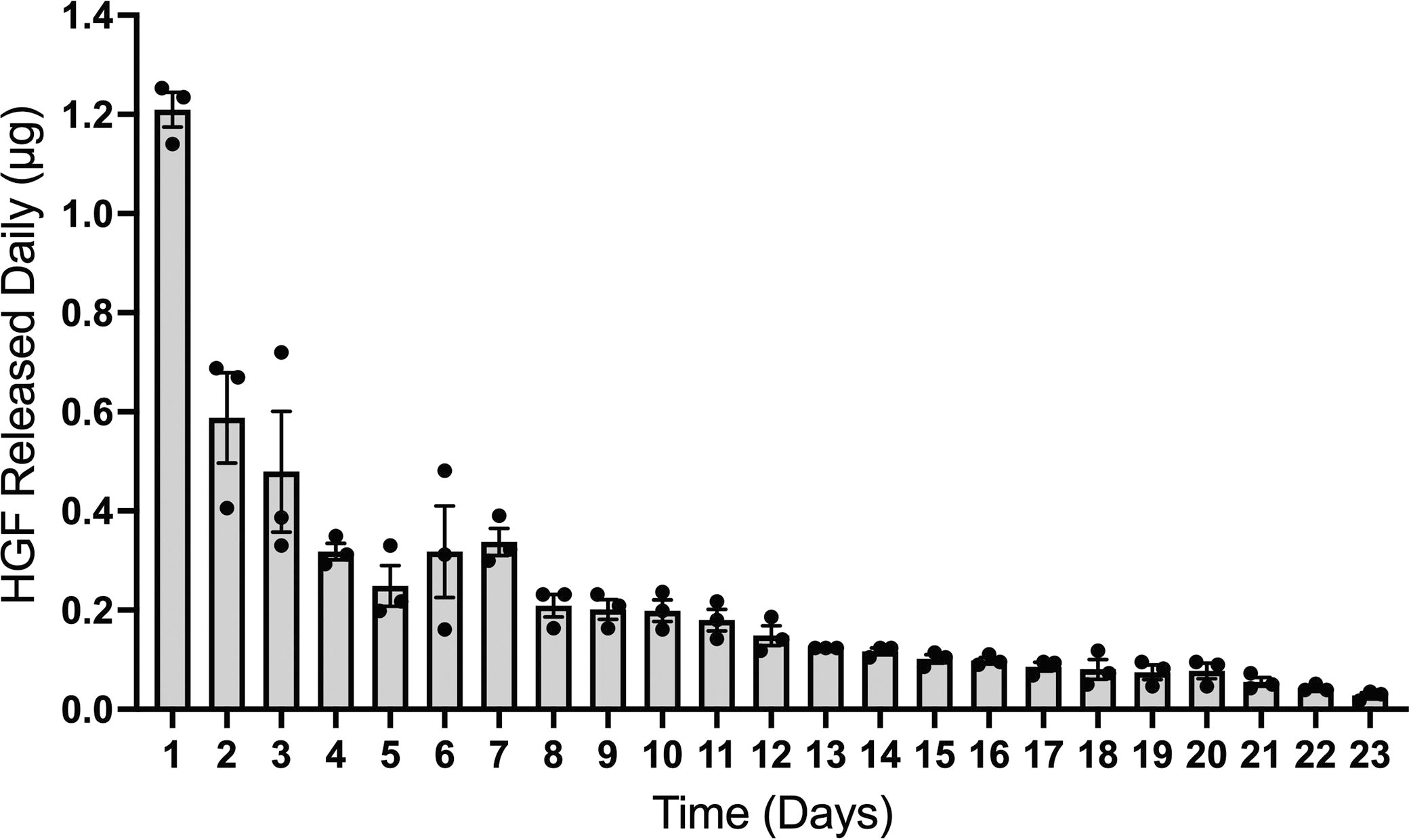
Characterization of HGF release from the hydrogel. HGF was released from an agarose hydrogel loaded with DS-HGF-CH complexes for up to 23 days. Data are presented as mean ± SEM (n = 3). CH, chitosan; DS, dextran sulfate; HGF, hepatocyte growth factor; SEM, standard error of the mean.
HGF hydrogel preserved diaphragm function following cervical contusion SCI
In a rat model of C5 hemi-contusion SCI, we delivered either HGF hydrogel or blank hydrogel control by subdural implantation at the contusion site immediately after injury. In anesthetized living animals during normal eupneic breathing, we assessed effects on diaphragm function by quantitatively measuring both inspiratory EMG burst amplitude at 56 days post-injury and evoked CMAP amplitude at both 14 and 28 days post-injury (Fig. 2A).
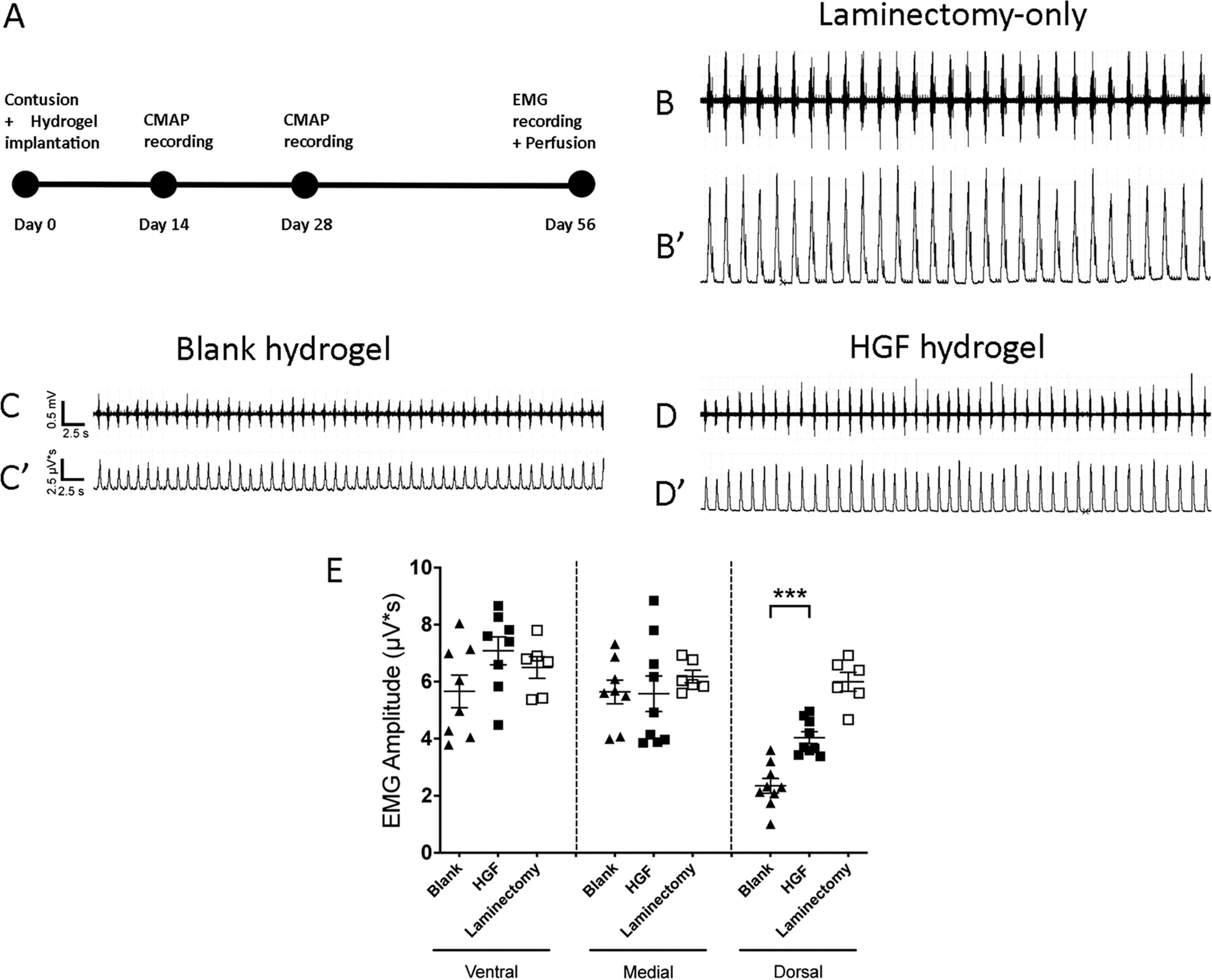
HGF hydrogel preserved diaphragm electromyography (EMG) amplitude. Timeline of experiment
We conducted intra-diaphragm EMG recordings separately from the three subregions of the hemi-diaphragm (i.e., ventral, medial, and dorsal) ipsilateral to contusion. PhMNs are located at the C3, C4, and C5 portions of the phrenic nucleus innervate ventral, medial, and dorsal muscle subregions, respectively. 62 In contusion animals implanted with blank hydrogel, we observed a significant reduction of EMG amplitude compared with laminectomy-only rats in only the dorsal subregion of the muscle (2.35 ± 0.26 μV/s for blank vs. 6.00 ± 0.33 for laminectomy-only; Fig. 2B–D), whereas both the medial and the ventral subregions showed no EMG amplitude reduction in contusion rats compared with laminectomy-only controls (traces not shown for medial and ventral recordings), which was expected given that the contusion was located at C5 and therefore did not target the C3–C4 PhMNs. In the dorsal subregion, HGF hydrogel resulted in significantly larger EMG amplitudes compared with blank hydrogel implanted control animals, though HGF hydrogel amplitude was still below laminectomy-only control (blank, 2.35 ± 0.26 μV/s; HGF, 4.04 ± 0.21) at 8 weeks post-SCI [dorsal: F(2,21) = 45.50; Tukey’s post-test; blank vs. HGF, p = 0.0002; blank vs. laminectomy, p < 0.0001; HGF vs. laminectomy, p = 0.0001; ANOVA; n = 6–8 rats/group; Fig. 2E]. We found no differences in EMG amplitudes among blank hydrogel, HGF hydrogel, and laminectomy-only rats in both the ventral (blank, 5.66 ± 0.66 μV/s; HGF, 7.08 ± 0.56; laminectomy, 6.50 ± 0.38) and the medial (blank, 5.64 ± 0.48 μV/s; HGF, 5.58 ± 0.76; laminectomy, 6.18 ± 0.22) subregions of the diaphragm [ventral: F(2,19) = 2.14; Tukey’s post-test; blank vs. HGF, p = 0.13; blank vs. laminectomy, p = 0.51; HGF vs. laminectomy, p = 0.72; medial: F(2,20) = 0.38; Tukey’s post-test; blank vs. HGF, p = 0.99; blank vs. laminectomy, p = 0.76; HGF vs. laminectomy, p = 0.70; ANOVA; n = 6–8 rats/group; Fig. 2D]. We also observed no differences in burst frequency (Table 1) or burst duration (Table 2) between the blank hydrogel and HGF hydrogel treatment groups at any subregion of the hemi-diaphragm. 56
Diaphragm EMG Burst Frequency
EMG burst frequencies, expressed in bursts per minute (BPM), recorded from the ventral, medial, or dorsal subregions of the ipsilateral hemi-diaphragm of eupneic rats. There were no differences in burst frequency across the three experimental groups in any of the diaphragm subregions, as determined by one-way analysis of variance (ANOVA) testing (n = 4–9 rats/group).
EMG, electromyography; HGF, hepatocyte growth factor; SEM, standard error of the mean.
Diaphragm EMG Burst Duration
EMG burst durations, expressed in seconds, recorded from either the ventral, medial, or dorsal subregions of the ipsilateral hemi-diaphragm of eupneic rats. There were no differences in burst duration across the three experimental groups in any of the diaphragm subregions, as determined by one-way analysis of variance (ANOVA) testing (n = 4–9 rats/group).
EMG, electromyography; HGF, hepatocyte growth factor; SEM, standard error of the mean.
We conducted CMAP recordings from the hemi-diaphragm at both 14 and 28 days after contusion/hydrogel implantation. Unlike EMG recordings, this approach allowed us to selectively quantify functional innervation of the diaphragm by PhMNs independent of the contribution of the descending bulbospinal drive, as we measured the CMAP response to supramaximal phrenic nerve stimulation. We recorded CMAP amplitude from the entire hemi-diaphragm using a surface electrode, unlike with the EMG recordings that allowed us to assess each muscle subregion individually. We performed CMAP recordings separately on the ipsilateral and contralateral hemi-diaphragm (Fig. 3A–C). We found significantly and persistently larger CMAP amplitudes in the ipsilateral hemi-diaphragm of the HGF hydrogel group (week 2, 5.51 ± 0.14 mV; week 4, 6.37 ± 0.44) compared with blank hydrogel control (week 2, 3.30 ± 0.17 mV; week 4, 4.33 ± 0.27) at 14 and 28 days post-injury [week 2: F(3, 30) = 57.01, Tukey’s post-test; blank vs. HGF, p < 0.0001; week 4: F(3, 30) = 15.64, Tukey’s post-test, blank vs. HGF, p = 0.0120; ANOVA; n = 8–9 rats/group; Fig. 3D]. Conversely, the contralateral CMAP amplitudes were not different between these two groups at either time point (blank: week 2, 6.80 ± 0.30 mV; week 4, 7.25 ± 0.47; HGF: week 2, 7.64 ± 0.33; week 4, 8.33 ± 0.54; Fig. 3D). In addition, while HGF significantly preserved CMAP amplitudes in the ipsilateral hemi-diaphragm compared with blank hydrogel control, ipsilateral amplitudes for the HGF animals (week 2, 5.51 ± 0.14 mV; week 4, 6.37 ± 0.44) were still significantly below contralateral hemi-diaphragm levels of the same animals (week 2, 7.64 ± 0.33; 8.33 ± 0.5) at both 2 [F(3, 30) = 57.01, Tukey’s post-test; p < 0.0001; ANOVA; n = 8–9 rats/group] and 4 weeks [F(3, 30) = 15.64, Tukey’s post-test; p = 0.0193; ANOVA, n = 8–9 rats/group], demonstrating a partial therapeutic effect of HGF (Fig. 3D).

HGF hydrogel preserved compound muscle action potential (CMAP) amplitude. CMAPs were recorded in both the ipsilateral
HGF hydrogel preserved PhMN innervation of the diaphragm
To quantify the numbers of PhMNs in the ipsilesional spinal cord innervating the ipsilateral hemi-diaphragm, we selectively labeled PhMN cell bodies by intrapleurally injecting the retrograde tracer, CTB. We performed these CTB injections 3 days prior to sacrificing rats at 8 weeks post-contusion. In transverse sections of the cervical spinal cord, CTB-labeled PhMNs in the ipsilateral ventral horn extended from approximately C3 to C5/6 (Fig. 4A–B). We quantified CTB-labeled PhMN numbers at specific distances relative to the epicenter of the lesion site. We analyzed multiple transverse sections at each distance, as the number of CTB-labeled PhMNs in the ventral horn of a single section is small given that the phrenic nucleus is arranged in the rostral–caudal axis of the spinal cord as a long thin column. Specifically, the lesion epicenter was defined as the zero distance, and four separate distance bins across the PhMN pool were examined (0–1000, 1000–2000, 2000–3000, and 3000–4000 μm, all measured rostral to the epicenter). Compared with blank hydrogel control rats, HGF hydrogel-implanted rats showed significantly larger numbers of retrogradely labeled PhMNs per distance bin (blank: 8.75 ± 1.11 vs. HGF: 24.50 ± 2.96, t-test, p = 0.002, n = 4 rats/group; Fig. 4C) but only at the 1000–2000 μm distance bin. No differences in PhMN numbers were observed between the two groups at the lesion site (the 0–1000 μm bin, where complete loss of PhMN was noted; blank: 0 vs. HGF: 0) or at either of the more rostral portions of the PhMN pool in the 2000–3000 μm bin (blank: 25.75 ± 1.25 vs. HGF: 29.75 ± 4.52) or the 3000–4000 μm bin (blank: 30.25 ± 1.65 vs. HGF: 31.00 ± 2.08).

Retrograde labeling of phrenic motor neurons (PhMNs). To quantify PhMN numbers of ipsilesional spinal cord innervating the ipsilateral hemi-diaphragm, we selectively labeled PhMN cell bodies by intrapleurally injecting the retrograde tracer, CTB. We quantified CTB-labeled PhMN numbers at specific distances relative to the lesion epicenter. The lesion epicenter was defined as the zero distance, and four separate distance bins across the PhMN pool were examined (0–1000, 1000–2000, 2000–3000, and 3000–4000 μm, all measured rostral to the epicenter). Compared with blank hydrogel (
Given the inverse relationship between neuron size and excitability, we also measured somal size of individual CTB-labeled PhMNs within the 1000–2000 μm distance bin, as effects on the size of spared PhMNs may impact their activation in response to bulbospinal input. We found that HGF hydrogel implantation significantly reduced the average cell body size of CTB-labeled PhMNs. These data were plotted either as the average value for all PhMNs within a given animal (blank, 558.25 ± 20.40 μm2; HGF, 419.5 ± 16.91 μm2; t-test; p < 0.0001; n = 4 rats/group; Fig. 4D) or for every individual PhMN across all animals within a group (blank, 581.67 ± 21.47 μm2; HGF, 428.95 ± 17.06 μm2; t-test; p < 0.0001; n = 38–58 neurons/group of 4 animals; Fig. 4E).
To quantify the effects of HGF hydrogel on PhMN innervation of the diaphragm using morphological assessment, we performed detailed analysis of individual NMJ phenotypes at the dorsal, medial, and ventral subregions of the ipsilateral hemi-diaphragm. 59,60 At 8 weeks post-SCI, we performed whole-mount immunohistochemistry on the ipsilateral hemi-diaphragm by labeling phrenic motor axons all the way to their terminals with neurofilament (SMI-312) and synaptic vesicle protein 2 (SV2) antibodies. In addition, we labeled post-synaptic nicotinic acetylcholine receptors in the muscle with Alexa Fluor 555-conjugated α-bungarotoxin. Compared with blank hydrogel (Fig. 5A), HGF hydrogel (Fig. 5B–E) significantly increased the percentage of fully innervated/intact NMJs (representative NMJ morphology shown in Fig. 5C) in the dorsal subregion (blank: 69.09 ± 6.11%; HGF: 94.47 ± 1.15%; p = 0.0025; t-test; n = 4–5 rats/group; Fig. 5F). In addition, compared with blank hydrogel, HGF decreased the percentage of both partially denervated NMJs in the dorsal subregion (representative NMJ morphology shown in Fig. 5D; blank: 20.60 ± 3.36%; HGF: 3.25 ± 1.16%; p = 0.0010; t-test; n = 4–5 rats/group; Fig. 5G) and completely denervated NMJs in the dorsal subregion (representative NMJ morphology shown in Fig. 5E; blank: 5.06 ± 1.46%; HGF: 0%; p = 0.0055; t-test; n = 4–5 rats/group; Fig. 5H). In alignment with the EMG data, we observed little-to-no denervation of the ipsilateral hemi-diaphragm at either ventral or medial subregions in either contusion groups, and we also found no differences between blank hydrogel and HGF hydrogel groups at ventral and medial subregions (Fig. 5F–H). We did not conduct NMJ analysis in the contralateral muscle because our previous work showed that NMJ innervation of the contralateral hemi-diaphragm is not significantly affected following unilateral cervical contusion SCI. 49 Collectively, these findings with both CTB retrograde labeling and NMJ morphological analysis demonstrate that HGF significantly enhanced diaphragm innervation by PhMNs following cervical SCI.

HGF hydrogel preserved diaphragm neuromuscular junction (NMJ) innervation. In the ipsilateral hemi-diaphragm at 8 weeks post-contusion/hydrogel implantation, we labeled phrenic motor axons and their terminals with neurofilament (SMI-312) and synaptic vesicle protein 2 (SV2) antibodies, respectively (both labeling in green). We labeled post-synaptic nicotinic acetylcholine receptors with Alexa555-conjugated α-bungarotoxin (red). Compared with blank hydrogel
HGF hydrogel reduced lesion size and motor neuron degeneration
To determine whether HGF hydrogel promoted neuroprotective effects centrally within the cervical spinal cord, we quantified both lesion size and motor neuron cell body degeneration in Nissl myelin-stained transverse sections at 8 weeks post-injury. Compared with blank hydrogel control (Fig. 6A–C), HGF hydrogel (Fig. 6D–F) decreased lesion area at multiple rostral–caudal distances from the injury epicenter (Fig. 6G), as well as significantly reduced overall lesion volume (Fig. 6H; blank: 4.56 ± 0.23 mm3; HGF; 3.61 ± 0.30; p = 0.0242; t-test; n = 7–8 rats/group). We also found that HGF hydrogel decreased the loss of motor neuron cell bodies in the ipsilateral ventral horn (Fig. 6I, J) at multiple rostral–caudal distances from the lesion epicenter (Fig. 6K), as well as significantly reduced overall loss of motor neuron cell body numbers across a 1 cm rostral–caudal expanse of spinal cord centered about the lesion epicenter (Fig. 6L; blank: 126 ± 10 motor neurons; HGF: 195 ± 15; p = 0.0017; t-test; n = 7–9 rats/group).

HGF hydrogel reduced lesion size and motor neuron degeneration. We quantified both lesion size and motor neuron cell body degeneration in Nissl myelin-stained transverse sections at 8 weeks post-injury. Compared with blank hydrogel control
HGF hydrogel reduced levels of scar-associated CSPG molecules
To assess levels of the axon growth-inhibitory glycosaminoglycan portion of CSPG, we performed immunohistochemistry on the cervical spinal cord using the CS56 antibody. Compared with blank hydrogel implanted animals (Fig. 7A), HGF hydrogel (Fig. 7B) decreased levels of CS56 immunostaining at multiple rostral–caudal distances from the lesion epicenter (Fig. 7C), as well as significantly reduced the overall volume of CS56 labeling across a 6 mm rostral–caudal expanse of spinal cord centered about the lesion epicenter (Fig. 7D; blank: 4.22 ± 0.20 mm3; HGF, 3.24 ± 0.20; p = 0.0059; t-test; n = 6–7 rats/group).

HGF hydrogel reduced levels of scar-associated CSPG molecules. We performed immunohistochemistry on cervical spinal cord using CS56 antibody. Compared with blank hydrogel-implanted animals
Discussion
Development of a novel HGF biomaterial delivery system
Neurotrophic factors have gained considerable interest as therapeutic agents for the treatment of central nervous system (CNS) injuries and degenerative diseases due to their promising regenerative and neuroprotective effects in animal models. However, practical application of these molecules in clinical settings has been hampered by significant translational challenges, including problems with drug delivery (limited CNS permeability and poor pharmacokinetics) 63 and serious adverse side effects associated with off-target systemic exposure. 64,65 Local drug delivery strategies offer a potential solution to these issues and may also help prevent or limit potential unwanted pleiotrophic effects associated with growth factors. To this end, we have introduced a novel approach for the targeted, local administration of HGF to the injured spinal cord by developing an implantable HGF-loaded hydrogel that can be safely applied to the injury site.
Several local delivery methods have been previously used to successfully administer HGF to the spinal cord, including bolus injections via lumbar puncture and the use of implantable intrathecal infusion pumps. 34,38,40,42 However, cerebrospinal fluid (CSF) undergoes frequent turnover and gets replaced approximately every 6 h. 66 This rapid replenishment means that drugs introduced directly into the CSF are swiftly cleared away. While implantable pumps address this issue by providing a steady infusion of drugs over a prolonged period of time, they come with significant drawbacks. These include their high cost, invasiveness, and increased risk for the development of complications such as infections, scarring, and pump dysfunction or failure over time. 67,68 Moreover, in clinical practice, intrathecal drug delivery via bolus injections or catheters to the spinal cord is typically performed in the lumbar space, which provides the largest intrathecal space to minimize the risk of spinal cord damage. This will cause HGF to diffuse throughout the CNS, significantly decreasing its localization at the injury site. This could lead to reduced efficacy and an increased potential for off-target effects. In contrast, our hydrogel can be subdurally implanted on top of the injured spinal cord, permitting localized release of HGF at the injury site over an extended period.
Our innovative delivery system provides a clinically viable strategy for locally administering HGF following cervical SCI. All materials used in this platform are natural polysaccharides that are biocompatible (i.e., nontoxic and nonimmunogenic), biodegradable, and have been safely used in humans for a variety of clinical applications. 69 –74 Furthermore, our HGF hydrogel can be directly implanted at the injury site without causing damage to the spinal cord, 75 and we have successfully used this method to deliver a number of other drug-loaded hydrogels in SCI models. 51,57,76 From a practical standpoint, treatment can be completed in a single application, which can conveniently be performed at the same time as other procedures routinely used to manage acute SCIs, such as spine decompression and stabilization surgeries.
HGF hydrogel has a neuroprotective effect on respiratory circuitry following SCI
The present study offers the first evidence of HGF’s beneficial impact on highly important respiratory circuitry following SCI. We found that, compared with the blank hydrogel control, our HGF hydrogel effectively maintained diaphragm function, as demonstrated by significant increases in EMG and CMAP amplitudes recorded in vivo from the ipsilateral hemi-diaphragm at multiple time points after injury. Although respiratory activity did not completely reach levels observed in laminectomy-only animals, our results nevertheless carry profound clinical implications for addressing respiratory dysfunction, a critical aspect of post-SCI recovery.
While HGF is known to exert both neuroprotective and regenerative properties in the CNS, our data suggest that its beneficial effects on respiratory function are primarily driven through neuroprotective mechanisms. We observed a decrease in lesion size and an increase in motor neuron survival following treatment with HGF hydrogel. This is consistent with previous findings describing the positive effects of HGF on neuronal survival in a variety of pathologies, including SCI, 33 ALS, 37,38 and cerebral ischemia. 77,78 HGF inhibits apoptosis through a number of molecular pathways, 25 including activation of Akt, 79 extracellular signal-regulated kinase (ERK), 80 and signal transducer and activator of transcription 3 (STAT3) 77 signaling. These prosurvival signals may help spare PhMNs after injury, and this central neuroprotective effect likely contributes to the decrease in diaphragm NMJ denervation we see in HGF hydrogel-treated animals. Our functional analyses also revealed robust effects on diaphragm function at relatively early time points post-injury, further suggesting that HGF is working to preserve existing function rather than by inducing restorative circuit plasticity. Additionally, although neurotrophic factors are known to promote axon growth, 81 –83 we did not observe this effect in our model. While we did not conduct an exhaustive assessment of axon growth, we did examine the potential for increased serotonergic fiber sprouting, as it is known to play an important role in modulating lower motor neuron excitability. 84,85 Such sprouting has the potential to strengthen excitatory bulbospinal inputs to PhMNs from spared or regenerating rVRG axons, thereby increasing respiratory function. 51,76 However, our data from this study did not reveal any significant differences between control hydrogel and HGF hydrogel on serotonergic axon density within the PhMN pool (data not shown).
Based on the lesion size analysis, the degree of tissue preservation was relatively small, though it was statistically significant. However, we do find significant protection of motor neuron numbers in the ventral horn and preservation of innervation at the diaphragm NMJ. Collectively, these data strongly support that the observed diaphragm functional effects were mediated (at least in large part) by some form of neuroprotective effect. That being said, we have not conclusively established which specific aspect of this neuroprotection is the mechanism(s) responsible for the functional outcomes. The combination of the preserved NMJ innervation (based on the NMJ morphology and CTB analyses) and the CMAP effects suggests that protection of the PhMN-to-diaphragm component of the circuitry played a major role. It is also possible that there was some sparing of descending axonal input into the PhMN pool, though we did not perform analyses to address this using—for example—anterograde labeling of bulbospinal respiratory axon pathways.
Although our results point toward a neuroprotective role of HGF in the context of respiratory circuitry, there are additional mechanisms by which it may also be acting to impact functional outcome. Interestingly, our results demonstrate that although the HGF hydrogel preserved PhMN number, it also led to a reduction in average somal size. As smaller size of motor neurons is associated with increased excitability, 86 this could translate into a physiologically beneficial change that facilitates increased PhMN activity and diaphragm activation. Other changes may also be at play. For example, HGF has been shown to modulate inflammation. HGF inhibits nuclear factor kappa-light-chain-enhancer of activated B cells signaling, 87 decreasing neutrophil infiltration 88 and the production of cytokine and chemokines after SCI. 35 These anti-inflammatory effects help attenuate secondary damage and improve functional outcomes. HGF also acts as a potent angiogenic factor. 89 The formation of new blood vessels post-SCI is important for recovery as they provide critical structural and trophic support for tissue repair and survival. 90 In addition, increases in blood flow 91 or vessel density have also been found to correlate with improvements in functional recovery. 92 Further studies are needed to fully explore the extent of these effects in the context of our HGF hydrogel treatment paradigm.
Combinatorial potential of HGF hydrogel with regenerative therapies
We demonstrate that treatment with HGF hydrogel induces a significant, yet incomplete, improvement in diaphragm function. Since our results indicate that neuroprotection rather than axon regrowth is the major contributor to this effect, a combinatorial approach encompassing other therapeutic strategies, especially ones that induce a strong regenerative response, may enhance overall treatment efficacy.
In this context, early administration of HGF hydrogel serves two important roles in creating favorable conditions for regeneration. First, it effectively preserves components of the respiratory circuitry, offering potential substrates for subsequent regenerative therapies to target. These spared elements could be induced to regrow and reconnect with their pre-injury targets, thereby restoring the original circuit. Additionally, they could be recruited into the formation of new circuits, which may also contribute to improvements in functional recovery. 93 Second, early HGF delivery can modulate the environment of the injured spinal cord, rendering it more permissible to axon regrowth. Of note, we observed a marked decrease in scar-associated CSPG molecules in HGF versus blank hydrogel implanted animals. HGF is known to possess anti-scarring properties 36,94 by reducing the secretion of transforming growth factor betas 1 and 2 from reactive astrocytes, decreasing the production of CSPGs. 36 CSPGs are a major component of the scar and potent inhibitors of axon growth; 95 therefore, reducing CSPG expression via HGF hydrogel is potentially an effective strategy for encouraging regrowth.
From a practical standpoint, there are a multitude of effective regenerative therapies 96 that can be potentially considered for use in conjunction with our HGF hydrogel following SCI. For example, recent clinical trials have highlighted the potential of cell implantation to enhance both motor and neurological function. 97 Of note, HGF has been shown to enhance differentiation, 98 survival, and synaptic connectivity of grafted neural stem cells after SCI, leading to improvements in functional recovery. 99 Similarly, in vitro studies have demonstrated that HGF, when combined with glial cell-derived neurotrophic factor, appears to promote the differentiation of bone-marrow-derived mesenchymal stem cells into neuron-like cells, 100 further highlighting the potential of HGF in combination with stem cell transplantation. Beyond cell-based therapies, another strategy could involve targeting intrinsic mechanisms of axon growth inhibition, such as phosphatase and tensin homolog (PTEN) signaling. We have previously shown that inhibition of PTEN is effective at inducing long distance axon regeneration in the respiratory circuitry after cervical SCI, 101 even in the setting of chronic injury. 102 This makes it another attractive candidate for combinatorial therapy with HGF hydrogel, although there are many other possible approaches beyond those discussed here.
Conclusions
In conclusion, our study introduces a novel and safe biomaterial platform for the localized delivery of HGF to the injured cervical spinal cord. Our findings demonstrate that HGF hydrogel preserves respiratory circuitry, maintains functional diaphragm innervation, reduces lesion size, and mitigates motor neuron degeneration. While the primary mechanism of HGF in this study appears to be neuroprotection rather than axon regrowth, it sets the stage for an additive or even synergistic approach with other regenerative therapies to enhance overall treatment efficacy. Our results may hold important implications for patients in mitigating respiratory dysfunction after SCI.
Transparency, Rigor, and Reproducibility Statement
We performed all statistical analysis using Prism 10 (GraphPad Software). All data were presented as mean ± SEM. Statistics were considered significant at p < 0.05. Sample sizes were determined based on a priori power analysis. Prior to beginning this study, we randomly assigned animals to the experimental groups, and we randomly distributed various surgery procedures used within a given experiment across these animals (and within a given surgical day). We conducted all surgical procedures, electrophysiological recordings, and histological analyses in a blinded manner. In the Results section and in the figure legends, we provide details of n’s, statistical tests used, and the results of all statistical analyses for each experiment and for all statistical comparisons. We also authenticated experimental regents to ensure that they performed similarly across experiments and to validate the resulting data.
Footnotes
Authors’ Contributions
Conceptualization: Y.Z. and A.C.L. Data curation: S.J.T., Y.Z., and A.C.L. Formal analysis: S.J.T., B.G., Z.W., M.Y., J.N., J.S., and M.C.W. Funding acquisition: Y.Z. and A.C.L. Investigation: S.J.T., B.G., Z.W., M.Y., J.N., J.S., and M.C.W. Project administration: Y.Z. and A.C.L. Supervision: Y.Z. and A.C.L. Validation: S.J.T., B.G., Z.W., M.Y., J.N., J.S., and M.C.W. Visualization: S.J.T., B.G., and M.C.W. Roles/Writing—original draft: S.J.T., B.G., and A.C.L. Writing—review and editing: S.J.T., B.G., Y.Z., and A.C.L.
Author Disclosure Statement
The authors have no competing interests to disclose.
Funding Information
This work was supported by the NINDS (