
Abstract
Trauma to the brain can induce a contusion characterized by a discrete intracerebral or diffuse interstitial hemorrhage. In humans, “computed tomography-positive,” that is, hemorrhagic, temporal lobe contusions (tlCont) have unique sequelae. TlCont confers significantly increased odds for moderate or worse disability and the inability to return to baseline work capacity compared to intra-axial injuries in other locations. Patients with tlCont are at elevated risks of memory dysfunction, anxiety, and post-traumatic epilepsy due to involvement of neuroanatomical structures unique to the temporal lobe including the amygdala, hippocampus, and ento-/perirhinal cortex. Because of the relative inaccessibility of the temporal lobe in rodents, no preclinical model of tlCont has been described, impeding progress in elucidating the specific pathophysiology unique to tlCont. Here, we present a minimally invasive mouse model of tlCont with the contusion characterized by a traumatic interstitial hemorrhage. Mortality was low and sensorimotor deficits (beam walk, accelerating rotarod) resolved completely within 3–5 days. However, significant deficits in memory (novel object recognition, Morris water maze) and anxiety (elevated plus maze) persisted at 14–35 days and nonconvulsive electroencephalographic seizures and spiking were significantly increased in the hippocampus at 7–21 days. Immunohistochemistry showed widespread astrogliosis and microgliosis, bilateral hippocampal sclerosis, bilateral loss of hippocampal and cortical inhibitory parvalbumin neurons, and evidence of interhemispheric connectional diaschisis involving the fiber bundle in the ventral corpus callosum that connects temporal lobe structures. This model may be useful to advance our understanding of the unique features of tlCont in humans.
Introduction
In humans, traumatic brain injury (TBI) involving the temporal lobes has unique features distinct from TBI involving other lobes of the brain. Unlike other lobes, the temporal lobes in humans are weakly protected by a thin overlying bone, the squamous portion of the temporal bone, that is easily depressed or fractured by blunt trauma to the side of the head. A severe blow to the temporal region can lead to a so-called “burst temporal lobe”—a temporal lobe contusion (tlCont) with an intracerebral hemorrhage in continuity with a subdural hemorrhage, which together predispose to brain herniation and high mortality. 1,2
A “computed tomography (CT)-positive,” that is, hemorrhagic, tlCont from nonfatal trauma can be associated with debilitating morbidity, as highlighted by a landmark study from the TRACK-TBI consortium. 3 The authors reported that, of patients with mild TBI complicated by a tlCont, fully 46% had 6-month disability graded as moderate or worse (Glasgow Outcome Scale Extended [GOSE] ≤ 6). By comparison, 6-month disability graded as moderate or worse was identified in only 26% of CT-positive subjects without tlCont, and in only 29% of CT-negative subjects. Moreover, 42% of subjects with tlCont were unable to return to work at 6 months, compared to 21% of CT-positive subjects without tlCont, and 20% of CT-negative subjects. The same study of mild-TBI patients showed that, on multivariable regression, the presence of a tlCont conferred an odds ratio (OR) of 3.17 (95% confidence interval [CI; 1.11–9.07], p = 0.031) for moderate or worse disability on GOSE and an OR of 4.48 ([1.49–13.51], p = 0.008) for inability to return to baseline work capacity, whereas frontal contusion, extra-axial injury, and other intra-axial injuries did not show statistically significant associations with disability on GOSE or return to work status. 3 These severe adverse findings may be due, in part, to the fact that patients with tlCont are at an elevated risk of memory dysfunction, anxiety, and post-traumatic epilepsy due to involvement of neuroanatomical structures unique to the temporal lobe, including the amygdala, hippocampus, and ento-/perirhinal cortex. 4 –6
Little progress has been made in elucidating the specific pathophysiology unique to tlCont, largely because no preclinical model of tlCont has been described. In mice, the temporal lobe lies deep to the ramus of the mandible, the root of the zygoma, the temporomandibular joint, and the thick temporalis muscle, immediately anterior to the ear (Fig. 1). These anatomical features in the mouse make it difficult to deliver an experimental traumatic impact directly to the temporal lobe. The frequently used techniques to induce TBI in rodents, including controlled cortical impact (CCI), 7,8 fluid percussion injury (FPI), 9 –11 and impact-acceleration 12 are readily applied to the parietal and frontal lobes. However, impacting the temporal lobe in mice with these techniques would require surgery that would harm structures critical for chewing, eating, and overall well-being.

Approach to the temporal lobe in humans versus mice. (
To overcome these anatomical limitations, we developed a tlCont model in the mouse with the contusion characterized by a traumatic interstitial hemorrhage. Our model utilizes a Fogarty balloon catheter to induce injury. The balloon is introduced extradurally via a small parietal craniectomy and is advanced anteroventrally into the middle cranial fossa lateral to the temporal lobe. Rapidly inflating/deflating the balloon induced a traumatic brain contusion with diffuse interstitial hemorrhage that was associated with low mortality and persistent neurological sequelae.
Methods
Ethics statement
Animal experiments comply with the Animal Research: Reporting of In Vivo Experiments (ARRIVE) guidelines and were performed under a protocol approved by the Institutional Animal Care and Use Committee of the University of Maryland School of Medicine and in accordance with the relevant guidelines and regulations as stipulated in the United States National Institutes of Health Guide for the Care and Use of Laboratory Animals.
Subjects
C57BL/6 mice, 22–28 g, 10–12 weeks of age, were obtained from Envigo (Indianapolis, IN, USA). Four series of mice were studied (Table 1), comprised of all males except in series 1. In series 1, 12 mice with tlCont were used to visualize the contusions at 15 min (two males) and 24 h (six males and four females). In series 2, 10 mice with tlCont and 10 uninjured controls were used for immunohistochemistry on day 7 (five mice/group) and day 45 (five mice/group); mice in this series did not have electrodes implanted. In series 3, 10 mice with tlCont and 10 uninjured controls were used for neurofunctional tests, including beam walk and accelerating rotarod on days –1, 1, 3, 7; open field test and novel object recognition on day 14; elevated plus maze on day 21, and Morris water maze on days 30–35; in this series, additional mice, three/group, were added for the novel object recognition test; mice in this series did not have electrodes implanted. In series 4, 10 mice with tlCont and 13 uninjured controls were used for pentylenetetrazol (PTZ) challenge on day 22, with 8 mice/group used for electroencephalogram (EEG) recording on days 7–21; in this series, 3 mice died and 7 mice had unusable EEG signals. For mice with planned survival >24 h, mortality post-tlCont was 0/10, 0/13, and 3/13 in series 2, 3, and 4, respectively.
Experimental Series
EEG, electroencephalogram.
Temporal lobe contusion
Mice were anesthetized (isoflurane: induction, 3%, maintenance, 1–1.5%), mounted in a stereotaxic apparatus (Stoelting, Wood Dale, Il), and allowed to ventilate spontaneously. Normal body temperature was maintained using a heating pad (Kent Scientific, Torrington, CT) and vital signs were monitored using a MouseOx (STARR Life Sciences, Oakmont, PA), which records O2 saturation, heart rate, and ventilation rate. Surgery was performed using aseptic/sterile techniques. Local anesthetic (1% lidocaine) was injected and a midline scalp incision was made. Using a high-speed drill with saline irrigation to prevent overheating, a 2-mm craniectomy was made in the parietal bone at the occipital crest (x, –4.0 mm; y, +3.5 mm). A blunt probe (0.3 mm diameter × 8 mm long) was introduced epidurally and advanced at ∼45-degree angle rostrally toward the floor of the middle fossa to strip the dura from the periosteum. Using a custom device (Fig. 2), the balloon of a 2-French Fogarty balloon catheter (Edwards Life Sciences, Washington, DC) was test-inflated in a 1 mL syringe containing sterile normal saline (NS) to confirm the desired inflated volume of 70–80 µL. The deflated balloon was gently introduced epidurally into the middle fossa, lateral to the temporal lobe, then was inflated for 2 sec to induce a contusion. The balloon catheter was removed and the scalp incision was closed. Balloon inflation/deflation invariably was accompanied by apnea, which resolved spontaneously after 6–12 sec. The entire surgical procedure to induce the contusion was completed within 12–15 min, following which the animals emerged from anesthesia and were returned to their home cages.
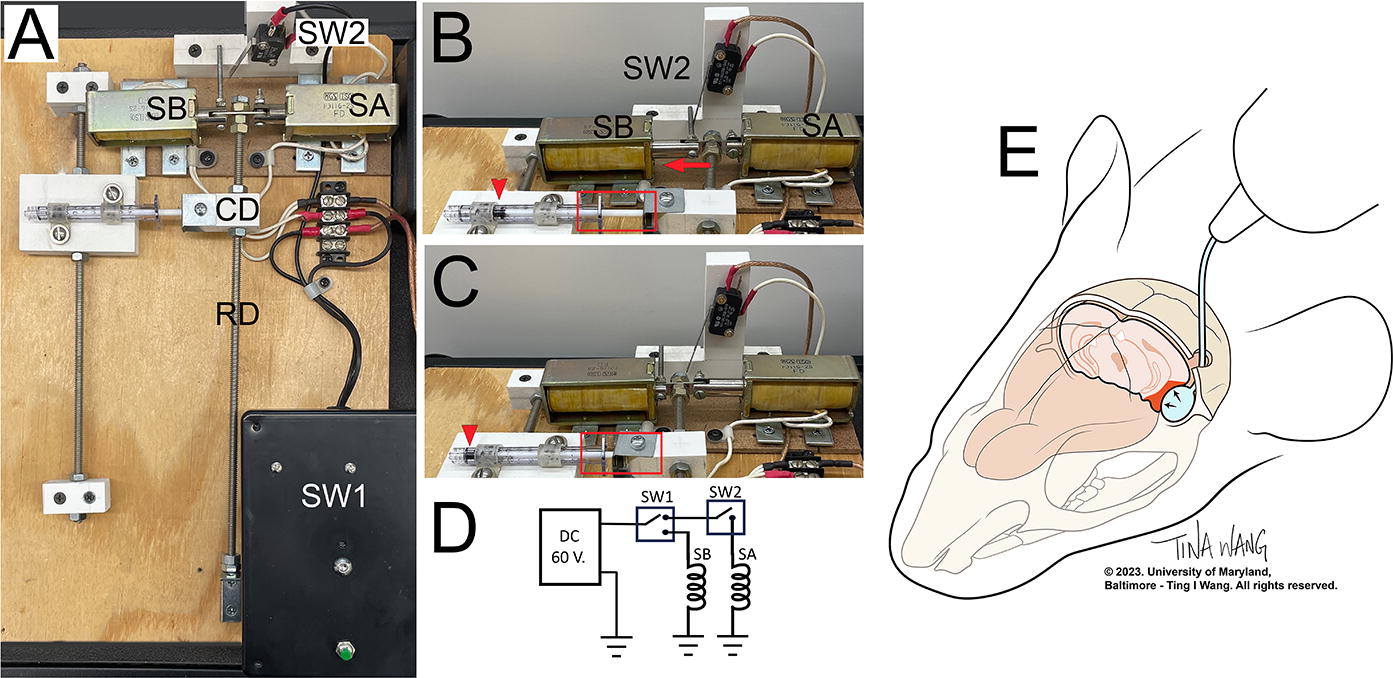
Custom device to rapidly inflate/deflate a 2-French Fogarty balloon catheter. (A)–(C) Reproducible balloon inflation to 70–80 µL is obtained using a 1 mL syringe whose plunger is controlled by a pair of solenoids, SA and SB. SA and SB pull in opposite directions on the end of a rod (RD), forcing the rod to its “resting” vs. “active” positions, respectively. The opposite end of the rod pivots, so that the horizontal traverse of the rod is minimal near the pivot point and maximum at the solenoids. The rod has a CD that connects it to the plunger of the syringe. The position of the coupling device and syringe can be moved along the rod, closer or farther from the solenoids, to increase or decrease the horizontal movement of the syringe plunger, thereby adjusting the volume of air that inflates the balloon. Due to intrinsic elasticity of the Fogarty balloon catheter, a greater volume of air must be injected from the syringe to obtain the desired change in balloon volume, measured by displacement of water. The photographs show the overall device
Electrode implantation surgery
For EEG recordings, mice were implanted with a head mount that included a single stainless-steel depth electrode, two epidural cortical contacts (stainless-steel screws), and a common/ground electrode (8201-DEP; Pinnacle Technology, Inc, Ontario, CA). Immediately following the procedure for tlCont, a small burr hole was made for stereotactic placement of the depth electrode into the ipsilateral hippocampus (x, –1.7 mm; y, +1.6 mm; z, –2 mm). Uninjured controls without tlCont also underwent the 2-mm parietal craniectomy, as above. The head mount was secured to the skull with four screws with the following approximate electrode locations (all ipsilateral): cortical electrode 1: 1 mm anterior, 0.5 mm lateral to bregma; cortical electrode 2: 3 mm posterior, 0.5 mm lateral to bregma; reference: 3 mm posterior, 3 mm lateral to bregma; ground: frontal bone. Electrode contact points were coated with silver epoxy, dental acrylic was applied over the screws, and the surgical incision was closed with sutures.
Beam walk
Mice were analyzed for motor coordination and balance. They were trained to walk 3 feet across a 0.5-inch-wide beam to enter a dark box through a small opening. 13 An annoying noise generated by shaking keys was used to encourage the mice to walk across the beam. The time required for mice to pass the 3-foot distance and enter the box was recorded. Three trials were conducted on each test day, each separated by 10 min of rest. The average latency of the three trials was recorded.
Accelerating rotarod
The accelerating rotarod test was used to assess coerced locomotor activity. 14,15 The mice were placed on the drum of the accelerating rotarod (IITC, Life Science, Woodland Hills, CA), starting at 4 rpm, accelerating at a rate of 2 rpm every 5 sec up to a maximum of 40 rpm, with three trials separated by 20 m administered on each day of testing. The drum surface was grooved to provide a better grip. We report the average latency to falling off the drum. The latency reported for each animal is the average of the three trials.
Open field test
The mice were placed in an open field made of a clear plexiglass box (40 × 40 × 100 cm) and equipped with a video tracking system (Any-Maze, Stoelting Co, Wood Dale, IL, USA), as described. 16,17 Each mouse was placed in the middle of the open field for a single 5-min session with the experimenter out of view. The uniformly illuminated floor space was divided into equal squares, corners, center, and intermediary regions. The measurement reported is time spent in the center. After each trial, fecal pellets were removed and the floor was wiped clean with 70% alcohol and dried.
Elevated plus maze
This test was used to assess anxiety-like behavior. 18,19 The apparatus (Coulbourn Instruments, Holliston, MA) consists of two open (50 × 10 cm) and two closed (50 × 10 cm) arms. The closed arms are surrounded by 30-cm-high walls painted black. Open and closed arms enclose a central region of 10 × 10 cm. The maze was elevated to a height of 50 cm above the floor. Each mouse was placed in the center of the maze facing an open arm and allowed to travel throughout the maze for 5 min. The time spent in the open arms of the maze, with the mouse exposing at least its head and body up to its forelimbs in the open arm, was quantified using Any-Maze video tracking software.
Novel object recognition test
This test was used to evaluate recognition memory. The test consisted of pretraining, training, and testing sessions performed as described. 20,21 Mice were habituated to the open field apparatus (40 × 40 × 100 cm) without any object for 5 min. The following day, mice were trained in the same field with two similar objects for 5 min. The objects used were of similar size and made of easy-to-clean plastic. During the acquisition phase, two identical objects were set in the field at a distance from each other. The mouse was placed in the testing arena and allowed to explore freely for 5 min, then was returned to its home cage. The objects were replaced by another set of objects, one identical and one novel; after 1 h, the mouse was returned to the arena and allowed to explore the objects for 5 min (the recognition phase). The time spent interacting with each object as well as the overall time exploring the objects, whether old or new (total exploration time), was measured. The recognition index was calculated as the time spent interacting with the novel object divided by total exploration time × 100. Animals that discriminate the new from the old object have a recognition index/exploratory preference >50%.
Morris water maze test
This test was used to assess spatial learning and memory. The apparatus consisted of a circular water tank (75 cm diameter) and a hidden circular rescue platform (8 cm diameter) about 1–1.5 cm below water surface located in the middle of the northwest quadrant. The pool was filled with tap water that was made opaque with nontoxic white tempera paint (Colorations, Discount School Supply®, Chicago, IL). The water temperature was kept at 21°C–24°C. The specific paradigms used for testing in this laboratory have been described previously. 22 Briefly, on days 1–5 of testing, mice are given four trials/day from four different entry spots (N, S, W, E), up to 2 min maximum each (30-min intertrial interval) to find the hidden platform. Mice are trained to sit on the platform for 25–30 sec before they are moved out of the pool. Animals that do not reach the platform during the trial are held by the tail on the platform. For incremental learning, daily latency was calculated as the average of the four trials of each of the 5 days. On day 6, the platform is removed and a memory probe is conducted by determining the time spent in the correct quadrant (where the platform was previously placed) during the 1-min test duration. Mice were tracked by Any Maze™ (Stoelting Co, Wood Dale, IL, USA) video tracking software through the video camera located on top of the apparatus.
Continuous EEG monitoring in mice and seizure threshold testing
One week after tlCont, mice were connected to a continuous video EEG monitoring system with a preamplifier (8406-SE). The EEG was sampled at 2 kHz during the 24-h live video recordings utilizing Sirenia Software 2.2.11 (Pinnacle Technology). Animals were recorded continuously on days 7–21 post-tlCont to monitor EEG and behavioral seizure activity. Electroencephalographic seizures were automatically detected and quantified using custom MATLAB scripts, with subsequent manual validation by two blinded observers. Interictal spikes were identified as voltage deflections exceeding six standard deviations above or below the mean baseline voltage. Seizures were defined by changes in EEG activity characterized by repetitive high voltage, high-frequency spike/wave discharges lasting 5 sec or more. On day 22 post-tlCont, we administered a single dose of PTZ (25 mg/kg) to evaluate acute seizure thresholds, observing the mice for 1 h. Behavioral responses to the PTZ challenge were assessed (unblinded) using the Racine Scale. 23
Histology
The brains of mice in series 1 were processed after tlCont to visualize contusions. Under deep anesthesia, mice were euthanized and underwent transcardiac perfusion with NS (20 mL). Brains were harvested and photographed. Ten brains were fixed in 10% neutral buffered formalin, cryosectioned and stained with hematoxylin and eosin.
Immunohistochemistry
For these analyses, we studied only mice that did not undergo electrode implantation. Under deep anesthesia, mice were euthanized, underwent transcardiac perfusion with NS (20 mL) followed by 4% paraformaldehyde (20 mL). Brains were harvested and postfixed. Tissues were cryoprotected with 30% sucrose, frozen in Optimal cutting temperature (OCT), and cryosectioned (12 µm). Immunohistochemistry was performed as we described. 24 –26 In some cases, sections were first processed for antigen retrieval in citrate buffer pH 6 Antigen Retriever (C9999; Sigma) solution in a microwave (Pelco Biowave Pro+ microwave, Ted Pella. Inc). 27 For all immunolabelings, sections were incubated at 4°C overnight with primary antibodies directed against: glial fibrillary acidic protein (GFAP) (1:300; #C9205, Sigma Aldrich, St. Louis, MO, USA), ionized calcium binding adaptor molecule 1 (Iba1) (1:500; #019–19741; Wako Chemicals, Richmond, VA, USA), cleaved caspase-3 (cCasp3) (1:200; #9661S, Cell Signaling Technologies, Beverly, MA, USA), β-amyloid precursor protein (β-APP) (1:200, #MAB343, Millipore, Temecula, CA, USA), or parvalbumin (1:200; #P3088, Sigma, St. Louis, MO, USA). After several rinses in phosphate-buffered saline, sections were incubated with species-appropriate fluorescent secondary antibodies (Alexa Fluor 488 and 555, Molecular Probes, ThermoFisher Scientific, Waltham, MA, USA) for 1 h at room temperature. Controls for immunohistochemistry included the omission of primary antibodies.
Unbiased assessments of specific labeling were obtained using NIS-Elements AR software (Nikon Instruments, Melville, NY, USA) from sections (one section per mouse) immunolabeled as a single batch. All images for a given signal were captured using uniform parameters of magnification, area, exposure, and gain. For all quantitative analyses, the background for each specific antibody was determined by segmentation analysis by computing a histogram of pixel intensity for a particular regions of interest (ROI). Specific labeling was defined as pixels with signal intensity >2× that of the background for parvalbumin and >3× that of background for GFAP and Iba 1. Object counts were performed on parvalbumin staining to determine the number of cells in the specific area. For the quantification of parvalbumin staining, ROI was defined as follows: the dentate gyrus included the granule cell layer and the dental hilus, whereas the hippocampus outlined the entirety of the hippocampal formation including the molecular layer of CA1, CA2, and CA3 in coronal sections.
Data analysis
Data are presented as mean±S.E. or S.D., as indicated in the text. Student’s t-test, 1-way ANOVA or repeated measures ANOVA with Bonferroni post-hoc comparisons, or Mann-Whitney U test, were used, as appropriate. Daily hippocampal spike rates were analyzed using linear regression and the cumulative incidence of mice with seizures was analyzed using the Kaplan–Meier estimator. Analyses were performed with Origin Pro V8 or GraphPad Prism 8.3. A value of p < 0.05 was deemed to be statistically significant.
Results
Temporal lobe contusion
Rapid inflation/deflation of a 2-F Fogarty balloon in the middle cranial fossa was accompanied by important changes in vital signs. Balloon inflation/deflation invariably was accompanied by apnea that typically resolved spontaneously within 6–12 sec. The measurement of vital signs showed a marked drop in O2 saturation that lasted ∼2 min (Fig. 3A), mild bradycardia that peaked at ∼3 min (Fig. 3B), and spontaneous ventilation that dropped abruptly during the first 2 min (Fig. 3C). All vital signs normalized fully by 10 min. The surgical procedure for tlCont alone without electrode implantation required only ∼15 min, after which mice recovered quickly from anesthesia. There was no intraprocedural mortality. For mice with planned survival >24 h, mortality post-tlCont was 3/36, with deaths occurring within the first week in mice that underwent the extended surgical procedure for tlCont plus electrode implantation.
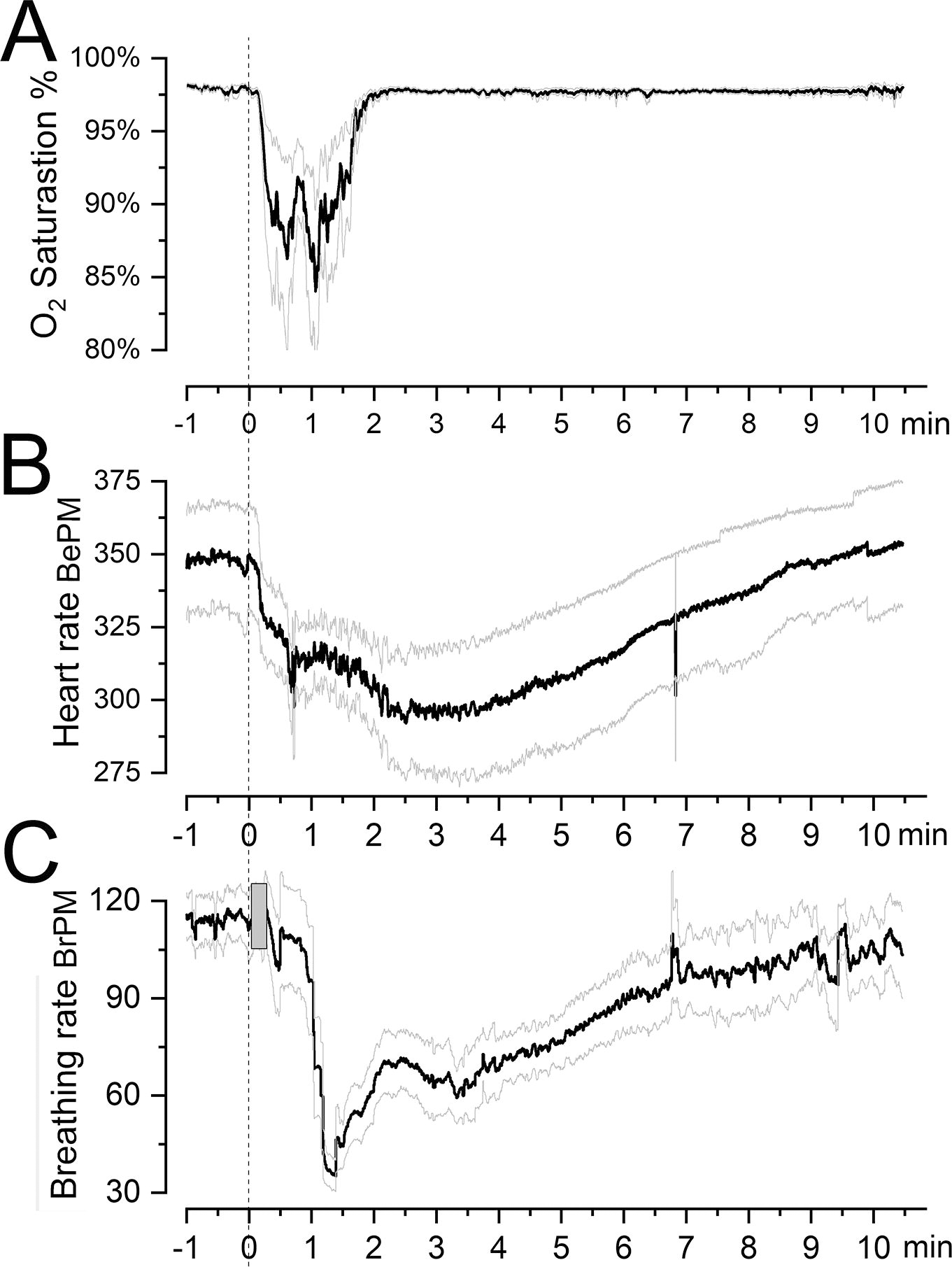
Changes in vital signs with induction of a tlCont. (
Rapid balloon inflation/deflation resulted in a contusion that was evident on the surface as well as within the temporal lobe (Fig. 4A,B). In eight consecutive mice, four were females and four were males, the contusion appeared reproducible, albeit perhaps somewhat less severe in females (Fig. 4C,D). Contusions examined 15 min after injury versus at 24 h showed no evidence of contusion expansion (Fig. 4E), suggesting that hemorrhagic progression of contusion, as seen with conventional CCI, 28 was not prominent in this model, at least within 24 h of trauma. TTC (2,3,5-triphenyltetrazolium chloride) staining suggested the presence of ischemia in the cortex superior to the contusion. The ischemic area involved the distal middle cerebral artery territory and extended up to the border zone of the anterior cerebral territory (Fig. 4E at 24 h), presumably reflecting trauma to the distal branches of the middle cerebral artery. Coronal sections stained with hematoxylin and eosin showed the damage and the cell loss in the ipsilateral compared to the contralateral temporal lobe cortex (Fig. 4F,G) and revealed no apparent injury to either dorsal hippocampus (Fig. 4H).
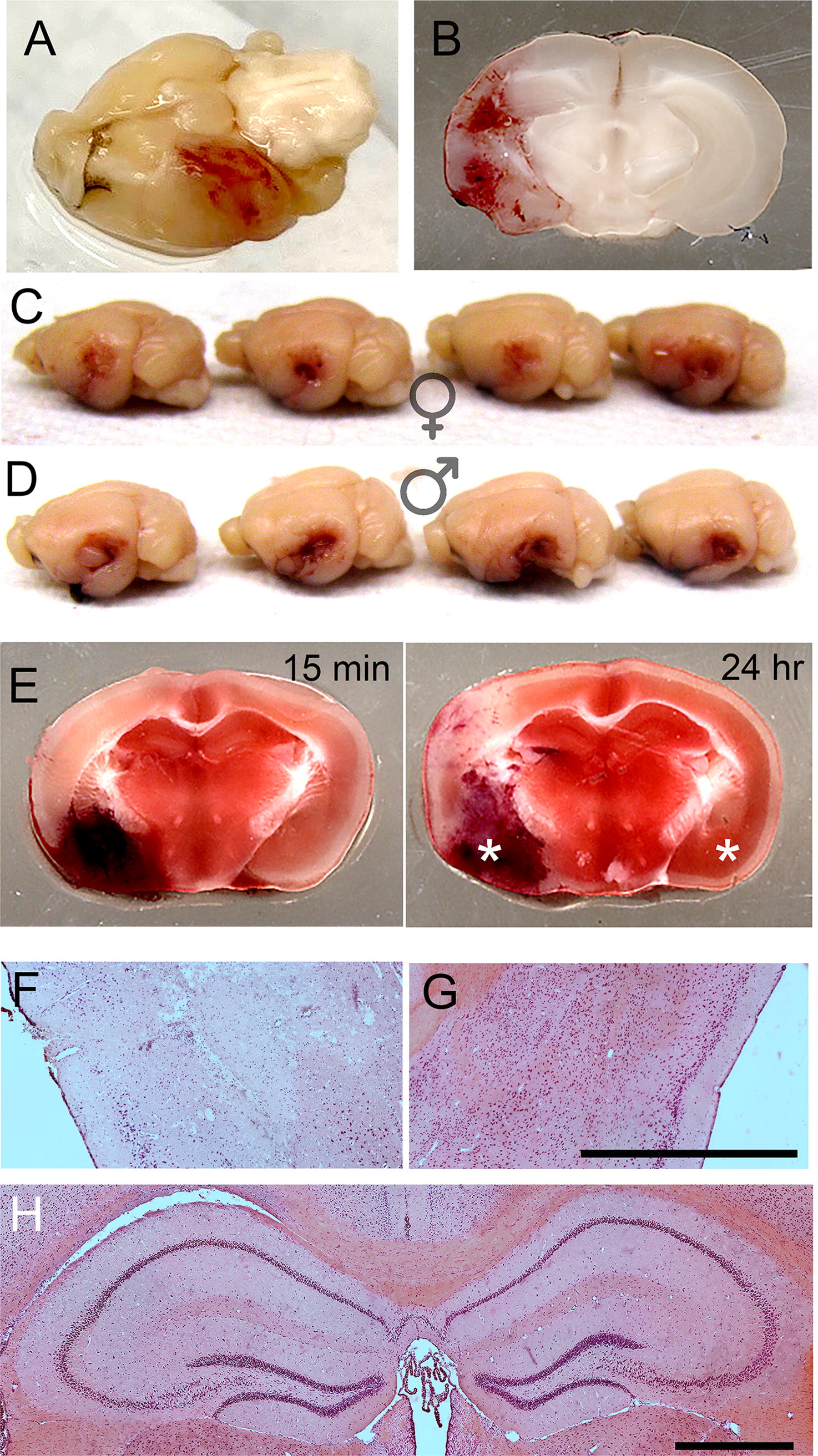
tlCont.
Sensorimotor function
Beam walk and accelerating rotarod were used to evaluate sensorimotor function. For both tests, measurements were compared to those obtained from uninjured control mice. Overall, tlCont was associated with only transient abnormalities in sensorimotor function that normalized rapidly over the course of 1 week (Fig. 5A,B). In both tests, significantly worse performance was observed 24 h post-tlCont but repeat testing on days 3 and 7 showed rapidly progressive improvements. On the accelerating rotarod, performance on day 7 was better than preinjury, consistent with practice-related improvement.
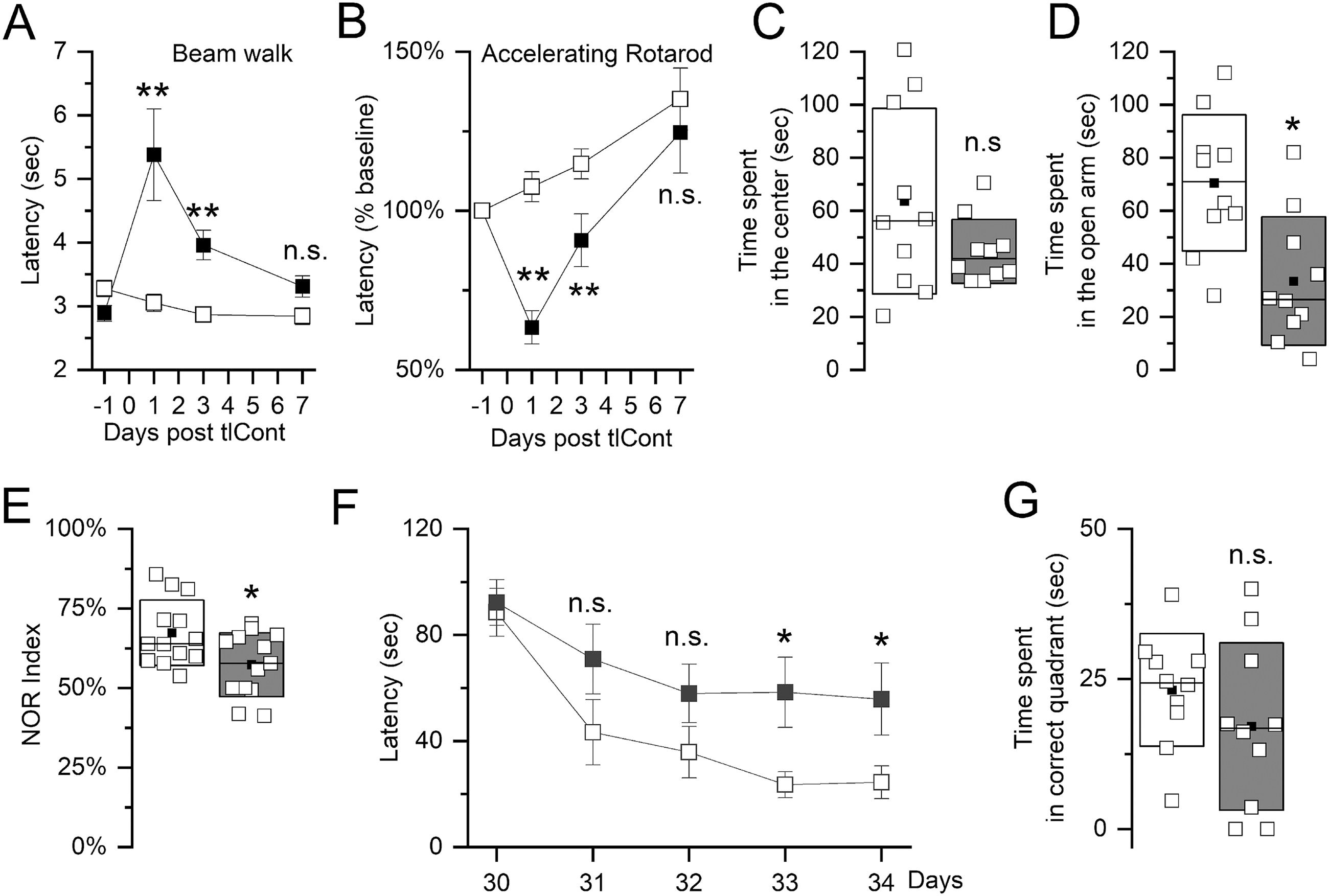
Neurological function following tlCont.
Anxiety
In the open field test, time spent in the center of the field is used as a measure of anxiety in mice. Control mice showed considerable variability on this test, whereas mice post-tlCont showed more uniform behaviors (Fig. 5C). Although average values in tlCont mice were lower, statistical significance was not reached.
In the elevated plus maze, time spent in the open arm is used as a measure of anxiety. In this test, variance was similar for both groups and mice with tlCont spent significantly less time in the open arm compared to controls (Fig. 5D).
Memory
The novel object recognition test assesses nonspatial object memory. In this test, variance was similar in both groups. Mice with tlCont spent significantly less time with the novel object compared to controls (Fig. 5E).
The Morris water maze was used to assess spatial memory. Mice with tlCont required more time to find the hidden platform on all test days compared to controls (Fig. 5F). Mice with tlCont also spent less time in the correct quadrant than control mice, although statistical significance was not reached (Fig. 5G).
Spontaneous seizures and interictal spikes
Continuous video EEG was used to investigate whether tlCont induced spontaneous seizure activity. We quantified the interictal spike rate and the incidence, number, and duration of electrographic seizures in the ipsilateral hippocampus (Fig. 6A,B). Compared with controls, tlCont mice had a significant increase in daily spike rate throughout the recording period (Fig. 6C). In addition, tlCont mice had significantly greater seizure incidence and total number of seizures (Fig. 6D,E), none of which were convulsive. The mean duration of seizures in tlCont mice was 23.7 ± 4.4 sec, while the sole seizure observed in a control mouse lasted 10 sec (Fig. 6F). The seizures remained focal in the hippocampus and there was no phenotypic or EEG evidence of seizure generalization.

Electroencephalographic abnormalities following tlCont.
Pentylenetetrazol challenge
To investigate seizure susceptibility after tlCont, we administered a PTZ challenge (25 mg/kg) and monitored behavioral seizure responses and interictal spikes for 1 h. tlCont mice had significantly greater seizure severity and interictal spike rates compared to controls (Fig. 6G,H). In addition, there was a nonsignificant trend towards an increase in the incidence of severe tonic-clonic seizures (Racine 4 or higher) in tlCont mice (control, 23%; tlCont, 60%; p = 0.08).
Immunohistochemistry
GFAP immunoreactivity on day 7 was used to assess astrogliosis. Compared to controls, GFAP immunoreactivity was widely increased after tlCont, more so on the side of injury but contralaterally as well (Fig. 7A,B). Compared with controls, GFAP immunolabeling was greatly increased in both the ipsilateral and contralateral dorsal hippocampus (Fig. 7C–E). In 5/5 specimens examined, there was no histological evidence of physical damage to the ipsilateral dorsal hippocampus. Quantification showed significant increases in GFAP in both the ipsilateral and contralateral parietal cortex, thalamus, and hippocampus (Fig. 7F). Although ipsilateral entorhinal cortex and amygdala were too damaged for analysis, quantification showed significant increases in GFAP in both contralateral structures (Fig. 7G). The GFAP-immunoreactivity of astrocytes following tlCont was more prominent than that of quiescent GFAP-positive control cells (Fig. 7H).

Astrocyte activation following tlCont.
Iba1 immunoreactivity on day 7 was used to assess microgliosis. The pattern of findings with Iba1 resembled that with GFAP. Compared with controls, Iba1 immunoreactivity was widely increased after tlCont, more so on the side of injury but also contralaterally (Fig. 8A,B). Compared with controls, Iba1 immunolabeling was greatly increased in both the ipsilateral and contralateral hippocampus (Fig. 8C–E). Quantification showed significant increases in Iba1 in both the ipsilateral and contralateral parietal cortex, thalamus, and hippocampus (Fig. 8F). Again, ipsilateral entorhinal cortex and amygdala were too damaged for analysis, but quantification showed significant increases in Iba1 in both contralateral structures (Fig. 8G). The plump-activated morphology of the microglia following tlCont was distinct from that of quiescent Iba1-positive control cells (Fig. 8H).
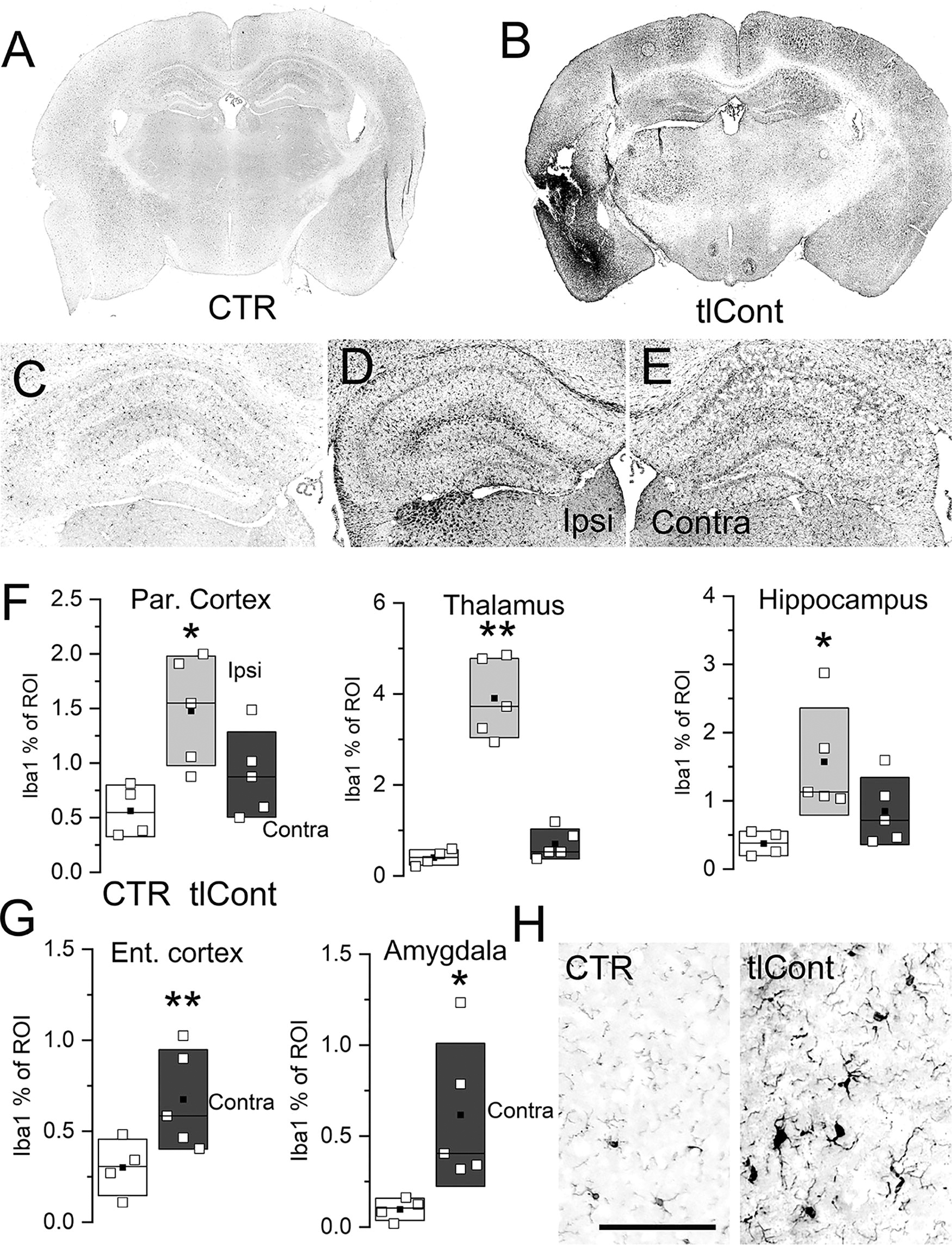
Microglial activation following tlCont.
We also evaluated GFAP and Iba1 immunoreactivity in the corpus callosum on day 7. Both markers were upregulated in the ventral portion of corpus callosum (Fig. 9), which contains fibers that carry projections between temporal lobe structures. 29 Immunolabeling for activated caspase-3 and β-amyloid precursor protein showed evidence of apoptosis and axonal degeneration in this portion of the corpus callosum (Fig. 9).
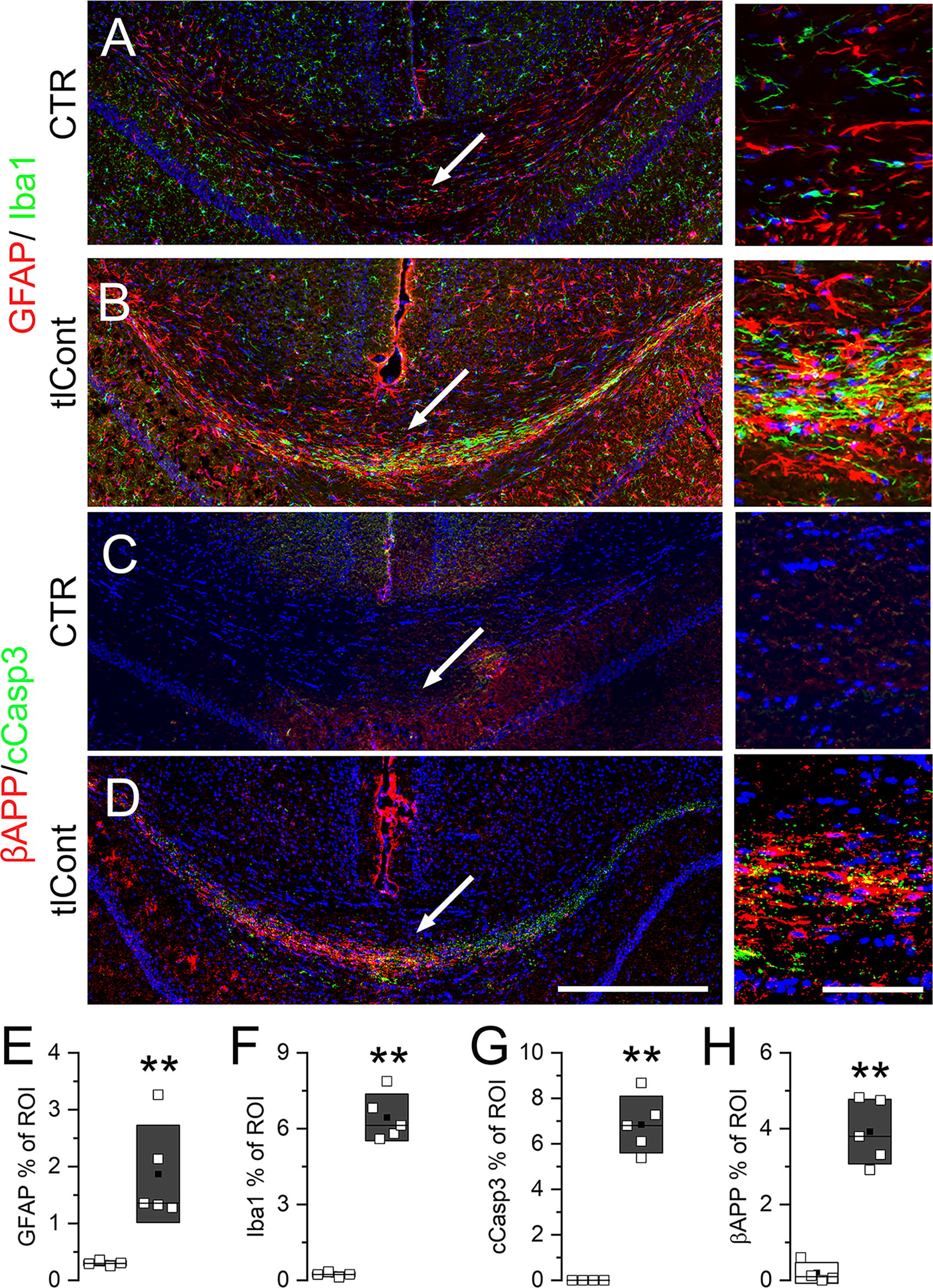
Pathological changes in the corpus callosum following tlCont.
We evaluated parvalbumin immunoreactivity, which identifies a subgroup of GABAergic inhibitory interneurons. 30 When examined 6 weeks following tlCont, both ipsilateral and contralateral dorsal hippocampi showed no overt evidence of injury but markedly fewer parvalbumin-positive cells compared to controls (Fig. 10A). Although ipsilateral temporal cortex could not be studied due to tissue damage, similar losses in parvalbumin-positive cells was found in contralateral parietal and entorhinal cortices, compared with controls (Fig. 10B,C). Quantification showed significant decreases in parvalbumin neurons in all these structures, with the greatest loss in the ipsilateral dentate gyrus (Fig. 10D).
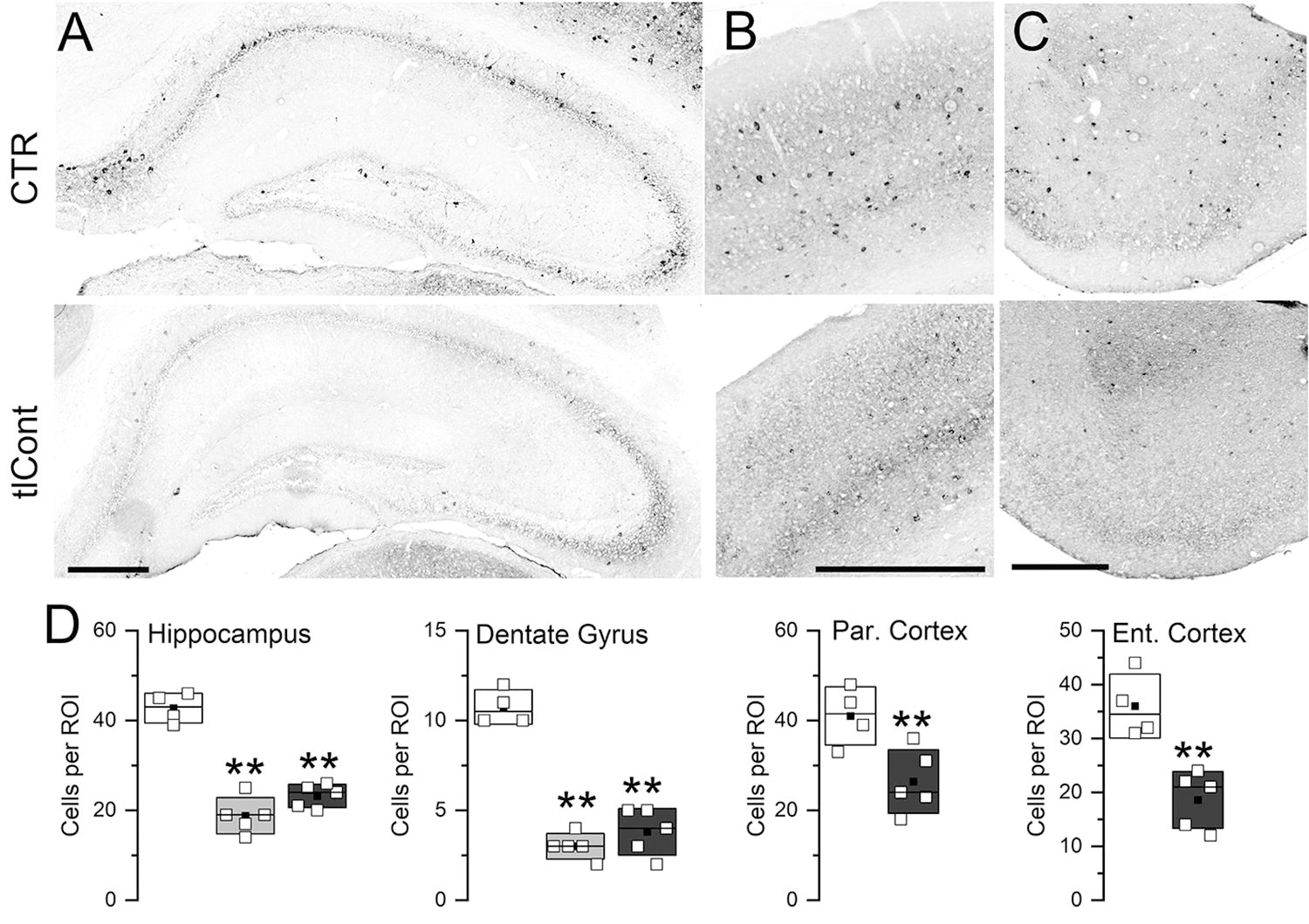
Parvalbumin neurons following tlCont.
Discussion
We describe the first rodent model that focuses specifically on tlCont. Most other models of focal TBI induce injury to the parietal lobe using CCI, 31 FPI, 9,32 or impact-acceleration, 12 with no stated intent of affecting the temporal lobes. Parietal lobe TBI models have been invaluable for elucidating numerous mechanisms of primary and secondary injury. 8 However, given the unique anatomical structures located within the temporal lobes, especially the limbic structures, conventional models focused on the parietal lobe may fail to reveal unique sequelae of injury identified in humans with tlCont. As reviewed in the Introduction, the experience in human TBI clearly shows that a CT-positive contusion in the temporal lobe is one of the strongest early biomarkers of poor neurological outcome and future post-traumatic epilepsy. The tlCont model we describe here may offer the ability to study these unique aspects of TBI.
The tlCont model was characterized by low mortality, transient sensorimotor abnormalities, and persistent deficits in anxiety and memory function. Transient sensorimotor abnormalities with prolonged memory impairment in the absence of hippocampal cell death was reported previously following midline FPI. 33 The dissociation of sensorimotor from memory abnormalities differs from TBI models that target the parietal lobe in which abnormalities in anxiety, memory, and sensorimotor function generally coexist. Severe (penetrating) parietal lobe CCI with a large lesion that encompasses much of the surrounding cortical tissue and ipsilateral hippocampus results in a similar degree of memory dysfunction (novel object recognition and Morris water maze) as the tlCont model but is accompanied by significant long-lasting sensorimotor abnormalities and high mortality (28%). 34 Co-association of sensorimotor and memory abnormalities also may be observed in murine models with impact acceleration. 35 These patterns of neurological dysfunction reflect well-known functional anatomy, point to the unique vulnerability of the temporal lobes, and underscore the unique neurological sequelae of tlCont.
Parvalbumin fast-spiking interneurons are GABAergic cells that comprise only a small fraction of the brain’s neuronal network, but drastically influence downstream signaling and thus ultimately affect multiple cognitive behaviors. 30 Parvalbumin-positive interneurons are highly represented in the hippocampal CA1 and CA3 regions as well as the dentate gyrus. 36,37 The hippocampal parvalbumin interneurons represent an especially vulnerable population of neurons in chronic stress, absent physical trauma, and their loss is considered to be of key importance in the development of depressive- and anxiety-like behaviors 38 as well as epileptogenesis. 39,40 The tlCont model was associated with major reductions in parvalbumin neurons in the ipsilateral and contralateral hippocampi as well as in contralateral cortex, along with significant anxiety-like behavior, as measured using the elevated plus maze, and spontaneous seizure activity, as detected by EEG. It may be that the trauma and its sequelae created a condition of chronic stress that was responsible for the loss of parvalbumin neurons and the accompanying anxiety-like behavior, although factors other than stress cannot be excluded. The loss of parvalbumin neurons also may have contributed to the increase in spontaneous seizure activity and the seizure responses to PTZ challenge observed in the tlCont model.
The tlCont model was characterized by widespread neuropathological changes, both ipsilaterally and contralaterally. We found astrogliosis and microgliosis in bilateral temporal lobes, thalami, hippocampi and other structures. Contralateral pathology as well as bilateral astrogliosis and microgliosis have been reported in CCI and FPI models. 7,34,41,42 In addition to upregulation of GFAP and Iba1, lateral FPI and CCI also reportedly cause bilateral upregulation of cyclooxygenase-2, 43 –45 a phenomenon not studied here.
Contralateral pathology that complicates ipsilateral damage likely has important functional consequences. In rodent studies on neuroanatomical function, it is established that the destruction or temporary inhibition of bilateral hippocampi and/or ento-/perirhinal cortices leads to major difficulties with learning and memory, including poor performance in novel object recognition 46,47 and Morris water maze. 47,48 Pathological involvement of one hippocampus is sufficient to adversely affect memory when the other hippocampus has been destroyed by trauma, 34 but not when the other hippocampus is spared. 49 When both hippocampi are largely intact, as in the tlCont model, bilateral neuropathological changes appear to predispose to significant underperformance in novel object recognition and Morris water maze.
The tlCont model was associated with apoptosis, 50 gliosis, and axonal degeneration in a discrete, ventrally-placed transcallosal fiber bundle that connects the temporal lobe cortices, 29 suggesting the presence of interhemispheric or transcallosal diaschisis. 51 –53 Diaschisis lesions were reported previously in the corpus callosum and contralateral cortex 7–21 days following parietal CCI. 54 Interhemispheric or transcallosal diaschisis is a poorly understood phenomenon that could be adaptive or maladaptive—contralateral changes that emerge may compensate for function that is lost by injury or it may worsen what is assumed to be a unilateral injury, transforming it into bilateral dysfunction. In humans, interhemispheric functional connectivity is reduced after TBI, 55 possibly reflecting diaschisis lesions. Further study of diaschisis in the tlCont model is warranted due to its relevance in human TBI.
The tlCont model also was characterized by EEG abnormalities in the hippocampus. In the cohort that we evaluated electroencephalographically, most mice showed nascent disturbances, including electrographic seizures and spikes that were more frequent than in controls and that persisted for 3 weeks. Given the location of the trauma with sparing of the sensorimotor cortex, it was not unexpected that the electrical disturbances would be nonconvulsive. Behavioral correlates were not observed but our recordings up to 3 weeks post-tlCont may have been too early for the electrical disturbance to manifest behaviorally. In their work on post-traumatic epilepsy after severe (penetrating) CCI to the parietal lobe, Golub and colleagues observed spontaneous recurrent seizures that developed in 87% of animals beginning with a mean latency of 53 ± 8 days. 34,56,57 Other investigators similarly have found that epileptiform activity takes a long time to develop post-TBI. 58 Future experiments with longer follow-up beyond 3 weeks may be needed to determine whether the EEG abnormalities observed in the tlCont model evolve eventually into worse electrical and behavioral disturbances.
The method used here to induce tlCont may be compared to methods used in conventional TBI models to induce focal or diffuse brain injury. A notable methodological innovation is the use of a Fogarty balloon catheter that is rapidly inflated/deflated epidurally to induce a brain contusion. However, the rates of impaction/de-impaction applied to the dura are 1000× slower and the area of impaction is larger in our model compared to conventional models. Nevertheless, the method replicates the traumatic interstitial hemorrhage typical of a contusion, prioritizing the characteristic hemorrhage over the usual fast biomechanics of injury. In humans, TBI is caused by a variety of forces with different biomechanical characteristics. The depth of impaction in CCI, the drop-height in impact-acceleration, and the force applied in FPI determine injury severity but otherwise, little empirical research has been conducted on the importance of most biomechanical factors including speed of impact in preclinical models. By convention, in the FPI and CCI models, the duration of the pressure transient or mechanical force applied to the dura is typically brief (∼20 msec), emulating the duration measured in human cadaver skulls during sudden impact. 9 Regardless of the biomechanical characteristics, what all models of focal injury have in common is that, if severe enough, they produce traumatic hemorrhages that evolve into necrotic lesions and eventual loss of tissue. In the clinical setting, the term “contusion” describes a spectrum of hemorrhagic lesions, regardless of the biomechanical characteristics of the force that caused the trauma. Here, our emphasis was not on implementing any specific injury-inducing biomechanical factor but on inducing a traumatic interstitial hemorrhage in the temporal lobe. Arguably, the presence of a traumatic hemorrhage, rather than the force that produced it, may be the most critical element that determines the tissue’s response.
The tlCont model has limitations, including some that are similar to those encountered in other pre-clinical models of focal TBI. Principal among these is the use of anesthesia during trauma, which could affect the response to trauma as well as the animal’s recovery from trauma. Also important is the requirement for a surgical craniectomy, although a 2-mm craniectomy is minimally invasive, is considerably smaller than in most other models, and is distant from the site of injury, precluding decompression of the contusion. The device we describe to rapidly inflate/deflate the Fogarty balloon in a reproducible manner to induce tlCont may seem complex, suggesting that the development of an alternative device may be warranted. As this is a new model of TBI, the specific details of the injury parameters may have to be changed as experience is gained and the full validity of the model will only become apparent after it has been reproduced by independent laboratories. The tlCont model did not exhibit hemorrhagic progression at 24 h post-trauma, whereas conventional CCI leads to a contusion that is typically accompanied by hemorrhagic progression during the hours following trauma. 28,59,60 This last difference may be due to the relatively slow inflation of the balloon, compared to CCI, and may have implications for therapeutic development.
In summary, we present our initial findings on a novel model of TBI that focuses on tlCont. Several features make this model especially appealing, including its direct relevance to a debilitating form of TBI in humans, the low mortality, significant disturbances in memory and anxiety with absence of persistent sensorimotor abnormalities, bilateral macro- and microglial responses, severe loss of hippocampal parvalbumin neurons, evidence of transhemispheric diaschisis, and prevalent EEG abnormalities. Given the clinical significance of tlCont in humans, the tlCont model may be a useful addition to the preclinical armamentarium. Additional work will be required to determine if this model recapitulates the unique spectrum of neurological and pathophysiological sequelae found in humans with tlCont.
Transparency, Rigor, and Reproducibility Summary
Surgeries and neurofunctional tests were performed by experienced investigators. Histochemical outcomes were obtained using objective methodologies with uniform parameters of tissue processing, image acquisition and analysis, including segmentation and stereology, by experienced investigators. Statistical analyses were conducted using Origin Pro V8 or GraphPad Prism 8.3 and included Student’s t-test, one-way ANOVA, or repeated measures ANOVA with Bonferroni post-hoc comparisons or Mann–Whitney U test, as appropriate. All tests were two-tailed with a statistical significance level set at p < 0.05.
Footnotes
Authors’ Contributions
J.M.S.: model conception, design and construction of the custom device for inflating/deflating the Fogarty balloon catheter, writing (lead), and funding; P.C., J.A.S., R.M.J.: consultation on the model, writing—review and editing (supporting); O.T., N.T.: animal surgery and husbandry; K.K., A.O., M.E.: neurofunctional tests; C.T.: immunohistochemistry, data analysis; M.M., J.L., Z.K., M.K., A.K.: surgery for electrode implantation, MATLAB script for EEG analysis, EEG recording, analysis; V.G.: data analysis, preparation of graphics.
Funding Information
This work was supported by an intramural pilot grant to A.K. and J.M.S. from the Department of Neurosurgery. J.M.S. is supported by a grant from the
Author Disclosure Statement
The authors have no competing interests to declare.