
Abstract
It is well documented that service members are exposed to repeated low-level blast overpressure during training with heavy weapons such as artillery, mortars and explosive breaching. Often, acute symptoms associated with these exposures are transient but cumulative effect of low-level repeated blast exposures (RBEs) can include persistent deficits in cognitive and behavioral health. Thus far, reliable diagnostic biomarkers which can guide countermeasure strategies have not been identified. In this study, rats were exposed to multiple field-relevant blast waves with 8.5 and 10 psi peak positive overpressures, applying one exposure per day for 14 consecutive days. micro-RNAs that can potentially be used as biomarkers for RBEs were assessed in blood, brain, and cerebrospinal fluid (CSF). RBE caused a differential pattern of changes in various miRNAs in blood, brain and CSF in an overpressure-dependent manner. Our key outcomes were decrease of mir-6215 and let-7 family miRNAs and increase of mir-6321 and mir-222-5p in brain, blood, and CSF. Expression pattern of these miRNAs is in concurrence with various neurological conditions such as upregulation of mir-6321 in focal ischemic injury and downregulation of mir-6215 in nerve injury model. Contrarily, Let-7 family miRNAs have neuroprotective role and their downregulation suggests progression of blast induced traumatic brain injury (bTBI) with RBE at 14× -8.5 psi. Repeated blast caused alterations in miRNAs that are likely involved in vascular integrity, inflammation, and cell death. These results indicate that miRNAs are differentially dysregulated in response to blast injuries and may represent better prognostic and diagnostic biomarkers than traditional molecules to identify blast-specific brain injury.
Introduction
Due to the increased use of improvised explosive devices and other explosives, blast-related mild traumatic brain injury (mbTBI) has become one of the most common types of injuries sustained by combatants in recent wars. 1 According to the U.S. Department of Defense, of the ∼380,000 TBIs reported in service members between 2000 and 2017, 82.3% were classified as mild (mTBI), 2 with the majority of these cases being attributed to blast exposure. 3 Unlike more severe TBI, mTBI is associated with normal structural imaging, brief or no loss of consciousness, and very rapid recovery of memory and mental state, as has been characterized in both pre-clinical and clinical studies. 4,5
Blast-induced TBI has been associated with long-term behavioral effects such as aggression, anxiety, cognitive deficits, depression, and suicide. 6 Similarly, it is well documented that cumulative effects of repeated blast exposures (RBE) can lead to behavioral and cognitive changes. 7,8 Oftentimes, these transient changes go unrecognized, but negative outcomes have been increasingly noted when an individual has record of RBE over their career. 9,10 Identifying specific diagnostic or prognostic biomarkers pertaining to RBE with adequate sensitivity and specificity can guide early intervention, potentially preventing the downstream pathophysiological mechanisms that can lead to long-term adverse outcomes.
Current standard blood-based biomarkers, including S100B, neurofilament protein (NFL), glial fibrillary acidic protein (GFAP), and ubiquitin C-terminal hydrolase L1 (UCHL-1), lack the required level of specificity to address the outcome of mTBI and are also not specific to blast-related brain injuries. 11 Recent studies have identified the potential utility of a class of biomolecules called microRNAs or miRNAs as biomarkers for mbTBI. miRNAs are small, noncoding RNA that regulate gene expression by silencing mRNA targets. 12,13 miRNAs are well-suited as biomarkers because they are stable, abundantly expressed, easily accessible, and have important biological functions including gene regulation and intercellular signaling. 12 Moreover, important characteristics of miRNAs as biomarkers include specificity to the cell type of origin. For instance, miRNAs differentially expressed in this study such as miR-128, miR-125, and miR-23 are examples of brain-enriched miRNA whereas miR-219 and miR-124 are examples of brain-specific miRNAs. 14 Other important characteristics of miRNAs as biomarkers include indications of the stage of the disease progression, and in the case of the brain, their ready accessibility since they easily cross the blood–brain barrier and can be identifiable in blood. 15 miRNAs can be isolated from bodily fluids such as blood, saliva, or cerebrospinal fluid (CSF), making measurements a simple minimally invasive and resource-effective procedure.
In this study, miRNA profiling was performed in a rat model following RBE to 14 daily 8.5 and 10 psi shock waves using an advanced blast simulator (ABS), which generates Friedlander-like shock waves. miRNA was profiled using next-generation sequencing (NGS) techniques in brain, CSF, and blood samples of rats. Our results indicate that various miRNAs are differentially expressed in brain, CSF, and blood following blast exposure, revealing their potential utility as biomarkers with RBE.
Materials and Methods
Experimental design
Material has been reviewed by the Walter Reed Army Institute of Research (WRAIR). There is no objection to its presentation and/or publication. The opinions or assertions contained herein are the private views of the author and are not to be construed as official, or as reflecting true views of the Department of the Army or the Department of Defense. Research was conducted under an IACUC-approved animal use protocol in an AAALAC International-accredited facility with a Public Health Services Animal Welfare Assurance and in compliance with the Animal Welfare Act and other federal statutes and regulations relating to laboratory animals. Male Sprague Dawley rats weighing 300–350 g (Charles River Laboratories, Wilmington, MA) were housed in group at 20°C–22°C (12 h light/dark cycle) with free access to food and water ad libitum. Animals were randomly divided into groups of 12 each for sham, 8.5 psi and 10 psi exposure groups.
Repeated blast exposure
Rats were anesthetized with isoflurane and subjected to survivable blast overpressures (BOPs) using an ABS located at the WRAIR. The ABS consists of a 0.5-ft-long compression chamber that is separated from a 21-ft-long transition/expansion test section. 16 Exposures in this study closely reflect the exposures in the training environment similar to breaching operations or shoulder-fired weapon systems (LAW or Carl Gustaf). 17 The anesthetized rat was secured in the test section in “off-axis,” that is, transverse orientation to the direction of blast. The compression chamber was pressurized with room air, causing membranes to rupture at a pressure that is dependent upon the thickness of the specific membrane sheet separating the two chambers, yielding a supersonic blast wave that impacts the experimental subject in the test section. To minimize polyorgan injury in these experiments, acetate membranes (Grafix Inc., OH, USA) were used to yield peak positive static pressures of 8.5 psig (positive pressure-impulse: 11.46 psig*ms, duration: 2.94 ms; negative pressure-impulse: −8.35 psig*ms, peak: −2.86 psig, duration: 5.89 ms), which has been identified as threshold for lung contusion for up to 30 daily exposures or 10 psig (positive pressure-impulse: 16.64 psig*ms, duration: 4.0 ms; negative pressure-impulse: −10.27 psig*ms, peak: −2.34 psig, duration: 7.07 ms) with a positive phase duration of 3–5 ms) multiple daily exposures to which caused around 1% lung injury as described in Heyburn et al. 18 In addition, low-level repeated exposures such as this simulate the military training environment which may lead to cumulative effect on brain pathophysiology. Animals (n = 12 per group) were exposed to a daily blast of 8.5 or 10 psi for 14 consecutive days; RBEs were separated by 24 h. All sham animals were subjected to identical isoflurane anesthesia, loading in the shock tube, and recovery procedures as described above, but were not exposed to BOP.
Brain, blood, and CSF collection
Twenty hours following final BOP exposure, animals were deeply anesthetized until the toe-pinch reflex was absent, euthanized by cardiac exsanguination and perfused with physiological saline followed by thoracotomy to ensure euthanasia. Blood was collected by cardiac puncture and stored in BDTM PAXgene blood RNA tubes for further analyses (Fisher Scientific, catalog # 23-021-01). The animals were then perfused with physiological saline and CSF was collected from the cisterna magna. Following blood and CSF collection, brain was divided into two hemispheres and flash-frozen on dry ice until further analysis.
Next-generation sequencing for miRNA
RNA extraction was performed using Exiqon miRCURYTM RNA isolation kit-Biofluids (Qiagen, Hilden, Germany) using manufacturer’s protocol. Sequencing sample preparation was performed using TruSeq Small RNA Library Preparation Kit (Illumina, San Diego, CA, USA) protocol. Libraries were multiplexed and gel purified for size selection. Sequencing was performed on illumina HiScanSQ platform using single read flow cell with 51 cycles for Read 1 and 8 cycles for index identification.
The sequence reads were first demultiplexed using bcl2fastq v 2.20 and reads were further quality filtered and adaptor trimmed using “cutadapt” software v1.16. Reads were further mapped against Rattus norvegicus (rn5, GCA_000001895.3) assembly for reference genome using Bowtie v 2.2.3.0 19 aligner. Mapped reads were further mapped to miRBase 20 -known miRNAs database for Rattus norvegicus (rn5, GCA_000001895.3) using miRDeep2 core algorithm for miRNA detection and de novo miRNA prediction. 21 The known miRNA quantification was done using miRDeep2 quantification function. Count data were further analyzed using edgeR V 3.6R bioconductor package. 22 A number of identified miRNA were filtered with less than 5 read counts (cpm >0.01) in 86% of sample population. Counts were further normalized by trimmed mean of M-values normalization method after library size factor calculations. 23 Sample library size distribution and per sample miRNA read counts matrix were generated to see overall quality of the data. For the identification of differentially expressed miRNA, the normalized data were further analyzed using empirical Bayes quasi-likelihood F-tests using contrast function. The DE miRNAs were identified using p value cutoff <0.05.
Results
miRNAs differentially expressed in brain
miR-6215 and miR-6321 were differentially expressed in brain at both 14×-8.5 psi and 14×-10 psi (Fig. 1). miR-6215 was significantly decreased with log fold change (FC) value of −2.75 (p = 0.02) at 14×-8.5 psi and −2.69 (p = 0.03) at 14×-10 psi. Differently, miR-6321 was significantly increased at both pressures with log FC value of 7.22 (p = 0.005) at 14×-8.5 psi and 6.048 (p = 0.01) at 14×-10 psi. The log FC, log CPM, and p values are listed in Table 1.

Differentially expressed miRNAs in hemi-brain at 14×-8.5 psi and 14×-10 psi BOP exposure. The bars represent significant (p < 0.05) fold change in the blast groups when blast groups are compared to sham. BOP, blast overpressure.
Log FC, Log CPM, and p Values of Differentially Expressed miRNA in Brain
miRNAs in CSF and blood at both exposures
Differentially expressed miRNA changes common to both CSF and blood at 14×-8.5 and 14×-10 psi included miR-200c-3p, miR-222-5p, miR-223-5p, miR-185-5p, and miR-3559-3p (Fig. 2). miR-200c-3p and miR-222-5p were significantly increased and miR-3559-3p was significantly decreased in both CSF and blood at 14×-8.5 and 14×-10 psi. miR-223-5p was significantly increased in CSF and significantly decreased in blood at 14×-8.5 psi and 14×-10 psi. In contrast, miR-185-5p was significantly decreased in CSF and increased in blood at both pressures. The log FC, log CPM, and p values are listed in Table 2.

Differentially expressed miRNAs in CSF and blood at 14×-8.5 psi and 14×-10 psi BOP exposure. The bars represent significant (p < 0.05) fold change in the blast groups when blast groups are compared to sham. CSF, cerebrospinal fluid.
Log FC, Log CPM, and p Values of Differentially Expressed miRNA in CSF and Blood
CSF, cerebrospinal fluid.
miRNAs changes in CSF and blood only at 14×-8.5 psi
Differentially expressed miRNA changes that were common to both CSF and blood at 14×-8.5 psi but not 14×-10psi pressure included miR-296-5p, miR-32-3p, and miR-6321 (Fig. 3). miR-296-5p and miR-6321 were significantly increased and miR-32-3p was significantly decreased in both CSF and blood. The log FC, log CPM, and p values are listed in Table 3.

Differentially expressed miRNAs in CSF and blood only at 14×-8.5 psi BOP exposure. The bars represent significant (p < 0.05) fold change in the blast groups when blast groups are compared to sham.
Log FC, Log CPM, and p value of Differentially Expressed miRNA on Blood and CSF at 14×-8.5 psi
miRNAs in CSF and blood only at 14×-10 psi
Differential expressions of miRNA that are common to both CSF and blood at 14×-10 psi pressure but not brain were miR-144-3p, miR-182, miR-298-3p, miR-3075, miR-32-3p, miR-350, miR-3542, miR-3542, miR-3558-5p, miR-3572, and miR-362-5p (Fig. 4). miR-144-3p was increased in CSF and decreased in blood. miR-32-3p was decreased in both CSF and blood. All other above-listed miRNAs were increased in both samples. The log FC, log CPM, and p values are listed in Table 4.
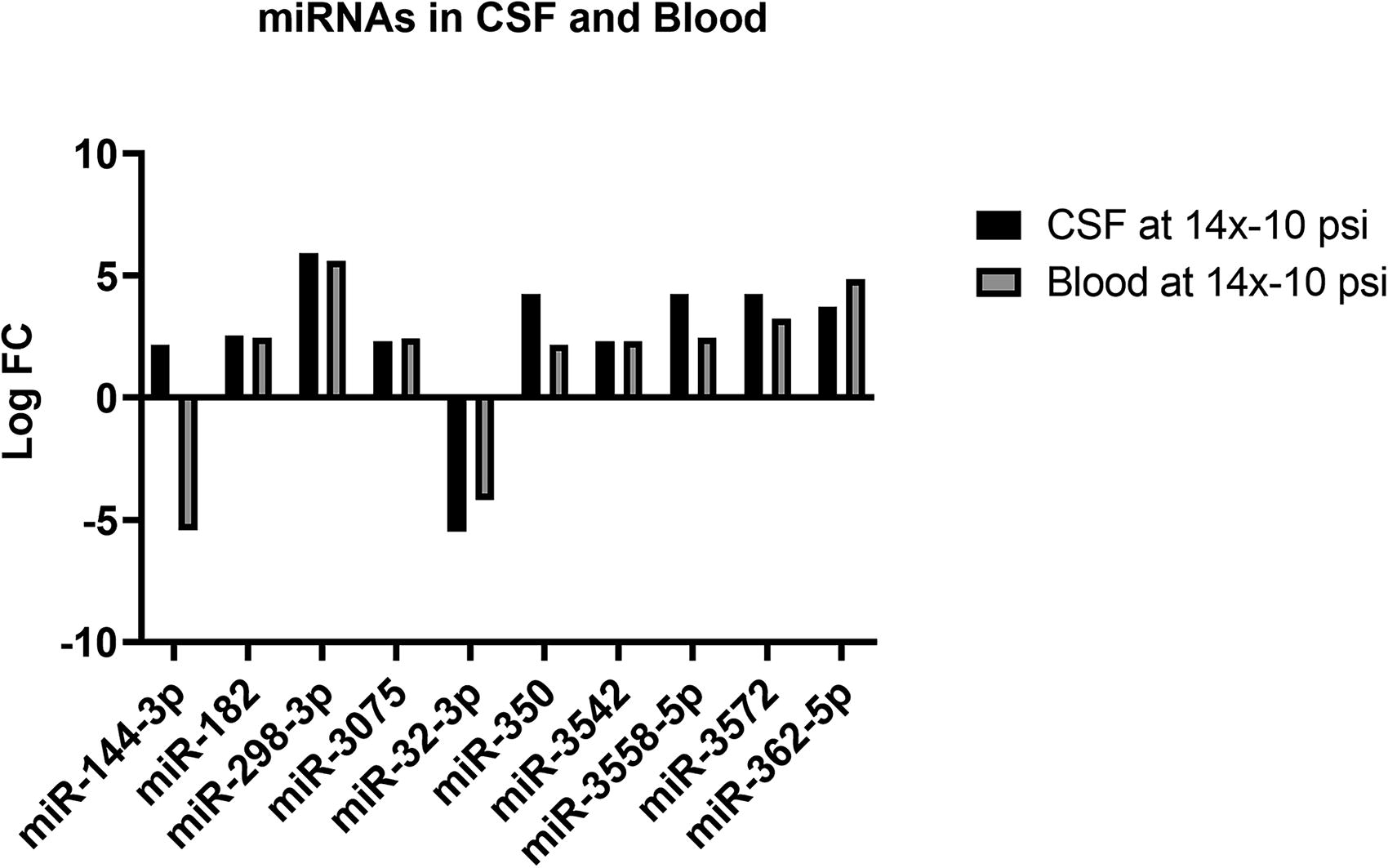
Differentially expressed miRNAs in CSF and blood only at 14× 10 psi BOP exposure. The bars represent significant (p < 0.05) fold change in the blast groups when blast groups are compared to sham.
Log F, Log CPM and p value of Differentially Expressed miRNA in CSF and Blood at 14×-10 psi
miRNA differentially expressed only in blood
miR-6215 was significantly increased in blood with log FC value 3.39 (p = 0.006) at 14×-8.5 psi when compared to sham.
Differential expression exclusive to each sample
Table 5 provides the number of miRNAs differentially expressed in CSF and blood samples after sorting out all the commonalities as stated in the Results section. The detail of miRNAs with fold change and p value is provided in Supplementary Data S1.
Number of Significantly Increased and Decreased miRNAs in CSF and Blood
Discussion
miRNAs are small noncoding RNA molecules with ∼22 nucleotides that act as regulators of post-transcriptional processes affecting gene expression and RNA production. These molecules have been shown to be highly stable and resistant to degradation which makes them attractive candidates for disease biomarkers. 24 –26 Studies have also shown their potential as biomarkers in Alzheimer’s disease, 27 Parkinson’s disease, 28 and epilepsy 25 among many other neurological disorders. Recent studies have shown that expression of miR-1, miR-22, and miR-29a can potentially be used to differentiate healthy patients from nontreated patients with Parkinson’s disease. 29
Since pathophysiology of bTBI is very complex, specific blood- or tissue-level diagnostic markers for bTBI have not yet been identified. Recently, proteins such as GFAP, UCHL-1, and NFL have been considered as markers of TBI, 30 –33 including our own study which has shown changes in GFAP in brain at 14×-10 psi but not in 14×-8.5 psi. 18 However, these markers have been only routinely used for decision-making whether to perform CT/MRI scans on patients with concussions. 34 In addition, these markers are not specific to blast-related TBIs, which have characteristically different mechanisms of injury. 35 miRNAs on the other contrary are highly conserved among species, which makes them highly translatable for clinical use. Our previous exploratory study using a mouse model of relatively high-intensity BOP exposure (17 and 20 psi) also suggested the alteration of brain-specific and brain-enriched miRNA in the blood plasma samples unique to number and intensity of exposures. 13 Therefore, in this follow-up study, we conducted a comprehensive measurement of miRNAs in the brain tissue, CSF, and blood samples that were collected from animals exposed to RBE (14×-8.5 psi and 14×-10 psi) using the NGS technique.
The markers uniquely originating in the brain that are measurable in CSF or blood have a higher prognostic utility as TBI biomarkers. 29 Specifically, miR-6215 was significantly decreased and miR-6321 was significantly increased in brain tissue samples from both exposure groups in association with similar changes in blood and CSF (Fig. 2). The potential biological role of miR-6321 following RBE largely remains unclear; however, it was shown to be upregulated in focal cerebral ischemic injury 36 and in animals exposed to transcranial stimulation, 37 and has also been implicated in the regulation of endothelial progenitor cell growth and migration by regulating c-Jun N-terminal kinase (JNK) pathway. 38 Interestingly, downregulation of miR-6215 was associated with neuropathic pain mediated by FosB in a spared nerve injury model. 39 Downregulation of miR-6215 can also activate microglia and contribute to inflammatory cascade through a FosB-mediated mechanism. 40,41 In the present study, miR-6321 was significantly increased in CSF and blood at 14×-8.5 psi (Fig. 1A) but similar changes were not observed in either at 14×-10 psi. Similarly, miR-6215 changes were not detected in CSF at both exposure conditions and increase in blood was observed only at 14× 8.5 psi (Fig. 1B). This could perhaps be indicative of inflammation progression and altered cellular signaling following 14×-8.5 psi RBEs. Notably, miR-6321 was shown to regulate signaling pathways involved in neurobehavioral outcomes such as anxiety/depression, a condition highly prevalent in the clinical setting from personnel exposed to blast. 31,42,43
When these data are compared to those of our previous study in mice employing higher psi overpressure exposures, 13 the differential expression of few specific miRNAs was consistently observed, such as miR-222-5p, which was consistently upregulated (Fig. 3), and let-7a-5p, let-7b-5p, and let-7c-5p which were consistently decreased in plasma in both studies. In addition, these markers were altered in CSF irrespective of the BOP intensity or the number of exposures in the present study. Upregulation of miR-222-5p has been implicated in accelerating vascular dysfunction and lipid deposition by targeting RB1. 44 Since altered lipid metabolism characteristically occurs in central nervous system (CNS) injuries, 45 miR-222-5p could also be an objective indicator of blast injuries, although a specific mechanistic role has yet to be established. Let-7 miRNAs are abundantly expressed in brain and regulate microglial function through TLR-7. 46 Let-7 family of miRNAs also play a crucial role in cell survival by regulating c-Myc and Ras pathways, which by mediating the p53 pathway, regulate apoptosis and cell death. 47 More specifically, let-7a-5p has been shown to promote neuronal growth in spinal cord-injured rats, 48 let-7b-5p is shown to have dual neuroprotective roles in multiple sclerosis 49 and memory problems, headache, and fatigue in postconcussive patients, 50 and let-7c-5p is also known to improve the neurological outcome in murine model of TBI by suppressing inflammation. 51 Since the overall role of these particular let-7 miRNAs is generally indicative of improving neurological outcomes, it potentially indicates that downregulation of these let-7 miRNAs points to brain injury evolution and negative outcomes associated with blast. In addition to these changes, miRNAs miR-200c-3p, 223-5p, 185-5p, and 3359-3p were differentially expressed in both CSF and blood in both exposure groups (Fig. 3). Although miR-200-c-3p has been reported to induce endothelial apoptosis and vascular disruption by promoting NaF transcription factor 52,53 and miR-223-3p, and miR-185-5p have been shown to act as negative regulators of inflammation by inhibiting NLRP3 inflammasome. 54,55 Interestingly, these miRNAs were unaltered in our previous study conducted with higher psi BOPs but were significantly altered with RBE at 14×-8.5 or 14×-10psi. 13 Based on these data, it can be speculated that a specific subset of miRNAs can be used to differentially diagnose brain trauma associated with various intensities of blast exposure. This evidence is further compounded by differential expression of miRNAs in CSF and blood such as miR-296-5p, miR-32-3p, and miR-6321, which were exclusive to the 14×-8.5 group, whereas upregulation of miR-182, miR-298-3p, miR-307, miR-350, miR-3542, miR-3542, miR-3558-5p, miR-3572, and miR-362-5p was exclusively observed in experimental subjects exposed to 14×-10 psi (Fig. 4). Among these miRNAs, miR-182 has been shown to cause mitochondrial dysfunction and its inhibition had protective effects in ischemia/stroke pre-clinical models. 56 Similarly, miR-298-3p has been protective against Aβ-deposition and Tau phosphorylation. 57
Overall, differential alterations observed in several miRNA profiles in two different subthreshold level RBE conditions collectively suggest that these molecules are sensitive to the intensity of BOP and potentially can be used for differential diagnosis of exposure effects of RBE or single BOP at varied intensities and can additionally differentiate tissue of origin. The majority of miRNAs altered in the present study are associated with vascular rupture along with systemic- and neuroinflammation, which can both appreciably influence the resulting behavioral changes. Taken together, these miRNA profiles associated with experimental RBE are suggestive of potentially promising blast injury diagnostic and prognostic markers that await validation in with clinical samples.
Transparency, Rigor, and Reproducibility Statement
All the institutional guidelines were followed in accordance with Department of Defense policies. We have meticulously documented our methodologies including statistical analysis to enable replication. For reproducibility, we will provide all the necessary resources other than those included in this document upon request.
Footnotes
Acknowledgments
The authors would like to acknowledge Ms. Irene Gist and Mr. Tung Tu for administrative support.
Authors’ Contributions
S.D.: Organized data, plotted figures, and drafted article. R.R.V.K., L.H., and D.W.: Performed blast experiments and specimen collection. R.K.: Performed miRNA data analysis. G.D.: Performed miRNA quantification. A.G. and R.H.: Supervised miRNA quantification and data analysis. J.B.L. and V.S.S.: Designed the study and contributed to writing.
Author Disclosure Statement
The author(s) have no competing interest to disclose.
Funding Information
This research was funded by the Military Operational Medicine Research Program, Medical Research and Development Command, United States Department of Defense, grant number
Supplementary Material
Supplementary Data S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.