
Abstract
Traumatic brain injury (TBI) after high-energy, behind helmet blunt trauma (BHBT) is an important but poorly understood clinical entity often associated with apnea and death in humans. In this study, we use a swine model of high-energy BHBT to characterize key neuropathologies and their association with acute respiratory decompensation. Animals with either stable or critical vital signs were euthanized within 4 h after injury for neuropathological assessment, with emphasis on axonal and vascular pathologies in the brainstem. The majority of cases were characterized by fractures of the cranium at or about the impact site, extensive subarachnoid hemorrhages, coup and contrecoup contusions, and primarily diffuse axonal and vascular lesions throughout the cerebrum, particularly in the brainstem. Absence of spontaneous respiration that was encountered frequently was associated with both severity of impact and the severity of brainstem axonal and vascular lesions. A focused regional examination of brainstem pathology indicated a link between adverse outcomes and diffuse axonal lesions within the medial medulla or vascular lesions within the anteroventral brainstem, a pattern suggesting that injury to brainstem respiratory centers may play a role in apnea following BHBT. In addition, while the overall burden of diffuse axonal and vascular pathologies correlated with each other, we found minimal overlap in their regional distribution. Our findings indicate that high-energy, blunt-force impact TBI causes diffuse lesions in axons and blood vessels associated with poor outcomes. They also suggest that axons and vessels may have distinct responses to tissue deformation and that commonly used markers of vascular pathology, for example, in diagnostic radiology, cannot be used as direct surrogates of diffuse axonal injury. In concert, our study underscores the role of regional axonal and vascular injuries in the brainstem in acute respiratory decompensation after high-rate blunt TBI, even in the presence of head protection; it also emphasizes the importance of detailed clinicopathological work in complex brains in the field of TBI.
Introduction
Blunt-force traumatic brain injury (TBI) associated with impact loading is by far the most common TBI scenario in civilian and military populations regardless of specific cause, including falls, motor vehicle collisions, struck-by-or-against accidents, assaults, tackling in collision sports, and ballistic insults. 1,2 Even in cases where there is prominent angular acceleration, head impact is regularly present. A major exception may be primary blast TBI, but tertiary blast injury that often co-occurs with primary blast does involve impact. Pure impulse (angular acceleration) loading as cause of TBI has been largely based on animal modeling 3 –6 that has contributed significant biological advances in the dissection of mechanisms, but does not represent the majority of TBI encountered in civilian, sports-related, and operational or combat situations. In addition, the commonly implied double dissociation between impact versus impulse (acceleration) loading on the one hand and focal versus diffuse TBI on the other, that is, the notion that impact loading causes focal TBI (contusions) and impulse loading causes diffuse axonal and vascular injury, is rather simplistic: even classical articles 7 –9 have touted the significant role of impact loading or translational movement in diffuse brain lesions. This insight is consistent with previously published data from our group showing that blunt-force impact TBI with limited rotational acceleration can indeed cause both diffuse vascular and axonal lesions. 10 –12
These notions are extremely important because in several cases of blunt-force TBI in civilian, sports-related, and operational or combat scenarios, there is an acute decompensation of vital signs attributed, among else, to dysregulated sympathetic activity 13,14 secondary to disconnection and loss of inhibitory control, although the precise mechanisms are still unclear. Therefore, clarifying mechanisms by which blunt force-related impact loading causes diffuse and nondiffuse TBI of all types is of enormous importance, potentially leading to better primary prevention with head and body protection or secondary prevention with drugs.
In the present study, we use brain tissues from swine exposed to high-energy behind helmet blunt trauma (BHBT), a form of high-energy blunt-force TBI. To the best of our knowledge, this is the first neuropathological characterization of this type of injury in a large animal model. First, we outline the main focal and diffuse pathologies, and then we engage in a more comprehensive study of diffuse axonal and vascular lesions in the brainstem to establish associations between regional pathology and vital sign decompensation shortly after injury. In addition, we explore potential mechanisms of vital deterioration in the first few hours after BHBT.
Materials and Methods
Procurement of tissues and experimental history
Minipig brains from the Army DEVCOM Chemical Biological Center were received in our laboratory in 10% formalin. Brains were obtained from an initial cohort of male Hanford miniature swine (n = 6) and a second larger cohort of male Yucatan swine (n = 38) at the age of 6–8 months. To obtain representative kinematics to humans, the skull thickness, the extent of the frontal sinus, and complete suture fusion are important features to consider in an animal model. Hanford pigs were initially selected because they demonstrated skull thicknesses similar to human frontal bones with relatively small sinus cavities. However, it was later discovered that they have incomplete fusion of the sutures at the age in which the desired skull thickness is reached, resulting in unrepresentative skull dynamics. Therefore, subsequent, more focused studies were performed in Yucatan pigs in whom the range of skull thickness and suture development are more representative of the adult human skull. 15 –18
Animals in both cohorts had been anesthetized with either a continuous infusion of propofol or with isoflurane inhalation for the duration of the injury and postinjury period until euthanized. The injury consisted of a variable ballistic impact to the helmeted head, with the head secured on a flat table. The impact was over the midline, approximately 15 mm anterior to the bregma and at a slight angle (<18 degrees) relative to a line perpendicular to the flat part of the frontal bone over the bregma. The impact line was determined based on preimpact X-ray, and in some animals requiring instrumentation (see below) bregma was directly visualized. The location of bregma was confirmed in all cases by postimpact computed tomography (CT). Information related to helmet and impact specifications cannot be disclosed due to their sensitive nature related to the U.S. Department of Defense. However, a group of subjects (n = 22) were fitted with instrumentation that allowed the estimation of impact severity in terms of helmet back-face velocity, which ranged from 180 to 360 m/s. Immediately after injury, the animals were checked for spontaneous respiration to decide on the need for further ventilator support. Based on this need, cases were categorized as either “critical” (no resumption of spontaneous respiration post trauma) or “non-critical” (resumption of spontaneous respiration).
In a subcohort of animals (n = 15), a DSI PhysioTel M11 digital telemetry implant (Data Sciences International, USA; measuring 40 × 26 × 14 mm; weighing 13.7 g) was used to monitor intracranial pressure (ICP), body temperature, electroencephalogram, and animal activity (derived from acceleration). This telemetry unit was installed on the day of the impact. The primary unit was placed within a subcutaneous pocket on the left side of the neck. The ICP catheter was subcutaneously tunneled to an incision made on the head and, using a stereotaxic frame and drill, was placed in the left ventricle. Heart rate and oxygen saturation were monitored using a pulse oximeter connected to the tail.
At necropsy, crania were carefully inspected for presence, as well as location, extent, and other features of fractures. To allow comparisons, binary scores of fractures were tabulated for each case based on presence or absence of fractures in orbital, frontal, parietal, temporal, and occipital bones, and a composite score was calculated as the sum of individual scores per case. Data on skull and skin thickness approximately at the impact location were also collected using CT scans.
Animals were transcardially perfused with 4% neutral-buffered formaldehyde 1.5–4 h postinjury. Two uninjured healthy Yucatan pigs were included as naive controls.
Mapping of subarachnoid hemorrhages
Brains were photographed from the top, bottom, and the two sides, and subarachnoid hematomas, present in the vast majority of cases, were traced from photographs using image processing (n = 36). To assess severity of subarachnoid hemorrhages (SAHs), two investigators (V.E.K., P.F.) independently categorized SAH as “Extensive,” “Moderate,” or “Mild-None” based on overall severity of hematomas; differences of opinion were discussed to reach consensus. To further characterize the distribution and extent of SAH for each case, a semi-quantitative score was assigned (0,1,2) to indicate, respectively, the absence of SAH and limited or extensive SAH in frontal, temporal, basal, and brainstem surfaces of the brain. The two control brains had no evidence of SAH.
Histochemical and immunohistochemical processing
Brains from the Yucatan cohort were selected for detailed neuropathological analysis (n = 22; including all 8 critical cases and 14 randomly selected noncritical cases). With the aid of a coronal swine brain slicer matrix (Cat. No #PBS050-1, Zivic Instruments, Pittsburgh, PA), brains were blocked at the coronal plane in 1 cm slices, except brainstem and genu-anterior segment of corpus callosum that were blocked in 5 mm slices. Based on placement in the brain matrix, brainstem slices extended from the thalamo-mesencephalic border to caudal medulla. Genu-anterior corpus callosum slices were blocked at a level corresponding to the anterior body of the fornix. Brainstem and callosal blocks were manually embedded in paraffin. Brains from the Hanford cohort were blocked in a similar manner, but in most cases caudal brainstem was not available. Along with some cases from the Yucatan cohort, these cases were the main source of tissues for the qualitative pathological analysis in the second section of the Results.
Paraffin-embedded blocks through the forebrain and brainstem were cut (10 μm) and serially stained for the following: cytoarchitectonics (Cresyl Violet); combined cyto-and myelo-architectonics (Cresyl Violet and Luxol Fast Blue-Cresyl Violet [Klüver–Barrera]); vascular lesions (Masson’s trichrome); and axonal lesions (amyloid precursor protein [APP] immunohistochemistry). APP immunohistochemistry utilized a rabbit polyclonal antibody against the C terminus of APP (CT695; 51–2700 from Invitrogen, AB_2533902 at 1:250). Paraffin sections were deparaffinized by incubation at 60°C for 30 min followed by sequential treatments with xylene and rehydrated in series of 100%, 90%, 75%, and 50% ethanol in dH2O. Endogenous peroxidase activity was blocked with 1% hydrogen peroxide. Sections were permeabilized and blocked with 0.3% Triton-X and 3% normal goat serum in phosphate buffered saline (PBS) and incubated overnight with the primary antibody at 4°C. Immunoreactive epitopes were visualized with a polymer-based HRP Kit (Super SensitiveTM Polymer HRP Kit, Cat No. QD430-XAKEN BioGenex, Fremont, CA) as per manufacturer’s instructions. Sections were dehydrated, cleared, and mounted with DPX Mountant (#13512, EMS, PA).
Quantitation of diffuse axonal and vascular pathology in the brainstem
APP-stained sections through 6 standard brainstem planes (Fig. 1, Supplementary Figs. S1, S2, and S3) were photographed at low magnification and, with the aid of adjacent cresyl violet-stained sections through the brainstem, divided into 116 predominantly gray or predominantly white matter regions. These maps were then used as guide for stereological analysis of axonal pathology per region of interest (ROI) as per next section. Quantitation of axonal injury utilized a fractionator probe applied in each separate ROI. Region boundaries were traced at 10× using an Axiophot microscope (Carl Zeiss Inc.) equipped with a motorized stage and Stereo Investigator© software (Microbrightfield Inc., Williston, VT). APP (+) lesions were counted at 20× magnification with the following counting parameters: counting frame size of 294 × 175 μm, grid size of 588 × 350 μm, and sampling fraction of 25%. Burden of axonal pathology was expressed per gray or white matter region by calculating the density of APP(+) lesions per ROI per section and side (left or right) and then averaging numerical scores per ROI per case. For example, the primary sensory nucleus of the fifth nerve (SNV) was encountered on both sides over more than one section, and a weighted average was calculated as the sum of total estimated lesions over the total area of ROIs belonging to the nucleus.

Six standard coronal planes of the swine brainstem containing most of the white and gray matter regions of interest (ROIs) used to assess vascular and axonal pathologies. AMB, nucleus ambiguus; bsc, brachium of the superior colliculus; cf, cuneate fasciculus; CG, central gray; cic, commissure of the inferior colliculus; CN, cochlear nuclei; cp, cerebral peduncle; cst, corticospinal tract; DM/PF, dorsomedial/parafascicular thalamic nucleus; DNX, dorsal motor nucleus of the vagus; DR, dorsal raphe; eml, external medullary lamina of thalamus; fr, fasciculus retroflexus; IC, inferior colliculus; icp, inferior cerebellar peduncle; II, lateral lemniscus; III, oculomotor nerve; ION, inferior olivary nucleus; IPN, interpeduncular nucleus; IRN, intermediate reticular nucleus; LG, lateral geniculate; LP, lateral posterior thalamus; LRN, lateral reticular nucleus; LVN, lateral vestibular nucleus; mcp, middle cerebellar peduncle; MG, medial geniculate; MgRN, magnocellular reticular nucleus; MH, medial habenula; LH, lateral habenula; ml, medial lemniscus; mlf, medial longitudinal fasciculus; MNV, mesencephalic nucleus of fifth, trigeminal nerve; MRN, mesencephalic reticular nucleus; mrV, motor root of the fifth nerve; MVN, medial vestibular nucleus; NIII, nucleus of third, oculomotor nerve; NP, nucleus prepositus; NTS, nucleus of the solitary tract; NVI, nucleus of the sixth, abducens nerve; NVII, nucleus of the seventh, facial nerve; NXII, nucleus of the twelfth, hypoglossal nerve; ot, optic tract; ParRN, parvicellular reticular nucleus; PBN, parabigeminal nucleus; pc, posterior commissure; PN, pontine nucleus; PRN, pontine reticular nucleus; PulV, pulvinar; py, pyramidal tract; RET, reticular formation; RN, red nucleus; RM, raphe magnus; RPHE, median raphe; SC, superior colliculus; scp, superior cerebellar peduncle; SNC, substantia nigra pars compacta; SNR, substantia nigra pars reticulata; SNV, sensory nucleus of the fifth nerve; SON, superior olivary nucleus; SpNV, spinal nucleus of the fifth nerve; sptv, spinal tract of the fifth nerve; SpVN, spinal vestibular nucleus; srV, spinal root of the fifth nerve; tb, trapezoid body; V, motor nucleus of the fifth nerve; VCN, ventral cochlear nucleus; VI, sixth nerve; VII, seventh nerve; VIII, eighth, vestibulocochlear nerve; VIIIv, vestibular branch of the eighth nerve; VRM, vermis; vsct, ventral spinocerebellar tract; VTA, ventral tegmental area; xscp, decussation of the superior cerebellar peduncle.
Microhemorrhages (microbleeds) were identified within each ROI and classified according to a semi-quantitative scale assigning scores from 0 to 3 (Supplementary Fig. S4). The intrarater reliability after 6 months was determined as excellent with Cronbach α of 0.90. As right and left hemispheres were counted separately and anatomical regions may span more than 1 tissue block, the maximum observed grade was assigned for each region. To assess total vascular pathology burden across anatomical planes and cases, we also calculated a composite vascular score (CVS) based on the frequency (f) and the severity of vascular lesions (G0 to G3) as follows: CVS = f(G0) × 0 + f(G1) × 1 + f(G2) × 3 + f(G3) × 9. The two control brains had no evidence of axonal or vascular pathology based on these criteria and were not further analyzed.
Statistical analysis
Comparisons between groups were performed using Student’s t test (t) with Welch correction for parametric data or Mann–Whitney (U) for nonparametric data. Associations between variables were assessed by Spearman’s correlation (ρ). Mixed effect analysis for APP densities and CVS was performed after log-transformation of data. For the assessment of frequency distributions of axonal and vascular pathologies, histograms were plotted for each case per critical versus noncritical group based on the measurements of individual ROIs. In the case of axonal lesions, frequency distributions for each case were fitted by log-normal models to estimate geometric mean and geometric standard deviations (SDs) as measures of average severity and variance of regional lesion densities. 19 For the assessment of vascular and axonal pathologies along the rostro–caudal axis of the brainstem, we performed mixed-effect analyses using log-transformed values. To assess regional differences in axonal and vascular pathologies and to identify anatomical areas whose involvement is predictive of clinical outcomes, we used Classification and Regression Tree (CART) analysis. 20 In brief, this method identifies predictive variables (anatomical regions) and quantitative criteria (amount of pathology) that can best classify and predict an outcome of interest (critical outcomes). This analysis was limited to a number of regions with at least 70% observations per group (n = 48). To facilitate CART analysis, missing values for APP density were imputed using the predictive mean matching method (with maxit = 20, seed = 600, and donors = 3) from the “mice” package in R. For imputing missing vascular grade data, a random forest-based imputation method was used from the “missForest” package. An iterative CART analysis was performed with the “rpart” package through recursive partitioning with the “gini” splitting criterion. A minimum bucket size of 4 was set for node splitting to enhance the reliability of outcomes. In each iteration, the variable identified as the first node was successively removed. For all statistical analysis, significance threshold (p) was set at 0.05. In addition, p values were not adjusted for multiple comparisons, unless otherwise stated. Graphs were plotted with GraphPad Prism 9.1 (GraphPad Software, La Jolla, CA, USA). Type of anesthesia (propofol or isoflurane) had no effect on clinical parameters or measures of total axonal or vascular pathology and was not considered further in any of the analyses.
Results
Clinical monitoring during and after BHBT and clinical outcomes
Cessation of spontaneous respiration was encountered after BHBT in the majority of cases. In the Hanford cohort, four out of six animals did not recover spontaneous respiration for the duration of the study. In the Yucatan cohort, on which we based our quantitative analyses, 34 out of 38 animals momentarily lost spontaneous respiration, whereas 8 cases never recovered spontaneous respiration for the duration of the study (referred hereafter as “critical” cases). In all injured cases we observed sinus tachycardia (193 beats per minute [bpm], SD 26) lasting up to 15 min postinjury. In a subset of Yucatan pigs (n = 15), ICP was monitored for up to 2 h by intraventricular sensors. BHBT was associated with an acute increase in ICP, with average peak at 34 (SD 20.3) mmHg above baseline in the first 15 min after injury (Table 1).
Characteristics of the Yucatan Swine Cohort
Subject information, injury parameters, and vitals are presented for the entire cohort (n = 38) and critical (n = 8), as well as noncritical, subgroups for all animals (n = 30) and for those used for neuropathological assessment (n = 14).
Intracranial pressure (ICP) measurements were obtained from a subset of critical (n = 5) and noncritical cases (n = 10). Data are expressed as mean values with SD in parentheses, unless otherwise specified.
HR, heart rate; ICP, intracranial pressure; SD, standard deviation.
General features of macroscopic and microscopic pathology
Skull fractures, configured as suture separation and osseous (transcranial or nontranscranial) fractures, were extremely common in our subjects (50% and 89% of suture separation or transcranial fractures, respectively) (Table 2). The majority of fractures involved the occipital (76% of cases), frontal (55%), and parietal bones (47%) as per samples illustrated in Figure 2. Orbital and temporal bone fractures were less common (32% and 21%, respectively). Critical cases had significantly higher total fracture scores compared with noncritical cases (Mann–Whitney U = 59, p = 0.025) due to a tendency for higher frequencies of parietal (Mann–Whitney U = 59, p = 0.016) and frontal fractures (Mann–Whitney U = 71, p = 0.053), which were in turn positively correlated with back-face velocity (for frontal, Spearman’s ρ = 0.49, p = 0.019, and for parietal, ρ = 0.43, p = 0.045). While occipital fractures were frequent and equally prevalent, in critical cases they were more often separated and closer to the foramen magnum, a pattern indicative of significant skull deflection opposite to the impact site.
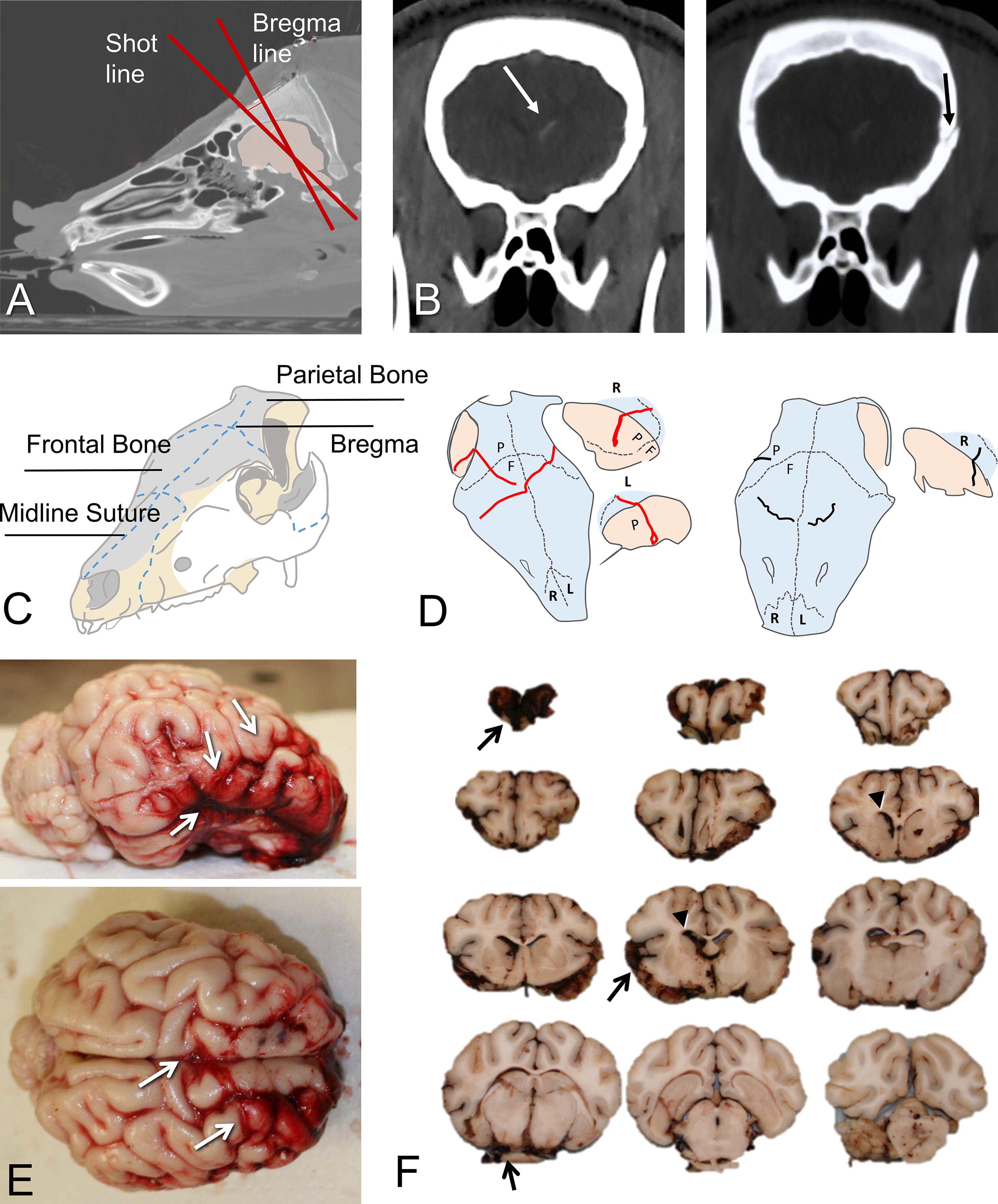
Gross pathology of skull, meninges, and brain associated with BHBT. Images and diagrams are from the Hanford cohort.
Skull Fracture Characteristics in the Yucatan Swine Cohort
Percentages of suture separation and osseous fractures are presented for the entire cohort, as well as critical and noncritical subgroups. Critical cases had significantly higher total fracture scores compared with noncritical cases (Mann–Whitney U = 59, p = 0.025) due to a tendency for higher frequencies of parietal (Mann–Whitney U = 59, p = 0.016) and frontal fractures (Mann–Whitney U = 71, p = 0.053).
The majority of cases in both the Hanford and Yucatan cohorts had SAHs. Photography-based maps of SAH for the Yucatan cohort (n = 36) are shown in Supplementary Figure S5. The majority of cases had extensive (65%) or moderate-size (19%) SAH mostly on top of the frontal lobes (92%) and sometimes extending to the temporal regions (51%) (Supplementary Fig. S5B). Basal SAHs were also very common (84%) with frequent extension around the brainstem (70%). The severity of SAH was highly correlated with back-face velocity (ρ = 0.54, p < 0.0008), as well as total fracture score (ρ = 0.58, p < 0.0001). Specifically, SAH severity was correlated with the presence of frontal (ρ = 0.45, p = 0.006), parietal (ρ = 0.47, p = 0.004), and occipital fractures (ρ = 0.42, p = 0.012). There were no significant differences between critical and noncritical cases in terms of distribution or severity of SAH.
There was extensive vascular and axonal pathology in a large number of cases. At least in the hemispheres, pathological severity was much higher and more frequent in the Hanford swine as illustrated in Figure 3. Hemorrhagic pathology included classical impact type, as well as more diffuse lesions. The former were coup lesions in medial frontal neocortex under the impact site (Fig. 3B, D) and, in some cases, less extensive contrecoup lesions in the olfactory tubercle region (Fig. 3E). These classical impact lesions 21 –24 were often associated with SAH located above or below the index pathology. Coup lesions were featured by microhemorrhages along the walls of small blood vessels in outer cortical layers, sometimes extending well into the surrounding parenchyma (Fig. 3C) and at times converging in gyral coronas that showed signs of ischemia (Fig. 3D). Many such lesions had a compressed or collapsed venule in the middle (Fig. 3C) and were adjoined by congested microvessels in the same region. Occasionally, there was continuity between SAH and a superficial microbleed (Fig. 4D, E). We also found diffuse microhemorrhages in gray or white matter away from the coup or contrecoup sites, with characteristics that were not very different from the ones in the superficial locations (Fig. 3F). In exceptional cases, we found extensive confluent microhemorrhages in the gray–white transition zone with the appearance of gliding contusions (Fig. 3G).

Representative patterns of vascular pathologies. Images are from sections prepared from the Hanford cohort and stained with Masson trichrome. Main panels are magnifications or outlined or pointed areas in insets.
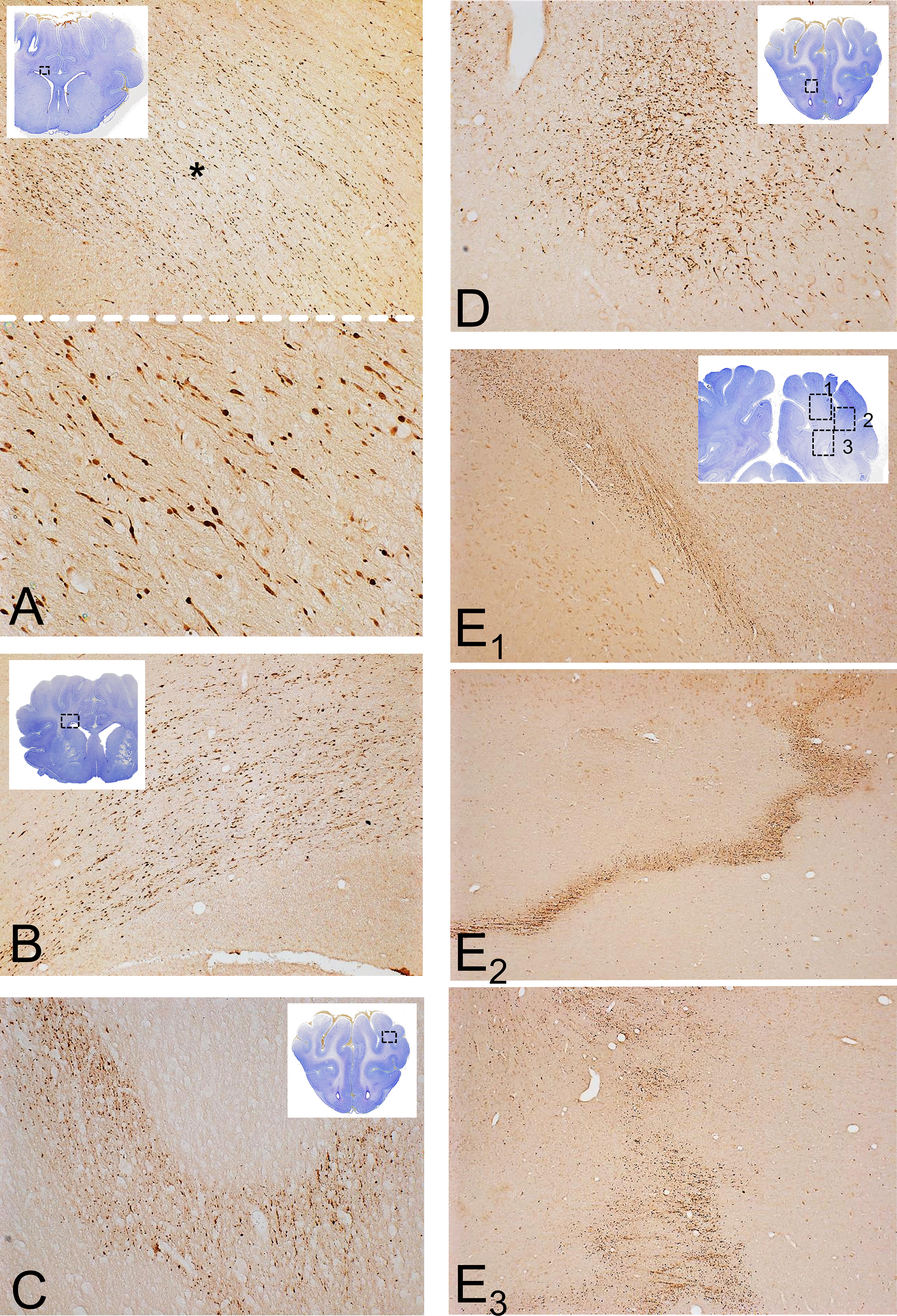
Representative patterns of axonal pathologies. Illustrations are from forebrain sections from the Hanford cohort immunostained for APP. For anatomical orientation, the images of Nissl-stained adjacent sections are included as insets in low magnification. (
Diffuse axonal injury (DAI) occurred in more than one pattern (Fig. 4). One was the classical pattern of populations of APP (+) distended or varicose axons and axon bulbs oriented parallel to native fiber bundles of axons in the corpus callosum, callosal radiation, anterior commissure (Fig. 4A–D), and in various locations in the diencephalon and brainstem as outlined in next section. Previous patterns were similar to ones reported in pure acceleration–deceleration scenarios. 3 –6 There was also punctate APP(+) axon labeling in several places in the subcortical white matter at the gray–white matter margin and, in some cases, an irregular “geographic” pattern of dense labeling in the deep white matter that is not coaxial with the orientation of fiber tracts and is classically associated with hypoxia 25,26 (Fig. 4E). As noted earlier, in the Yucatan cohort, axonal and vascular pathology was more limited. In the case of the forebrain, there were no significant differences of APP(+) lesions in the corpus callosum and subcortical white matter between critical and noncritical cases.
Brainstem pathology: axonal and vascular lesions and association with critical outcomes
To assess the distribution of axonal pathology in the brainstem following BHBT, we measured the density of APP(+) lesions across six standard anatomical planes. To address clinicopathological correlations involving critical outcomes, assessment was conducted on all 8 critical and 14 randomly selected noncritical cases serving as controls. While the total burden of axonal pathology in the brainstem did not significantly differ between critical and noncritical cases (Fig. 5A), the critical group was characterized by lesions per ROI that were on average more severe (median geometric mean density of 4.5 compared to 1.7 lesions per mm2 in noncritical cases, p = 0.016), as well as variable (median geometric SD of 4.3 compared to 2.8 lesions per mm2 in noncritical cases, p = 0.0002) (Fig. 5B).
With respect to the anatomical distribution of axonal pathology, there was a significant variation in pathological burden across the rostro–caudal axis of the brainstem (F3.2,57.3 = 15.8, p < 0.0001), as well as between the two groups (F1,20 = 4.4, p = 0.048) (Fig. 5C). For a more detailed regional analysis, we restricted comparisons to 63 anatomically defined regions observed in at least 4 cases per group. A heatmap of pathological burden is shown in Figure 5D. Regions with the highest densities of APP(+) axonal lesions across cases were the substantia nigra (both pars reticulata and compacta) and the cerebral peduncles. Thalamus also showed substantial pathology, both in major fiber bundles (e.g., external medullary lamina and fasciculus retroflexus) and in bundles or axons embedded in thalamic nuclei (reticular, mediodorsal/parafascicular, lateral posterior and medial geniculate nucleus, and the pulvinar). Major brainstem tracts, such as the medial lemniscus and medial cerebellar peduncle, were also involved and so were axon groups associated with the ventral tegmental area, basis pontis, and raphe nuclei.
To establish anatomical patterns of axonal pathology in critical and noncritical cases, we first generated a heatmap to visualize fold differences in axonal burden across ROIs (Fig. 5E). Interestingly, regions characterized by high burden of axonal pathology were similarly involved in the two groups, a pattern suggesting either shared propensity for these regions to be affected regardless of clinical outcome or a plateau effect. In contrast, there was a handful of regions with moderate or low pathological burden that clearly showed higher involvement in critical compared with noncritical cases. Such regions include the dorsal motor nucleus of the vagus (DNX), superior colliculus (SC), parvocellular reticular nucleus (PaRN), and the inferior olivary nucleus (ION).
To further explore patterns of DAI distribution, we conducted an iterative CART analysis. CART is a decision tree-based machine learning method that can identify anatomical patterns of pathology as predictors of clinical (critical or noncritical) outcomes. As shown in Table 3 (iteration 1), axonal injury in the PaRN (>9.1 lesions per mm2) is highly predictive of persistent loss of spontaneous respiration. To further assess how stable and how reliant CART predictions are on particular anatomical regions, we successively removed the best predictor and repeated the CART analysis for each iteration. Therefore, after excluding PaRN from the analysis, we find that the second-best predictor is ION, followed by nucleus of the solitary tract (NTS), pyramidal tract (py), intermediate reticular nucleus (IRN), magnocellular reticular nucleus (MgRN), SC, SNV, and so on (see Table 3). The first 8 iterations of CART have similarly strong discriminative potential in differentiating critical from noncritical cases, a pattern indicating an association of critical outcomes with increased axonal pathology in the medial medulla.
Iterative Classification and Regression Tree Analysis for Prediction of Critical Cases Based on Anatomical Distribution of Pathological Axonal Burden (Lesions per mm2)
The first 15 iterations are shown. The primary predictive criterion indicates the first node of the decision tree, which best differentiates between critical versus noncritical cases. In the majority of cases, there is no second criterion. Classification error, area under the curve of receiver operator curve (AUC-ROC), sensitivity, and specificity are calculated for the original data based on the complete decision tree for each iteration. See text for details.
Microhemorrhages or difuse vascular injuries (DVI) were also frequent throughout the brainstem. Their distribution and severity were assessed with the use of a semiquantitative scale applied to the same ROIs as axonal pathology. Composite Vascular Scores (see Materials and Methods) were calculated to enable comparisons across anatomical regions and cases.
Critical cases had significantly higher total burden of vascular pathology in the brainstem (Mann–Whitney U for CVS, U = 22, p = 0.02) (Fig. 6A). This increased burden was characterized not only by an increased frequency of grade 1 lesions (1.8 times more frequent in critical cases) but also a disproportionate increase in grade 3 lesions (32 times more frequent) (Fig. 6B). As with axonal pathology, mixed effect analysis along the 6 standard planes revealed a significant variation of pathological burden across the anteroposterior axis favoring caudal most planes (F3.34,59.6 = 5.6; p = 0.001) and also variation between groups (F1,20 = 7.4; p = 0.013) (Fig. 6C). A heatmap of pathological burden is shown in Figure 6D. Areas most affected included the nucleus prepositus (NP), central gray (CG), superior olivary nucleus (SON), reticular formation (median raphe, RPHE, MgRN, interpeduncular nucleus, IPN), and white matter tracts such as the medial lemniscus (ml) and medial longitudinal fasciculus (mlf). To define patterns of vascular pathology between critical and noncritical cases, we generated a heatmap showing differences in vascular scores among anatomical regions. This analysis revealed that areas with higher vascular pathology in all cases are also the ones that are most severely affected in critical cases (Fig. 6E). Iterative CART analysis identified the SNV and the cerebral peduncle (cp) and to a lesser extend the mesencephalic reticular nucleus (MRN) and the medial geniculate (MG) nucleus, as those regions differentiating critical from noncritical cases (Table 4), a pattern suggesting an association of critical outcomes with increased vascular pathology in the anterior ventral brainstem.
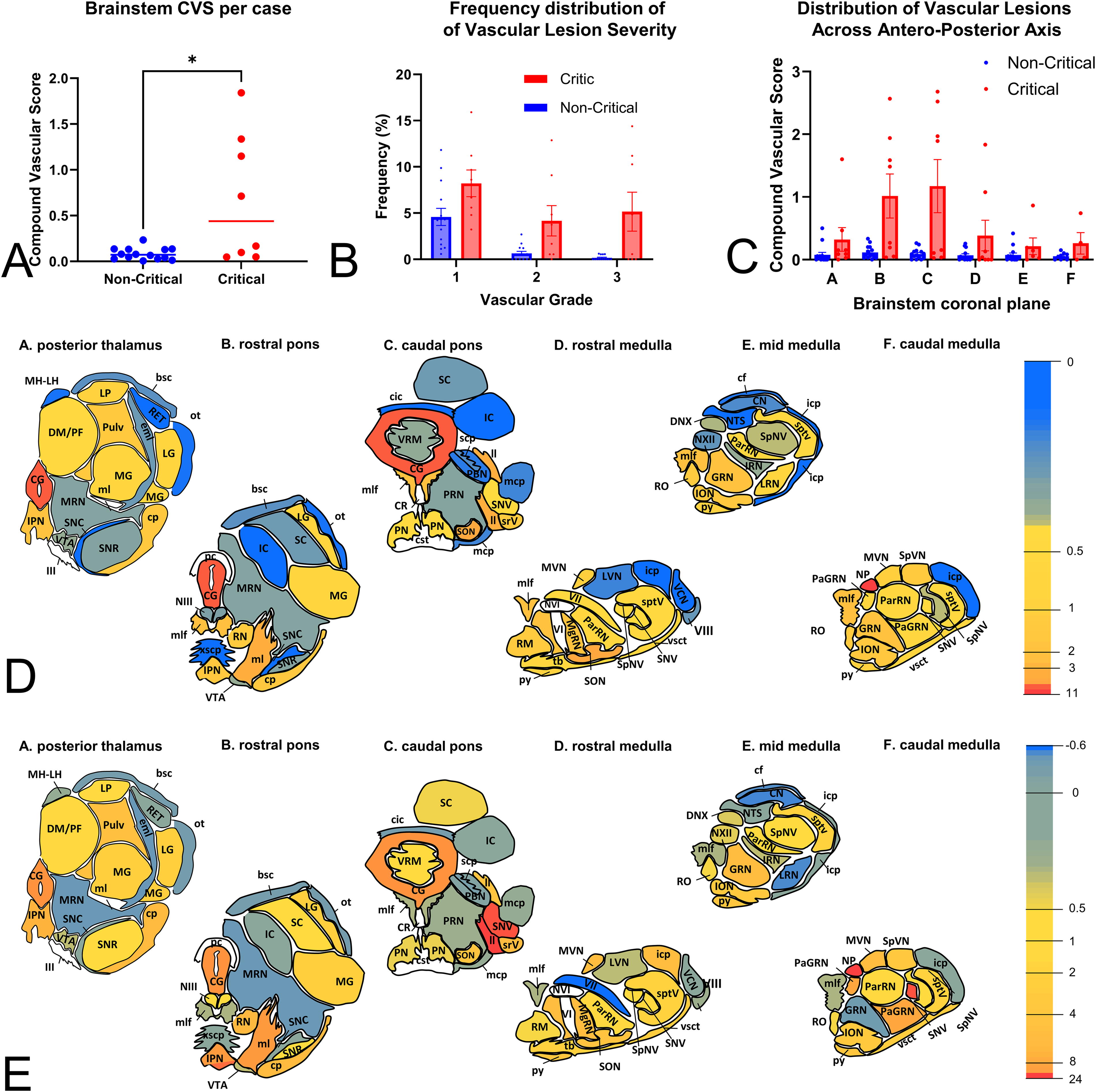
Total and regional severity of vascular pathologies in brainstem after BHBT.
Iterative Classification and Regression Tree Analysis for Prediction of Critical Cases Based on Anatomical Distribution of Pathological Vascular Burden (Grade)
The first 15 iterations are shown. The primary predictive criterion indicates the first node of the decision tree, which best differentiates between critical versus noncritical cases. In the majority of cases, there is no second criterion. Classification Error, area under the curve of receiver operator curve (AUC-ROC), sensitivity, and specificity are calculated for the original data based on the complete decision tree for each iteration. See text for details.
We subsequently asked whether regional axonal or vascular pathologies would better predict persistent apnea. For this, we performed the same analysis, including both axonal and vascular pathology measures. We found that axonal measures of regional pathology consistently emerged as the main predictors of apnea in up to 9 iterations (PaRN, ION, NTS, py, IRN, MgRN, SC, SNV, eighth vestibulocochlear nerve [VIII]).
Brainstem pathology: other clinicopathological correlations
To better understand how axonal and vascular pathology may relate to each other and to other neuropathological or clinical measures, we also explored potential associations between relevant variables across individual cases.
First, we explored injury-related predictors of brainstem pathology. We found that the total burden of brainstem axonal pathology was highly correlated with helmet back-face velocity (ρ = 0.79, p = 0.003). Total vascular pathology was also positively correlated with back-face velocity (ρ = 0.63, p = 0.032) but not with other factors (Supplementary Fig. S6). Second, we explored correlations of axonal pathology in brainstem with other pathologies, that is, fracture score, SAH severity, brainstem vascular lesions, and forebrain pathology. We found that total axonal pathology in the brainstem was strongly correlated with both SAH severity (ρ = 0.53, p = 0.005) and with the composite vascular score (ρ = 0.68; p = 0.001).
Although microhemorrhages, such as those described here, are commonly considered surrogates of axonal pathology 27 since total vascular and axonal pathology burden are often correlated with each other, colocalization of vascular and axonal pathologies was uncommon in our material. Therefore, to gain deeper insight into potential relationships between vascular and axonal pathologies, we assessed their co-occurrence within each ROI across all cases (n = 4303 individual regions). We found that regional correlation of vascular and axonal pathologies was very weak with Spearman’s ρ of 0.11 (p = 3.6 × 10−12), indicating that the regional distribution of vascular lesions is not a good predictor of the distribution of axonal lesions (and vice versa).
Discussion
Summary of key findings and significance of the study
Behind helmet blunt trauma is a significant clinical entity not only due to its potential to cause severe acute neurological morbidity but also because of a broad significance for military, as well as civilian, athletic and other settings, where helmets are utilized as protective head gear. In the current study, we have utilized a swine model of high-energy BHBT to characterize, for the first time, key neuropathologies and their association with acute respiratory decompensation, one of the main causes of acute mortality in high-energy blunt TBI in humans. The presentation of BHBT is complex: The majority of cases involve fractures of the cranium at both the impact site and away from it, extensive SAHs, as well as focal lesions in the form of coup and contrecoup contusions, and diffuse vascular and axonal lesions throughout the cerebrum, especially in the brainstem. Absence of spontaneous respiration is frequently encountered and is associated with higher back-face velocity, higher total fracture score, as well as more severe axonal and vascular lesions in the brainstem. A focused regional examination of brainstem pathology unveiled a potential critical link between adverse outcomes and diffuse axonal lesions within the medial medulla or vascular lesions within the anteroventral brainstem. Vascular pathology (microbleeds), although correlated with DAI severity across cases, did not appear to be colocalized in the exact same areas as DAI and therefore cannot be used as surrogate marker of traumatic axonal injury.
Blunt-force TBI is extremely common in civilian, as well as military, populations and occurs in a variety of conditions, including falls, motor vehicle accidents, contact and collision sports, construction work, and in similar scenarios involving military personnel both in combat and in barracks. Behind helmet blunt trauma is a special type of blunt-force TBI, in which the impact of high energy ballistics that would normally penetrate the skull is exerted locally to the skull through the rapid back-face deformation of the helmet. 28 Therefore, BHBT is a unique and interesting scenario that has implications for both the design of protective gear and the management of injured patients in both military and civilian settings.
Traditionally, impact TBI has been associated with coup and contrecoup contusions, whereas diffuse vascular and axonal injuries encountered in TBI have been linked, primarily based on work on nonhuman primates and swine, to impulse forces in the form of rotational acceleration. However, in the clinical setting, impact forces are nearly ubiquitous and commonly associated with diffuse lesions, especially in motor vehicle accidents and high-impact falls. 29,30 The present swine model of high-energy BHBT replicates several features of BHBT in the human head, 31 –33 including the generation of linear and depressed fractures in the vicinity of or away from the impact site. In addition, our study confirms that blunt-force impact TBI with minimal, if any, rotational acceleration can cause diffuse lesions in axons and blood vessels that are associated with poor outcomes.
General mechanisms of DAI and DVI
Diffuse axonal injury is one of the most common pathological features of TBI of a variety of causes and is thought to result from rapid deformation (dynamic loading) to the axons, primarily from rotational acceleration of the brain. 3,4,7,34 –36 The principal role of impulse loading in the induction of DAI was first demonstrated in pure rotational acceleration models by Gennarelli, Meaney, Smith, and colleagues. 3 In the swine, inertial loading is sufficient to produce DAI throughout the white matter along with petechial hemorrhages and tissue tears and can result in loss of consciousness depending on the plane and magnitude of acceleration. 4,37,38 Although high-energy BHBT injury is achieved by high-velocity impact loading, transmitted forces through the helmet may result in both direct local deformation of the skull and translational and possibly rotational acceleration of the head, although the magnitude of the latter (if it occurs) cannot be compared with this of the previously cited swine model of the University of Pennsylvania. As expected from inertia models, total burden of DAI after BHBT correlated well with impulse loading, at least as inferred by impact velocity. Similarly, captive bolt pistol experiments in sheep (with freely rotating head) have produced contusions, SAH, DAI, and DVI. 39 –41
In view of the limited experimental manipulation in subjects of this study, we cannot state with certainty what the causes of DAI and DVI are here or why BHBT was universally associated with DAI and DVI and more so in the brainstem than in the forebrain. However, in our BHBT model, the impact was delivered approximately through the center of gravity of the head, and the helmeted head of the swine was secured on a flat table; therefore, rotational acceleration of the head would not be significant. We hypothesize that, in contrast to the low-rate low-energy impacts studied in traditional models of blunt TBI in which the skull would more likely demonstrate rigid-body motion, high-rate, high-energy, and low-mass impacts are more likely to cause local deformation of the skull around the impact point and propagation of high tensile strains remotely from the impact site. 42 While there is limited work on the biomechanics of BHBT, early observations from unprotected human skull injury models are consistent with the idea that unless the impact results in perforation or comminution, tensile strains may be higher and reach the threshold of failure away from the impact site. 43 We have previously also shown that in a human cadaver model, higher energy BHBT impacts are also associated with distal linear fractures. 33 Similarly, simulations of control cortical impact have indicated that larger impactors and faster impacts lead to more diffuse involvement of regions that experience high strains associated with damage. 44
In our model, this hypothesis is supported by the pattern of fractures, especially in critical cases. Compared to other high-energy impacts where a lot of the energy is released through a high degree of local bone comminution, in BHBT there can be a lot of energy still available in the form of kinetic energy that can generate a lot of displacement and, therefore, strain in the brain tissue. 45 The presence of parietal fractures suggests a large degree of skull deflection that is sufficient to reach tensile strain thresholds away from the impact site. At the same time, the presence of separated fractures in the occipital bone close to the foramen magnum suggests that the ventral side was subjected to higher tensions than the dorsal side. Of note, in our preliminary analyses of the Hanford cohort, we found that overall pathology and clinical outcomes were more severe than in the main Yucatan cohort, and we also found that the Hanford swine had incomplete suture fusion. Hence some of the differences in outcome severity could be potentially explained by a further increase in skull deflection after impact due to incomplete suture fusion.
We surmise that high degree of skull deflection impacts a large area of the cranium and may lead to high levels of brain tissue strain distal and opposite to the original impact site and toward ventral aspects of brainstem. Thus, this mechanism may explain the significant presence of DAI and DVI lesions at these sites. While in the current study we are not able to conclude what underlies the consistent involvement of the brainstem, the potential vulnerability of the brainstem was first suggested in early studies by Gurdjian and Lissner who found that impacts may lead to increased strains in the craniospinal junction. 46 This was later explained by impact-related pressure gradients in the cranial cavity and relative movement of the brain after impact. 47 More recently, simulations of controlled cortical impact in rodents have revealed that larger and faster impacts of the brain itself can also be associated with higher levels of strain and damage occurring at the brainstem. 44 However, differences between skull and brain anatomy among models, and types of injury, make drawing any parallels difficult. Notwithstanding, we cannot exclude that some of the axonal pathologies observed here, for example, the ones with geographic appearance, may be secondary to hypoxic injury. 25,26
Further commentary on pathogenic mechanisms
Similar to DAI, DVI is thought to be caused by the dynamic deformation of blood vessels resulting in hemorrhages around arterioles, venules, and capillaries. 48 Mechanisms of DVI have not been as well characterized as those of DAI, but DVI has been reported in moderate-to-severe TBI and has been associated with severe DAI. 30 In our study, DVI was observed across the cerebrum and particularly in the rostral brainstem. Early neuropathological studies have emphasized the occurrence of microhemorrhages in the brainstem after human blunt impact TBI (especially with frontal blows in the rostro–caudal direction) and their more frequent occurrence in the periaqueductal gray matter and lateral brainstem structures such as the colliculi and cerebellar peduncles, 48,49 which is consistent with our findings.
Diffuse axonal and vascular injuries have been thought to arise from similar injury mechanisms, and historically, the presence of DVI, as revealed by susceptibility weighted imaging, has been used in the clinic for the diagnosis of DAI. 27 This idea has been reinforced by the Adam’s DAI classification, in which higher grades of DAI are associated with macroscopic hemorrhagic lesions in the corpus callosum and brainstem. 30 In our study, brainstem DAI burden was modestly correlated with brainstem DVI burden (DVI predicted 36% of the variability of DAI). However, when we looked at the regional distribution of DVI and DAI, the correlation broke down (only 4% of regional DAI variability could be predicted by DVI), a pattern indicating that the responses of axons and vasculature to regional tissue deformation may be different. While we have not assessed whether DVI may eventually cause secondary axonal injury, imaging studies in more chronic cases have found no correlation between measures of white matter microstructure and the presence of vascular lesions after adjusting for injury severity. 50 –52 Therefore, we conclude that the presence of DVI cannot be used to infer the distribution of DAI and may not always predict its severity.
The relationship of SAH with focal and diffuse lesions
Of potential mechanistic importance in our study is the nearly ubiquitous presence of SAH. Traumatic SAH is distinct from aneurysmal SAH and can be caused by tearing of bridging veins or pial vessels or leakage of blood into the subarachnoid space from adjacent cerebral contusions. 53 Traumatic SAH is very common and has been observed in up to 80% of “mild” TBI. 54 Due to limitations in our experimental material that had not allowed the manipulation of many injury parameters, we cannot be certain of the role and consequences of SAH after BHBT. However, the severity of SAH was highly correlated with the severity of the impact. In addition, the variable location of SAH with respect to the impact site and some relationships with contusional microhemorrhages may point to multiple mechanisms. Interestingly, despite a correlation between fracture and SAH severities, we did not find a spatial relationship between the two, perhaps indicating independent mechanisms in their generation. With respect to the role of SAH in brain injury, SAH may interrupt venous return, cause vasoconstriction and, thus, raise ICP. The contribution of traumatic SAH to axonal injury should also be considered, especially because spontaneous SAH has already been associated with reduced white matter integrity in humans 55 and experimental SAH in mice has also been associated with multifocal axonal injury. 56 Our findings suggest that SAH may be a good indicator of overall injury severity, but its relationship with other pathologies is not straightforward. This assertion is also in register with data from clinical series. 57,58
Clinicopathological aspects of critical deterioration
Acute cardiorespiratory decompensation, particularly in the first 10 min after TBI (“critical phase”), has been long recognized as a major cause of morbidity and mortality, although mechanisms are still poorly understood. 59,60 Cessation of spontaneous respiration after TBI, also referred to as “Impact Brain Apnea” (IBA), was a nearly ubiquitous finding in our model of high-energy BHBT. Apnea following TBI has been reported in multiple animal models and has been correlated with the magnitude of inertial or impact loading. 60 Pathophysiological changes in brainstem respiratory centers have been implicated as a possible cause. 59,60 Our focused analysis to identify brainstem vascular and axonal pathology in animals that experienced apnea after BHBT revealed that the inability to recover spontaneous respiration was predicted by the selective presence of DAI in regions such as the PaRN, ION, and NTS and not the overall burden of DAI in the brainstem or cerebrum, a finding that is consistent with the previous idea. Although we did not have the ability to directly assess injury in particular respiratory centers, automatic breathing depends on a central pattern generator that is distributed in the dorsolateral pons, NTS, and ventrolateral medulla, 61,62 areas that largely overlap with the predominant location of DAI in critical cases. Interestingly, the regions that appear to relate to critical outcomes were not the ones with the highest burden of DAI, a pattern indicating that location rather than burden of DAI may be more important. Indeed, a key difference between critical and noncritical cases is that, while both groups have the highest burden of pathology in the midbrain and pons, in critical cases pathology extends to more caudal brainstem levels. In contrast to DAI, the burden of vascular lesions in critical cases favors more rostral brainstem regions. Overall, the distribution of vascular lesions does not predict clinical outcomes, as well as that of DAI. We, therefore, believe that the higher burden of vascular pathology in critical cases may simply reflect the overall higher severity of injury.
A better understanding of the mechanisms behind brainstem injury and postinjury apnea is urgently needed. This understanding is especially critical since timely resuscitation is not possible in many cases of IBA, even when the injury itself is not immediately fatal or particularly severe. This is highlighted by clinical reports of postinjury apnea occurring even with minimal radiological or histological evidence of injury. 59,60,63
Implications for protective head gear and the study of human TBI
At this juncture, helmets are mostly helpful in preventing fractures more so than TBI (particularly concussion). 64 –66 The relationship between cranium/skull and brain protection is not straightforward and needs to be better understood. Cranial and parenchymal mechanical responses to blunt force are quite different and the cranium itself is a dynamic mechanical entity that may behave differently at different levels/intensities of impact loading. In addition, BHBT studies using cadaver heads or surrogates have shown that outcomes (e.g., fractures) in ballistic (high energy) BHBT cannot be predicted or extrapolated from previous modeling from lower energy blunt injury models, for example, relevant to vehicle collisions or sports injuries. Crucially, there is dearth of information on the effect of such high-energy injuries on the brain parenchyma. 28 Therefore, understanding some of the key pathological mechanisms via which blunt-force TBI with and without protection of the cranium with helmet causes focal and diffuse injuries is a high research priority. This need is further emphasized by findings indicating that the location of the injury, rather than the overall injury burden, is more closely associated with critical outcomes. These insights highlight both the need for more detailed and nuanced mechanical models and surrogates and the inherent complexities in translating findings from animal and cadaver models and developing protection strategies. Indeed, while the current study provides important insights in the neuropathology of high-energy, high-rate BHBT in the swine and potential mechanisms of brain impact apnea, findings may not be directly applicable to human BHBT. Skull deflection, fracture patterns, and critical brain regions along the impact axis will be dependent on the skull and brain geometry and, thus, may vary between species. Unfortunately, a detailed neuropathological understanding of high-energy BHBT in humans remains incomplete, and more studies are needed to better address these issues.
Transparency, Rigor, and Reproducibility
This study was not preregistered. Analyses of experimental materials were performed by investigators blinded to relevant characteristics of the subjects. Information related to helmet and projectile specifications cannot be disclosed due to their sensitive nature related to Department of Defense. Further information on equipment and analytical reagents used to perform experimental manipulations is available upon request. The statistical tests and assumptions are detailed in the Materials and Methods section. A limited number of histological samples from each of the experimental groups are available for future analyses on request. The authors have agreed to publish the article using the Mary Ann Liebert Inc. “Open Access” option under appropriate license.
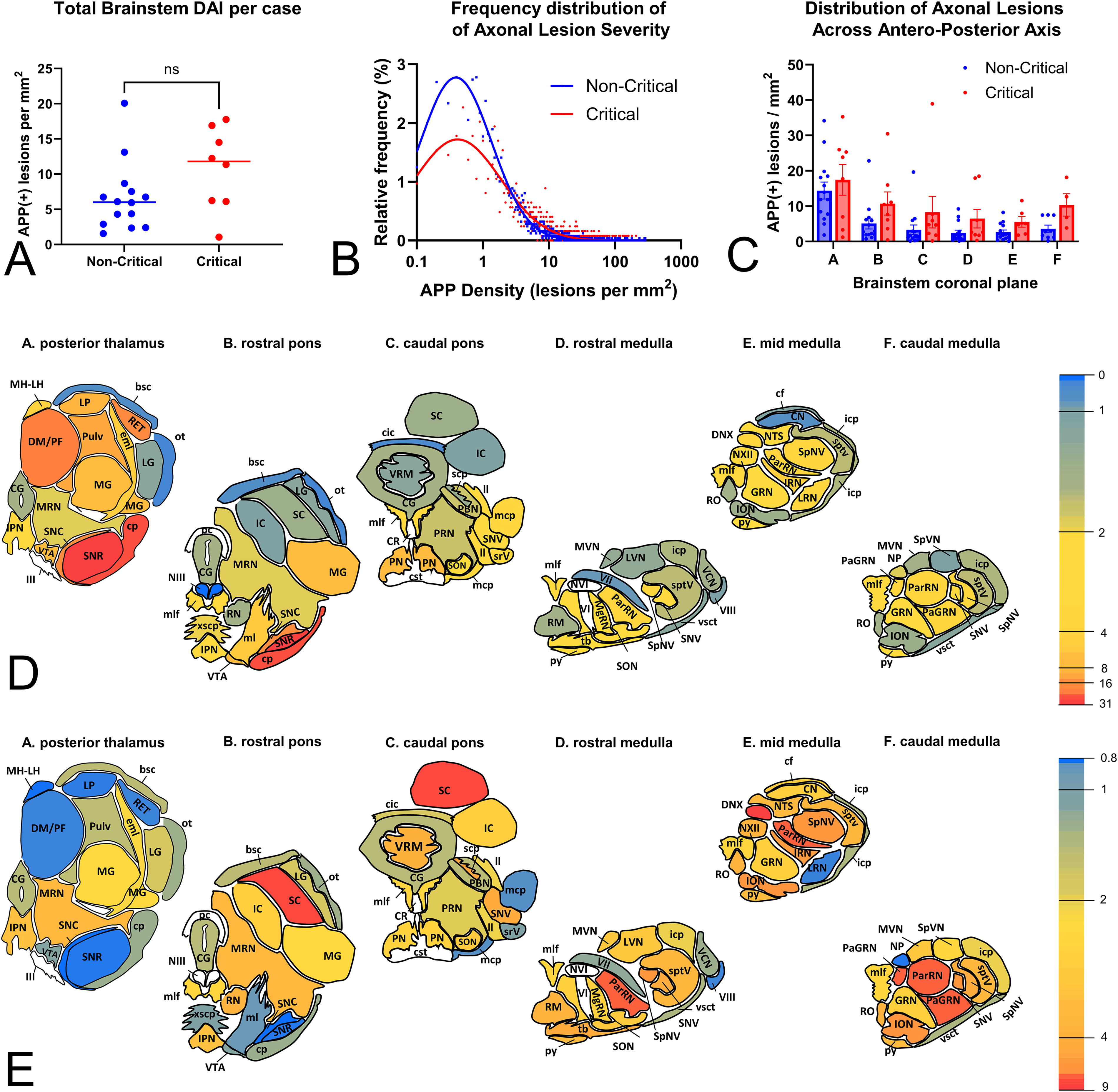
Total and regional severity of axonal pathologies in the brainstem based on APP immunohistochemistry (Yucatan cohort).
Footnotes
Authors’ Contributions
A.S.A.: Conceptualization, data curation, formal analysis, investigation, methodology, visualization, writing—original draft, and writing—review and editing; K.R.: Conceptualization, data curation, formal analysis, investigation, project administration, resources, and writing—review and editing; M.H.: Data curation, investigation, project administration, and resources; S.W.: Data curation, formal analysis, investigation, project administration, and resources; J.B.: Investigation and methodology; P.F.: Investigation and methodology; J.R.: Data curation and investigation; C.E.F.: Formal analysis and writing—review and editing; D.I.: Investigation and writing—review and editing; D.P.: Conceptualization and funding acquisition; and V.E.K.: Conceptualization, formal analysis, funding acquisition, methodology, project administration, supervision, writing—original draft, and writing—review and editing.
Author Disclosure Statement
The opinions expressed herein are those of the authors and not necessarily representative of those of the Uniformed Services University of the Health Sciences (USUHS), the Department of Defense (DOD), the United States Army, Navy, or Air Force, any other U.S. government agency, and Henry M. Jackson Foundation for the Advancement of Military Medicine, Inc. (HJF) unless so designated by other authorized documents. The authors have no conflicts of interest to declare.
Funding Information
The study was performed in connection with contract/instrument W15P7T-19-D-0126 with the Combat Capabilities Development Command (DEVCOM) Army Research Laboratory and sponsored by the Military Operational Medicine Research Program of the U.S. Army Medical Research and Development Command and by the U.S. Army Medical Research and Materiel Command via a subcontract between Johns Hopkins University and Henry Jackson Foundation/Uniformed Services University.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.