
Abstract
Human neural stem cells (hNSCs) possess significant therapeutic potential for the treatment of traumatic brain injury (TBI), a leading cause of global death and disability. Recent pre-clinical studies have shown that hNSCs reduce tissue damage and promote functional recovery through neuroprotective and regenerative signaling and cell replacement. Yet the overall efficacy of hNSCs for TBI indications remains unclear. Therefore, this systematic review aims to evaluate hNSC interventions compared with controls in pre-clinical TBI models. Through this process, variations in hNSC administration protocols were consolidated, and key knowledge gaps were identified. Meta-analysis was applied to primary outcomes of lesion volume, Morris Water Maze (MWM) performance, modified Neurological Severity Scores (mNSS), and the rotarod task. Narrative review of secondary outcomes included hNSC survival and differentiation, endogenous neuron survival, axonal injury, and inflammation. Overall, hNSC intervention reduced lesion volume, enhanced MWM performance, and led to trending decreases in acute and chronic neurological deficits at acute and chronic time points. These results suggest hNSCs demonstrate clear efficacy in pre-clinical TBI models. However, further studies are needed to address key questions regarding optimal hNSC administration (e.g., dosing, treatment window) and underlying mechanisms of action prior to progressing to human clinical trials.
Introduction
Traumatic brain injury (TBI) is a leading cause of death and disability, affecting over 1.5 million Americans annually and resulting in 190 TBI-related deaths daily. 1,2 Roughly 2% of the current population lives with TBI-related impairments such as learning, memory, motor function, and behavioral deficits, which can persist long-term and disrupt an individual’s quality of life. 1 –4 Primarily caused by falls, motor vehicle accidents, or sports-related injuries, TBI is classified by injury severity as indicated by the physician-assigned Glasgow Coma Scale (GCS; mild 13–15, moderate 9–12, severe <9). 4,5 TBI is first characterized by the primary injury endured at the moment of impact in which external mechanical forces induce tissue shearing, deformation, stretching, and tearing. 6 The secondary injury then develops from minutes to weeks post primary injury, and persists even over years. 7 This phase, commonly referred to as the secondary injury cascade, represents a series of molecular, cellular, and tissue changes due to excitotoxicity, inflammation, necrosis, and apoptosis. 8 Although the neuropathological and functional deficits induced by the primary injury are unavoidable, the secondary injury cascade represents a therapeutic target that could be dampened or reversed, thus limiting long-term patient deficits. Despite the significant prevalence of TBI and associated mortality, there are currently no Food and Drug Administration (FDA) approved treatments.
A growing body of literature evaluating the therapeutic potential of human neural stem cells (hNSCs) across various neurodegenerative diseases and injuries including Alzheimer’s Disease, 9 –11 Huntington’s Disease, 12 –14 stroke, 15 –18 and TBI 19 –22 has shown that hNSCs have significant therapeutic capabilities. The promise of hNSC therapy is due to multimodal therapeutic effects including the production of neuroprotective and regenerative factors, the ability to differentiate into neurons, astrocytes, and oligodendrocytes, and serve as a source for cellular replacement for damaged tissue. 14,19 –24 In rodent TBI models, hNSC treatment led to the mitigation of pathological progression including decreased neuroinflammation, apoptotic cell death, and axonal injury. 20,25 –27 Moreover, hNSCs promoted tissue regeneration with hNSC-treated animals showing increased endogenous Nestin+ NSCs and doublecortin+ (DCX) neuroblast populations indicative of neurogenesis. 28 These dynamic cellular changes in pathophysiology resulted in improved survival, cognition, behavior, and motor function in rodent models. Despite these promising results, there has been significant variability with respect to cellular and functional outcomes, raising questions pertaining to the efficacy of hNSC intervention for TBI patients. However, this observed variability may be due to a lack of standardization between studies evaluating hNSC TBI treatment including differences in cell source, dose, delivery route and location, administration interval, and immunosuppression regimen. The recognition of this limitation in the field highlights the opportunity to perform a meta-analysis of hNSC TBI treatment to better understand the best practices that lead to the most beneficial outcomes.
The purpose of this systematic review was to assess the efficacy of hNSC therapy compared with vehicle administration in pre-clinical TBI models and provide a comprehensive review of the literature. Meta-analysis was applied to the primary outcomes of interest: lesion volume, Morris Water Maze (MWM), modified Neurological Severity Score (mNSS), and rotarod performance. Lesion volume was used to assess tissue damage. MWM was used to evaluate cognitive function, specifically spatial memory. The mNSS assessment quantified neurological deficits. The rotarod task measured changes in motor function. Qualitative analysis of secondary outcomes included hNSC survival, engraftment, differentiation, and endogenous neuronal preservation, axonal injury, and inflammation. Additionally, important technical hNSC administration variables including cell source, dosage, and administration time points were evaluated.
Methods
Prior to searching the literature, the study protocol was identified and finalized including the definition of the objectives, search strategy, inclusion/exclusion criteria, data collection, and outcomes of interest. The reporting of this systematic review was performed in accordance with the Cochrane Handbook for Systematic Reviews 29 and based on the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) statement. 30
Search strategy and study selection
A comprehensive search of online databases was performed within PubMed, ISI Web of Science, and Cochrane Library to identify relevant publications that evaluated hNSCs in pre-clinical TBI models published from October 1, 2002 to June 1, 2024. The search strategy included a combination of keywords: “traumatic brain injury,” “brain contusion,” or “brain concussion” AND “neural stem cells,” “neural progenitor cells,” “induced pluripotent stem cells,” or “neural precursor cells.” Selected studies were limited to original research articles published in English with full text available through the University of Georgia online access.
Identified articles were imported into Endnote (Version 21.3, Clarivate Analytics) and the duplication identification command removed identical articles automatically. The remaining studies were evaluated through initial title screening followed by a secondary in-depth screening of both title and abstracts within Covidence systemic review software (2024, Veritas Health Innovation) to identify therapeutic intervention of hNSCs within pre-clinical TBI models. If initial screening of the titles or abstracts did not clearly identify the cell origin, articles were moved to full-text screening for further evaluation. The remaining studies underwent full-text screening to determine if the publication fit within the pre-determined inclusion and exclusion criteria of the systematic review.
Inclusion and exclusion criteria
The PICO framework to identify the desired Patient or Population, Intervention, Comparison, and Outcomes (Table 1) was used to develop the inclusion and exclusion criteria (Table 2).
PICO Framework Outlining the Population, Intervention, Comparison, and Intended Outcomes for the Development of the Research Question and Search Protocol
MWM, Morris Water Maze; mNSS, modified Neurological Severity Score; TBI, traumatic brain injury.
The Inclusion and Exclusion Criteria Used to Identify Relevant Studies of Interest
hNSC, human neural stem cell; hNPC, human neural progenitor cell; TBI, traumatic brain injury.
Data extraction
Data extracted from each publication included study characteristics of author, year, species, gender, age or weight, TBI model and parameters, TBI location and severity, immunosuppressant protocol, hNSC origin and cell line, cell dose, intervention time frame, route, relevant outcomes, and the planned end of the study. Lesion volume, MWM, mNSS, and rotarod assessments were selected as primary outcomes. Lesion volume was used to evaluate tissue damage and was reported in individual articles as an estimated volume in cubic millimeters or as a percentage of the ipsilateral hemisphere. MWM performance measured cognitive spatial memory. Within the literature, MWM is reported utilizing numerous outcomes (e.g., latency to reach the platform, distance to the platform, path length to the platform, swimming speed, immobility, and time spent in each quadrant) and may be evaluated multiple times within the same study. For this systematic review, MWM performance was recorded as the latency to reach the platform at the final time point provided. Due to repeat outcomes reported across various days for mNSS (neurological function) and latency to fall within the rotarod task (motor function), results were extracted at both an acute time point (ranging from 5 to 8 days post-transplantation) and a chronic time point (the last time point reported). For all outcome measures, the sample size, mean, and standard deviation (SD) or standard error of the mean (SEM) were manually extracted and recorded within Review Manager (RevMan Web; 8.1.1).
Nearly all publications represented data in figures or graphs only. For these studies, PlotDigitizer (2024; v3) was used for interpolation of continuous data including mean, SD, or SEM. If measures of variance were not clearly indicated as SD or SEM, the measure was assumed to represent standard error to prevent overestimation of effect sizes. Based on the Cochrane guidelines, if graphical data was presented as the mean and interquartile range, it was removed from the meta-analysis as the assumption of normality was inappropriate and therefore determination of the required SD was not possible. 29
Quality assessment
The quality of each included study was assessed using the Systematic Review Center for Laboratory Animal and Experimentation (SYRCLE) risk of bias (RoB) tool for animal studies. 31 This is a well-recognized RoB tool based on the original Cochrane RoB tool adjusted specifically for animal intervention studies. SYRCLE contains 10 entries focused on selection bias, performance bias, detection bias, attrition bias, reporting bias, and other biases (Supplementary Table S1). Through a series of signaling questions, bias was reported for each study as either “low risk,” “unclear,” or “high risk.” Graphical summaries of the SYRCLE tool were visualized using Robvis. 32
Data analysis
A combination of meta-analysis and descriptive narrative presentation is provided within this systematic review to present an accurate and robust synthesis of the literature with the data available within the identified publications. Primary outcome measures required a minimum of three studies with retrievable data to be considered eligible for meta-analysis. Review Manager software (RevMan Web; 8.1.1) was utilized for all statistical analyses of the primary outcomes. The sample size, mean, and SD for each treatment and control group were recorded. If SEM was provided within the original article, RevMan calculated the SD based on the mean and sample size input. The inverse variance method of meta-analysis was applied with all primary outcomes presented as continuous variables. Due to high levels of statistical heterogeneity confirmed by I 2 values >50%, a random effects model was used to determine the standardized mean difference (SMD; Hedge’s g) with a 95% confidence interval (CI). The overall effect for each outcome was evaluated using the Z-test and considered statistically significant if p < 0.05. Cohen’s general rule of thumb for interpreting the absolute value of SMDs was applied: small effect (<0.2), moderate effect (0.21–0.8), and large effect (>0.8). 29 Forrest plots were used for visualization of the corresponding results for each outcome.
Additional outcomes of interest that were deemed important but did not meet the minimum requirements due to their statistical and pre-clinical heterogeneity for inclusion in a quantitative meta-analysis included exogenous cell survival, engraftment, and differentiation, endogenous neuron populations, axonal damage, and measures of inflammation. These secondary outcomes were presented in a qualitative descriptive manner.
Results
Search outcomes
The electronic database search presented a total of 715 non-duplicated original research articles that investigated NSCs within pre-clinical TBI models. Initial results were narrowed down to 126 studies by screening of titles and abstracts followed by full-text screening of 42 articles. Finally, a total of 20 articles were identified as meeting the pre-determined criteria and therefore included within this systematic review. Specific information detailing the search process can be found within the PRISMA flow chart (Fig. 1) and in the “Methods” section.
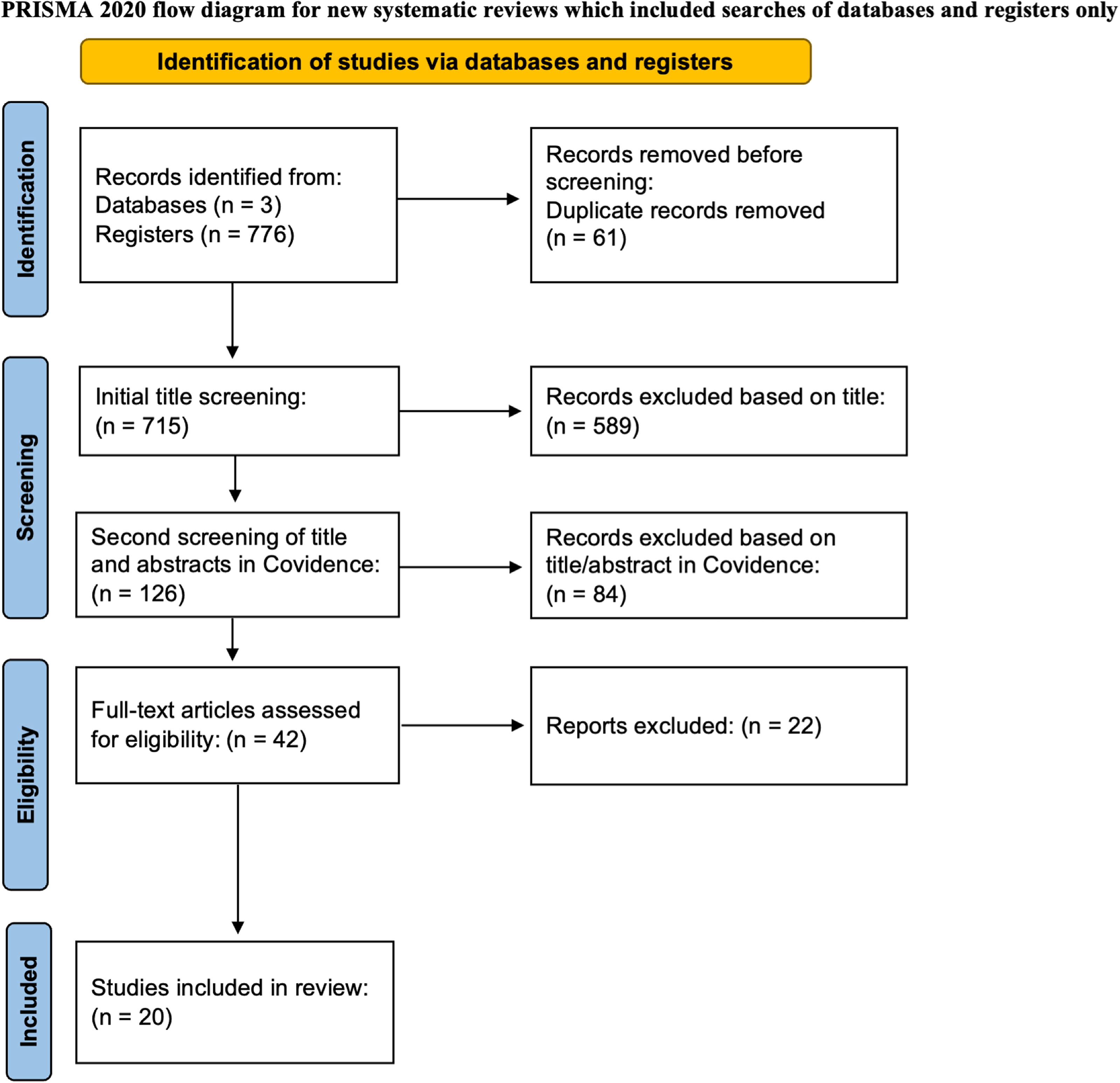
Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) 2020 flow diagram depicting the article identification, screening, and selection protocol.
Study characteristics
The 20 studies included within this review were published between 2006 and 2023 and their characteristics are summarized in Table 3. Of these, 13 originated from the United States (65%), 3 from Iran (15%), 2 from China (10%), and 2 from Germany (10%; Fig. 2A). The predominant model species were rats (85%), with mice comprising the remainder (15%; Fig. 2B). Male animals were used most frequently (75%), while only two studies reported the use of both sexes (10%) and one study used females (5%; Fig. 2C). Notably, the animal’s sex was unclear in 2 of the 20 studies (10%). Various methods were used to induce TBI, with controlled cortical impact (CCI; 50%), penetrating TBI via probe (pTBI; 20%), and punch biopsy (15%) being the most common (Fig. 2D). Fluid percussion injury (10%) and blunt instrument induction (5%) models were employed less frequently. A total of 14 studies (70%; Fig. 2E) reported the use of immunosuppression within their animal model. Of these 14, 12 reported immunosuppression through drug administration (60%), while the remaining 2 studies utilized an immune-compromised animal model (10%). Two studies (10%) explicitly stated the use of an immunocompetent model and the remaining four (20%) did not address immunosuppression within the study protocol. hNSCs originated from fetal tissue (60%), embryonic stem cells (20%), induced pluripotent stem cells (15%), or adult tissue (5%; Fig. 2F; Supplementary Table S2).

Study characteristics included within the systematic review. Data regarding the publication origin
Study Characteristics Related to the 20 Identified Publications Evaluating hNSC Efficacy in Pre-Clinical TBI
TBI parameters are reported as the penetration depth of depression (mm), the velocity of which the injury is induced (m/s), and the dwell time in which the impactor remains in contact with the tissue (s).
atm, standard atmospheric pressure; ATN rats, athymic nude rats; CCI, controlled cortical impact; FPI, fluid percussion injury; hESC, human embryonic stem cell; hiPSC, human induced pluripotent stem cells; hNSC, human neural stem cell; mNSS, modified Neurological Severity Scale; MWM, Morris Water Maze; PT, post-transplantation; SD rats, Sprague Dawley rats; TBI, traumatic brain injury.
Multiple studies included more than one hNSC experimental group for direct comparison of hNSC administration protocols including variations in cell dose, administration route, transplantation location, and intervention time window. Across the 20 studies, a total of 28 different experimental groups were identified. The timing of hNSC administration following injury varied substantially and was categorized into four periods: hyperacute (<30 min), acute (30 min to 24 h), subacute (72 h to 3 weeks), or chronic intervention (>3 weeks). Interestingly, 50% of the experimental groups received subacute transplantation, 32% received acute, 11% received hyperacute, and 7% received chronic intervention (Fig. 3A–B; Supplementary Table S3). Two studies began treatment 7 days post-TBI and repeated administration alternating daily from 9 to 17 days and were therefore classified within the acute administration group based on the first day of treatment. The dose of hNSCs varied among the 28 treatment groups and was classified as low (≤1 × 105), moderate (>1 × 105 to <1 × 106), or high (≥1 × 106) based on the total number of cells administered. The most commonly administered dose was moderate (50%), followed by low dose (32%), and high dose (18%; Fig. 3C; Supplementary Table S4). Nearly all experimental groups received hNSCs through direct brain injection (86%), while a small subset received systemic intravenous (IV) administration (7%), or intranasal (IN) delivery (7%; Fig. 3D; Supplementary Table S5). Among the experimental groups that received direct brain transplantation, hNSCs were delivered intralesionally in 54% of the groups and perilesionally in the remaining 46% (Fig. 3E; Supplementary Table S5). A full description of study characteristics and groupings based on administration protocols can be found in Table 3.
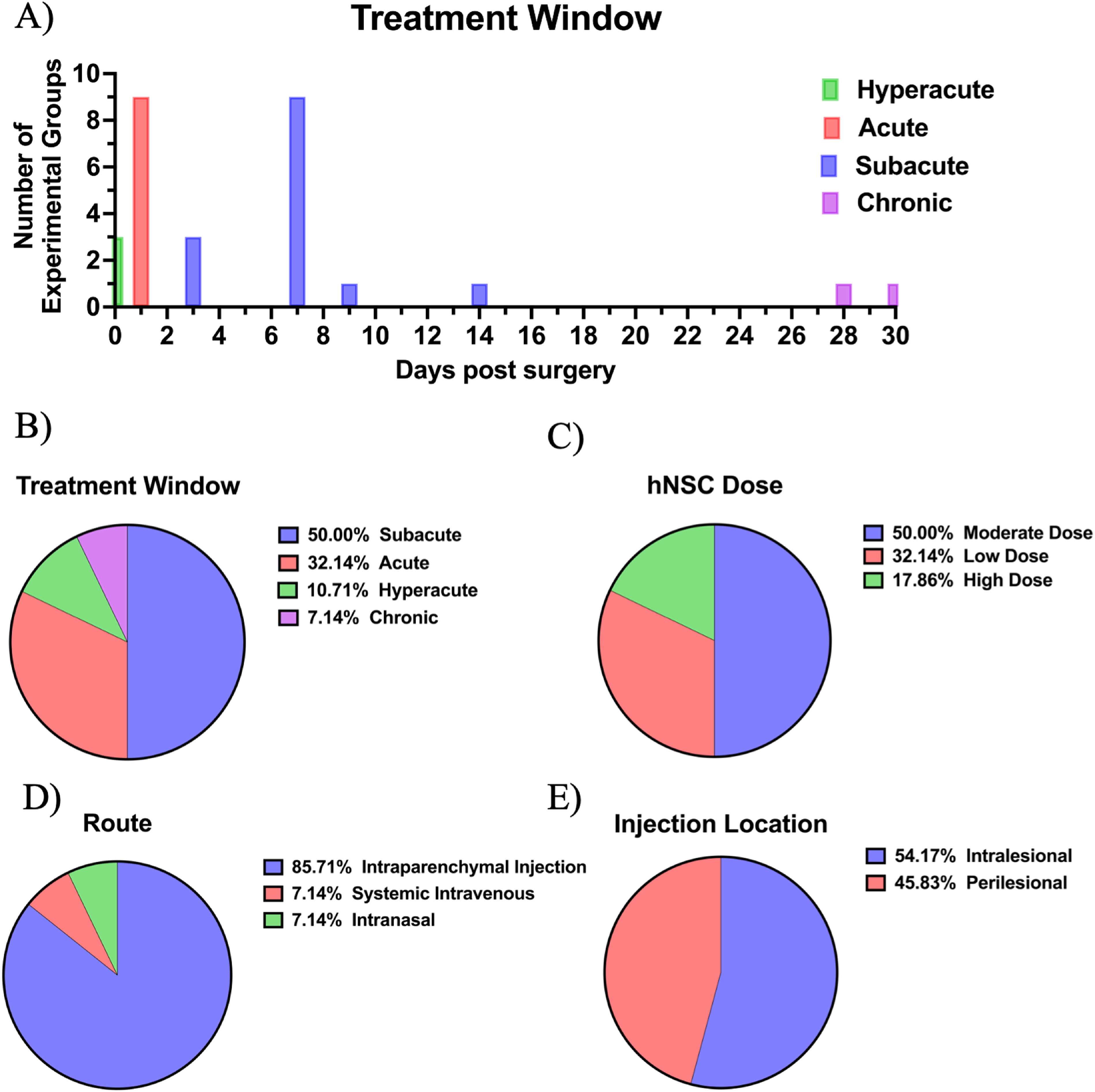
Study characteristics included within the systematic review related to hNSC administration. Data regarding the injury treatment window
Risk of bias
The SYRCLE’s RoB showed 75% of the included studies represented an unclear RoB based on incomplete or missing information, while 15% indicated high RoB (Fig. 4). Overall indications of bias for each article were assumed “high” if more than one individual parameter was identified as high risk. Specifically, details regarding the concealment of animal allocation within experimental groups and the randomization of animal housing were absent in all studies. The majority of studies specified random assignment of experimental groups; however, blinding of such groups varied across studies. Collectively, an unclear level of bias was determined. However, it is worth noting that these risks are likely attributed to the lack of detailed reporting in publication protocols as opposed to poor scientific design often due to publication limitations (e.g., word count, figure limits).

SYRCLE RoB tool revealed 16 studies with an “unclear” risk of bias and 4 of “high” risk of bias visualized per study
Meta-analysis and effect size of primary outcomes
The effect of hNSC treatment on TBI compared with vehicle administration was evaluated through a meta-analysis of four primary outcomes: lesion volume, MWM performance, mNSS functional scores, and rotarod performance. The SMDs between hNSC intervention groups and control groups were pooled across studies to identify the overall effect size. Z-scores revealed an overall positive, yet variable, effect of hNSC treatment on TBI-induced deficits (Table 4). Large effects were observed on lesion volume along with acute and chronic mNSS assessment scores. MWM and rotarod performance indicated a moderate effect in favor of hNSC intervention. Statistical heterogeneity ranged from 0% to 73%.
Summary of Mean Effect Sizes for Primary Outcomes
CI, confidence interval; mNSS, modified Neurological Severity Score; MWM, Morris Water Maze; SMD, standardized mean difference.
hNSC treatment leads to a reduction in lesion volume
Of the 20 studies, 11 reported the effect of hNSC treatment on lesion volume following TBI. However, two studies 20,33 were excluded due to insufficient data reporting within the publication and therefore a total of nine studies remained within the meta-analysis. 25 –27,34 –39 Lesion volume was expressed in five studies as an estimated volume in cubic millimeters 27,34,35,38,39 whereas the remaining four studies presented lesion volume as a percentage of the ipsilateral hemisphere. 25,26,36,37 All nine studies indicated an effect size in favor of hNSCs. Overall, the meta-analysis revealed a statistically significant reduction of TBI lesion volume following hNSC therapy with a pooled SMD of −1.09 (CI: [−1.74, −0.43], p = 0.001; Fig. 5). Significant statistical heterogeneity was observed (I 2 = 73%).
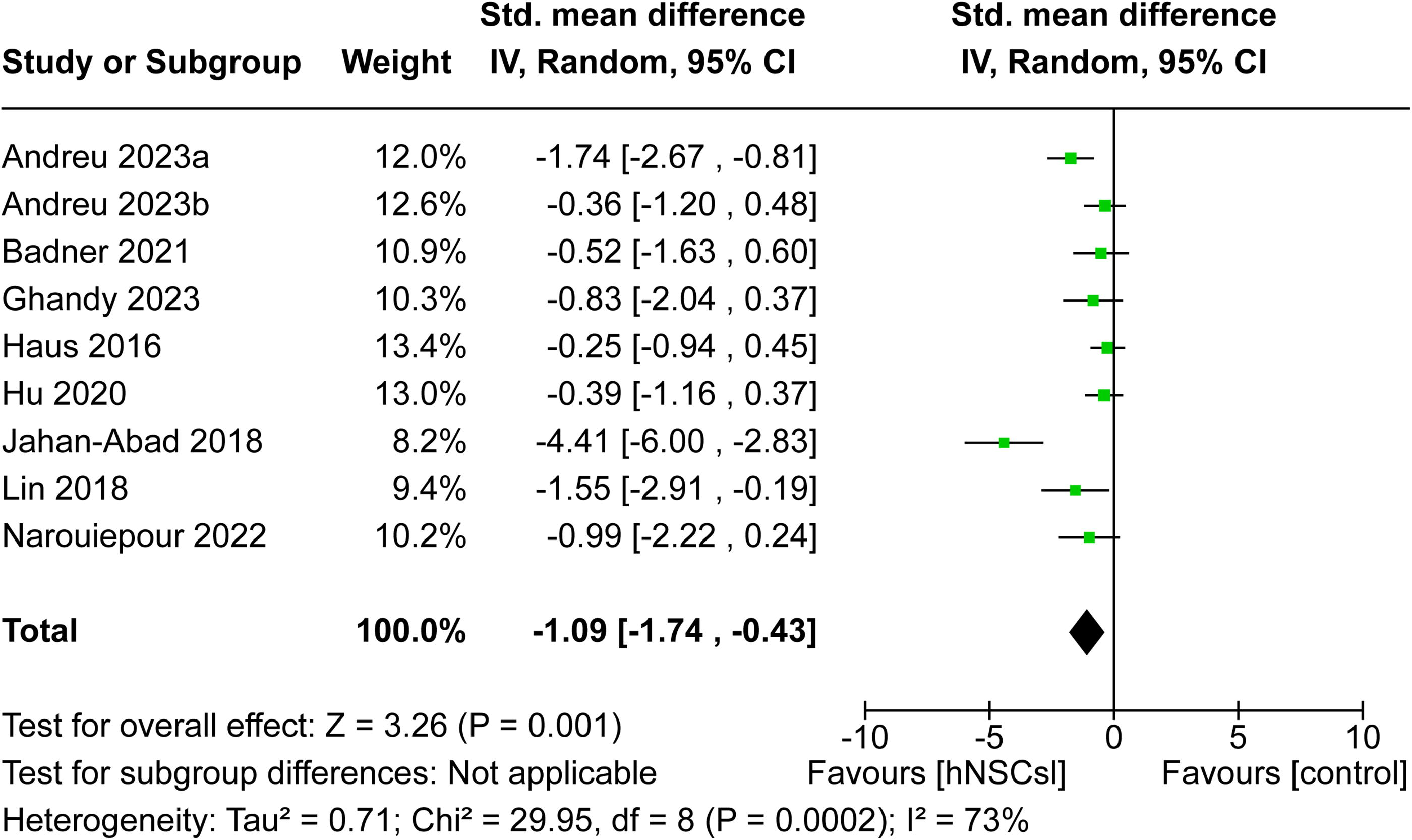
Overall effect of hNSCs on lesion volume after TBI. Forest plot revealed hNSCs significantly reduced lesion volumes in pre-clinical studies of TBI (SMD −1.09, CI: [−1.74, −0.43], p = 0.001). CI, confidence interval; hNSCs, human neural stem cells; SMD, standardized mean difference; TBI, traumatic brain injury.
hNSC treatment improves MWM performance
MWM performance was reported in six studies 19,34 –36,40,41 ; however, numerical values were unable to be retrieved from one study and therefore it was removed from the meta-analysis. 34 Escape latency on the last day of performance was extracted from each study for meta-analysis. The total time allotted for each animal to complete the MWM task varied among studies, as three studies allotted 60 sec 35,36,41 compared with 120 sec in two studies. 19,40 The meta-analysis forest plot revealed that hNSCs reduced the escape latency in each individual study, indicating an increase in spatial memory performance within the MWM task. The overall effect size of SMD −0.70 (CI: [−1.07, −0.33], p = 0.0002; Fig. 6) demonstrated that the MWM performance statistically favored hNSC intervention, suggesting hNSCs significantly improved cognitive recovery post-TBI. Statistical heterogeneity was not observed (I 2 = 0%).
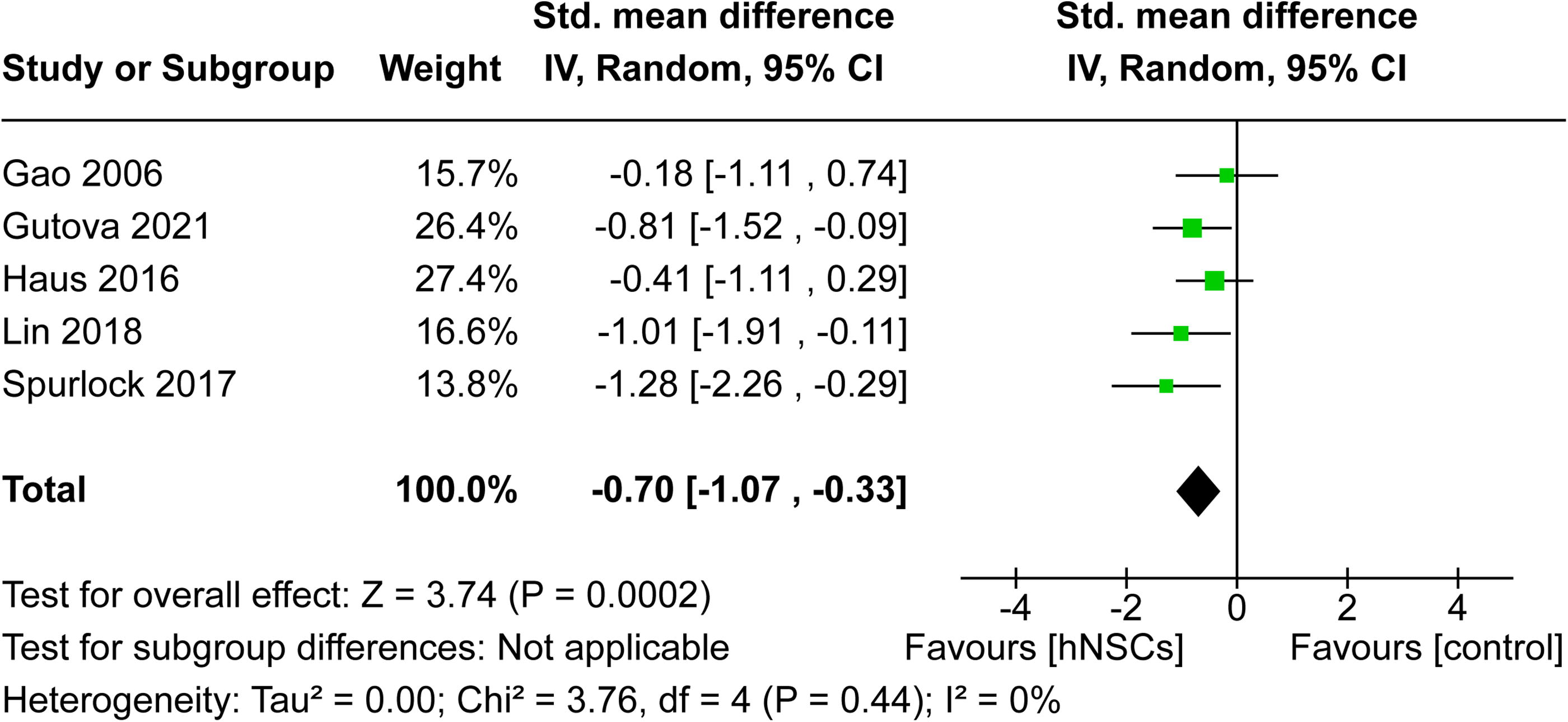
Overall effect of hNSC intervention on MWM performance post-TBI. Forest plot showed MWM performance was significantly improved by hNSC therapy (SMD −0.70, CI: [−1.07, −0.33], p = 0.0002). CI, confidence interval; hNSC, human neural stem cell; MWM, Morris Water Maze; SMD, standardized mean difference; TBI, traumatic brain injury.
hNSC treatment results in trending improvements in mNSS scores
A total of three studies were eligible for meta-analysis of the mNSS functional assessment. 27,39,42 Notably, three additional publications reported mNSS, but were excluded due to data extraction limitations. 33,38,43 mNSS was collected multiple times within each study. Therefore, the overall effect was analyzed at two time points: acute (5–8 days post-transplantation) and chronic (final time point reported). Two studies indicated favorable effects on mNSS scores at both time points in response to hNSC administration. 27,42 Alternatively, one study indicated mNSS scores were unaffected by hNSC intervention at acute and chronic time points. 39 The overall pooled effects revealed hNSCs reduced mNSS scores acutely (SMD −0.91, CI: [−1.85, 0.04]; Fig. 7A) and chronically (SMD −1.20, CI: [−2.47, 0.07]; Fig. 7B); however, the Z-test indicated these differences to be a non-significant trending difference (p = 0.06). This trending difference is likely due to the small sample size and significant statistical heterogeneity (I 2 = 70%).
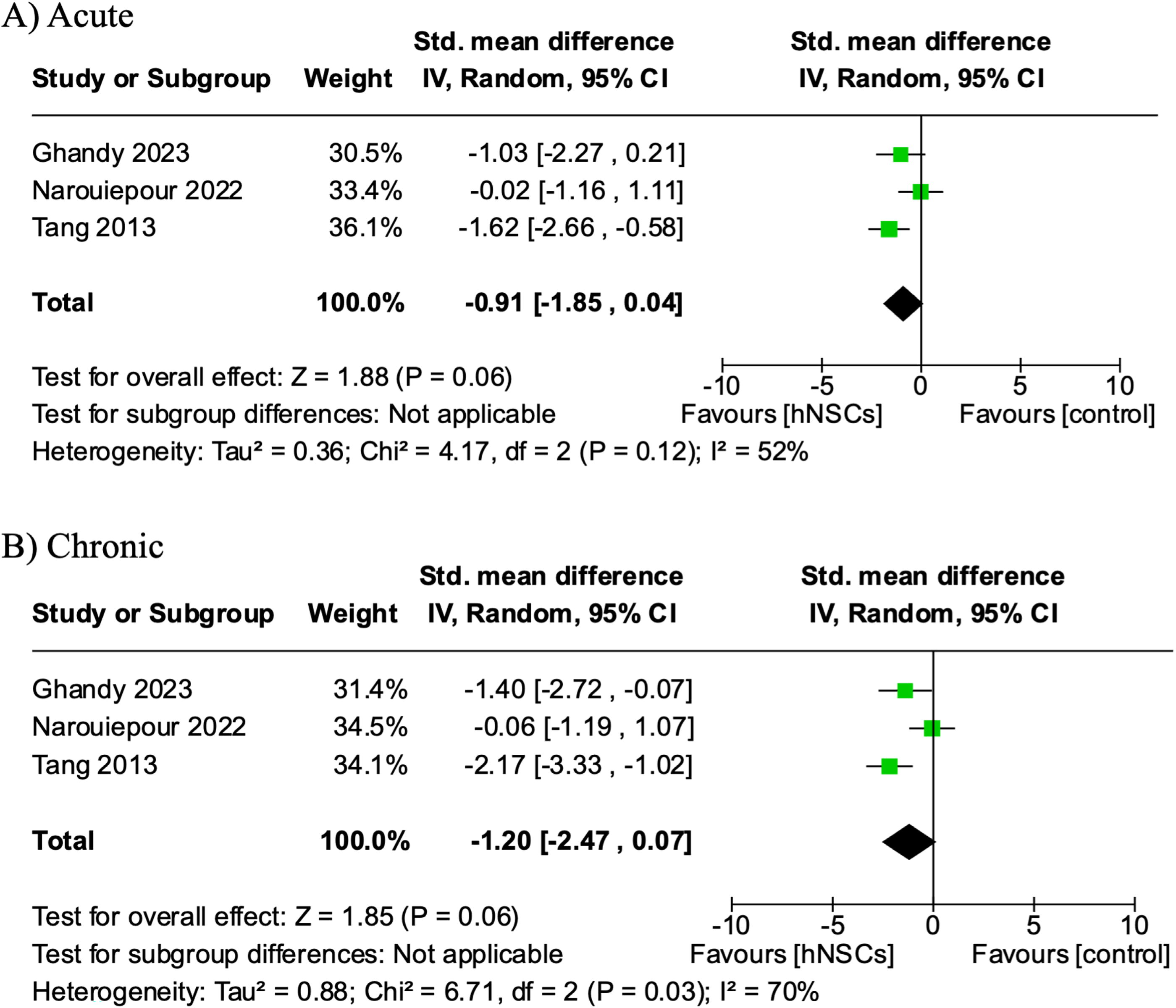
The effect of hNSC treatment on the mNSS neurological function assessment at acute and chronic timepoints post-TBI. A trending improvement in neurological function in TBI animals treated with hNSCs at acute (SMD −0.91, CI: [−1.85, 0.04, p = 0.06])
Rotarod task performance was not significantly improved by hNSC treatment
The rotarod task was used to evaluate motor function recovery at multiple time points post-TBI in five studies. 27,28,36,39,42 Performance was recorded as the latency to fall in four studies 27,28,36,39 and as an endurance score in one study. 42 Rotarod performance was analyzed at two distinct time periods: acute (5–8 days post-transplantation) and chronic (last reported evaluation). Meta-analysis revealed motor function performance favored (not statistically significant) hNSC intervention to vehicle administration at acute time points (SMD −0.29, CI: [−0.69, −0.10], p = 0.15; Fig. 8A). Similarly, evaluation of chronic rotarod performance indicated five out of 6 studies individually favored hNSC treatment, yet again this was not statistically significant with an overall effect of −0.35 SMD (CI: [−1.09, 0.38]; p = 0.34; Fig. 8B). I 2 values indicated low acute heterogeneity (0%) and moderate chronic heterogeneity (66%).
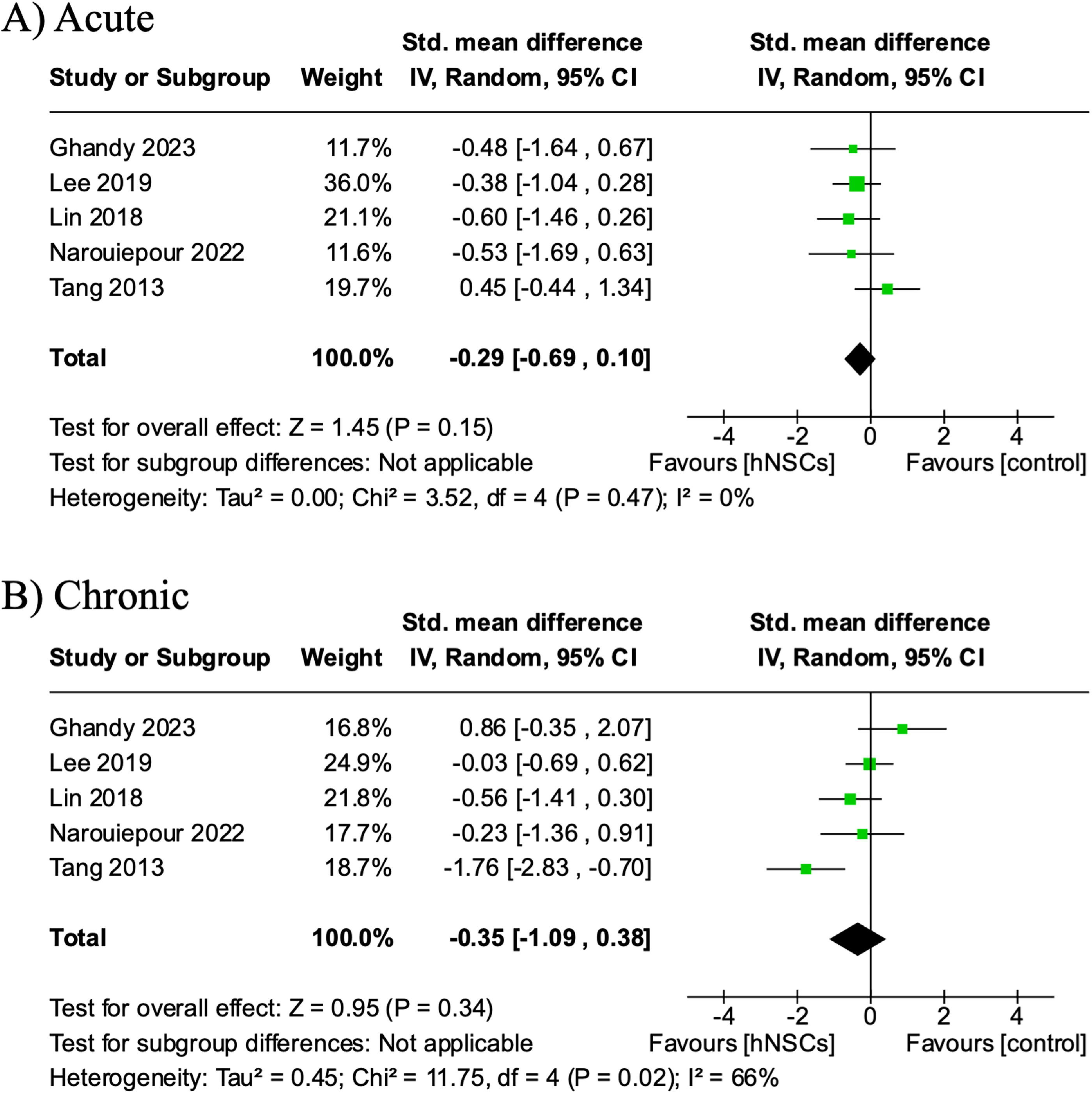
The effect of hNSCs treatment on rotarod motor function performance at acute and chronic timepoints post-TBI. Rotarod performance at acute (SMD −0.29, CI: [−0.69, 0.10], p = 0.15])
Changes in cellular outcomes in response to hNSC treatment
A number of important cellular outcomes found to be ineligible for meta-analysis due to heterogeneity across studies and lack of adequate reporting included exogenous hNSC survival, engraftment and differentiation within the host TBI microenvironment, neuroprotection of neural cell populations, axonal integrity following injury, and inflammation. These outcomes are important measures of the multimodal mechanism of action (MOA) underlying hNSC intervention and are therefore important to consider in this review despite being unable to undergo meta-analysis.
hNSCs survive and differentiate into neural cell types within the rodent TBI brain
The survival, migration, and differentiation patterns of exogenous hNSCs were reported in 17 of the 20 studies (Supplementary Table S6). Notably, 15 publications reported successful survival of hNSCs upon completion of the study and two studies were unable to visualize engraftment. 33,43 Skardelly et al. identified hNSCs in TBI brain tissue when delivered via intraparenchymal transplantation. 33 However, in the same study, hNSCs administered systemically were not found in TBI brain tissue, suggesting a delivery modality-dependent effect on engraftment and survival 84 days post-treatment. Within a follow-up study, intraparenchymal and systemically delivered human neural progenitor cells (hNPCs) were unidentifiable on days 7, 28, or 84 post-delivery despite the successful identification of TAMRA+ hNPCs within the pilot arm of the study. 43 The methods and reported outcome measures utilized to evaluate hNSC survivability post-TBI varied immensely across the included studies. Specifically, nine studies visualized survival and engraftment through pre-labeling of cells with green flourescent protein (GFP), BrdU, red flourescent proteins DS-Red or PKH-26, tetramethylrhodamine (TAMRA), or superparamagnetic iron oxide (SPIO). Alternatively, seven studies identified engraftment through immunohistochemical analysis utilizing human-specific antibodies including STEM121, lysosomal associated membrane protein 2 (LAMP2), anti-human Nestin, anti-SC121, neural cell adhesion molecule (NCAM), HuNu, or HNA. Exogenous hNSC survival, migration, and differentiation were evaluated at various time points post-administration, ranging from acute (0 days) to long-term (20 weeks). Within the 17 articles, 2 reported subacute hNSC engraftment (6 days and 2 weeks post-transplantation), 9 reported long-term hNSC engraftment (5, 12, and 20 weeks post-administration), and six evaluated hNSC survival repeatedly at more than one time point ranging from 3 days to 16 weeks with all six studies showing engraftment at their latest time points. Furthermore, the percentage of surviving hNSCs ranged from 3.7% to 53.4%. However, most publications reported qualitative evidence of hNSC survival and engraftment within the host tissue rather than quantitative metrics.
The differentiation capacity of hNSCs in the TBI environment was evaluated in nine studies, 19,20,33 –37,41,44 five of which did so quantitatively 34 –37,41 (Supplementary Table S6). These studies showed that the majority of hNSCs differentiated into DCX+ (22.9% to 78.1%) neuroblasts or neuronal nuclei + (NeuN) (18.0% to 43.0%) or beta tubulin III + (Tuj1) (44.0%) neurons. These studies also demonstrated that hNSCs had a limited capacity to differentiate into astrocytes or oligodendrocytes with four studies identifying glial fibrillary acidic protein + (GFAP) astrocytes populations ranging from 13.0% to 22.9% and two studies finding Olig2+ oligodendrocytes populations ranging from 11.0% to 16.4%. Furthermore, three studies identified Nestin+ hNSC populations ranging from 3.4% to 29.2% suggesting that a number of hNSCs remained in an undifferentiated state even after 20 weeks of differentiation.
Independently, seven studies evaluated the effect of treatment protocols (i.e., hNSC dose, location, passage, route) on cell survival and engraftment 25,26,28,33,35,37,43 (Supplementary Table S6). Haus and colleagues interestingly observed low passage hNSCs (passage 7) increased, although not statistically significant, cell survival at 20 weeks post-transplantation compared with high passage hNSC (passage 27) from 8.7% to 24.9%, respectively. 35 Andreu et al. noted a dose-dependent response to engraftment, in which the high dose (1.6 × 106 hNSCs) significantly increased the percentage of GFP+ hNSCs located in the ipsilateral (TBI affected) hemisphere compared with the low dose (1.6 × 105 hNSCs). 25 In a follow-on study, Andreu et al. differentially concluded the administration interval relevant to injury did not statistically alter cell survival as hNSCs delivered 1, 2, or 4 weeks post-TBI demonstrated similar quantities of GFP+ hNSCs. 26 Similarly, Hu et al. demonstrated cell engraftment was unaffected by transplantation location, intralesionally versus perilesionally. 37 Taken together, these studies suggest passage number and cell dose could influence hNSC engraftment in addition to administration timeframe or transplantation location, however future contemporary studies directly evaluating these variables are required.
hNSCs induce neuroprotection and promote endogenous neuron survival and axonal integrity
hNSC-induced neuroprotection of endogenous neuronal populations was evaluated in six studies. 28,33 –35,43,44 Of these, 50% utilized Nissl or Cresyl violet staining and 50% stained for NeuN. Endogenous neuron populations were quantified as the positive cell count within a specific region, the percentage of positive staining compared with the contralateral hemisphere, or the percentage of positive cells within a specified area. Overall, the six studies indicated a positive effect of hNSC treatment on increasing endogenous neuron survival compared with vehicle controls (Supplementary Table S7). Badner et al. reported hNSC treatment significantly increased ipsilateral cornu ammonis (CA) neurons. 34 Haus et al. observed neuron survival to be affected by passage number with high passage hNSCs (passage 27) significantly increasing endogenous neuron populations within the ipsilateral DG and CA regions and low passage hNSCs (passage 7) inducing no change in neuron survival. 35 Lee et al. indicated a dose-dependent effect on ipsilateral neuron populations, where moderate and high doses significantly increased neuron survival, whereas the low dose resulted in a non-significant increase compared with controls. 28 Skardelly et al. found contradictory results in two studies evaluating hNSC treatment effect on neuron survival in TBI tissue. 33,43 In 2011, Skardelly et al. found systemic hNSC administration significantly preserved neuron counts at the lesion border; however, locally administered hNSCs resulted in a non-significant increase compared with controls at 12 weeks post-treatment demonstrating a delivery-route effect. 33 Later in 2014, a follow-on study by Skardelly et al. showed neuron density was reduced in systemic and intraparenchymal injection hNSC treatment groups compared with control groups. 43
Axonal damage was reported in three studies, all of which reported a comprehensive reduction of axonal injury following hNSC administration 20,21,26 (Supplementary Table S8). Measurements of axonal damage included acute accumulation of amyloid precursor protein (APP) immunoreactive profiles at 3 or 6 days post-transplantation 20,21 or chronic silver staining density at 12 weeks post-administration. 26 Furthermore, two studies quantified acute APP expression through western blot analysis at 3 or 6 days post-transplantation. 20,21 All three studies demonstrated hNSC treatment significantly decreased profiles of axonal damage identified through silver staining density or APP accumulation. Interestingly, western blot analysis produced conflicting results as Gao et al. observed a significant reduction of APP expression in response to hNSC treatment 19 ; however, Wang et al. concluded APP expression to be unaffected by hNSCs following TBI despite reduced APP density. 21 Andreu et al. reported the reduction of axonal injury corresponded with a shorter intervention period. 26 Specifically, hNSC delivery at 1 and 2 weeks post-TBI significantly decreased silver staining density; however, mitigation of axonal damage was unaffected by chronic hNSC administration 4 weeks post-TBI when compared with control animals. Collectively, these results suggest hNSC intervention reduces axonal injury potentially in a delivery time frame dependent manner.
hNSCs mitigate the TBI immune cascade
The immunomodulatory properties of hNSC treatment were explored in 11 studies; however, the heterogeneity of time points, methods, and animal immune states prevented a formal statistical analysis 20,25,27,28,33,34,38,39,43 –45 (Supplementary Table S9). The immune system of the model species plays a critically relevant role in the context of post-TBI inflammation with increased inflammation resulting in higher levels of cell death. Immunosuppression was used in 5 of the 11 studies, 4 did not specify, and 2 utilized immunocompetent animals. Within the 11 studies, 5 clearly concluded hNSC treatment significantly dampened the inflammatory cascade post-TBI. The remaining six studies alternatively presented neutral or negative effects of hNSCs on TBI-induced inflammation. Amirbekyan et al. concluded hNSCs downregulated inflammatory genes and pathways; however, hNSC treatment did not affect CD68+ or CD163+ macrophage cell counts compared with vehicle animals. 45 Similarly, two studies reported by Ghandy et al. and Narouiepour et al. observed reduced toll-like receptor 4 (TLR4) and nuclear factor kappa B (NF-kB) signaling molecules, yet Iba1+ microglia populations showed no difference between treated and non-treated animals. 27,39 Skardelly et al. observed hNSCs transiently mitigated inflammation through acute reductions of GFAP expression, but the observed effect was absent by 84 days post-transplantation. 33 hNSC therapeutic concerningly induced a pro-inflammatory response in two studies. 43,44 A sex-dependent response was observed by Nieves et al., where hNSC-treated females had increased populations of reactive astrocytes compared with controls, yet a slight decrease was observed in males compared with controls. 44 The opposite effect was observed in Iba1+ microglia populations as males demonstrated increased microglia presence compared with an observed reduction in the females. Likewise, Skardelly et al. revealed systemic delivery of hNSCs significantly increased CD68+ and CD11b+ macrophage recruitment to the lesion site and basal ganglia at chronic time points. 43 Overall, these results support the suggested anti-inflammatory mechanism of hNSCs within the TBI brain. However, future studies are needed to elucidate specific pathways involved to fully understand the sex-dependent response observed.
Discussion
Numerous studies support that hNSC treatment has multimodal neuroprotective and regenerative capabilities resulting in improved functional recovery in TBI animal models. However, the ability of hNSCs to perform as a cell replacement therapy (including hNSC survival, engraftment, and differentiation) and to create a neuroprotective and regenerative environment through paracrine signaling has been in question due to discrepancies in results between pre-clinical studies. In this systematic review and meta-analysis, hNSC treatment led to improved tissue preservation (lesion volume), cognitive function (MWM), and trending improvements in neurological recovery (mNSS) following TBI. Additionally, hNSCs were capable of long-term survival, differentiation, and engraftment within TBI tissue and modulation of the TBI microenvironment to enhance endogenous neuron survival, reduce axonal damage, and decrease the immune response. The therapeutic effect of hNSCs in TBI was observed across several publications utilizing cells from four unique sources (e.g., fetal, iPSC) at different doses and routes of administration in varying types of TBI models (e.g., CCI, pTBI) in both rat and mouse models. The significant therapeutic effect of hNSC treatment in such a highly variable group of TBI studies indicates that hNSCs are likely a robust therapeutic option capable of moving toward human clinical trials.
hNSCs act through neuroprotection
This systematic review revealed abundant evidence supporting the neuroprotective effect of hNSCs in rodent TBI models, likely through the release of trophic factors. Within the clinical setting, lesion volume is acknowledged as a predictive indicator of long-term recovery as it is highly correlated with the GCS, neuropsychological index scores, and cognitive function in TBI patients, and therefore is a valuable measure of therapeutic efficacy for pre-clinical models. 46 –48 This meta-analysis of relevant studies showed an overall significant reduction of lesion volume in hNSC-treated animals compared with non-treated controls. The largest effect size (SMD −4.41) was observed following immediate administration of hNSCs post-TBI 38 and the smallest individual study effect sizes (SMD −0.25, −0.36, −0.39) were observed following subacute or chronic hNSC administration. 26,35,37 hNSC TBI intervention also correlated with improved endogenous neuron survival and decreased axonal damage. Overall, the evaluated studies indicated hNSC treatment increased neuronal survival within hippocampal regions and the ipsilateral cortex as well as significantly reduced axonal injury.
Although the studies identified here provide substantial evidence of the neuroprotective effect of hNSC intervention on lesion volume, neuronal survival, and axonal integrity post-TBI, most failed to sufficiently address the specific MOAs apart from immunomodulation. The current body of literature reports the secretion of neurotrophins such as brain-derived neurotrophic factor (BDNF), glial cell line-derived neurotrophic factor (GDNF), and nerve growth factor from hNSCs within the injured brain induce the “bystander effect” that prevents the progression of the secondary injury cascade and improves functional recovery. 49,50 Evidence supporting the secretion of neurotrophic factors by hNSCs was found explicitly within the included studies here, which revealed integrated hNSCs not only express GDNF and BDNF in vitro but also significantly increase rat endothelial cell antigen expression, an indirect measure of angiogenesis. 19,21,33 These findings are consistent with the body of literature which suggest NSCs preserve vascular networks in the brain and promote angiogenesis through vascular endothelial growth factor, and angiopoietin-1. 15,51 However, future in vivo studies evaluating hNSC MOAs are needed to fully uncover the signaling factors at play.
hNSCs act as a source of cellular replacement
The TBI brain is characterized by both necrosis, the passive disruption of homeostasis leading to cellular death, and apoptosis, the active programmed cell death. 52 Inevitable neuronal loss following TBI is linked to chronic functional deficits and mortality. 53,54 Therefore, the ability of hNSCs to act through cellular replacement highlights a key strength which remains a limitation for many other therapeutic interventions. The long-term survival, migration, and differentiation of transplanted hNSCs within the rodent brain following TBI were first reported by the Mathiesen group in 2004. 24,55 Providing insights into the feasibility of hNSC survival in the rodent TBI brain, Wennersten et al. and Nimer et al. administered hNSCs immediately after injury. However, the full therapeutic capacity of hNSCs remained ambiguous due to the relatively short study period (6 weeks) and lack of functional outcomes evaluated. Over the next 20 years, studies identified within this review collectively elucidated a much broader scope of hNSC survival, migration, and differentiation within the TBI microenvironment. Notably, 15 out of the 20 included publications, all with various treatment administration protocols ranging from immediate to 4-week transplantation intervals, reported sufficient hNSC survival and differentiation up to 20 weeks post-transplantation. Not only were engrafted hNSCs identified at the transplantation site and lesion border, but it is also well-documented that hNSCs migrate beyond these locations by crossing the midline as far as the contralateral hippocampus and cortex, likely following axonal tracts. Acutely, hNSCs possessed markers of proliferation (Ki-67) and early differentiation (DCX) within the host. As the interval between administration and histological analysis progressed, markers of proliferation and early neural differentiation decreased and hNSCs reached terminal differentiation identified by NeuN+ and Tuj1+ expression. Lin et al. further revealed that engrafted hNSCs successfully integrated into the host neuronal network through calcium imaging and neural tracing experiments. 36 Unlike the well-documented differentiation of transplanted hNSCs into neurons, reports of oligodendrocytes and astrocytes originating from the engrafted hNSCs varied with further evaluation of glial differentiation being needed to advance the field.
Despite robust evidence supporting hNSC engraftment and differentiation in the TBI brain, the extent to which hNSC cellular replacement contributes to corresponding tissue preservation and functional recovery is highly debated. Here, meta-analysis revealed a non-significant improvement in acute neurological and motor function in favor of hNSC treatment through analysis of the mNSS assessment and rotarod performance 5–8 days post-transplantation. Additional functional measures reported significant improvements observed in the swing bias, paw grasp, and open field testing as early as 7 and 14 days post-transplantation. 28,39 This observed functional recovery likely occurs prior to engrafted hNSCs achieving functional integration. Findings from Andreu et al. revealed a significant relationship between lesion volume and transplanted cell dose, but not between lesion volume and GFP+ engraftment volume. This suggests an alternative hNSC MOA, rather than cellular replacement, is responsible for mitigating tissue atrophy. 25 Hu et al. similarly showed lesion volume reduction to be unrelated to engraftment volume. 37 In this study, the results indicated perilesional delivery significantly decreased lesion volume compared with intralesional transplantation. However, GFP+ cell counts representative of overall engraftment at 12 weeks post-transplantation did not statistically differ between the two administration locations. Taken together, these studies suggest the observed hNSC-induced tissue preservation and functional recovery post-TBI likely stems from a paracrine signaling mechanism(s) rather than cellular replacement as the primary contributor.
hNSCs modulate the immune response
Targeting of the inflammatory cascade is a key component of many pre-clinical trials aimed at developing therapeutic interventions for TBI patients. Moreover, immunomodulation is one of the most widely accepted neuroprotective mechanisms of hNSC intervention. The diversity of the reported methods and outcomes evaluating inflammation within this systematic review prevented the use of a meta-analytic approach. However, including it as a secondary outcome provides critical insight into the therapeutic efficacy of hNSCs in pre-clinical TBI models. Within minutes following initial mechanical impact, damage-associated molecular patterns are rapidly released in response to cellular damage, BBB breach, and disruption of the central nervous system vasculature. 56,57 Consequentially, resident microglia, the endogenous immune cells of the brain, activate thus producing cytokines and chemokines such as interleukin 1 beta (IL-1β), interleukin 6 (IL-6), and C-X-C chemokine receptor type 4 (CXCR-4), which promote peripheral immune cell infiltration toward the damaged tissue. 58 This injury-induced inflammatory cascade promotes both the M1 classical pro-inflammatory and alternative M2 anti-inflammatory response. 20 The resulting inflammatory injury can persist chronically for months, even years post-TBI, further contributing to neuronal death and axonal degeneration which are responsible for chronic neurological deficits. 20,59
The literature here supports the hypothesis that hNSCs modulate the post-TBI inflammatory response by reducing resident microglia activation and promoting the transition of a pro-inflammatory M1 phenotype to an anti-inflammatory M2 phenotype. The unique methodology employed by Andreu et al. demonstrated a significant reduction in microglia activation through quantification of morphological presentation using Sholl analysis. 25 Specifically, vehicle animals exhibited reduced Sholl intersections as activated microglia in the ameboid state lack filament projections. In contrast, sham and high-dose hNSC-treated animals demonstrated increased Sholl intersections suggestive of the inactivated ramified morphology. This supports the role of hNSCs in mitigating microglia activation at both acute (as early as 6 days post-transplantation) and chronic (12 weeks post-transplantation) time points. Gao et al. further elucidated that the observed reduction of Iba1+ microglia coincided with a 48.4% reduction in M1 phenotypic cells and an 81.8% increase in M2 phenotypic cells. 20 The authors corroborated this hNSC-mediated transition with the reduction in axonal injury as pro-inflammatory M1 activity was recently linked to axon damage and M2 activity was linked to axon regeneration following spinal cord injury. 60 Only two other studies evaluated axonal damage; however, neither of which also explored the link to inflammatory outcomes. Therefore, further research is needed to explore if this proposed relationship between inflammatory phenotypes and axonal damage holds true in TBI and if hNSC-reduced axonal injury occurs through inflammatory modulation.
It should be noted that two alternative studies evaluated the presence of macrophages post-TBI and reported hNSC treatment produced no change or even increased CD68+, CD11b+, or CD163+ cells within the lesion area. Amirbekyan et al. intranasally delivered hNSCs beginning 7 days post-TBI and quantified CD68+/CD163+ macrophage populations 32 days after the start of treatment. 45 Although hNSC treatment yielded no change in macrophage presence compared with vehicle animals, nanostring analysis revealed hNSC treatment downregulated inflammation-related genes specific to microglial function and cytokines (i.e., CDh1, Lyz2, PDGFRb). One explanation for the lack of changes in macrophage populations observed in this study may be that the time point of evaluation was beyond the typical acute macrophage infiltration period. Comparatively, Skardelly et al. observed a pro-inflammatory hNSC response as systemic delivery of hNSCs significantly increased CD68+/CD11b+ macrophages 84 days post-treatment. 43 Interestingly, intralesional delivery of hNSCs in this study transiently induced a non-significant increase of macrophage presence, yet significantly reduced GFAP+ reactive astrocytes at the lesion border. The authors speculated this was due to an ongoing graft rejection as this observation corresponded with a significant decrease in NeuN+ neurons at the lesion border. Moreover, it is imperative to acknowledge these results in the context of immunosuppression, as over 70% of the included studies noted the use of an immunosuppressant and an additional 20% do not clearly provide the model’s immune state. Further research is therefore required to fully elucidate the full capacity of hNSCs to modulate the TBI inflammatory cascade in models more representative of the human immune system.
Identification of knowledge gaps
Through the consolidation of this systematic review and meta-analysis, several gaps within the literature arose. It must be acknowledged that the goal of these pre-clinical studies is to provide sufficient evidence to support moving hNSCs into the clinical setting for TBI patients who are currently without access to FDA-approved therapeutics. Therefore, it is imperative to discuss the limitations of the included studies and gaps within the field that ultimately prevent hNSC progression to the bedside. One of the largest challenges preventing this progression is the variability in cell survival and migration, with numerous studies reporting insufficient engraftment within the neurotrauma field. 16,33,43,61 Notably, the first in-human clinical trial evaluating hNSCs in a patient population with a neurological disorder (neuronal ceroid lipofuscinoses) did not observe any transplantation-related adverse effects. 62 However, the transplanted cells showed limited migration beyond the transplantation site. Although the studies highlighted in this review demonstrate successful long-term engraftment and sufficient migration beyond the injection site into the contralateral hemisphere, it is important to acknowledge the fundamental species differences, particularly the overall size and therefore required distance of migration, between rodent and human brains which could be driving the observed data discrepancies. Specifically in pre-clinical TBI models, studies evaluating hNSC efficacy have not yet expanded beyond small animals. Utilizing rodents in biomedical research is beneficial in that they are relatively inexpensive, easy to handle, and well-validated across various disease paradigms and outcomes measures. However, the majority of neuroregenerative therapies tested in rodents fail to successfully translate to humans. Pertaining to TBI, this is likely due to the lissencephalic structure and reduced white-to-gray matter ratios that lead to unique pathology and recovery responses in rodents compared with humans. 63,64 One opportunity to fill this gap and accordingly increase the translatability of hNSCs is to use large animal TBI models such as pigs or non-human primates that possess gyrencephalic brains and more similar neural anatomy and physiology to humans. 65,66
Second, xenotransplantation of hNSCs into non-human models may result in outcomes that are not reflective of allogeneic hNSC transplantation into human patients due to species-specific differences in cell signaling proteins, receptors, signaling doses, spatial and temporal controls, innate capabilities of transplanted cells (e.g., neurite length and branching), and other key aspects that are crucial for hNSC differentiation and engraftment. Exploration of allogeneic NSC treatment with mouse NSCs (mNSCs) in mouse TBI models supported the findings of hNSC xenotransplantation TBI studies where mNSCs led to significant mitigation of TBI pathophysiology and enhanced functional recovery in mouse models. 67 –70 These studies showed mNSC treatment led to improvements in MWM, mNSS, and rotarod task performance, which were primary outcomes of this systematic review. Moreover, mNSC treatment led to significant improvements in secondary outcomes including improved endogenous neuron survival and decreased axonal injury and inflammation. However, more in-depth cross-species comparisons must be made to better understand innate species differences, particularly those in which cell origin and recipient recovery responses and cross-species immune rejection are thoroughly evaluated. This further highlights the need for testing in more predictive large animal TBI models in order to gain a better understanding of the therapeutic potential of hNSCs for patients.
Within the field, the optimal administration protocol for hNSC delivery lacks consensus. Very few studies perform direct comparisons of administration techniques, such as cell dose, treatment window, administration route, or transplantation location, while controlling for all other variables. Drawing conclusions across publications is further limited as high heterogeneity between study designs typically results in multiple variations of therapeutic intervention (i.e., cell dose and delivery interval). However, six studies identified here independently evaluated cell dose, administration time, location, and route. However, typically only one or two studies experimentally compared each therapeutic intervention variable (e.g., cell dose).
The characteristics related to the animal models within the identified studies failed to accurately represent the population of human TBI patients. Particularly, 15 of the 20 studies utilized adult male animals only. Although TBI prevalence is higher in males, females account for 41% of TBI-related medical visits in the United States and often display differences in TBI pathology and outcomes in relation to males. 44,71,72 Females remain heavily under-represented accounting for about 40% of participants in early-stage clinical trials, 73,74 despite recent efforts by the National Institutes of Health and other governing bodies to include both sexes in pre-clinical and clinical studies. Furthermore, age is reported as one of the highest independent predictors of mortality and recovery following TBI. 75,76 Most pre-clinical therapeutic intervention studies focus on adults despite the increased prevalence of TBI in pediatric and geriatric populations. The immature and geriatric brains display age-dependent differences related to neurodevelopment, programmed cell death, synaptic reorganization, neuroinflammation, and neurogenesis. These unique responses following TBI have been historically overlooked in the study design of many TBI pre-clinical studies. 77,78 The last aspect of population characteristics neglected within pre-clinical studies identified here, and within the literature as a whole, is the presence of co-morbidities prior to injury. Future studies must take these factors into consideration and perform direct comparisons in order to identify sufficient treatment protocols for the diverse population of TBI patients.
Limitations
Several limitations were identified within the systematic review. First, significant heterogeneity was observed across the evaluated studies. Variations of study design (i.e., species, injury induction, immunosuppression), hNSC administration protocols (i.e., origin, delivery interval, route and location, dose), and outcome measures (i.e., time point, technique, reported units) further confounded the magnitude of the meta-analysis due to relatively small samples sizes. Moreover, the meta-analysis was performed using SMD rather than the mean difference (MD) technique as the evaluated outcomes reported various units across all studies. There exists substantial debate surrounding SMD versus MD. However here, where the outcome scales are not easily translatable directly to patients, SMD may be more meaningful in determining the biological significance. The SYRCLE assessment likewise identified an unclear RoB pertaining to treatment group blinding and failure to report protocol details in the article. Finally, this systematic review did not address combination therapies that support NSC survival post-transplantation including the use of novel biomaterials, nanoparticles, and scaffolds. In recent years, co-transplantation of NSCs and scaffolds has gained significant attention, particularly in an effort to enhance cell engraftment and viability following transplantation. 27,38,79 –82 Specifically, scaffolds have been shown to significantly enhance the retention of NSCs by adhering cells to the site of injury and preventing them from being removed by the flow of cerebral spinal fluid. 38,81 Moreover, scaffolds can be manufactured with a number of TBI recovery factors including BDNF and GDNF that promote NSC survival in the cytotoxic TBI microenvironment. 82 However, the ability of such complementary biomaterial approaches to enhance the therapeutic efficacy of hNSCs and specifically address a major gap in the field of enhancing cell retention and survival highlights the continued need for ongoing studies and an opportunity for future systematic reviews to expand upon.
Conclusion
For the first time, this review systematically identified studies assessing the efficacy of hNSC therapy in pre-clinical TBI models from October 1, 2002 to June 1, 2024. Meta-analysis revealed hNSC TBI treatment led to significant and favorable effects on neuropathology and cognitive and neurological functional recovery. Additional evaluation of secondary outcomes showed hNSC engraftment and integration into the host network increased neuronal survival, reduced axonal damage, and decreased the inflammatory response. However, pre-clinical trials must address concerning levels of internal and external bias moving forward. This can be achieved by the standardization of protocol reporting, improving study design with appropriate blinding, and encouraging researchers to reference the SYRCLE guidelines in their publications. Additionally, future studies evaluating hNSC efficacy, underlying MOAs, and optimal administration techniques conducted in animal models with increased translational characteristics that are more representative of patients (i.e., large animals with appropriate white-to-gray matter ratios, split-sex, pediatric or geriatric age, and co-morbidities) are necessary. Together, the therapeutic efficacy of hNSC intervention demonstrated within these pre-clinical studies combined with follow-on investigations addressing the limitations and knowledge gaps identified here may aid in moving hNSC therapy into the clinical setting in the near future and providing TBI patients with an effective treatment option.
Transparency, Rigor, and Reproducibility Statement
The reporting of this systematic review was performed in accordance with the Cochrane Handbook for Systematic Reviews and based on the PRISMA statement. Additionally, the analysis plan was pre-specified by the lead and corresponding author prior to the literature search using the PICO framework. The resources and software used in this study are clearly identified and openly available. Statistical analysis, including the handling of missing data, was performed according to the guidance provided within the Cochrane Handbook. The primary outcome measures used are well-established in the field. A replication study has not yet been planned at the time of writing. This article will be published under a Creative Commons Open Access license, and upon publication will be freely available.
Footnotes
Authors’ Contributions
S.L.S.: Conceptualization, methodology, software, validation, formal analysis, data curation, writing, visualization. K.J.D.: Writing—reviewing and editing, supervision. E.E.K. and F.D.W.: Conceptualization, resources, writing—reviewing and editing, supervision, project administration.
Author Disclosure Statement
The authors declare no potential conflicts of interest.
Funding Information
There was no funding provided for this research.
Supplementary Materials
Supplementary Data
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
Supplementary Table S6
Supplementary Table S7
Supplementary Table S8
Supplementary Table S9
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.