
Abstract
Respiratory failure is one of the greatest causes of morbidity and mortality after cervical lesions, the most common type of spinal cord injury (SCI). Fortunately, several pre-clinical and clinical studies have shown spontaneous, but limited, respiratory recovery after injury. However, there are still many unanswered questions about what is driving this recovery, so there is a growing need to further elucidate the neuroplastic potential of the phrenic network. Here, we investigated the structural plasticity of the right and left phrenic networks by analyzing perineuronal net (PNN) changes after a C2 hemisection (C2Hx) in mice. For this purpose, the right and left phrenic systems were traced with a pseudorabies virus, a trans-synaptic retrograde tracer applied to the diaphragm muscle, labeling the entire phrenic motor network. We found most PNN-bearing neurons within the ventral horn in naïve animals, specifically around phrenic motoneurons (PhMNs), but not phrenic spinal interneurons. Right, but not left, C2Hx resulted in a significant increase in PNNs and glutamatergic synapses around ipsilateral PhMNs, suggesting that the right C2Hx requires greater neuroplasticity to overcome respiratory dysfunction. The results from this study uncover profound anatomical and functional asymmetries in left- and right-sided phrenic networks, underlying the complex nature of the spinal respiratory system, and contribute to a more advanced understanding of how the phrenic network adapts to trauma. Overall, this work underscores the importance of studying neuroplasticity and how it holds the potential to help improve outcomes for individuals living with SCI.
Introduction
Neuroplasticity is defined as persistent anatomical and functional changes in the nervous system in response to stimuli or injury. In the spinal cord, plasticity has been mainly studied in the locomotor system to promote recovery after a spinal cord injury (SCI) (reviewed in the study by Loy and Bareyre 1 ). However, other spinal circuits such as the phrenic motor system have also shown spontaneous recovery after injury.2,3 Phrenic motoneurons (PhMNs) are located at spinal cervical levels 3 − 6 (C3−C6) and innervate the diaphragm, the primary muscle of inspiration. 4 High- and mid-cervical SCIs denervate PhMNs from upper brainstem centers and generate dysfunctional respiration, a primary contributor to morbidity and mortality in SCI patients. 5 Despite this devastating outcome, clinical and experimental research has shown some spontaneous plasticity that promotes limited functional recovery after cervical SCI. 6
Lateral hemisection at the C2 level (C2Hx) has become the most widely used pre-clinical SCI model to study respiratory plasticity, since it was first described by Porter in 1895. 7 A C2Hx interrupts descending bulbospinal pathways on one side of the spinal cord and immediately silences the activity of the ipsilateral hemidiaphragm. However, within minutes to hours after the injury, anatomical plasticity occurs that can lead to spontaneous functional recovery weeks or months later.6,8,9 Spinal interneurons have been identified as a key player in this plasticity following a left C2Hx, able to anatomically reorganize their connectivity within the injured (left) network and modulate PhMN activity.10–13 However, the neuroplastic potential of these spinal interneurons and PhMNs after right C2Hx injury is unknown.
Perineuronal nets (PNN) are structures rich in chondroitin sulfate proteoglycans that ensheathe neurons, mediating neuronal axonal growth and synapse formation in an activity-dependent manner (reviewed in the study by Fawcett et al. 14 ). Changes in neural activity can alter the expression of PNN components, and consequently promote stability or plasticity in the central nervous system (CNS). Reductions in PNNs open a window of plasticity that favors reorganization of circuits, whereas increases in PNNs suppress neuroplastic potential, attenuate new synapse formation, and stabilize the existing synaptic contacts. 15 Thus, levels of PNNs surrounding neurons in the CNS can reflect relative stability or plasticity within specific neural circuits. We previously showed that 5 weeks after a thoracic SCI, PNNs decreased around lumbar motoneurons, which favored the formation of new synaptic contacts. After 11 weeks, PNN density recovered normal values, closing the plasticity period, and stabilizing the newly formed synapses. 16 Although locomotor and respiratory circuits share some similarities, their window of plasticity might differ, given the increased drive for sustained activity in breathing networks. The neural networks controlling breathing must be more flexible to any physiological or environmental challenge, due to their biological significance. Hence, this study aimed to analyze changes in PNN density around phrenic neurons after a C2Hx. We used a retrograde, trans-synaptic tracer, pseudorabies virus (PRV), to label either the left or right phrenic motor circuit and analyzed changes in PNN density at 1 or 2 weeks post left- or right-sided C2Hx. Our results reveal changes unique to each side of the spinal cord, offering important insights into differing neuroplastic potential between the two sides that have not been previously explored. Defining the PNN dynamics in the phrenic network of the intact and injured spinal cord is crucial in elucidating the contribution of these neural populations in the spontaneous plasticity that mediates respiratory recovery.
Materials and Methods
Experimental design
Adult female C56BL6/J mice (n = 51; 2–3 months) were used for the present study. Mice were housed at the animal care facility at Drexel University College of Medicine (Philadelphia, USA) and in the animal care facility of the Universitat Autònoma de Barcelona (UAB, Spain). Mice were kept on a 12:12 light/dark cycle and received food and water ab libitum in any case.
Sham mice (n = 8) were used to characterize the left and right phrenic motor circuits. Specifically, we compared PNN density, synaptic proteins, and the number of motoneurons on each side of the spinal cord. To assess PNN density in the phrenic spinal circuit after a SCI, mice were separated into a sham (n = 7) and injured group (n = 36). The latter received a spinal cord hemisection at the second cervical spinal level (C2Hx) either on the left (n = 30) or right (n = 6) side of the cord. Animals that received left C2Hx survived for 7 (n = 16) or 14 days postinjury (dpi) (n = 14) prior to neuroanatomical tracing and perfusion. Animals that received right C2Hx survived for 7 dpi (n = 6) prior to tracing and perfusion (Fig. 1), since these mice showed an unusual high rate of attrition within the first days after the C2Hx. Histological analysis in SCI animals was performed on the ipsilateral side below the injury, therefore, the right side was explored after right C2Hx and the left one after left C2Hx.

Schematic representation of the experimental groups and design.
Experimental procedures were conducted with approval from the Institutional Animal Care and Use Committee of Drexel University and the UAB Experimental Ethical Committee. These procedures follow the National Research Council Guidelines for the Care and Use of Laboratory Animals (USA) and the animal welfare guidelines of the European Communities Council Directive (2010/63/EC).
Spinal cord injury
Cervical spinal cord hemisection at C2 level was performed under isoflurane anesthesia (induction 4%, maintenance 1–2%) and on a water-circulating heating pad. The skin of the dorsal part of the neck was shaved, sterilized with ethanol and betadine, and an incision was made from the base of the skull to the shoulder blades. Surrounding musculature was retracted to reveal the vertebral column prior to a partial laminectomy at the C2 vertebral level. Using a No. 11 scalpel blade, a C2Hx was induced immediately caudal to C2 roots, from the middle to the lateral edge of the spinal cord. To ensure a complete and reproducible hemisection, the scalpel was drawn from the midline to the lateral edge of the spinal cord three consecutive times. Surrounding muscles were sutured, and skin was closed using wound clips. Postoperatively, lactated ringers’ solution (3 mL) and buprenorphine (0.03 mg/mL; 0.05 mL) were provided subcutaneously (s.q.).
Neuronal tracing
To study PhMNs that innervate the left and right hemidiaphragm, mice were retrogradely traced with True-Blue Chloride (1%; TB, Setareh Biotech). Mice were anesthetized with ketamine (90 mg/kg) and xylazine (10 mg/kg) prior to aseptic preparation of the abdomen. A laparotomy was performed to expose the diaphragm, and TB was applied topically (9 µL) on the left or right hemidiaphragm. The abdominal muscles and skin were sutured, and analgesia was provided s.q. with buprenorphine (0.1 mg/kg). Animals were euthanized six days after tracing.
To trace the entire left or right phrenic motor circuit, animals were anesthetized with isoflurane (induction 4%, maintenance 0.5–2%) prior to aseptic preparation of the abdomen. A laparotomy was performed to expose the diaphragm, and a trans-synaptic retrograde PRV, 40 µL) 17 was topically applied to either the left or right hemidiaphragm, as described previously in mouse 18 and rat. 19 Either PRV614 (8 × 108 plaque-forming units [pfu]/mL; expressing red fluorescent protein), PRV152 (5 × 109 pfu/mL; expressing green fluorescence protein), or PRV Bartha (1 × 109 pfu/mL; without a fluorescence protein) was used for these tracing studies. In animals with cervical hemisection, tracer was applied to the ipsilateral side of the hemidiaphragm. In sham animals, tracer was applied to the left. The abdominal muscles were sutured, the skin closed, and lactated ringers’ solution (1 mL s.q.) and buprenorphine (0.03 mg/mL; 0.05 mL s.q.) were provided. Animals were euthanized 72 h after the neuronal tracing. 19
Histology
All mice were intracardially perfuse-fixed with 4% paraformaldehyde (w/v in 0.1 M phosphate-buffered saline (PBS), and the cervical spinal cords were harvested and postfixed in 4% paraformaldehyde over 2 h. Long-term tissue storage was in 2% paraformaldehyde at 4°C. Tissues were cryoprotected in 15–30% sucrose solution in PBS (4°C), sectioned longitudinally on a cryostat (20 µm thick) and collected onto serial gelatin-coated glass slides.
A series of sections 40 µm apart from animals that received TB tracing were used to quantify the number of TB-positive motoneurons on each side of the spinal cord. Images were taken with a 10x upright microscope.
To assess the distribution and density of PNNs and synaptic changes, a series of sections 60 µm apart, were immunohistochemically labeled with antibodies against aggrecan, serotonin (5-HT), vesicular glutamate 2 (VGlut2), and PRV (see Table 1), as previously described.19,20 Secondary antibodies were conjugated with fluorophores as described in Table 1. Immunolabeled sections were washed and cover-slipped with fluorescence mounting medium (Fluoromount medium; Southern Biotech). Images were captured with a confocal laser-scanning microscope (around 30 steps, z-step size of 0.5 µm, Leica TCS SP5). For each animal, all PRV-positive neurons were imaged, maximal projections of the z-stacks were created, and the background was corrected. To quantify aggrecan and VGlut2 labeling, a 4 µm band around the PRV-stained cell body was delineated to measure the integrated density of that region. For 5-HT analysis, the immunolabeling intensity of the whole picture was quantified. Then, the data obtained were clustered according to dorso-ventral spinal cord regions (dorsal horn, intermediate region, and ventral horn).
List of Primary and Secondary Antibodies

PRV, pseudorabies virus; VGlut2, vesicular glutamate transporter 2.
Statistical analysis
Data were analyzed using GraphPad Prism 7 software and reported as mean ± standard error of the mean (SEM). Differences between groups were considered significant when p value <0.05. Normal distribution was confirmed using Shapiro–Wilk test for each variable/group. The histological analysis performed for the phrenic motor pool characterization was analyzed by Student’s t-test (unpaired), since variables presented equal variances. Differences in the expression of aggrecan before the injury in the different spinal cord regions were assessed by one-way analysis of variance (ANOVA) followed by Bonferroni’s multiple comparison test. The changes in PNNs and synaptic markers (VGlut2 and 5-HT) at 7 and 14 dpi were evaluated with one-way ANOVA with same post hoc as above.
Results
Left and right phrenic motoneurons do not differ in PNN density nor synaptic content
To evaluate how spinal PNNs are modulated after injury, we first studied PNN density and synaptic content around TB-labeled PhMNs in the naïve (uninjured) spinal cord (Fig. 2A, B). Integrated density of the synaptic markers 5HT and VGlut2 around PhMNs revealed no significant differences in serotoninergic and glutamatergic inputs between the left and right PhMN pools. Similarly, there were no significant differences in the density of aggrecan-positive PNNs between left and right PhMN pools.
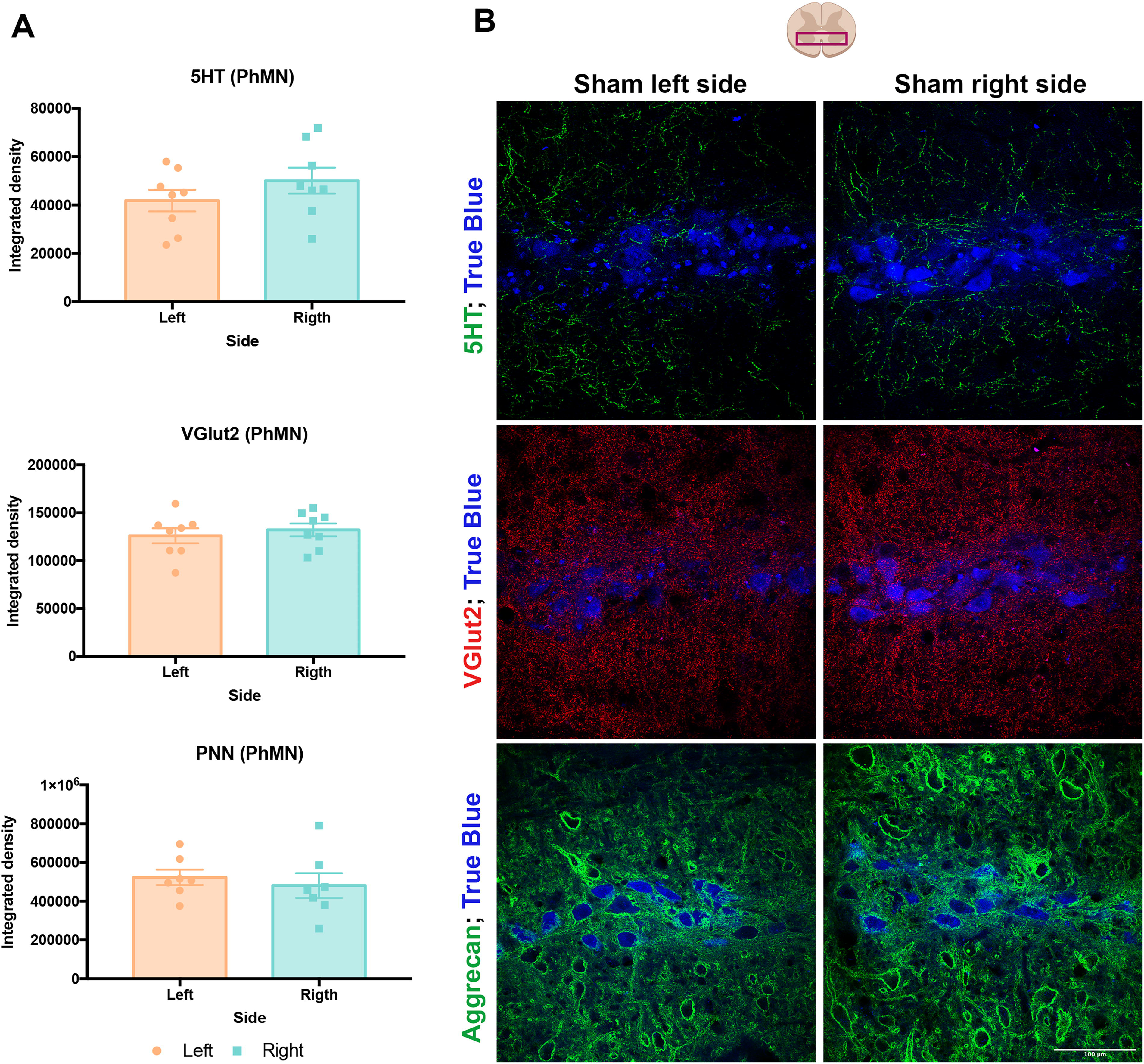
The left and right phrenic motor pools do not present significant differences in PNNs, VGlut2, or 5-HT levels.
PNN density in the spinal phrenic circuit increases from dorsal to ventral spinal regions
Phrenic interneurons were identified with PRV-positive labeling and were located throughout the dorso-ventral neural axis. We immunohistochemically measured the density of aggrecan-positive PNNs around phrenic neurons and interneurons in intact mice in the dorsal, intermediate, and ventral regions of the spinal cord (Fig. 3A, B). We found the least number of PNN-bearing neurons within the dorsal horn, and therefore normalized the data to the dorsal horn. This analysis revealed higher density of PNNs around PRV-positive phrenic neurons within intermediate gray region, albeit not reaching significance. In contrast, PhMNs located within ventral region of the spinal cord showed a statistically significant higher density of PNNs than those in the dorsal horn (p < 0.01).
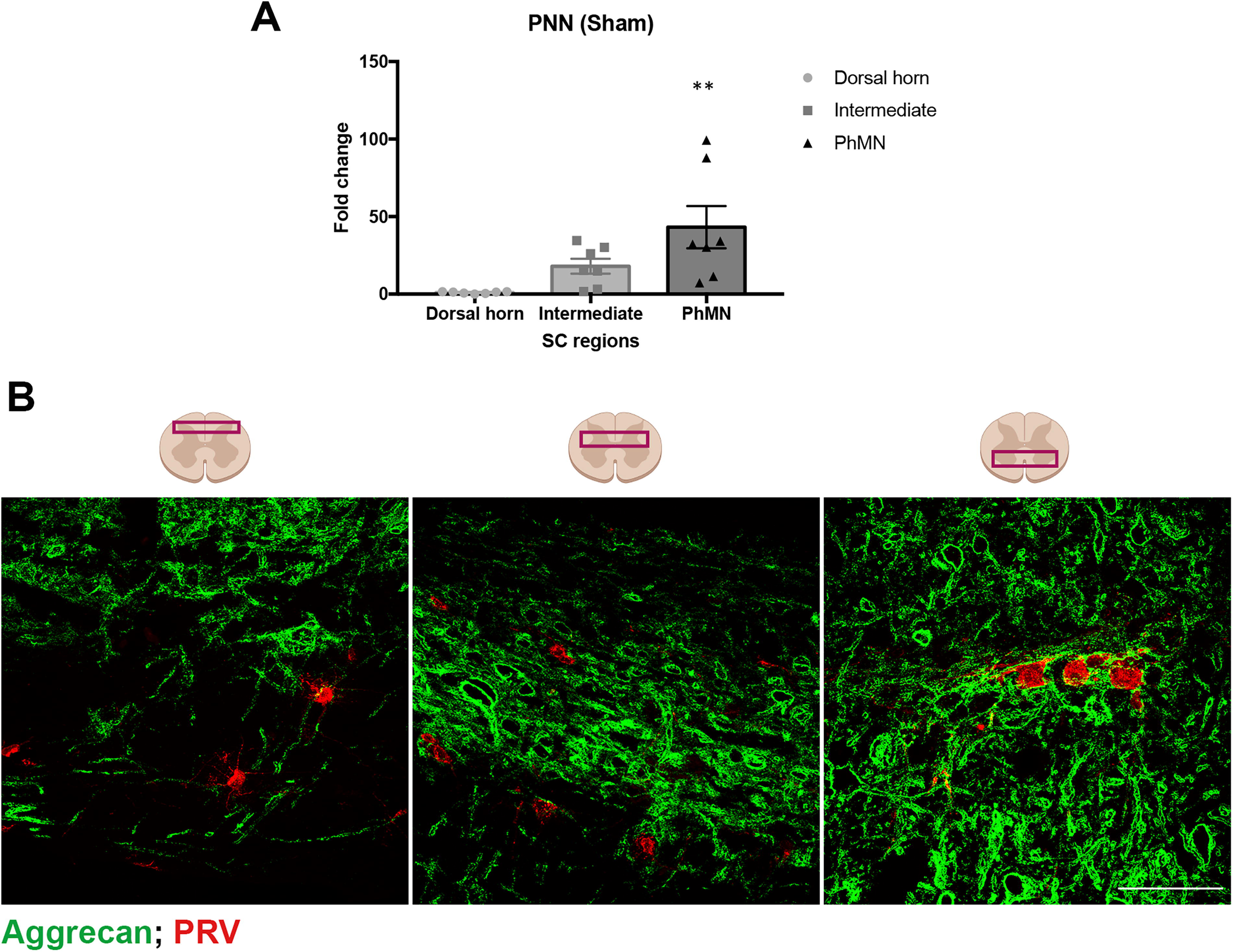
PNNs are mainly found around PhMNs, whereas very few spinal interneurons are coated by them.
Regulation of PNNs differs depending on the side of SCI
We observed an unusual high rate of mortality in animals receiving a right C2Hx. Three animals went into respiratory arrest after the injury, and four animals did not survive 7 days postinjury. Therefore, we just followed the right C2Hx animals for 1 week. Thus, we evaluated the differences in plasticity of PNNs and synaptic content between left and right phrenic circuits 7 days after a left or right C2Hx. We found that a C2Hx on the right side of the spinal cord resulted in higher PNN density around ipsilateral neurons in the dorsal horn compared to the sham group (p < 0.05; Fig. 4A, C). In contrast, after a left-sided C2Hx, ipsilateral PNNs were not significantly increased. Similarly, ipsilateral PhMNs after a C2Hx on the right side showed a significant increase in PNN density compared to the sham and left-side injured groups (p < 0.001 vs. sham and 7 dpi left). We did not observe significant changes in PNNs surrounding phrenic neurons in the intermediate region of the spinal cord.
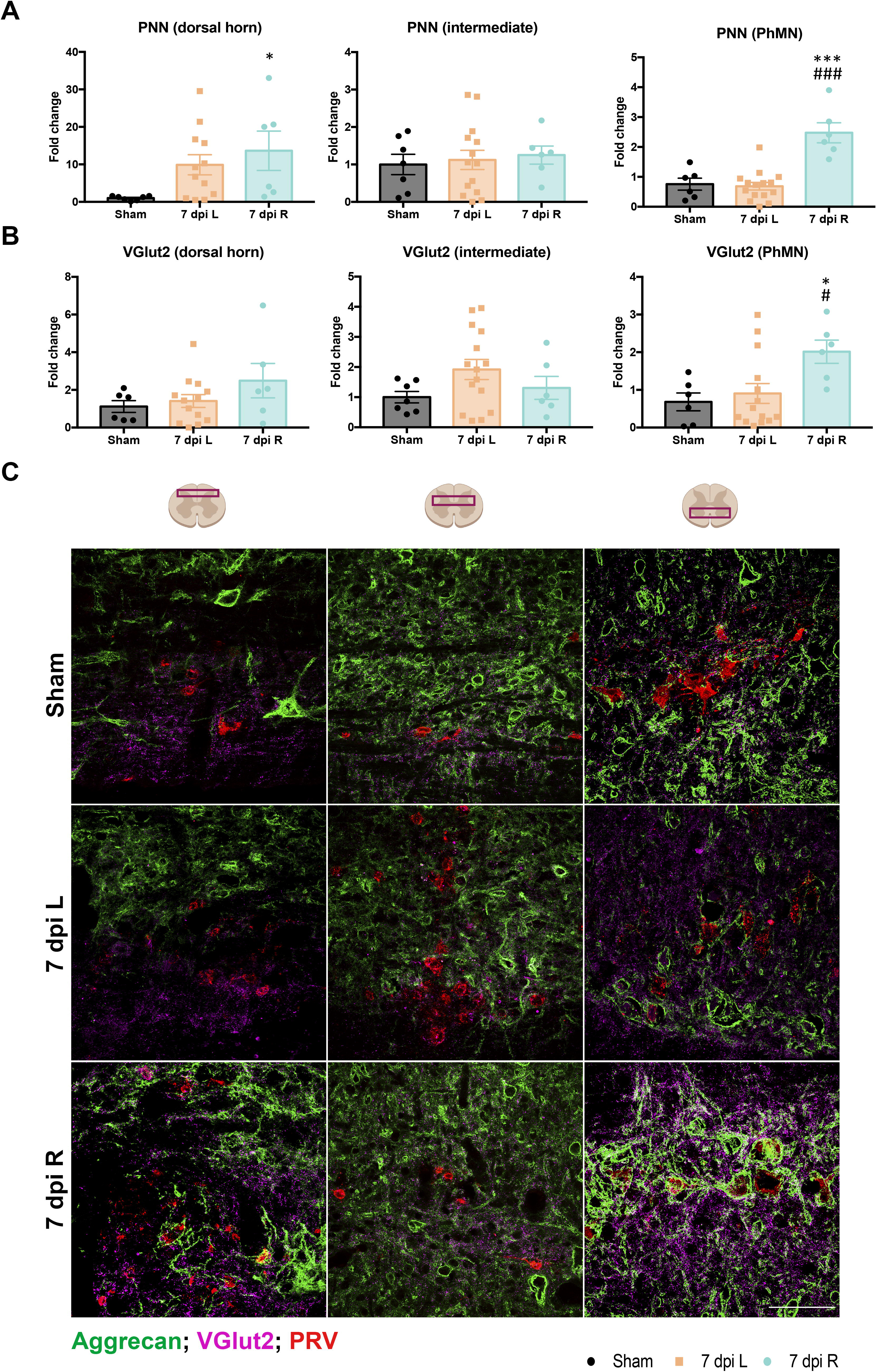
PNNs and VGlut2 are differently regulated depending on the side of the C2Hx. Quantification of the intensity of PNNs
To investigate synaptic plasticity between left and right phrenic circuits, we used a marker of glutamatergic synapses (VGlut2) to label excitatory synapses in sham and injured animals (Fig. 4B, C). We observed a significant increase in VGlut2 intensity around ipsilateral PhMNs after a C2Hx on the right side, compared to the sham and left-side C2Hx injured groups (p < 0.05 vs. sham and 7 dpi left). We did not observe significant changes in VGlut2 staining around phrenic neurons 7 days after a left C2Hx.
To evaluate differences in bulbospinal serotonergic innervation on the phrenic network along the dorso-ventral neural axis, we measured 5-HT intensity around phrenic neurons in the dorsal, intermediate gray, and ventral regions of the spinal cord. Neither a left-sided nor right-sided C2Hx resulted in significant 5-HT changes in any of the spinal cord regions (Fig. 5A, B). When analyzing the evolution of synaptic contents from 7 to 14 dpi in left C2Hx animals, we detected a significant decrease of serotoninergic inputs in the ipsilateral ventral horn after injury, compared to sham (Supplementary Fig. S1).
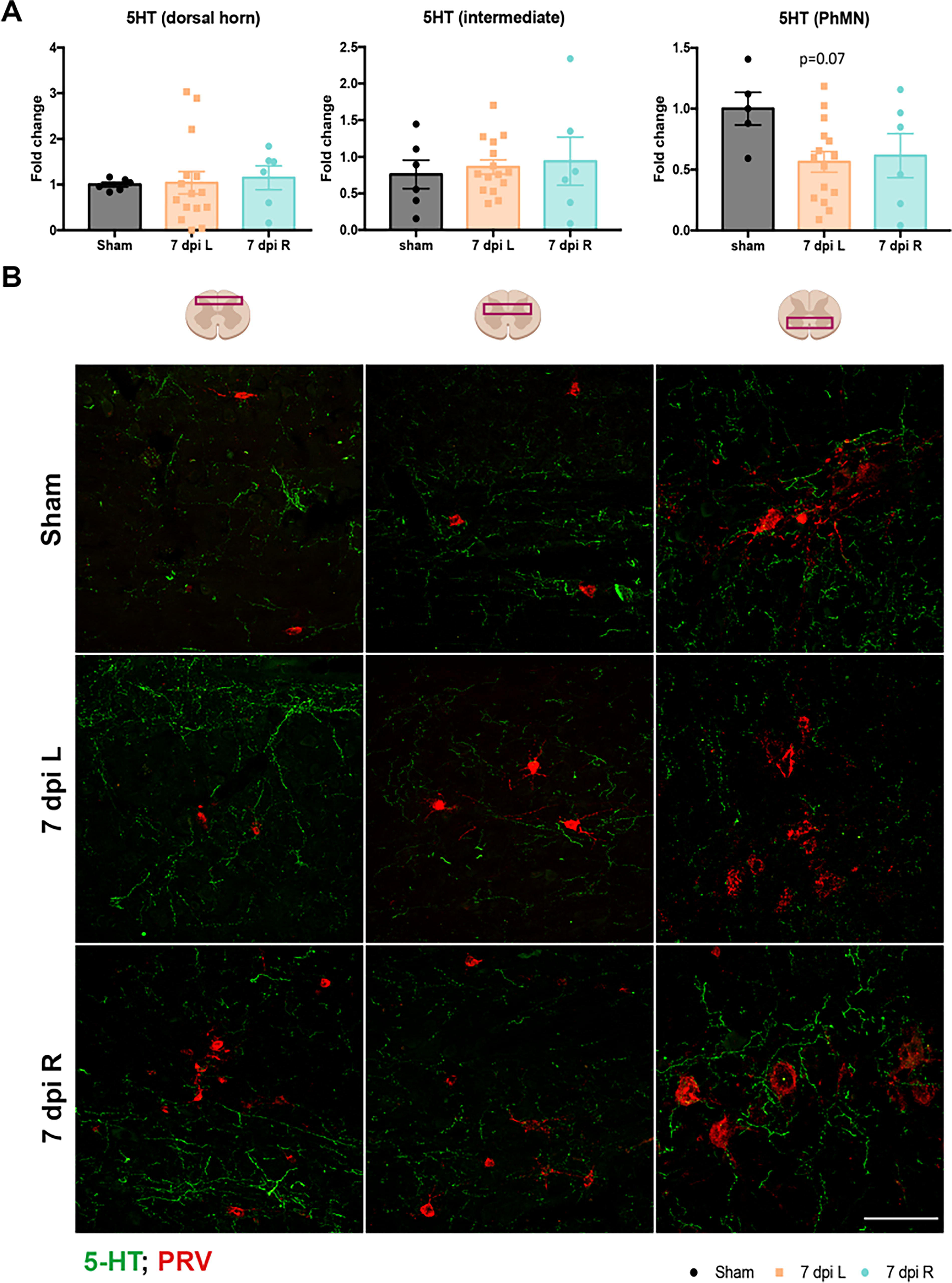
No significant changes in 5-HT at 7 dpi following either a left or right C2Hx.
The right phrenic motor pool presented more phrenic motoneurons
To explain the differences in PNN and excitatory synaptic contacts between sides after injury, PhMNs were counted in the left and right phrenic circuits of naïve mice (Fig. 6A, B). Interestingly, the right phrenic motor pool showed 23% more MNs than on the left side (p < 0.01).

The right phrenic network has more PhMNs.
Discussion
Since structure and function are closely tied, understanding structural plasticity within the phrenic circuit and its temporal change after SCI is crucial to predicting its contribution to functional recovery postinjury. In this study, structural plasticity was assessed by analyzing PNN changes around phrenic interneurons and motoneurons following C2Hx injury. While this and the majority of other studies have used a left-sided C2Hx injury to observe anatomical and structural changes within the phrenic network,20–25 here, we have additionally described the neuroplastic changes that occur following injury to the right phrenic circuit. These results offer valuable insight into how each side of the phrenic motor network may respond differently to injury and denervation.
PNN density confirms the plastic potential of the phrenic motor system
To get a better understanding of PNN dynamics, we first sought to understand the basal state of PNNs around the phrenic neural population. Aggrecan was used as a marker of PNN since it has been shown that in the spinal cord, it labels more than 90% of PNN around alpha MN, being a better label for spinal PNN than Wisteria floribunda agglutinin. 26 In normal conditions, aggrecan immunolabeling revealed that PNNs are rarely present around neurons in the dorsal horn. Similarly, interneurons located in the intermediate region showed limited PNNs. In contrast, PhMNs have thicker PNNs, but without significant differences between sides. This supports the notion that interneurons could have greater plasticity and thus can be more readily recruited under specific physiological and environmental demands.10,13,20,27 As PhMNs provide stable and synchronized motor output to activate the diaphragm, they likely require more stable synaptic input. 28 Worthy of consideration, however, in comparison to other spinal motor circuits like the lumbar circuit, the phrenic network can be considered extremely malleable. In the lumbar spinal cord, we previously observed more PNN-bearing neurons in both the ventral horn and intermediate gray region than within the cervical spinal phrenic network. 29 This notion highlights the biological significance of the potential synaptic flexibility of the phrenic circuit in comparison to other spinal circuits.
Structural plasticity differs on each side of SCI
Injury to the spinal cord rearranges circuits into new anatomy. In our work, we observed a difference in neuroplastic changes that were predominantly dependent on the side of the spinal cord that was injured. Interestingly, while the left C2Hx did not modify PNNs, the same injury on the right side significantly increased PNN density around interneurons in the dorsal horn and PhMNs. Furthermore, glutamatergic inputs also significantly increase around right ipsilateral PhMNs when compared to sham and the left-injured group. The observed increase of PNNs around denervated cervical neurons was unexpected, as our previous reports show a reduction around denervated lumbar neurons following thoracic SCI. 16 The loss of synaptic contacts reduces PNNs, whereas the return of these inputs increases PNNs, which strengthens the existing synaptic connections while limiting the formation of new ones. 14 Given this background, we expected a reduction in PNNs around denervated PhMNs due to the decline of inputs after hemisection. However, activity-dependent plasticity does not follow the same rules in all neural systems, and our results suggest that in constitutively active systems such as the respiratory network, plasticity is regulated by other activity-independent mechanisms that guard the system against inactivity (since the consequence of respiratory failure is death). This compensatory mechanism, inactivity-induced phrenic motor facilitation, consists of a burst of inputs around PhMNs after a period of activity deprivation.30,31 The C2Hx injury compromises all direct ipsilateral input from respiratory neurons in the medulla that innervate PhMNs, and a loss of these inputs results in ipsilateral hemidiaphragm paralysis acutely after injury. However, this immediate paralysis initiates a compensatory mechanism that increases glutamatergic synapses from spared pathways (contralateral bulbospinal axons and spinal interneurons that innervate the PhMNs). Predictably, this sprouting consequently increases PNNs around the PhMN, but surprisingly, only after injury to the right side of the cervical spinal cord. Since the boost of synaptic contacts is proportional to the magnitude of inactivity, 30 it is possible that denervation of the right and left phrenic spinal network has distinct impacts on PhMN activity. Notably, diaphragm innervation is also not symmetric. In fact, prior studies have shown that the right phrenic nerve contains 20% more axons than the left nerve, innervating a larger area of the diaphragm. 32 This is consistent with the neural tracing and MN labeling in the present work, in which we observed 23% more PhMNs on the right side compared to the left side in naïve mice (Fig. 6). Consequently, our hemisection did not disconnect from upper centers 50% of the phrenic neurons in each case, but rather 30% or 70% when we injured the left or right side, respectively. The differential denervated states can be extrapolated into PNN changes. Since the left-side injury might have a smaller impact on diaphragm control, little spinal reorganization was needed, and thus, PNNs remained unchanged. However, after the right-side injury, diaphragm activity could be compromised, activating the iPFM and increasing PNNs around disconnected PhMNs. This histological finding was corroborated at the functional level, since the right-side injury had an extremely high mortality rate within the first days after the injury, compared to the left-side injury.
While the neuronal circuits mediating this spontaneous respiratory recovery after C2Hx are challenging to identify, concerted efforts over the past decade have anatomically and electrophysiologically identified two key substrates that appear to be involved: spared bulbospinal axons from the contralateral spinal cord and cervical spinal interneurons.10,13 The activation of latent, crossed, and monosynaptic projections from the ventral respiratory column, although possibly the most effective route to enhance the excitatory drive to PhMN, 33 hardly contributes to the spontaneous recovery found after C2Hx. 11 Instead, retrograde and trans-synaptic tracing experiments have revealed an increased number of labeled interneurons connecting the ipsilateral phrenic network after injury.10,13,19,34 This observation indicates a key role of spinal interneurons in spontaneous respiratory recovery after cervical SCI. Among spinal interneurons’ diversity, excitatory interneurons seem to be recruited within the first weeks after the injury and participate in respiratory recovery. 26 Specifically, glutamatergic V2a interneurons are a promising candidate for facilitating breathing after SCI. 20 While V2a spinal neurons are integrated with the phrenic network in naïve mice, their connectivity to the spinal phrenic network significantly increased after C2Hx. Notably, these prior studies showed this change following a left C2Hx, while VGlut2 immunofluorescence did not significantly change after a left C2Hx in the present work. Although the overall glutamatergic input measured by VGlut2 quantification did not significantly change, one consideration is that the increase in interneuronal glutamatergic input may have compensated for the loss of bulbospinal glutamatergic input, resulting in no net change. In contrast, we measured significantly greater glutamatergic contacts around PhMNs after a right C2Hx. The origin of this input, however, remains unknown. One possibility is that there is even greater activation of these inputs when the right spinal phrenic system is denervated by a lesion rostral to the motor pool, resulting in greater glutamatergic sprouting and subsequently greater expression of PNNs surrounding ipsilateral PhMNs.
Another key anatomical component of plasticity in the injured spinal cord, especially within the respiratory networks, is the serotonergic projections that modulate spinal neuronal network function and recovery. While several prior studies have shown that serotonergic input contributes to ipsilateral phrenic motor plasticity after a C2Hx,35,36 here we show that the extent of serotonergic input to the spinal networks caudal to a left C2Hx remains below what is seen in naïve (uninjured) animals. In fact, ipsilateral serotonergic input is significantly reduced 2 weeks following a left-sided injury. In contrast, we observed that 5-HT fluorescence was unchanged at 7 dpi after a right C2Hx, which could reflect a contribution to plasticity on that side. In addition, V2a interneurons receive serotoninergic (5-HT) inputs. 20 It has been reported that 5-HT plays a neuromodulatory role, contributing to respiratory recovery by enhancing neuroplasticity in the phrenic system.8,37
A limitation in the present study is that we could not analyze histological changes in mice injured on the right side at 14 dpi, mainly due to the high mortality rate after injuring the right phrenic circuit. This is an important result, suggesting that right-sided injuries impact phrenic motor function to a greater extent. Future studies could aim to assess this more acutely following SCI. In the left C2Hx group, since few anatomical changes were observed at 7 dpi, it is not surprising that at 14 dpi, the same markers remained unchanged. Functional plasticity is first observed in the second-week postinjury.20,23 Thus, while structural and anatomical plasticity likely starts within this acute period, there is also evidence for ongoing changes at more subacute and chronic times. 13 Accordingly, a more in-depth assessment of these chronic changes will be important to identify possible targets for treating the chronically injured spinal cord.
Overall, our data demonstrate an asymmetrical distribution of PhMNs between spinal sides, and a differential PNN modulation after injuring the right or left spinal cord at the cervical level. These anatomical findings suggest a predominant phrenic circuitry in the right side, which is expected to undergo a stronger plastic reorganization after injury.
Conclusions
This study demonstrates differences between the right and left phrenic circuitry that coincide with the neuroplastic outcomes after SCI. These data provide a new perspective on how we should consider phrenic motor function in naïve and injured conditions, with important implications for how the laterality of injury could result in significantly different functional outcomes. These differences may also have important considerations for developing and applying treatments that target repair and activity within these distinct networks. These results highlight the need for further research into the functional implications of affecting both phrenic circuits. A better understanding of phrenic circuitry will be essential to harness the spontaneous and limited recovery observed in this system and potentiating it through therapeutic interventions.
Transparency, Rigor, and Reproducibility Summary
The characterization of PNN in the phrenic circuit and the impact of left or right cervical hemisection on PNN was designed prior to the initiation of the experiments. However, due to an unexpectedly high mortality of animals injured on the right side of the cord, those with right hemisection were terminated at a single time-point: 7 days. The planned simple size was determined based on data from similar studies. However, as mentioned above, the higher mortality when injuring the right spinal cord reduced the “n” in that group. Finally, to further evaluate potential differences between the left and right phrenic circuitry, estimation of the number of PhMNs on the right and left sides was performed. Statistical analysis was performed using GraphPad Prism 7 software, and data were considered significant when p value <0.05. Normal distribution was confirmed using Shapiro–Wilk test. One-way ANOVA followed by Bonferroni’s multiple comparison test was used to compare the expression of aggrecan in the different spinal cord regions and to evaluate the changes in the spinal phrenic circuit after injury, whereas an unpaired Student’s t-test was used in the characterization of the phrenic motor pool. All the histological analysis was performed blinded to the experimental group. Data are available from the corresponding author upon request.
Authors’ Contributions
J.S.V., M.A.L., and E.U. contributed to conceptualization, formal analysis, investigation, methodology, validation and writing the original draft, and revised the final version. M.A.L. and E.U. also contributed to supervision, project administration, resources, and funding acquisition. K.A.S., T.F., and L.V.Z. contributed to conceptualization, investigation, and revised the article.
Footnotes
Acknowledgments
Finally, the authors would like to acknowledge that most of the studies described in the article were part of the PhD thesis of the first author, J.S.-V., under the supervision of senior author E.U., entitled “The role and modulation of PNNs in the healthy and injured spinal cord,” defended in 2023 in Universitat Autònoma de Barcelona. The authors thank members of the Udina and Lane laboratories for discussion and feedback on these studies. The authors also thank the Drexel University Laboratory Animal Resources facility at Queen Lane College of Medicine for support with animal housing and care. The authors thank Kathryn Claiborn of the Gladstone editorial team for her helpful feedback and article revisions.
Author Disclosure Statement
None of the authors has any competing or conflicting interests.
Funding Information
This work was funded by the Fundació La Marató-TV3 (TV3-201736-30-31). The author’s research was supported by funds from CIBERNED and TERCEL networks, cofunded by European Union (ERDF/ESF, “Investing in your future”). J.S.V. holds a predoctoral fellowship of the AGAUR, Secretaria d’Universitats i Recerca del Departament d’Empresa i Coneixement de la Generalitat de Catalunya, cofunded by European Social Funds. T.F. was supported by NIH (NINDS, F31 NS125975). M.A.L. was supported by the NIH (NINDS, R01 NS104291) and the Pennsylvania State Fund for Spinal Cord Injury Research. L.V.Z. was supported by the Lisa Dean Moseley Foundation, NIH (NINDS F32 NS119348), and CIRM DISC2-14180.