
Abstract
Influenza viruses A and B cause widespread infections of the human respiratory tract; however, existing vaccines and drug therapy are of limited value for their treatment. Here, we show that bispecific short-hairpin small-interfering RNA constructs containing an 8-nucleotide intervening spacer, targeted against influenza virus A or influenza virus B, can inhibit the production of both types of virus in infected cell lines. This multiple vector showed remarkable ability to cope with both influenza viruses A and B. Furthermore, the Autographa californica multiple nuclear polyhedrosis virus can infect a range of mammalian cells, facilitating its use as a baculovirus vector for gene delivery into cells. In this study, baculovirus-mediated bispecific short-hairpin RNA expression markedly inhibited both influenza viruses A and B production.
Introduction
I
Small-interfering RNAs (siRNAs) that are 21–25 base pairs (bp) in length with 3′ overhangs of 2 nucleotides can cause RNA interference (RNAi; Elbashir et al., 2001). siRNAs are a powerful tool for sequence-specific post-transcriptional gene silencing, and have potential therapeutic and prophylactic applications against cancer, as well as infectious diseases (Haasnoot and Berkhout, 2006). siRNAs have also been used recently to interfere with the replication of an influenza virus (Ge et al., 2003, 2004; Tompkins et al., 2004). Baculoviruses have previously been employed to deliver genes into mammalian cells (Matsuura et al., 1987). The Autographa californica nuclear polyhedrosis virus (AcNPV) has long been used as a biopesticide and as a tool for efficient recombinant protein production in insect cells (Matsuura et al., 1987; Luckow and Summers, 1988). The host specificity of this baculovirus was originally thought to be restricted to cells derived from arthropods; however, with an appropriate eukaryotic promoter, it can express foreign genes in several types of mammalian cell (Boyce and Bucher, 1996; Barsoum et al., 1997; Shoji et al., 1997; Condreay et al., 1999; Sarkis et al., 2000). The baculovirus may also be applied in conjunction with other viral vector to escape either pre-existing or therapy-induced anti-viral immunity.
Here, we report on the identification of baculovirus vector-mediated bispecific short-hairpin siRNA expression vectors (Sioud and Leirdal, 2002; Anderson et al., 2003) targeting the NP gene sequence of influenza virus A (Ge et al., 2003) and influenza virus B. This multiple vector showed remarkable ability to cope with either influenza virus A or B. The baculovirus vector is unable to replicate and do not have a cytopathic effect (CPE) in mammalian cells, even at a high multiplicity of infection (moi), this vector is now recognized as a useful viral vector, not only for the expression of foreign proteins in insect cells but also for gene delivery to mammalian cells.
Materials and Methods
Cell lines
Madin-Darby canine kidney (MDCK) cells were maintained in Dulbecco’s modified Eagle’s medium (DMEM; low glucose, Sigma Chemical Co., St. Louis, MO) supplemented with 10% heat-inactivated fetal bovine serum (FBS) and 1% penicillin-streptomycin.
Influenza virus strains
Influenza virus A/Puerto Rico/8/34 (A/PR/8/34) and influenza virus B/Ibaraki/2/85 (B/Ibaraki) were propagated following previously published procedures (Abe et al., 2003).
Plasmid construction
The pSV2neo-U6 plasmid vectors and pVL1393-based baculovirus vectors (Invitrogen, Carlsbad, CA) were used in this study. The NP genes of influenza viruses A and B were used as targets, and the shRNA expression plasmid vectors were constructed under the control of the human U6 Pol III promoter. We searched for siRNAs targeting the sequences in the influenza B virus NP gene. The selected sequences were used in a BLAST search against the human genome sequence to ensure that the latter was not targeted. To construct the shRNA expression cassette, the following DNA oligonucleotides were synthesized (here the lowercase letters at the 5′ end represent the template plasmid pSVneo-hU6 KpnI site sequence, the uppercase letters represent the NP sequence, which includes the sense and antisense sequences of a particular NP gene region, the lowercase letters indicate the linker (9 nt), the stretch of 5 thymidines at the 3′ end (Pol III terminator) is underlined, and the lowercase letters at the 3′ end represent the template plasmid pSVneo-hU6 BamHI site sequence BNP-52, 5′-cCCAGAAGAAACAAC TTCTGttcaagagaCAGAAGTTGTTTCTTCTGG
Design of bispecific short-hairpin siRNA constructs
The bispecific short-hairpin siRNA constructs contained 2 shRNAs, separated by a spacer sequence. We experimented with 2 different spacer sequences: UUCAACUU (Anderson et al., 2003) and UUCAAGAGA (Anderson et al., 2003). The shRNA constructs were targeted against sequences in the influenza A virus NP gene (ANP-1496) and the influenza B virus NP gene (BNP-52). To construct the bispecific short-hairpin siRNA expression cassette, the following DNA oligonucleotides were synthesized: AB1sh, 5′-cGGATCTTATTTCTTCGGAGttcaacttCTCCGA AGAAATAAGATCCTTCAACTTCCAGAAGAAACAAC TTCTGttcaacttCAGAAGTTGTTTCTTCTGG
Inhibition of influenza virus replication by shRNA-expressing plasmid
MDCK cells were plated at 2.5 × 105 cells/well in a 6-well plate and cultured for 24 hours. Each shRNA expressing plasmid (1 µg) was transfected with the FuGENE6 transfection reagent (Roche Diagnostics, Basel, Switzerland). After 8 hours, the MDCK cells were washed with phosphate-buffered saline (PBS) and infected with A/PR/8/34 or B/Ibaraki at a multiplicity of infection (moi) of 0.01 at 37°C for 1 hours. After infection, the cells were washed with PBS and cultured in MEM containing 0.5% heat-inactivated FBS and 4 µg/mL trypsin (Invitrogen) for 3 days. The culture supernatants were harvested, and the virus titers were determined by a plaque assay.
Preparation of baculoviruses
The baculovirus transfer vector, pVL1392-shRNAs, was constructed by inserting the shRNA expression cassettes into the cloning site of the baculovirus transfer vector pVL1392. The recombinant baculovirus containing shRNA of the NP genes of the influenza viruses A and B (rBV-shRNAs) was generated by homologous recombination of the transfer vector and linearized baculovirus DNAs (BD Biosciences, San Jose, CA) following previously published procedures (Abe et al., 2003).
Inhibition of influenza virus replication by baculovirus-mediated bispecific short-hairpin siRNA
MDCK cells were plated at 2.5 × 105 cells/well in a 6-well plate and cultured for 24 h. The MDCK cells were washed with PBS and infected with A/PR/8/34 or B/Ibaraki (moi = 0.01) at 37°C for 60 minutes. After 6-h incubation, MDCK cells were washed with PBS and infected with shRNA-expressing baculovirus (moi = 50 or 100) at 37°C for 60 minutes. After infection, the cells were washed with PBS and cultured in DMEM containing 0.5% BSA and 4 µg/mL trypsin (Invitrogen) for 4 days. The supernatants were harvested and the titers of the viruses were measured by a plaque assay.
Western blot analysis
The cells were washed with PBS and lysed on ice with cell lysis buffer (containing 0.2% NP-40 in PBS and protease inhibitors). Protein concentrations were determined using the BCA protein assay kit (Pierce, Rockford, IL). Equivalent amounts of cell lysate were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene fluoride (PVDF) membranes. The NP was detected and subsequently incubated with anti-influenza A NP mouse monoclonal (QED Bioscience, Inc. San Diego, CA) or anti-influenza B NP mouse monoclonal antibodies (Japan Millipore, Tokyo, Japan). After washing and subsequent incubation with horseradish peroxidase coupled to anti-mouse immunoglobulin G (IgG), the immunoreactive proteins were visualized by an ECL detection kit (Amersham Biosciences, Piscataway, NJ).
Northern blot analysis
Total RNA was extracted from AB-1-, BNP-52-, and ANP-1496-shRNA-expressing MDCK cells after using Trizol reagent according to the manufacturer’s instructions (Invitrogen). Small RNAs <200 nt were separated concentrated using PureLink™miRNA Isolation Kit according to the manufacturer’s instructions (Invitrogen). Small RNAs 5 µg samples were loaded onto a 15% (w/v) polyacrylamide/7 M urea gel. After transfer to a Hybond-N™ nylon membrane (GE Healthcare Bio-Sciences Corp., Piscataway, NJ), synthetic LNA/DNA oligonucleotides (ANP-1496: 5′-GAATCTTATTTCTTCGGAG-Biotin-3′ and BNP-52: 5′-CCAGAAGAAACAACTTCTG-Biotin-3′) complementary to the antisense strand of the AB-1-shRNA were used as probes. The membranes were prehybridized for 1 hours in North2South hybridization buffer (Pierce) at 55°C and hybridized overnight to the 3′-biotin labeled LNA/DNA probe (30 ng/mL of hybridization buffer). Four posthybridization washes were done at 20 minutes each at 65°C with 2× SSC (1× SSC is 0.15 M NaCl plus 0.015 M sodium citrate–0.1% sodium dodecyl sulfate (SDS). Detection of LNA/DNA/RNA hybrids was carried out using the North2South chemiluminescent detection system (Pierce).
PCR and analysis of amplified products
Total DNA was purified from MDCK cells and the presence of AcNPV (moi = 50) or rBV-ANP-1496, BNP-52, AB1sh, and AB2sn (moi = 50) was detected by polymerase chain reaction (PCR) amplification of the viral genes vp39 and gp64. The sequences of the specific primers were as follows: vp39, 5′-TATGGCGCCGCGACAAATGAGAGTT-3′ (sense) and 5′-TTCGCCTGCAACACCAGGCGCAGG-3′ (antisense); gp64, 5′-GGGTCCGTACAAGATTAAAACTTGGACAT-3′ (sense) and 5′-CGCTCAGACTGGTGCCGACGCCGCCAAAT-3′ (antisense); G3PDH, 5′-TCCACCACCCTGTTGCTGTA-3′ (sense) and 5′-ACCACAGTCCATGCCATCAC-3′ (antisense).
Results and Discussion
Construction of influenza B NP gene-targeted mono- and bispecific short-hairpin siRNA–expressing vectors
The NP protein is required for elongation and antitermination of nascent complementary RNA (cRNA) and vRNA transcripts (Beaton and Krug, 1986; Shapiro and Krug, 1988). Without newly synthesized NP, further viral transcription and replication are blocked, as is virion production. Therefore, we designed siRNA specific for the NP gene. To determine whether influenza B virus NP gene expression could be inhibited by this siRNA, we designed six 21-nucleotide (nt) siRNA sequences for DNA from different regions of the NP gene, according to the criteria used in influenza A NP gene-targeted siRNA (ANP-1496), and cloned them under the control of the U6 promoter (Lee et al., 2002) into a pSV2-neo vector. The resultant plasmids (pSV2neoU6-BNP-52, pSV2neoU6-BNP-273, pSV2neoU6-BNP-873, pSV2neoU6-BNP-1178, pSV2neoU6-BNP-1237, pSV2neoU6-BNP-1492, pSV2neoU6-ANP-1496, and pSV2neoU6-ran-shRNA) synthesized siRNA under the control of the U6 promoter (Lee et al., 2002; Suzuki et al., 2008), and directed the synthesis of a Pol III–specific RNA transcript, starting with guanosine at position one, as required for efficient transcription initiation from the U6 promoter. The 5 thymidines on the plasmid DNA that functioned as a termination signal for Pol III were added at the 3′ end. The resulting RNAs had a guanosine residue at the 5′ end, and 2 complementary 19-nt sequence motifs in an inverted orientation, separated by an 8-nt loop of nonhomologous sequence (5′-UUCAAGAGA-3′; Anderson et al., 2003). This RNA was predicted to fold back to form an shRNA with a 3′ overhang of 2 uridines and a 9-nt loop, and, when cleaved with endonuclease was expected to generate an siRNA of 21 bp with a guanosine at the 5′ end and 2 uridines overhanging at the 3′ end (Fig. 1A and 1B).
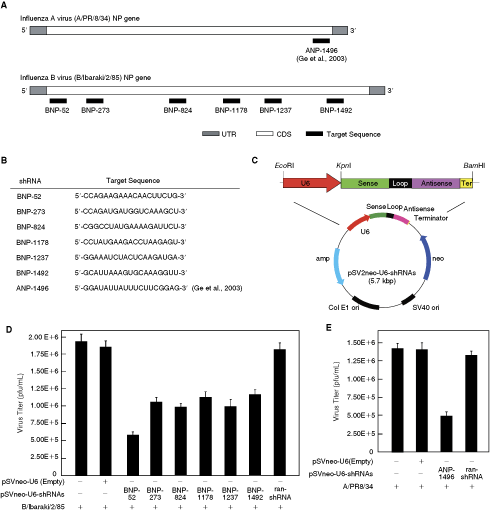
siRNA interference with the production of influenza viruses A and B in Madin-Darby canine kidney (MDCK) cells. (
Inhibition of influenza B NP gene expression by shRNA
To determine whether the influenza B NP gene-targeted siRNAs could suppress the production of NP protein, MDCK cells were transfected with the shRNA-expressing vector (pSV2neoU6-BNP-shRNAs) using FuGENE 6, followed by B/Ibaraki virus infection, and the virus titers in the culture supernatants were measured. High virus titers were detected in cultures with the control vectors pSV2neoU6-ran-shRNA and pSV2neoU6-ter. Although inhibition of NP protein production occurred with all 6 constructs, the greatest inhibitory effect occurred with the siRNA targeting the 52–71 nt region of the influenza virus B NP gene (65%; Fig. 1D). We also examined the effect of the siRNA targeting the 1,496–1,515 nt region of the influenza virus A NP gene. As expected, low virus titers were detected in cultures transfected with pSV2neoU6-ANP-1496 (Fig. 1E). By contrast, the control vectors (pSV2neoU6-ran-shRNA and pSV2neoU6-ter) had no inhibitory effect on influenza A/PR/8/34 virus. These results point to a sequence-specific inhibitory effect of shRNA.
Downregulation of influenza viruses A and B by bispecific short-hairpin siRNAs
Next, we designed bispecific short-hairpin siRNA constructs against influenza NP genes to inhibit the expression of the corresponding viral protein. Bispecific short-hairpin siRNA constructs, containing an 8-nt intervening spacer (Anderson et al., 2003), targeted against either B/Ibaraki NP or A/PR/8/34 NP, were expressed by pSV2neoU6-ANP-1496/BNP-52 (AB1sh and AB2sh). Intracellular processing of bispecific short-hairpin siRNA constructs was reported previously (Sioud and Leirdal, 2002; Anderson et al., 2003). More recently, Gupta et al. reported that chimeric siRNA-riboxyme interfered strongly with the expression of target gene expression both RNA and protein levels (Gupta et al., 2008). The designs of the bispecific short-hairpin siRNA constructs are shown in Figure 2A. The resultant plasmids (pSV2neoU6-AB1sh and pSV2neoU6-AB2sh) synthesized shRNA under the control of the U6 promoter. To investigate whether influenza A and B virus NP gene targeting bispecific short-hairpin RNAs can be digested to monospecific products of expected size, monomeric siRNAs were performed by Northern blot analysis in AB1sh expressing MDCK cells. After 48 h, bispecific-short-hairpin siRNAs yielded products ∼20 nt, the expected size of monomeric siRNAs [Fig. 2B, i (oligonucleotide probe, ANP-1496, lane 1: ANP-1496; lane 2: AB1sh); ii (oligonucleotide probe, BNP-52, lane 1: BNP-52; lane 2: AB1sh)].
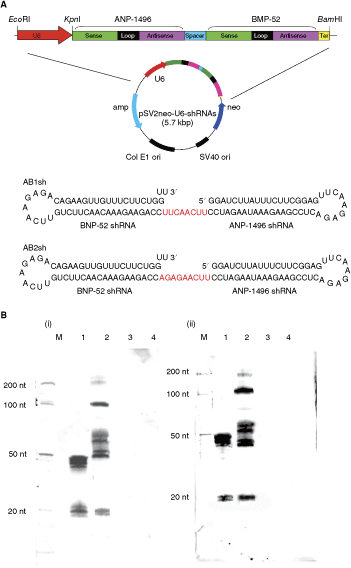
Effects of bispecific short-hairpin siRNA on influenza A and B virus production in Madin-Darby canine kidney (MDCK) cells. (
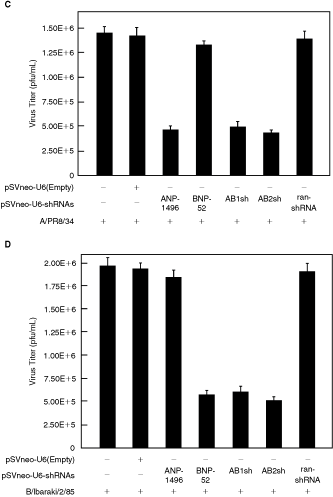
(Continued) (
Next, to determine whether bispecific short-hairpin siRNAs targeting influenza A and B virus NP genes could suppress the expression of both influenza A and B virus NP proteins, MDCK cells were cotransfected with plasmids expressing the shRNA, bispecific short-hairpin siRNA using FuGENE 6, followed by A/PR/8/34 virus or B/Ibaraki virus infection, and the virus titers in the culture supernatants were measured. Figure 2C shows that AB1sh and AB2sh, and ANP-1496 inhibited virus production in the A/PR/8/34 virus-infected MDCK cells, but not BNP-52. By contrast, the control vectors (pSV2neoU6-ran-shRNA and pSV2neoU6-ter) had no inhibitory effect of virus production in the A/PR/8/34 virus-infected MDCK cells. Furthermore, we examined the inhibition of virus production in the B/Ibaraki-infected MDCK cells. The NP gene targeting U6-shRNAs showed the sequence-specific inhibitory effects of the B/Ibaraki virus, but not ANP-1496 (Fig. 2D). By contrast, the control vectors (pSV2neoU6-ran-shRNA and pSV2neoU6-ter) had no inhibitory effect of virus production in the B/Ibaraki virus. These results suggest that the bispecific short-hairpin siRNA induced the sequence-specific inhibition of either influenza A or B virus. We also tested the 2 different spacer sequences, and found no significant differences between them (Fig. 2C and 2D). Thus, our results confirmed the ability of the bispecific constructs to effectively downregulate both the influenza viruses A and B.
Downregulation of influenza viruses A and B by baculovirus-mediated short-hairpin siRNA
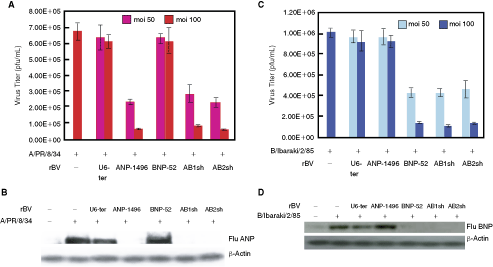
To characterize the efficiency of baculovirus as a vehicle for gene therapy, we selected the NP gene of influenza viruses A and B as the target site (Fig. 3A), and constructed a recombinant baculovirus vector expressing the shRNAs and bispecific short-hairpin siRNAs. Figure 3A shows the baculovirus transfer vectors used in this study. The baculovirus transfer vectors pVL1393-ANP-1496 and pVL1393-BNP-52 (monospecific-shRNAs), and pVL1393-ANP-1496/BNP-52 (AB1sh and AB2sh) carried NP-shRNA under the control of the Pol III, U6 promoter. The recombinant baculovirus containing the shRNA genome (Ac-shRNA) was propagated following previously published procedures (Abe et al., 2003; Suzuki et al., 2008). The viruses were produced at high titers, ranging from 1.5 × 108 to 1.2 × 109 pfu/mL, and the expression of the gp64 and vp39 genes was detected in the baculovirus vector–infected MDCK cells by PCR analysis (Fig. 3B).
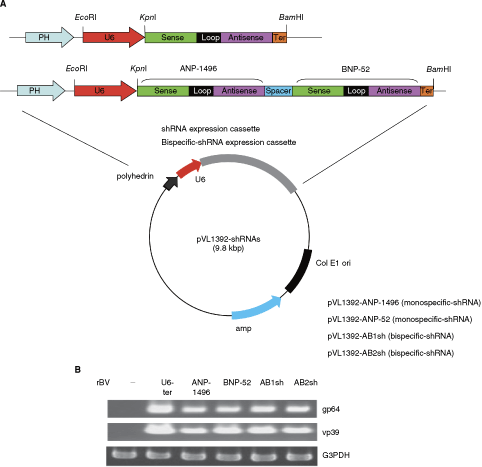
(
To determine whether bispecific short-hairpin siRNAs targeting influenza A and B virus NP genes can suppress the production of these viruses, MDCK cells were infected with A/PR/8/34 or B/Ibaraki (moi = 0.01). After 6 hours, the influenza virus–infected MDCK cells were infected with shRNA or bispecific short-hairpin siRNA expression baculovirus (moi = 50 or 100). At 4 days postinfection, the culture supernatants were harvested and assayed to determine the virus titers by plaque assay. Figure 4A showed that rBV-AB1sh and AB2sh, and ANP-1496 strongly inhibited virus production in the A/PR/8/34 virus-infected MDCK cell, whereas high virus titer was detected with rBV-BNP-52 infected cells. The influenza B virus production was significantly reduced by infection with rBV-BNP-52, rBV-AB1sh, and rBV-AB2sh, but not with rBV-ANP-1496 (Fig. 4C). These results indicate that the NP-targeting bispecific short-hairpin siRNA significantly inhibited the production of both influenza viruses A and B, whereas the empty baculovirus vector alone has no such effect. Furthermore, there was a direct correlation between the level of the virus and NP protein production (Fig. 4B and 4D).
Baculovirus-mediated bispecific short-hairpin siRNAs inhibit the accumulation of influenza A and influenza B virus NP mRNA. (
In conclusion, we have demonstrated the inhibition of influenza viruses A and B production by baculovirus-mediated bispecific short-hairpin siRNA expression vectors. This multiple vector showed remarkable ability to cope with both influenza virus A and B. Baculovirus-mediated bispecific short-hairpin siRNAs targeting the NP proteins of influenza viruses A and B have been shown to be particularly potent in inhibiting influenza virus production in cell lines, whereas that of the empty baculovirus vector showed no such effect. Now baculovirus can be used as an efficient vector for gene delivery mammalian cells for a wide variety of applications.
Footnotes
Acknowledgments
We are grateful to Miss M. Ishikawa and A. Ohno for excellent technical assistance. We wish to thank Dr. M. Kitajima (Benaroya Research Institute, Seattle, WA) for helpful discussions. This work was supported, in part, by a grant from the Research and Development Program for New Bio-industry Initiatives, by a Grant from Supporting Program for Creating University Ventures from Japan Science and Technology Agency, and by Grant-in-Aid for High Technology Research No. 09309011 from the Ministry of Education, Science, Sports, and Culture, Japan.