
Abstract
Cotransfection of a mixture of siRNAs species is typically used when simultaneous targeting of more than one mRNA is required. However, competition between siRNAs could occur and reduce the activity of some siRNAs within the mixture. To further study the factors affecting the degree of competition between siRNAs, we cotransfected luciferase targeting siRNAs with various irrelevant (ie, nonluciferase targeting) siRNAs into cells and examined differences in their competition profiles by assessing the effect on luciferase expression. We show that the degree of competition varies between irrelevant siRNAs and occurs at the point of RISC loading. Although the competition profile appears to be related to the calculated RNA-induced silencing complex (RISC) loading potential, empirical testing is required to confirm the competitive effects. We also observed reduced competition with siRNAs in the Dicer-substrate format, presumably due to more efficient RISC loading as a consequence of the physical transfer of the processed siRNA from Dicer.
Introduction
P
While it is generally possible to target more than one mRNA in the cell by cotransfecting a mixture of siRNAs, several studies have shown that the knockdown activity of some siRNAs in the mixture is compromised due to competition between siRNAs (Koller et al., 2006). Even inactive siRNAs have been shown to compete with active siRNAs, resulting in reduced knockdown of the latter in a dose-dependent manner (Holen et al., 2002; McManus et al., 2002; Formstecher et al., 2006; Stierle et al., 2007). The factors determining the extent of competition between siRNAs are presently unknown although it has been suggested that this effect may vary, depending on the cell type (Vickers et al., 2007). Although most studies looking at siRNA competition have only examined a limited number of siRNAs, a recent report has suggested that the sequence composition of the 5′ region of the guide strand may be a determining factor (Yoo et al., 2008).
In this study, we wish to identify differences in competition profile between luciferase specific siRNAs and a range of irrelevant siRNAs when cotransfected in cells expressing the firefly luciferase target gene. The degree of competition was assessed by the effect on luciferase knockdown activity. We found that siRNAs have different competitive characteristics that appear related to their RISC loading potential as assessed by scores of thermodynamic duplex asymmetry. We also show that competition between siRNAs in the Dicer-substrate format is reduced and that this type of siRNA is more competitive when co-delivered with siRNAs of the conventional 21 bp format.
Materials and Methods
SiRNAs
SiRNAs targeting different sites of the firefly luciferase (Luc2—genebank accession # AAV52875) mRNA were designed using Invitrogen’s web-based conventional siRNA design algorithm. We also selected a series of siRNAs, mostly from the literature, which do not target the firefly luciferase (Luc2) mRNA to be used as the irrelevant siRNAs in the competition studies (Supplementary Table 1; Supplementary materials are available online at http://www.liebertpub.com/). We specifically avoided siRNAs that target endogenous genes as these might have nonspecific effects in the cell that could indirectly alter luciferase expression. Based on a luciferase targeting siRNA (ie, siGL4.1-1), we synthesized an siRNA duplex by base inversion at the 9th, 10th, and 11th positions (relative to the 5′ ends) [ie, siGL4.1-1 (mut)] to inhibit the mRNA cleavage activity. The effect of this siRNA in competition was assessed. In addition, we also designed and synthesized Dicer substrate siRNAs version of siGL4.1-1 and several of the irrelevant siRNAs according to Amarzguioui et al. (2006; Amarzguioui and Rossi 2008).
Cell culture and transfection
A549 and U87 cells were purchased from American Type Tissue Culture and maintained in DMEM containing 10% fetal bovine serum and penicillin/streptomycin. A549 cells stably expressing the firefly luciferase target gene (ie, A549-Fluc) were generated by transfection with the Luc2 gene under CMV promoter control, followed by selection and maintenance of the cells in the above media supplemented with puromycin. For siRNA competition studies, A549-Fluc cells were seeded at 6000 cells/well in a white luminescence plate (Greiner Bio-one, Germany). The next day, each well was replenished with 80 μL of fresh culture media (without puromycin) and cells were transfected with siRNAs either alone or in combination (at 5–45 nM final concentration) in OptiMEM containing 2 μg/mL of Lipofectamine2000 (Invitrogen, Carlsbad, CA) using manufacturer’s instructions. All experiments included Lipofectamine2000-alone transfected cells (ie, mock-transfected) as controls and transfections were performed in triplicate. Luciferase expression was analyzed 48 hours post-transfection.
Luciferase expression analysis
The luciferase expression was determined using the Luciferase Assay System® (Promega, Madison, WI) as per manufacturer’s instruction for the 96-well assay format with the exception of using 30 μL of lysis buffer and luciferase substrate solutions. Luciferase measurements were performed on a FluoStar Optima plate reader (BMG LabTech). With the values averaged and expressed as a percentage of the mock-transfected sample.
Calculation of siRNA strand loading potential
The thermodynamic stability of siRNA ends was estimated using a sequence-based method as previously described (Tafer et al., 2008) except we used the first 4 nucleotides of the 5′ ends on each strand of the siRNA duplex as these positions are critical in determining the strand loading bias (Schwarz et al., 2003). Briefly, for each siRNA, the number of G/C residues of the first 4 nucleotides at the 5′ ends was determined and the difference relative to the passenger strand 5′ end was calculated. The analysis typically generated positive or negative scores suggesting the preferential loading of the guide or passenger strands, respectively.
Results
Competition profile varies with siRNAs
To examine the potential for competition between siRNAs, we performed our co-transfection studies in A549 cells stably expressing the firefly luciferase gene (A549-Fluc). As expected, transfection of A549-Fluc cells with up to 40 nM of the irrelevant siRNAs alone showed no effect on luciferase activity. In contrast, transfection with 5 nM of the luciferase-specific siGL4.1-1 effectively suppressed reporter activity by 90% relative to the untransfected controls (Fig. 1A). Cotransfection of cells with siGL4.1-1 and the irrelevant siRNAs produced a range of inhibitory effects that were dependent on the concentration ratio and the nature of the irrelevant siRNA (Fig. 1B). Specifically, some of the irrelevant siRNAs such as siGC47, NP_1496, siHTLV-1, and siLuc+ (white bars) had little effect on siGL4.1-1 activity even at the highest concentration ratio tested (ie, 40 nM irrelevant siRNA + 5 nM siGL4.1-1), indicating that these siRNAs are weak competitors. In contrast, siEGFP, siCyan1, and siCyan2 were strong competitors and cotransfection with these siRNAs affected siGL4.1-1 activity at all of the ratios tested (black bars). Of these, siCyan2 (black hatched bar) was the strongest competitor as its effect on siGL4.1-1 knockdown activity was clearly apparent even at the lowest ratio of cotransfected siRNAs (1:1). The siGL4.71-1, siGL4.7-2, siHIV-Tat, and GFP_949 siRNAs (grey bars) produced intermediate effects on siGL4.1-1 activity, with competition apparent only at the higher concentration ratios.

Varying degree of competition between Luciferase targeting and irrelevant siRNAs. A549-Fluc cells were transfected with (
We repeated the experiments using a targeting siRNA designed to a different site on the luciferase mRNA sequence. The siRNA siGL4.1-2 also caused ∼90% knockdown of luciferase activity at 5 nM (Fig. 1A). Cotransfection experiments using siGL4.1-2 and the irrelevant siRNAs produced a similar pattern of competition of luciferase knockdown activity as observed with siGL4.1-1, although the level of luciferase reduction in this set is slightly reduced (Supplementary Fig. 1). The results suggest that the competition depends on the siRNA present in the mixture and is independent of the target site.
SiRNA RISC loading is a point of competition
Competition between siRNAs is likely to occur at the level of incorporation of the guide strand into the RISC machinery and may, therefore, be influenced by the kinetics of strand loading. To show that RISC loading is a prominent site of competition between siRNAs duplexes, we performed siRNA competition assays using siGL4.1-1 (mut) and RISC-Free siRNA (Dharmacon) as competitors. The former is identical to siGL4.1-1, except for mutations introduced at positions 9 to 11. This mutant version should be equally efficient in RISC incorporation but has no target cleavage activity of the siRNA (Coburn and Cullen, 2002) (Fig. 2A). Similarly, the RISC-Free siRNA has no RNAi activity as it is designed not to be loaded by the RISC complex (Fig. 2A). The modification or sequence of this siRNA that results in the lack of RISC uptake is not knows as it is proprietary. However, it has been shown that the 5′ end of the guide strand must be phosphorylated for RISC incorporation and siRNA activity and this could be prevented by the presence of modifications such as methoxy groups (Nykanen et al., 2001; Martinez et al., 2002). Therefore similar modification (and possibly others) maybe present in the RISC-Free siRNA sequence to prevent its recognition by RISC as a bona fide substrate. When siGL4.1-1 and siGL4.1-1 (mut) were used in the competition assay, a decrease in knockdown efficacy of the luciferase target with increasing ratio was observed, similar in profile to an intermediate-strong competitor (Fig. 2B, white bars). On the other hand, competition assay with the RISC-Free siRNA showed no effect of siGL4.1-1 knockdown efficacy even at the highest concentration of competitor used (Fig. 2B, black bars). We expect that in this latter scenario, that the RISC complex predominantly incorporates the siGL4.1-1 siRNA in all of the different ratios tested. Altogether, these results indicate that the efficacy of loading into RISC is a major factor influencing competition between siRNA duplexes.
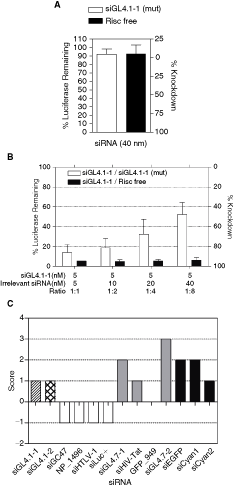
Competition between siRNAs occurs at the point of RISC loading and is related to siRNA asymmetry scores. A549-Fluc cells were transfected with siGL4.1-1 (mut) and RISC-Free siRNAs (see text for details) either (
To understand the different competitive potential of the various irrelevant siRNAs we tested above (Fig. 1B), we examined their relative efficacy of RISC loading by examining the 5′ end thermodynamic stability. It has been shown that the thermodynamic stability of the siRNA duplex ends is a major determinant for RISC incorporation efficacy with the strand having the lowest stability in its 5′ region of the guide strand being preferentially loaded (Khvorova et al., 2003; Schwarz et al., 2003). Our analysis confirmed that, in general, there was a tendency for the irrelevant siRNAs with intermediate to strong competitive ability (except GFP_949) to have positive asymmetry scores similar to siGL4.1-1 or siGL4.1-2 (Fig. 2C). Weakly competitive irrelevant siRNAs had negative asymmetry scores suggesting less preferential loading of the guide strand of these duplexes into the RISC complex, which could be outcompeted by the more efficiently loaded siRNA.
Dicer-substrate siRNAs have reduced competitive potential
In the experiments above, we used 21 bp siRNAs with 2 base 3′ overhangs (ie, “conventional siRNAs”) that are likely to be loaded directly into RISC without prior processing by Dicer. More recently, RNAi activity has also been achieved using Dicer-substrate siRNAs (Kim et al., 2005). These siRNAs are 25–27 bp dsRNA that are recognized and processed by Dicer prior to loading into RISC. Dicer processing has been postulated to result in enhanced potency through “hand-off” transfer from Dicer to RISC. This would be expected to increase the competitive potential relative to conventional siRNA as these could be preferentially loaded into RISC. To examine this hypothesis, we used Dicer-siRNA formats corresponding to the luciferase-targeting siRNA (ie, D-siGL4.1-1) and siRNAs representing weak (D-NP_1496), intermediate (D-siGL4.7-1), and strong (D-siCyan2) competitors. Transfection of A459-Fluc cells with the Dicer substrate version of siGL4.1-1 (D-siGL4.1-1) resulted in ∼90% suppression of luciferase activity at 5 nM (Fig. 3A and 3B), similar to siGL4.1-1. Importantly, the Dicer substrate versions of the irrelevant siRNAs alone did not inhibit luciferase activity (data not shown). We performed competition experiments using D-siGL4.1-1 when cotransfected with each of the irrelevant Dicer substrate siRNAs at the highest ratio. Overall, although some degree of competition was observed (Fig. 3A), it was clearly not as extensive as that observed in the corresponding experiments using the conventional format siRNAs (Fig. 1B). In particular, D-siGL4.1-1 retained strong activity even in the presence of D-siGL4.7-1 and D-siCyan2 (∼70% suppression of luciferase). The reduction in competition was not limited to the siRNAs used above as we also observed similar increase in D-siGL4.1-1 activity (≥60% knockdown) in the presence of other the Dicer-substrate version of irrelevant siRNAs (Supplementary Table 1). The ability of Dicer substrate siRNA to retain activity in the competition experiments suggests that these are more efficiently loaded, consistent with the “hand-off” mechanism proposed (Tomari and Zamore, 2005). To examine whether Dicer substrate siRNA are more resilient to competition from conventional format siRNA, we performed competition experiments between the luciferase-targeted D-siGL4.1-1 and the conventional irrelevant siRNAs (ie, NP_1496, siGL4.71-1, and siCyan2). Indeed, the Dicer-substrate siRNA had more activity in the presence of the irrelevant siRNAs as compared to siGL4.1-1 (Fig. 3B). The sustained knockdown activity was most apparent in cotransfections of D-siGL4.1-1 and the strong competitor siCyan2, which resulted in ∼40%–50% luciferase knockdown. In contrast, there was a more modest effect in cells cotransfected with the weak and intermediate competitors. We also tested the reverse scenario in which the Dicer-substrate formats of selected irrelevant siRNAs were cotransfected with siGL4.1-1. As expected, we observed that the D-NP_1496, D-siGL4.7-1, and D-siCyan2 Dicer-substrate siRNAs caused at least the same or more reduction in the siGL4.1-1 knockdown activity (Fig. 3C) than their corresponding conventional formats.
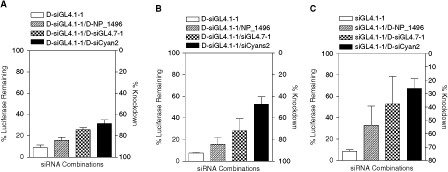
The effect of siRNA formats on competition. A549-Fluc cells were transfected with (
Discussion
To date, several reports have shown that cotransfection of siRNAs could result in reduced knockdown activity compared to singly-transfected siRNAs. It is thought that this is caused by competition between siRNAs—most likely for the RNAi machinery—which is still not well understood. However, it is not clear whether all siRNAs possess similar competitive ability or the competition is a function of the siRNAs present. In this study, we examined 11 different nonluciferase targeting siRNAs and found that these siRNAs have varying ability to compete (ie, negatively affect) with luciferase-targeting siRNAs activity in a dose-dependent manner. We also showed that the competition is most pronounced for siRNAs with high end-asymmetry scores, which is thought to reflect the relative ability of the guide strand for incorporation into RISC. Schwarz et al. (2003) have shown that the thermodynamic stability of the siRNA 5′ ends has a large influence on the selection and efficiency of loading of the siRNA guide strand into the RISC complex (Schwarz et al., 2003). Since 21 bp siRNAs could directly associate and be incorporated into the RISC complex, it could be expected that siRNAs with favorable 5′ end thermodynamic properties would be preferentially loaded. Even so, it is likely that empirical testing is required to identify the degree of competition between specific siRNAs. In particular, the direction of the loading bias may not be relevant to the studies of competition, particularly when the target sequence is not present and it may be that the absolute value of the loading bias score is more relevant.
Our study showed that irrelevant siRNAs produce a concentration-dependent competitive effect on the luciferase siRNA activity when cotransfected. While this may simply reflect relative affinities of the various siRNA for the RISC machinery, it may also reflect a capacity limitation (ie, saturation of RISC). Indeed, a recent study has shown that competition is reduced when siRNAs are expressed at very low levels using microRNA backbones (Castanotto et al., 2007). Therefore, cotransfection using highly potent 21 bp siRNAs is desirable to avoid saturating concentrations that might accentuate the effects of competition.
Recently, it was shown that cellular level of Ago2 is another factor on the degree of competition between siRNAs (Vickers et al., 2007). We have tested our competition assays in cells with low and high levels of Ago2 (similar to that used by Vicker’s et al.) but we did not observe significant differences in the competition (data not shown). It is possible that the siRNA formats used in the assays—that is, 21 bp with 2 base 3′ overhangs (used by us) versus the 19 bp blunt siRNA as used by Vickers et al. (2007)—may account for the discrepancy in the competition. Since the PAZ domain has the lowest affinity toward binding of blunt ended duplexes (Parker and Barford, 2006), siRNA in this format maybe more sensitive to reduced levels of Ago2. This difference in siRNA format uptake and the influence on siRNA competition should be further examined.
We also observed that competition between siRNAs could be reduced by cotransfecting with a Dicer-substrate format. This result is consistent with the fact that entry of these siRNAs into the RNAi pathway is not occurring at the limiting RISC incorporation step and, therefore, discrimination of siRNAs based on RISC loading efficiency is reduced (Tomari and Zamore, 2005). As a result of the physical transfer from Dicer processing into the RISC complex, Dicer-substrate siRNA could be expected to be more competitive as compared to conventional siRNAs. Although these Dicer-substrate siRNA may, therefore, offer a more efficient way of targeting multiple mRNA species, the advantages of this approach have to be explored against other factors. It is interesting to note that different irrelevant Dicer-substrate siRNAs caused varying effects on D-siGL4.1-1 activity suggesting that preferential uptake between these siRNAs exist.
Combinatorial siRNA approach is likely to be a desirable approach for regulating multiple targets associated with different human diseases. Like most other studies looking into siRNA competition, our assays were carried out using a highly expressed exogenous target. The results reported here are, therefore, relevant to situations where highly abundant mRNA targets are present (eg, during viral infections). We see that a combinatorial siRNA approach could potentially be useful in simultaneously protecting the cell against multiple infectious agents (eg, lung cells from various cold-causing viruses). However, further studies are required in using this approach to knockdown multiple endogenous genes with varying levels of expression.
This study indicates that the competition between siRNAs depends not only on the specific siRNAs present in the cocktail but also the format used. The competition between conventional siRNAs appears to be dictated by the efficiency of the guide strand loading into the RISC complex. In contrast, Dicer-substrate siRNAs appeared to display reduced level of competition and better activity in the presence of conventional siRNAs. Although more studies are needed, it is possible that a favorable activity could also be seen with siRNA in other formats (eg, shRNA and miRNA backbones), particularly those which reduce competition for certain components of the RNAi pathway. Overall, utilizing different siRNA formats maybe necessary to balance the competitive nature of some siRNA species and achieve an efficacious combinatorial RNAi approach.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.