
Abstract
The oligonucleotide Dz13, a DNA enzyme that cleaves c-Jun mRNA, is capable of inhibiting various model tumors in mice. However, to date, a thorough examination of its target specificity in tumor cells has not been performed. In addition, an evaluation of its safety in a mammalian whole organism system has not been carried out. Dz13 mutated oligonucleotides were designed and tested in a proliferation assay. Dz13 was also tested for its safety in vivo when administered intravenously in a bolus dose, or systemically in an in utero assay. While Dz13 down-regulated target gene (c-Jun) expression in human tumor cells, c-Jun siRNA failed to cause cell growth inhibition. Furthermore, alteration of contiguous G motifs in Dz13 flanking arms inhibits cell death activity, but removal of the same from the catalytic core can increase cell death activity. A 20mer (truncated) derivative Dz13 exhibited similar activity. Dz13 was not toxic to blood and solid tissues in adult or fetal mice, though slight hepatotoxicity was noted with histology. It was also void of adverse effects to the physiological processes of angiogenesis and apoptosis. Collectively, the data support the safety of Dz13 and its activity attributed to off-target effects.
Introduction
In human dermal microvascular endothelial cells (HMEC-1) and rat SV40-transformed rat smooth muscle cells, Dz13 is potently able to kill cells at doses as low as 0.1 μM (Rivory et al., 2006). RT-PCR was used to rule out c-Jun down-regulation by Dz13, and no attempt to examine levels of protein were made. The authors concluded that this cell growth inhibitory effect could be due to the G-rich 5′ sequence, though formation of a G quadruplex was ruled out. The importance of a G-rich 5′ sequence was confirmed later in HMEC-1 cells (Goodchild et al., 2007), which also demonstrated the requirement of a high proliferative index of human retinal pigmented epithelium (ARPE-19) cells for Dz13 activity to be exhibited at 0.1 μM oligonucleotide concentration. Dz13 was found to be selective to proliferating cells as opposed to those cells that had been contact-inhibited. Finally, the cell growth inhibitory effect of Dz13 on HMEC-1 cells was attributed to nonapoptotic programmed cell death, with cell death effectors from necrosis, apoptosis, and autophagy implicated (Gozar et al., 2008).
The above results with HMEC-1 cells (Gozar et al., 2008) contrast findings in various human cancer cells in which a clear indication of apoptosis is seen in tumor cells of OS (Dass et al., 2008a, 2008b, 2008c and LS (Dass et al., 2008d). Depending on the cell line, chemical inhibitors were employed to show that caspase-1, -2, -8, or -10 or combinations thereof (Dass et al., 2008a, 2008b, 2008c, 2008d,) were involved in causing tumor cell death, though these results were not confirmed in any other way.
Here, we demonstrate that c-Jun down-regulation via siRNA does not cause cancer cell growth inhibition. Instead, we observed the off-target effects of Dz13 in various tumor cell types, always causing a potent cell death. In addition, we demonstrate the safety of this oligonucleotide in adult and fetal mice, in blood and in organs. We also have discovered a shorter variant of Dz13, DzM14, which has equal or more activity against cancer cells, depending on the cell type.
Materials and Methods
Cells
Human prostate cancer cell line PC3, breast cancer cell line MDA-MB231, osteosarcoma cell line SaOS-2, and osteosarcoma cell line 143B were from the ATCC (Virginia, USA). Human osteosarcoma cell line SJSA-1 and liposarcoma cell line SW872 were from D. Thomas (Peter MacCallum Cancer Institute, Australia). Growth media (α-MEM) was supplemented with 10% FBS and 1% antibiotic–antimycotic. Cell lines were maintained up to 20 passages in a 37°C/5% CO2 incubator. Cells were ensured to be >95% viable prior to experiments.
DNAzyme transfection
Oligonucleotides were synthesized and prepared as before (Mitchell et al., 2004). For the “Dz13 sequence” studies, oligonucleotides were synthesized by Sigma. DNAzyme (0.8 μM) and other oligonucleotides were transfected with Fugene-6 liposomes (Roche Diagnostics, Indianapolis, IN) in completer medium for 24 hours prior to assaying. The FITC-Dz13 uptake study was performed as before in complete medium (Dass et al., 2002).
siRNA transfection
All siRNAs were from Santa Cruz Biotechnology, and transfected 1 hour prior to DNAzymes with Fugene-6 at 0.4 μM in complete medium. Cells were transfected for 25 hours prior to assaying.
Cell proliferation assay
Treatments were added to preseeded (2000 cells/well, 96-well plates, 24 hours) SaOS-2 cells in triplicates (Dass et al., 2008e). Treated cells were incubated for 24 hours before quantitation of cell viability by staining and subsequent counting of cell nuclei with 10 ng/mL DAPI and observation under glycerol and ×100 magnification using a Nikon Eclipse TE2000-U microscope (Nikon, Sydney, Australia) and imaged with SPOT Advanced software (SciTech, Melbourne, Australia). Alternatively, cell viability was assessed with the CTBlue assay (Promega, Sydney, Australia) using an absorbance of 570 nm on a BioRad Model 680-plate reader and Microplate Manager software. Blank readings at 600 nm were always subtracted as background.
Immunocytochemistry
To detect levels and localization of c-Jun and c-Fos proteins in the transfected human cancer cell panel, a published protocol (Tinel et al., 2007) was followed with minor modifications. Primary antibodies were from Santa Cruz Biotechnology, and secondary antibodies from Dako. After secondary antibody incubation, samples were treated with the Vectastain ABC kit (Vector Labs, Burlingame, CA). Samples were developed with DAB for 5 minutes and observed under glycerol.
Western blotting
To detect levels of c-Jun in the transfected human cancer cell panel, a published protocol (Ek et al., 2007a, 2007b) was followed. Post-transfection, cells were lysed with RIPA (150 mM NaCl, 50 mM Tris, 1 mM EDTA, 0.1% SDS, 1% Triton X-100, pH 7.4) buffer containing complete protease inhibitors (Roche). All primary antibodies were from Santa Cruz Biotechnology, and all HRP-conjugated secondary antibodies were from Dako. Bis–Tris (12%) polyacrylamide gels (Invitrogen, Carlsbad, CA), polyvinylidene fluoride (PVDF) membrane (Invitrogen), and an ECL kit (GE Healthcare, Sydney, Australia) and ECL hyperfilm (GE Healthcare) were used for electrophoresing, blotting, and detection of proteins, respectively.
Intravenous Dz13 safety study
Prior ethics approval was obtained from the St. Vincent's Health Animal Ethics Committee, adhering to the “Principles of Laboratory Animal Care” (NIH publication #85-23, revised in 1985). Five-week-old Balb/c mice were administered for 4 weeks, at once per week, a bolus of 5 mg/kg Dz13 or DzScr in a volume of 0.1 mL. This dose was chosen as it represents a dose that is normally used for systemic DNAzyme evaluation in vivo (Sun et al., 2002). At the end of the study, blood was analyzed on a Sysmex XE2100 instrument (Fahmy et al., 2003). Immediately posteuthanasia, animals were dissected and the states of livers and spleens were noted. In addition, as performed routinely for soft tissues (Dass et al., 2006; Ta et al., 2009) spleens of animals were observed histologically.
In utero safety study
Balb/C pregnant mice at E10 were subcutaneously implanted with model 1002 osmotic pumps (Durect Corp, Cupertino, CA) filled with Dz13 to deliver at a rate of 5 mg/kg/day. Pumps were primed for 12 hours prior to implantation (Walker et al., 1998). Mice were left for 4 days before fetuses, placentas, and tissues were processed histologically. Skin wounds were inflicted by a surgical (7 mm) incision on mice mid-dorsum and observed 4 days later and processed histologically as performed previously (Ek et al., 2007a). Limbs, tumors, and the juxtapositioned inguinal lymph nodes, livers and spleens were processed histologically (Ek et al., 2007b).
Statistical analysis
All data were analyzed using the 1-way Student's t-test with unequal variances.
Results
Dz13 chemistry, not target gene down-regulation, induces tumor cell death
To gauge the level of DNAzyme uptake (transfection efficiency), a FITC-conjugated Dz13 was employed. It enabled tracking of Dz13 inside cells (Fig. 1A), and a typical speckled, punctate cytoplasmic localization (Dass et al., 2002) was noted. To evaluate whether the target (c-Jun) is down-regulated in vitro, Dz13 was transfected into human cancer cells in culture. Reduced c-Jun levels mainly in the nucleus (Fig. 1B) and in a time-dependent manner in whole cell lysates were noted (Fig. 1C). However, when a c-Jun siRNA was employed, no reduction in cell number was noted (Fig. 2A) when the c-Jun target was down-regulated (Fig. 2B).
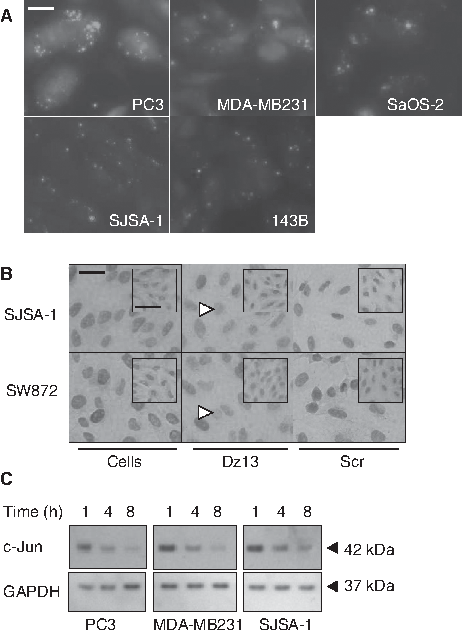
Dz13 down-regulates c-Jun in human cancer cells. (
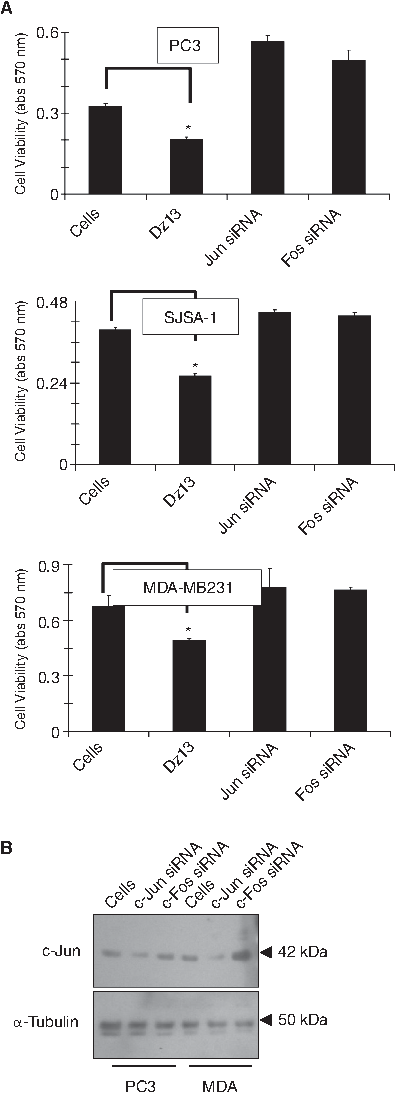
c-Jun down-regulation does not cause cell growth inhibition. (
Repositioning of contiguous G motifs in Dz13 flanking arms abrogates activity
To test whether cell growth inhibition is due to Dz13's unique sequence and chemistry, a 2-tiered series of Dz13-based oligonucleotides was designed. The first was aimed at contiguous G motifs (bolded for Dz13) with such motifs being altered at either one of the 5′ terminus or 3′ terminus arms flanking the catalytic core (Table 1). The alteration of contiguous G motifs in the Dz13 sequence inhibited cell death activity in 3 cell lines (Fig. 3A), even though G contiguity was simply relocated along the Dz13 sequence at either one of the flanking arms. Dz13M, a mutated catalytic core version of Dz13, demonstrated equal cell death activity as the parent molecule. Lengthening (11-mer catalytic core flanking sequence arms) or shortening (7-mer arms) of the sequence also decreased Dz13 (9-mer arms) potency. Two DNAzymes shown also to have antitumor effects, DzXL (Sun et al., 2002) and DzUPAR (Dass and Choong, 2008a), were not growth inhibitory, suggesting that the catalytic core was not the direct cause of the activity. As13, which is known to down-regulate c-Jun (Zhang et al., 2004), was also noninhibitory.

Potency of Dz13 is due to G motifs in flanking arms but not in catalytic core. (
Ti, inverted T base linkage.
In addition, a Dz13 sequence with an altered base in the catalytic core (DzM), the antisense versions of Dz13 and its scrambled sequence (Dz13Scr), a longer and shorter version of Dz13 based on arm length, and DNAzymes to Bcl-xL and the urokinase plasminogen activator receptor (uPAR) were tested alongside Dz13 and Dz13Scr (also called Scr elsewhere in this article).
Truncated catalytic core reveals importance of removal of G motifs for potency
Since Dz13M was as effective as Dz13, a second series of oligonucleotides, such that the catalytic core was truncated in either direction [starting from the G (bolded in Dz13 sequence) that was converted to C to alter Dz13 into Dz13M], with the flanking arm sequences constant, was designed and tested (Table 2). Interestingly, when the only 2 possible contiguous catalytic core G motifs were removed (bolded in DzM3 and DzM13), the sequence potently promoted cell death (Fig. 3B). A 20-mer sequence (DzM14) was identified that had equal activity to Dz13 in a p53-functional cell line (SJSA-1), but was more potent in a p53-null background (SaOS-2). Incidentally, the only difference between DzM14 and As13 (causes growth inhibition in the PC3 cell line only, Fig. 3A) is the replacement of a C in position 10 of the latter by GA in the former (Fig. 3C). A shorter oligonucleotide like DzM14 would be more cost effective for mechanistic and therapeutic studies in future, especially given its p53-non-discriminating action, possibly being able to exhibit antagonistic activity against the whole spectrum of neoplastic disorders. For the purpose of this study, further evaluation of DzM14 was not undertaken, since our focus was on the molecule Dz13, which has shown considerable activity in vivo (Dass et al., 2008a, 2008b, 2008c, 2008d; Zhang et al., 2004, 2006).
Ti, inverted T base linkage.
When the contiguous GG motifs (bolded) are removed, the sequence assumes a more toxic disposition.
Intravenous, bolus administration of Dz13 is nontoxic in adult mice
While several studies have shown the antiangiogenic (Zhang et al., 2004) and direct antitumor (Zhang et al., 2006; Dass et al., 2008a, 2008b, 2008c, 2008d) activity of Dz13, the safety profile of this molecule has not been reported. This data is critical if clinical studies are to be performed in future. Mice were administered for 4 weeks, at once per week, a bolus of 5 mg/kg Dz13 or DzScr intravenously. At the end of the study, no significant differences in blood cell composition (Fig. 4A) and clinical biochemistry (Fig. 4B) parameters, and spleen (Fig. 4C) as indicators of oligonucleotide toxicity were noted. The phenomena, observed for phosphorothioated (PO) oligonucleotides, of increased white blood cells (WBCs) and reduced red blood cells (RBCs) (Henry et al., 2004a) were not noted with Dz13 administration, while there was a tendency for reduced platelet counts, though this was not significant.

Intravenous (bolus) administration of Dz13 is nontoxic to blood in mice. (
Systemic, continuous Dz13 is nontoxic in pregnant and fetal mice
In a more sensitive assay for genotoxicity, pregnant mice were administered Dz13 continuously and systemically with the aid of osmotic pumps. The fetus of Balb/C pregnant mice administered systemically with Dz13 or DzScr neither died nor showed signs of gross toxicity (Fig. 5A). Similar findings were reported for a PO antisense molecule where no oligonucleotide was detected in the murine (Henry et al., 2004a) and rabbit (Henry et al., 2004a) fetal kidney and liver, and very little in expressed milk and fetal plasma. Thus, exposure to the fetus is expected to be minimal, but until now, this had not been directly evaluated for a 3′-inverted T oligonucleotide.
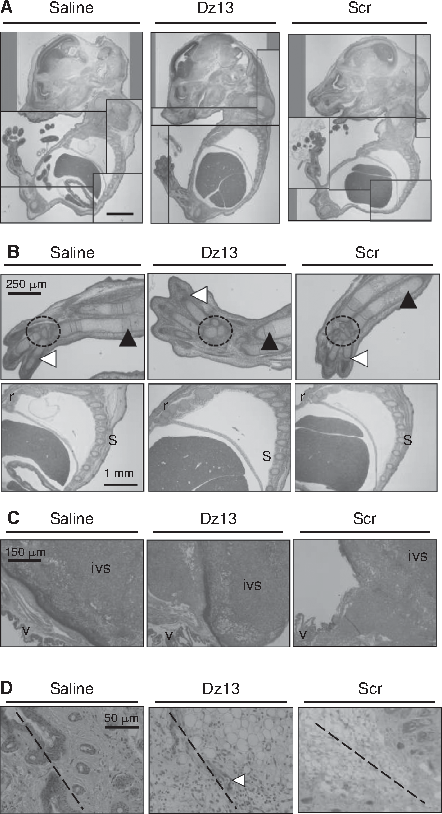
Systemic and continuous administration of Dz13 is nontoxic to mice in utero. (
Since Dz13 is capable of inhibiting tumor angiogenesis (Zhang et al., 2004), and as a measure of the effects of Dz13 on physiological angiogenesis in the mouse, 3 tissues were analyzed. These included fetal bone (Fig. 5B), placenta (Fig. 5C), and skin wound (Fig. 5D). No evidence of perturbed angiogenesis was seen in all 3 cases. Apoptosis is vital to proper tissue modeling in the young including the removal of interdigital web tissue in developing mouse paws (Jacobsen et al., 1996). No signs of derailed apoptosis were noted either (Fig. 5A).
Then attention shifted to the adult tissues. Bone was chosen as the cell lines evaluated in this study were of tumors arising in or metastasising to the bone. Dz13 may well be administered in bone in future to control these neoplasms. Skeletal muscle was chosen for evaluation as bone tumors often breach through the cortex and invade into the surrounding muscle. Thus, it is possible that Dz13 may be administered in muscle in future for therapy of such tumors. The liver, spleen, and lung were chosen since they constitute the major part of the reticuloendothelial system (RES), involved in immune surveillance in the body, and the liver accounts for most of the oligonucleotides administered in vivo (Agrawal et al., 1991). Kidney was evaluated as oligonucleotides are mostly excreted through this organ as are the metabolites (Agrawal et al., 1991). Thus, the tissues analyzed included the adult bone (Fig. 6A), skeletal muscle (Fig. 6B), liver (Fig. 6C), spleen (Fig. 6D), lung (Fig. 6E), and kidney (Fig. 6F). Only in the liver was toxicity noted, mainly as cell death, most probably via necrosis (Fig. 6C), though this was not widespread and hepatomegaly, as noted with other oligonucleotides (Henry et al., 2004a, 2004b) was absent. Furthermore, there was a mild toxic effect with the scrambled oligonucleotide as well. Nevertheless, because of the hepatotoxicity, a systemic route of administration was not chosen for further studies.

Systemic and continuous administration of Dz13 is nontoxic in adult mice. (
Discussion
As mentioned above, in the field of cancer research, and particularly in nucleic acid-based cancer therapeutics, Dz13 is an interesting molecule due to its antagonistic activity against a range of neoplasias (Zhang et al., 2004, 2006; Dass et al., 2008a, 2008b, 2008c, 2008d). To advance a potential lead compound further toward clinical evaluation, amongst other things, the acquisition of 2 major datasets are mandatory: safety studies and defining exact mechanisms of activity. This study aimed to do just this.
As reported numerous times before (Dass and Choong, 2008b), this study confirms the activity of the Dz13 molecule toward its target (c-Jun) in both cells that were actively growing continuously, or that were stimulated to grow post-serum starvation. Initially, this was thought to be causing the potent cell death effect in tumor cells. However, when siRNA to c-Jun was employed, growth was not inhibited. Thus, a nonspecific mechanism (off-target) effect was thought to be responsible for causing this cancer cell growth inhibition. Interestingly, this effect seems to be relevant to tumor cells selectively, as Dz13 administered in situ does not have adverse effects on bone or muscle in mice (Dass et al., 2008e), nor does it affect the ability of primary human bone marrow cells to proliferate in vitro (Dass et al., 2008b). However, a preliminary search for complementary sequences on BLAST (http://blast.ncbi.nlm.nih.gov/Blast.cgi) failed to highlight suspects that both Dz13 and its shorter derivative, DzM14, could be binding to.
To directly test this, and in view of the results with HMEC-1 cells (Rivory et al., 2006; Goodchild et al., 2007; Gozar et al., 2008), we embarked on designing a 2-tiered series of oligonucleotides, mutated from the parent Dz13 molecule. The first aimed at contiguous G motifs with such motifs being altered at either one of the 5′ terminus or 3′ terminus arms flanking the catalytic core. Similar to the findings with HMEC-1 cells, the alteration of contiguous G motifs in the Dz13 sequence inhibited cell death activity, even though G contiguity was simply relocated along the Dz13 sequence at either one of the flanking arms. In the HMEC-1 studies, only the 5′-terminal G-rich motifs were implicated to be important for cell growth inhibition (Goodchild et al., 2007). Dz13M, a mutated catalytic core version of Dz13, demonstrated equal cell death activity as the parent molecule, which is akin to the findings in HMEC-1 cells (Rivory et al., 2006). In addition, lengthening or shortening of the sequence decreased Dz13 potency in the present study.
Since Dz13M was as effective as Dz13, a second series of oligonucleotides, such that the catalytic core was truncated in either direction with the flanking arm sequences constant, was tested. Contrary to expectation, when the only 2 possible contiguous catalytic core G motifs were removed, the sequence potently promoted cell death, at least at an equal level to Dz13. Interestingly, a 20-mer sequence (DzM14) was identified that had at least equal activity to Dz13 in a p53-functional cell line (SJSA-1), but was more potent in a p53-null background (SaOS-2). A shorter oligonucleotide like DzM14 would be more cost effective for mechanistic and therapeutic studies in future, especially given its p53-nondiscriminating action, possibly manifesting a therapeutic affect over the whole spectrum of neoplastic disorders. As is commonly known, p53 is a gene that is frequently mutated in cancers (Lane, 1994) and its protein is required for the proper functioning of various therapeutic agents currently in clinical use or being tested (Steele and Lane, 2005).
Off-target effects due to gene-silencing oligonucleotides have led to an entire new field—moligonucleotide-mediated immunomodulation (Dorn and Kippenberger, 2008), though this is more relevant in a whole organism in which the effect of immune cells postadministration of oligonucleotides can be gauged. In vitro, strict off-target effects absent of an immune component can be observed for a variety of oligonucleotides, including siRNA and antisense (reviewed in Alvarez-Salas, 2008). To take one of the initial promising antisense molecules as an example; the cell growth inhibitory effects of the Bcl-2 down-regulating antisense, G3139, can be attributed to specific CpG motifs in its sequence (Lai et al., 2003). In fact, it has been found that in melanoma cells, the pro-apoptotic activity of G3139 is independent of its ability to down-regulate its target gene (Lai et al., 2005; Benimetskaya et al., 2006). Thus, there is a definite need to properly examine both target and off-target effects of each oligonucleotide being developed into a potential therapeutic agent, which includes a thorough safety examination in appropriate models in vivo.
The safety profile of Dz13 has been largely unexplored. This data is critical if primate and then clinical studies are to be performed in future. Mice were administered for 4 weeks, at once per week, a bolus of 5 mg/kg Dz13 or DzScr intravenously. At the end of the study, no significant differences in blood parameters (cellular composition and liver function) and spleen were noted. The spleen was chosen as it serves as a sensitive organ that changes its tissue architecture when challenged with an oligonucleotide capable of inducing an immune response (McIntyre et al., 1993). The phenomena, observed for phosphorothioated (PO) oligonucleotides of increased WBC and reduced RBC (21), were not noted with Dz13 administration, while there was a tendency for reduced platelet counts, though this was not significant.
In a further sensitive test, when pregnant mice were administered Dz13 continuously and systemically with the aid of osmotic pumps, the fetus of Balb/C pregnant mice failed to exhibit signs of gross toxicity. Similar findings were reported for a PO antisense molecule in mice (Henry et al., 2004a) and rabbits (Henry et al., 2004b). In mice, exposure to the fetus is expected to be minimal, but until now, this had not been directly evaluated for a 3′-inverted T oligonucleotide.
Dz13 effects on tumor cell angiogenesis (Zhang et al., 2004) and apoptosis (Dass et al., 2008a, 2008b, 2008c, 2008d) have been reported. Its effects on physiological processes of angiogenesis and apoptosis have never been examined; so to address this, 3 tissues were analyzed in the in utero assay. These included fetal bone, placenta, and skin wound. No evidence of perturbed angiogenesis was seen. These results somewhat contradict the in vitro findings in HMEC-1 cells (Rivory et al., 2006; Goodchild et al., 2007; Gozar et al., 2008), where reproducible cell death was shown in this cell line in all 3 studies by the same group. This can be attributed to the fact that in vivo, other major factors are in play in a tissue that is rapidly undergoing angiogenesis, and in this scenario, Dz13 is not able to cause cell death to endothelial cells, even though they are rapidly dividing and highly activated. In addition, the effect of the SV40 transformation on the originally derived (primary) HMEC cells may render these cells more susceptible to such insults. A study with primary HUVEC cells would have addressed this in the earlier studies. Apoptosis is vital to proper tissue modeling in the young including the removal of interdigital web tissue in developing mouse paws (Jacobsen et al., 1996). No sign of derailed apoptosis was noted in the limbs of unborn mice.
In adult mice, several tissues were assessed, including bone, skeletal muscle, liver, spleen, lung, and kidney. Only in the liver was toxicity noted, mainly as cell death, most probably via necrosis, though this was not widespread and hepatomegaly, as noted with other oligonucleotides (Henry et al., 2004a, 2004b) was absent. Furthermore, there was a mild toxic effect with the scrambled oligonucleotide as well. Nevertheless, because of the hepatotoxicity, a systemic route of administration was not chosen for further studies. Hepatocyte necrosis has been noted previously with PO oligonucleotides administered intravenously at 30 mg/kg/day over 14 days (Agrawal et al., 1997). Interestingly, in that study ALP levels were perturbed and renal tubular structure was affected, but not in the present study.
Conclusion
Summarily, this article establishes that Dz13 is capable of down-regulating c-Jun in human cancer cells. However, c-Jun down-regulation is not the cause for the cell growth inhibitory effects seen. For Dz13, certain motifs within the oligonucleotide sequence endow it with its cell inhibitory ability. In vivo, in general, no toxic effects were observed in a wound healing assay, an in utero assay, and adult mice bolus and continuous systemic dosing studies. A shorter version of Dz13 was identified that could make potential therapy in future more economical and practical, provided it passes the preclinical tests that Dz13 has undergone and is currently undergoing.
Footnotes
Acknowledgments
We thank Maya Kansara for assistance with hematology and Levon Khachigian for Dz13 and Scr. This study was supported by funds from St. Vincent's Hospital Melbourne (Orthopaedics Department funds). The authors wish to dedicate this article to the memory of the late Rod Walsh, part of our research support team at SVH.
Author Disclosure Statement
None declared by the authors.