
Abstract
MicroRNAs (miRNAs) are noncoding RNAs approximately 22 nucleotides in length that play a major role in the regulation of important biological processes, including cellular development, differentiation, and apoptosis. Antisense oligonucleotides against miRNAs are useful tools for studying the biological mechanisms and therapeutic targets of miRNAs. Various antisense oligonucleotides chemistries, including peptide nucleic acids (PNAs), have been developed to enhance nuclease-resistance and affinity and specificity for miRNA targets. PNAs have a greater specificity and affinity for DNA and RNA than do natural nucleic acids, and they are resistant to nucleases—an essential property of an miRNA inhibitor that will be exposed to cellular nucleases. However, the main limiting factor in the use of PNAs is their reduced penetration into cells. Recently, several cell-penetrating peptides (CPPs) have been investigated as a means to overcome the limited penetration of PNAs. Here, we evaluated the ability of 11 CPPs to transport PNAs inside cells in the absence of transfection reagents and then investigated the ability of these CPPs to inhibit miRNAs. Of the 11 CPPs tested, Tat-modified-conjugated PNA showed the most effective penetration into cells in the absence of transfection reagents and most effectively inhibited miRNAs. Our data demonstrate that Tat-modified-conjugated CPP is the most suitable for supporting PNA-mediated miRNA inhibition.
Introduction
PNAs are DNA mimics consisting of an N-(2-aminoethyl)-glycine backbone that has a neutral character (NIELSEN, 1991; Buchardt et al., 1993). They are resistant to degradation by nucleases and bind to complementary DNA or RNA sequences with high affinity and high specificity. Given these advantages, PNAs have strong potential as antisense and gene-targeting drugs; however, there are certain limits to their application. In particular, PNAs have low solubility in aqueous solutions and, because of their neutral charge, do not easily enter cells. Additionally, PNAs do not interact with transfection reagents, such as cationic lipids. Several delivery methods have been suggested for overcoming these limitations, including electroporation, microinjection, and cotransfection with DNA (Koppelhus and Nielsen, 2003). Alternatively, PNAs can be modified by conjugation with specific materials to enhance intracellular delivery (Rasmussen et al., 2006; Shiraishi et al., 2008; Abes et al., 2009). Several modifications of PNAs that increase their association with cationic lipid complexes have been suggested, including conjugation with lipophilic ligands (eg, fatty acids) that introduce a negative charge in the backbone, or the use of PNA–DNA heteroduplexes or PNA–DNA chimeras (Shiraishi et al., 2008). In addition, several peptides, known as cell-penetrating peptides (CPPs) or protein-transduction domains, have been shown to have strong cell-penetrating properties that can be conferred to conjugated cargo molecules (Turner et al., 2005; Bøe and Hovig, 2006; Abes et al., 2007; Foged and Nielsen, 2008; Mae et al., 2009). Conventional antisense oligonucleotides, such as phosphorothioate- or 2′OMe-modified forms, require transfection reagents for intracellular delivery. However, cationic lipid transfection reagents are cytotoxic to sensitive cell types, such as primary cells and stem cells (Mae et al., 2009). For such applications, it is desirable to eliminate the cytotoxicity of transfection reagents. Recently, Stein et al. reported the development of a method for promoting intracellular penetration of oligonucleotides that does not require the use of transfection reagents. Here, we analyzed numerous CPPs for their ability to intracellularly deliver PNAs without the use of transfection reagents and support the function of these PNAs as miRNA inhibitors. Using flow cytometry to monitor internalization of fluorescently labeled CPP-conjugated PNAs, we show that CPP-conjugated PNAs effectively entered cells in the absence of transfection reagents. The inhibitory effects of CPP-conjugated PNAs on miRNAs were confirmed by luciferase assays and real-time reverse transcription–polymerase chain reaction (RT-PCR).
Materials and Methods
Design and synthesis of anti-miRNA PNA antisense oligonucleotides
Human miRNA sequences were obtained from the miRBase Sequence Database (http://microrna.sanger.ac.uk, Release 10.1). The sequences of anti-miRNA PNAs used in this study were as follows: PNA21, N-tcaacatcagtctgataagcta-C; PNA24, N-ctgttcctgctgaac-C; PNA30b, N-gtgtaggatgtttac-C; PNA31, N-tgccagcatcttgcc-C; PNA181a, N-gacagcgttgaatgt-C; PNA222, N-acccagtagccagat-C. PNAs, synthesized as previously described (Lee et al., 2007), contained an O-linker at the N-terminus to improve solubility and were conjugated to a modified peptide at the N-terminus for effective transfection into cells. The following 11 different CPPs were tested: Pen, RQIKIWFQNRRMKWKK-PNA; Tat, GRKKRRQRRRPPQ-PNA; 47Tat57, GGGGYGRKKRRQRRR-PNA; Cationic, KKKK-PNA; C-myc tag, EQKLISEEDLNA-PNA; H region, AAVALLPAVLLALLA-PNA; PTD-4, YARAAARQARA-PNA; Tp-10, AGYLLGKINLKALAALAKKIL-PNA; SSBP(I), PKKKRKV-PNA; Tat-modified, RRRQRRKKR-PNA; and Lys, K-PNA-KKK (Koning et al., 2003; Abes et al., 2007; Fabani and Gait, 2008).
Cell culture and transfection
HeLa, A549, and MCF7 cells were maintained in Dulbecco's modified Eagle's medium or Roswell Park Memorial Institute medium (RPMI) supplemented with 10% fetal bovine serum, penicillin, and streptomycin. Before experiments, cells were plated in either 24- or 6-well plates and maintained in antibiotics-free medium for 24 hours. For lipid-based transfections, PNAs and target gene plasmids were mixed with Lipofectamine 2000 (Invitrogen) and then added to cells. After incubating for 48 hours, cells were harvested, and luciferase activity in cell lysates was assessed. For delivery of PNAs in the absence of a transfection reagent, PNAs were mixed with Opti-MEM (Invitrogen) and then incubated for 15 minutes at room temperature. PNA-Opti-MEM mixtures were directly added to cells.
Luciferase assay
Luciferase reporters were constructed by subcloning nucleotides complementary to the miRNA into the 3′-UTR region of the firefly luciferase gene in the pGL3 control vector (Promega). The miRNA-complementary nucleotide sequences were as follows: hsa-miR-21, 5′-tcaacatcagtctgataagcta-3′; hsa-miR-24, 5′-ctgttcctgctgaactgagcca-3′; hsa-miR-30b, 5′-agctgagtgtaggatgtttaca-3′; hsa-miR-31, 5′-agctatgccagcatcttgcct-3′; hsa-miR-181a, 5′-actcaccgacagcgttgaatgtt-3′; and hsa-miR-222, 5′-acccagtagccagatgtagct-3′. Cells were cotransfected with PNAs and both firefly luciferase reporters and renilla luciferase expression constructs (Promega). After 48 hours, firefly luciferase activity was measured in cell lysates and normalized to transfection efficiency, measured by renilla luciferase enzyme activity (Dual Luciferase Assay, Promega).
Fluorescence-activated cell sorting analysis
PNAs conjugated with N-terminal fluorescein isothiocyanate (FITC)-labeled CPP were incubated with HeLa cells in the absence of a transfection reagent. After 48 hours, the cells were washed with phosphate-buffered saline and harvested using trypsin-ethylenediaminetetraacetic acid. The cells were suspensed with cold phosphate-buffered saline containing 10% fetal bovine serum. Cellular internalization of FITC-labeled, CPP-conjugated PNAs was analyzed using a FACSCalibur (Becton Dickinson) flow cytometer with detection at excitation and emission wavelengths of 488 and 575 nm, respectively. Fluorescence was detected using the FL1-H channel, and data were analyzed using FACSCalibur 110VAC (Becton Dickinson).
Real-time RT-PCR analysis
CPP-conjugated PNAs were incubated with cells in the absence of transfection reagents. After 48 hours, miRNAs were extracted using a PureLink miRNA Isolation Kit (Invitrogen), as described by the manufacturer. cDNA was synthesized by RT-PCR using 10 ng of miRNA and 3 μl of RT primer in a total reaction volume of 12 μl, as described by the manufacturer of the RT-PCR kit (Applied Biosystems). MiRNA levels were quantified using a TaqMan MicroRNA Assay Kit (Applied Biosystems) and a CFX96 Real-Time PCR System (Bio-Rad).
Results and Discussion
Screening of CPPs for anti-miRNA efficacy of conjugated PNAs
Previous studies have investigated the use of CPPs to increase cellular penetration of PNAs (Geoffrey et al., 2001; Hallbrink et al., 2004; Barany-Wallje et al., 2007; Awasthi and Nielsen, 2008; Fabani and Gait, 2008; Lundin et al., 2008; Yoo et al., 2009). To determine which CPP was most appropriate for efficient intracellular delivery of anti-miRNA PNAs, we first tested the anti-miRNA effects of PNAs conjugated to 11 different CPPs. To accomplish this, we cotransfected HeLa cells with a luciferase reporter construct for the target gene of miRNA-24 and CPP-conjugated PNAs using the transfection reagent Lipofectamine 2000. Assays of cell lysates for luciferase activity performed 48 hours later showed that all CPPs tested, except Cationic, C-myc tag, H region, and PTD-4, increased cellular uptake compared with naked PNA (Fig. 1). Of these, the most effective miRNA inhibition was obtained using Tat-modified-conjugated PNA.

Assessment of miRNA-inhibitory effects of CPP-conjugated PNAs. HeLa cells were transfected with 200 nM of each CPP-conjugated PNA24 using Lipofectamine 2000. After 48 hours, cell lysates were analyzed by luciferase assay. No CPP: naked PNA. miRNA, microRNA; CPP, cell-penetrating peptide; PNA, peptide nucleic acid.
Uptake of CPP-conjugated PNAs in the absence of a transfection reagent
To confirm penetration of PNAs, we N-terminally labeled each of the 11 types of CPP-conjugated PNAs with FITC. HeLa cells were incubated with the fluorescence-labeled CPP-conjugated PNAs in the absence of transfection reagents. After 48 hours, cells were harvested, and cellular uptake of fluorescently labeled CPP-PNA conjugates was measured by fluorescence-activated cell sorting (FACS) analysis (Fig. 2). The most effective CPP was Tat-modified, which increased the mean value of fluorescence by approximately 10.9-fold compared with naked PNA (51.06 vs. 4.69). The second-most effective CPP for promoting cell penetration was 47Tat57. In fact, cell penetration of 47Tat57-conjugated PNA was similar to that of Tat-modified-conjugated PNA. However, results of luciferase assays showed that Tat-modified-conjugated PNA was more effective in inhibiting miRNA than was 47Tat57-conjugated PNA. Consistent with the results of lipofectamine-mediated transfection experiments, FACS data showed that the localization of FITC fluorescence for the H region-conjugated PNA was similar to that of control, although the mean value of fluorescence was elevated (39.96). One possible explanation could be that the H region-conjugated PNA did not evenly penetrate cells. The Tat-conjugated PNA also effectively mediated penetration into cells, but its miRNA inhibitory effect measured in luciferase reporter assays was less than that of Pen-conjugated PNA. Collectively, these data show that certain CPP-conjugated PNAs are effectively transported into cells in the absence of a transfection reagent that will be able to cause cytotoxicity and are capable of inhibiting miRNA.

Fluorescence-activated cell sorting analysis of cell penetration by fluorescently labeled CPP-conjugated PNAs. HeLa cells were incubated with 1 μM of each CPP-conjugated PNA24 in the absence of a transfection reagent. After 48 hours, cells were harvested and analyzed by fluorescence-activated cell sorting. No CPP: naked PNA; black peaks: control cells; gray peaks: PNA-transfected cells.
Real-time RT-PCR analysis of the inhibitory effect of CPP-conjugated PNAs
To confirm the miRNA-inhibitory function of intracellular PNAs, we cultured CPP-conjugated anti-miRNA PNAs with HeLa cells in the absence of a transfection reagent and then assessed the expression of miRNA-24 using quantitative real-time RT-PCR. These analyses showed that Pen-, Tat-, 47Tat57-, Tp-10-, and Tat-modified-conjugated PNAs inhibited miRNA expression more effectively than naked PNA (Fig. 3). These results correspond well with FACS analyses (Fig. 2) and confirm that these CPPs promote cellular uptake of PNAs in the absence of a transfection reagent. Real-time RT-PCR analyses showed that Tp-10- and Tat-modified-conjugated PNAs exerted similar miRNA-inhibitory effects. However, luciferase assays and FACS data showed that Tat-modified is a better CPP for PNA than Tp-10.
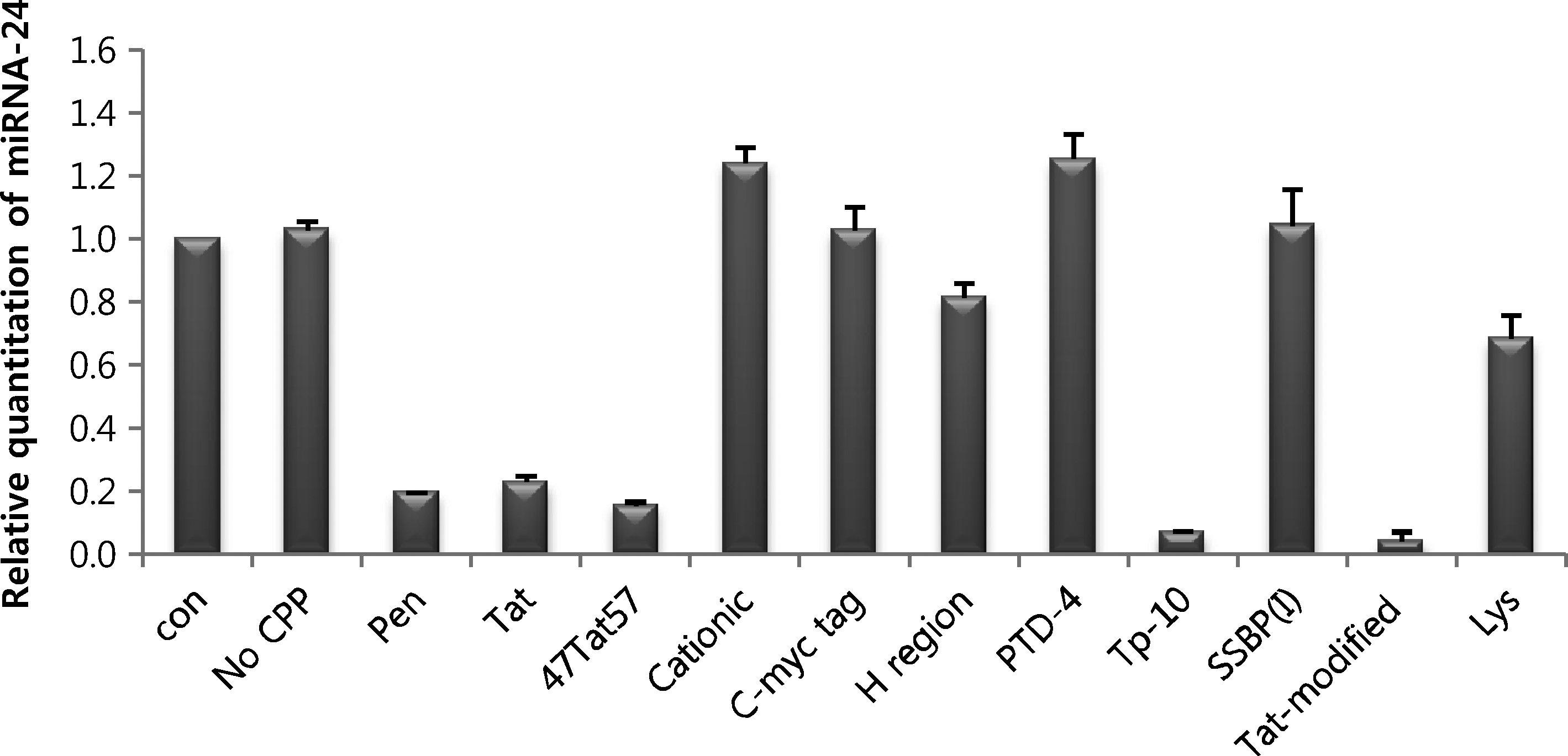
Assessment of miRNA-inhibitory effects of CPP-conjugated PNAs. HeLa cells were cultured with 1 μM of each CPP-conjugated PNA24 in the absence of a transfection reagent. After 48 hours, cell lysates were analyzed by real-time RT-PCR. Con: no treatment; No CPP: naked PNA. RT-PCR, reverse transcription–polymerase chain reaction.
Cell specificity of CPP-conjugated PNAs
The CPPs tested here have been recently used to increase PNA uptake into cells (Geoffrey et al., 2001; Zaffaroni et al., 2003; Hallbrink et al., 2004; Barany-Wallje et al., 2007; Awasthi and Nielsen, 2008; Lundin et al., 2008; Yoo et al., 2009). One recent study showed that a high concentration of KKKK-conjugated PNA directly inhibits miRNA expression (Fabani and Gait, 2008), and Tat-modified-conjugated CPP has been shown to be a useful tool for target-directed anticancer therapeutic interventions (Zaffaroni et al., 2003). Here, we found that the miRNA-inhibitory effectiveness of lysine-conjugated PNAs was lower than that of Pen-, Tp-10-, and Tat-modified-conjugated PNAs.
On the basis of the results described above, we selected the highly cationic CPPs, Pen and Tat- modified, and the amphipathic CPP, Tp-10 for tests of cellular specificity. To accomplish this, we first treated HeLa, MCF7, and A549 cell lines with these CPP-conjugated PNAs in the absence of a transfection reagent, and then we analyzed miRNA expression by real-time RT-PCR. Figure 4 shows that miRNA-24 expression was more effectively inhibited by each of these 3 CPP-conjugated PNAs than by naked PNAs in all tested cell lines. Consistent with the results described above, the Tat-modified-conjugated PNA was more effective than the other CPPs for PNA in these cell lines. Subsequent experiments confirmed that in each of 15 cultured cell lines tested, including stem cells, Tat-modified-conjugated PNAs inhibited miRNA (data not shown). Accordingly, we propose that Tat-modified is the most appropriate CPP for promoting cell penetration of the tested PNAs. Importantly, this CPP promoted effective transcellular transport of PNAs in the absence of a transfection reagent, thereby eliminating the cytotoxicity associated with these reagents and facilitating miRNA functional analyses.

Analysis of miRNA-24 expression in cells treated with CPP-conjugated PNAs. Each cell type was cultured with CPP-conjugated PNA24 in the absence of a transfection reagent. After 48 hours, miRNA was extracted, cDNA was synthesized, and miRNA was analyzed by real-time RT-PCR. No CPP: naked PNA.
Optimal concentration of Tat-modified-conjugated PNAs for miRNA inhibition in the absence of a transfection reagent
In the experiments described above, we identified Tat-modified as the most effective CPP for PNAs. To determine the optimally effective concentration of Tat-modified-conjugated PNA, we treated cultured cells with 10–1000 nM of different Tat-modified-conjugated PNAs in the absence of a transfection reagent and analyzed miRNA inhibition by real-time RT-PCR (Fig. 5). In the case of Tat-modified-conjugated PNA21 and PNA24, miRNA inhibition was detected at concentrations as low as 200 nM and 100 nM, respectively. However, the optimal concentrations of Tat-modified-conjugated PNA181a and PNA222 were 1000 nM and 500 nM, respectively, indicating that the appropriate concentration of Tat-modified-conjugated PNA for miRNA inhibition may differ depending on the target miRNA. We speculate that marked differences in the anti-miRNA effects of PNAs may reflect differences in the kinetics of each PNA.
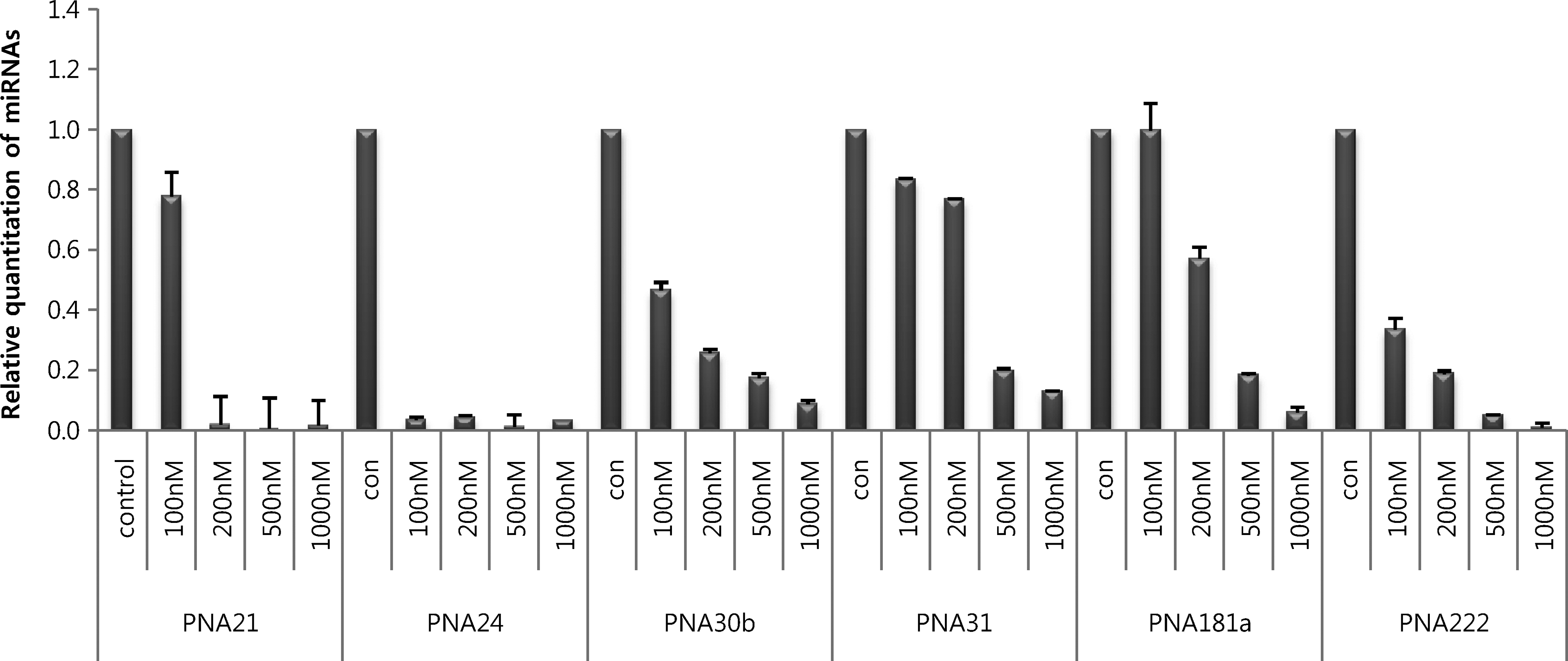
Optimization of Tat-modified-conjugated PNA concentration in HeLa cells. HeLa cells were cultured with 100–1000 nM of Tat-modified-conjugated PNAs in the absence of a transfection reagent. After 48 hours, miRNA was extracted, cDNA was synthesized, and miRNA was analyzed by real-time RT-PCR. Con: no treatment.
Importantly, no cytotoxicity was observed in cells treated with up to 4 μM CPP-conjugated PNA (data not shown). Our results demonstrate that Tat-modified is an effective and appropriate CPP for anti-miRNA PNAs, and CPP-conjugated PNA exhibits different optimal concentrations for each target miRNA.
Footnotes
Acknowledgments
This work was supported by Small and Medium Business Administration grant funded by the Korean Government (S1037791).
Author Disclosure Statement
No competing financial interests exist.