
Abstract
RNA and DNA aptamers that bind to target molecules with high specificity and affinity have been a focus of diagnostics and therapeutic research. These aptamers are obtained by SELEX often requiring many rounds of selection and amplification. Recently, we have shown the efficient binding and elution of RNA aptamers against target proteins using a microfluidic chip that incorporates 5 sol–gel binding droplets within which specific target proteins are imbedded. Here, we demonstrate that our microfluidic chip in a SELEX experiment greatly improved selection efficiency of RNA aptamers to TATA-binding protein, reducing the number of selection cycles needed to produce high affinity aptamers by about 80%. Many aptamers were identical or homologous to those isolated previously by conventional filter-binding SELEX. The microfluidic chip SELEX is readily scalable using a sol–gel microarray-based target multiplexing. Additionally, we show that sol–gel embedded protein arrays can be used as a high-throughput assay for quantifying binding affinities of aptamers.
Introduction
Additional gains in the SELEX process might also be achieved by miniaturization using microfluidic technology. Microfluidics refers to systems that handle small volumes of liquid (10−9–10−18 L) using micrometer-sized channels. Resulting small volumes and reduced matrix surface offer high-speed partitioning by decreasing diffusion time and nonspecific binding to the partitioning matrix. Microfabrication allows integration of fluidic elements such as micropumps, microvalves, microheaters, and related devices, on a single chip, and controlling such devices by computers can provide efficient automation of these processes. For those reasons, microfluidics have been broadly utilized in the field of chemistry, biology, medicine, and engineering to analyze samples at high speed and with high throughput (WHITESIDES, 2006).
Recently, we demonstrated that microheaters positioned under nanoporous sol–gel droplets were effective in selectively eluting aptamers that were bound to specific proteins embedded in the sol–gel droplet (Kim et al., 2006; Park et al., 2009). A microfluidic device with this arrayed configuration of sol–gel droplets and microheaters, named “SELEX-on-a-chip,” can be used to perform parallel aptamer selections using small sample volumes and thereby minimizing consumption of expensive reagents. Sol–gel processes offer a relatively mild route for the immobilization of biomolecules such as proteins, which are entrapped in the growing covalent silica gel network rather than being chemically attached to an inorganic material (Gill and Ballesteros, 1996, 2000). The sol–gel process involves the evolution of inorganic networks through the formation of a colloidal suspension (sol) and gelation of the sol to form a nanocomposite network in a continuous liquid phase (gel). At the functional group level, 3 reactions are generally used to describe the sol–gel process: hydrolysis, alcohol condensation, and water condensation. However, the characteristics and properties of a particular sol–gel network are related to a number of factors that affect the rate of hydrolysis and condensation reactions (COLOMBO, 2006; Dietze et al., 2006). Many studies have described the encapsulation of a variety of biomolecules, including enzymes, antibodies, regulatory proteins, membrane-bound receptors, nucleic acid aptamers, and even whole cells, using a wide range of sol–gel derived nanocomposite materials (Reetz et al., 1996; Frenkel-Mullerad and Avnir, 2005). With regard to stability, proteins entrapped in sol–gels typically exhibit improved resistance to thermal and chemical denaturation, and increased storage and operational stability over months or even longer (Kim et al., 2005; Pastor et al., 2007). Additionally, the nanoporous structure of the sol–gel matrix can allow diffusion of some molecules, such as aptamers, but keeps target molecules (protein or chemicals) immobilized in the pores. Sol–gel has been optimized by screening from over 100,000 formulations for a formulation that provides optimal retention of targets of various sizes, optimal access of probes, and low assay backgrounds (Kim et al., 2006). That can be explained by the experimental results that sol–gel droplet can hold proteins inside a dual nanoporous structure–small pores for immobilizing proteins and bigger pores or channels as molecular passages to bind specifically to entrapped proteins inside. Lee et al. have used sol–gel-based chip material for detecting antigen–antibody or antigen–aptamer interactions with higher sensitivity than other currently available protein chip materials (Lee et al., 2007; Ahn et al., 2008; Kwon et al., 2008).
Here, using this sol–gel-based microfluidic SELEX-on-a-chip, several aptamers for TATA-binding protein (TBP) (Yokomori et al., 1994) were selected, including aptamers previously selected by conventional SELEX (Shi et al., 2007) as well as new high-affinity aptamers, demonstrating the general utility of the system as a high-throughput SELEX protocols (Fig. 1). Our microfluidic SELEX system greatly improved selection efficiency, reducing the number of selection cycles needed to produce high-affinity aptamers by 5–8 rounds, and producing high-affinity aptamers to TBP that were identical or homologous to those isolated previously by conventional filter-binding SELEX (Fan et al., 2004). Moreover, this process is readily scalable to larger arrays of sol–gel-embedded proteins.

Experimental plan for comparing microfluidic to conventional SELEX. TBP aptamers isolated by microfluidic SELEX on-a-chip were compared with aptamers from 11th round of conventional filter binding SELEX. The microfluidic SELEX was performed after 2 rounds of conventional SELEX. TBP aptamers from micro-fluidic cycles ms 3, ms 4, ms 5, and ms 6 were sequenced. TBP aptamer sequences between from the microfluidic each round and from the 11th round of conventional SELEX were compared and classified into 2 groups (matched and newly selected aptamers). TBP, TATA-binding protein.
Materials and Methods
Protein preparation
Full-length His-tagged versions of yeast TBP (yTBP), transcription factor-II B (yTFIIB), and human heat-shock factor (hHSF) were all purified from BL21-DE3 cells according to a standard His-tagged protein purification protocol (Fan et al., 2004; Zhao et al., 2006; Sevilimedu et al., 2008). In the case of yeast transcription factor-II A (yTFIIA), recombinant proteins were purified by using a protocol in which subunits Toa1 and Toa2 were expressed separately in Escherichia coli, denatured in 8 M urea, combined, and renatured by dialyzing out the urea (Hahn et al., 1989). The purified target protein fractions were dialyzed following the methods provided by the manufacturer (Spectrum). To confirm the dialyzed protein remained intact, SDS-PAGE analysis was performed.
Microfluidic SELEX-on-a-chip preparation using sol–gel protein droplet immobilization
For on-chip aptamer selection, we used our microarray platform of nanoporous sol–gel droplets partitioned over microheaters as in our previous study (Park et al., 2009). Our chip consists of a glass slide with a set of aluminum electrodes and a polydimethylsiloxane (PDMS) lid (microchannel part) and sol–gel droplets (300 μm diameter and 20 μm height) connected by a microfluidic channel. As shown in Supplementary Fig. S1A (Supplementary Data are available online at www.liebertonline.com/oli), the microfluidic device consists of 5 hexagonal chambers with a side length of 1 mm and each chamber is connected by 6 microchannels with 170 μm deep and 8 mm length. The dimensions of final PDMS chip are 7.5 cm in length, 2.5 cm in width, and 1 mm in height.
Briefly, before bonding the PDMS lid with microheater part, the sol–gel droplets containing proteins were spotted onto the center of the patterned PMMA surface on top of Al-electrode microheater using a Stealth Solid pin (Digi Lab). Sol–gel materials and proteins in sol–gel were prepared according to the method described previously (Kim et al., 2006). After overnight gelation, the microheater part with sol–gels and the PDMS lid part were bonded under oxygen plasma treatment as described previously (Park et al., 2009).
In vitro selection of RNA aptamer using microfluidic SELEX chip device
Initial filter binding SELEX
Using starting aptamer pool (∼1015 different RNA molecules) as described previously (Fan et al., 2004; Shi et al., 2007), the first 2 cycles of selection and amplification were performed according to the conventional nitrocellulose filter binding assay (Fan et al., 2004; Sevilimedu et al., 2008). In detail, the random pool (420 pmol) was applied to 4 different proteins (42 pmol of TBP, TFIIA, TFIIB, and hHSF1) separately. Each RNA–protein mixture was incubated in 1 × binding buffer (12 mM HEPES pH 7.9, 100 mM NaCl, 10 mM MgCl2, and 1 mM DTT), partitioned using a nitrocellulose filter, and the bound RNA was recovered and amplified to yield an enriched pool for the next cycle.
Microfluidic SELEX on-a-chip
After the second cycle of filter binding SELEX, 4 consecutive cycles of in vitro selection and amplification were performed with the microfluidic SELEX platform. All reactions and washing procedures were performed in microfluidic chip using a syringe pump as described below. Five sol–gel droplets containing yTBP, yTFIIA, yTFIIB, and hHSF1 proteins were spotted evenly along the microfluidic channels (Supplementary Fig. S1B). Each sol–gel droplet can entrap ∼30 fmol protein, so that a total of 120 fmol (for 4 proteins) of the protein were immobilized in one microdevice. Around 12 pmol of RNA species was introduced to the single microfluidic device through the microchannel and chambers for on-chip SELEX in the direction from spot 4 to spot N.
Individual elution and recycling of selection of bound aptamers
After 1-hour incubation of selected RNA pool, the sol–gel droplets in the microfluidic chambers were washed with a 90 μL/minute flow of a binding buffer. Optimal electric powers (2 minutes, 1.5 V, 450 mW) using Keithley 2400 source meter (Keithley) were applied to the aluminum electrodes for heat elution, starting from hHSF1 droplet (N4) to TBP droplet (N1) (Supplementary Fig. S1B) to avoid unwanted heat effect in subsequent chambers. Each eluted RNA aptamer was retrieved, reverse-transcribed to cDNA, amplified, and then transcribed to RNA aptamer for the next-round selection. The reverse transcription reactions were carried out using reverse transcription kit (Invitrogen) according to the manufacturer's protocol. This cDNA was amplified by the polymerase chain reaction (PCR) step using 2 primers. The sequence of the forward primer is 5′-GTAATACGACTCACTATAGGGAGAATTCAACTGCCATCTAGGC-3′ and that of the reverse primer is 5′-ACCGAGTCCAGAAGCTTGTAGTACT-3′. Each PCR product was purified using QIAquick PCR Purification Kit (Qiagen) and then converted into RNA aptamers using a MEGAshortscript kit (Ambion). Equimolar RNA aptamers against yTBP, yTFIIA, yTFIIB, and hHSF1 were introduced into microfluidic chip for the following selection step.
Aptamer identification and sequence comparison between aptamers from conventional SELEX and microfluidic SELEX-on-a-chip
Eluted pools from every round of microfluidic SELEX were reverse transcribed to DNA. Amplified double-strand DNAs were cloned into the pGEM-T vector (Promega) and clones were all sequenced after selection cycles, ms 3, ms 4, ms 5, and ms 6, respectively (Solgent). The comparisons of DNA sequences were performed with Clustal X (Higgins and Sharp, 1988). All selected aptamers were compared with previously isolated TBP aptamers from conventional SELEX (TBP Apt No. 1, TBP Apt No. 13, and TBP Apt No. 17) (Fan et al., 2004).
Quantitative measurement of aptamer from each round
To understand the process of aptamer enrichment, the amount of single aptamer was measured from each round pool (Library, first, second, ms 3, ms 4, ms 5, and ms 6) using quantitative realtime PCR (RT-qPCR) experiment according to the manufacture's protocol (AB StepOnePlusTM Real-Time PCR System; Applied Biosystems) (Fig. 2). In detail, a specific primer set (specific forward primer 5′-GAGATCACGAAAAAGCGGAATTG-3′ and reverse primer 5′-ACCGAGTCCAGAAGCTTGTAGTACT- 3′) was designed against the selected aptamer, ms 3.3 (TBP Apt No. 1) (Fig. 2A for detailed sketch). For quantification of ms 3.3 aptamer in each round, the each eluate from the first and second round of filter binding SELEX, and ms 3, ms 4, ms 5, and ms 6 pool of microfluidic SELEX was used as templates for assays under the following conditions: 95°C for 10 minutes followed by 40 cycles of 95°C for 10 seconds and 60°C for 1 minutes. All experiments were done in triplicate. Best fit lines were generated by the StepOne™ software v2.1 (R2 > 0.928) (Applied Biosystems).
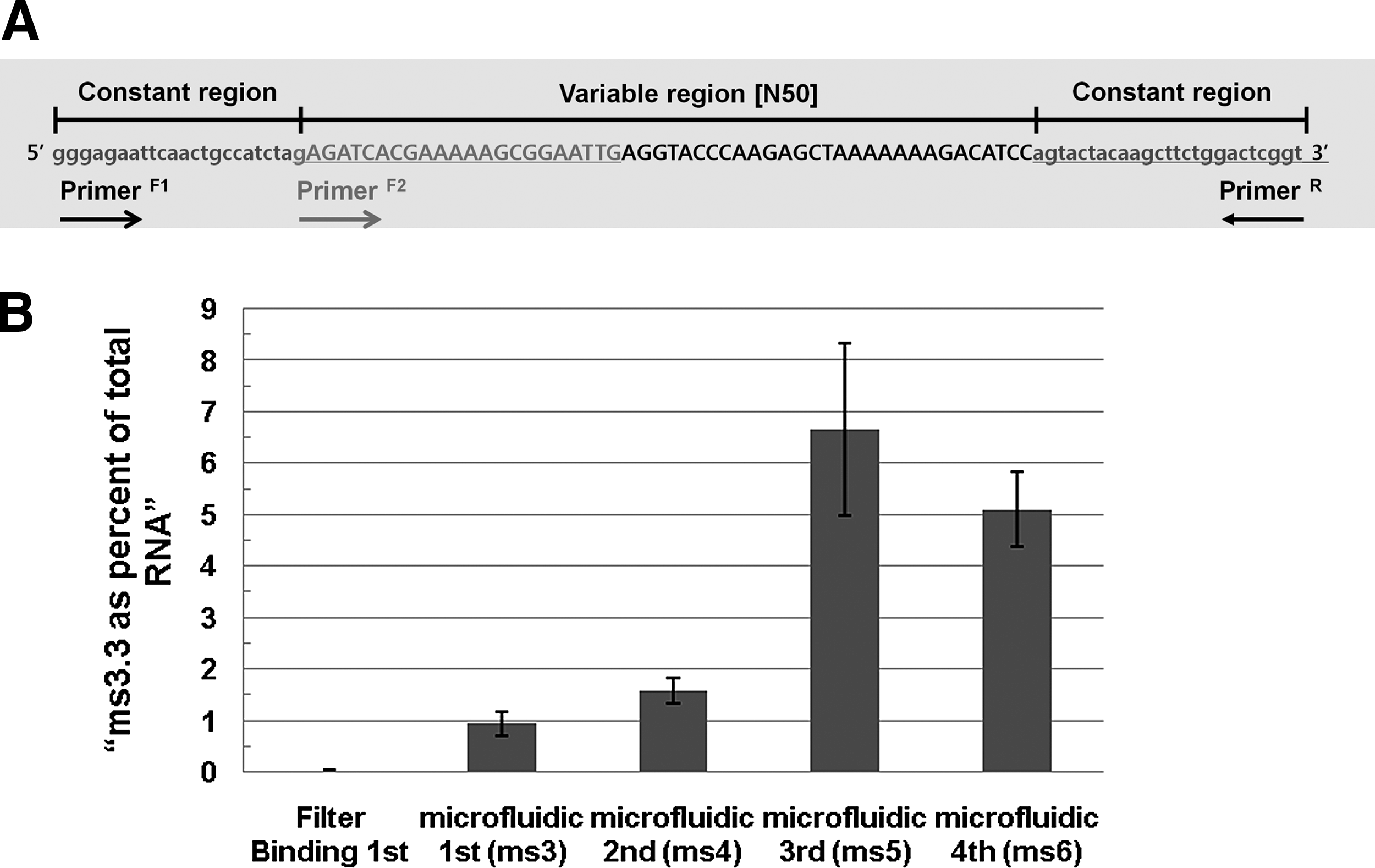
Monitoring the enrichment of aptamer during microfluidic SELEX. Specific forward primer was designed for ms 3.3 (TBP Apt No. 1) including a portion of 5′ constant sequence
Protein–aptamer binding assay using sol–gel-based array chip
RNA aptamer synthesis
The selected individual aptamers against TBP were transcribed using a MEGAshortscript kit (Ambion). Briefly, after PCR amplifying the aptamer template construct, 1 μg of the amplified DNA templates was in vitro transcribed to RNA according to the manufacturer's protocol. Finally, the samples were incubated at 37°C for 20 minutes in the presence of 1 μL of DNase I.
End labeling of RNA aptamer
The RNA aptamer was incubated for 4 hours at 37°C with 100 μM Cy3-dUTP (E-Biogen), 50 μM terminal deoxynucleotidyl transferase (TdT), (Fermentas), 1 × TdT buffer [200 mM potassium cacodylate, 25 mM Tris/HCl (pH 6.6), 4 μM bovine serum albumin, and 5 mM CoCl2], 0.5 mM deoxynucleotide triphosphate, and 10 U RNase inhibitor (Roche). The reaction was stopped by addition of EDTA. The labeled RNA was extracted by phenol/chloroform/isoamylalchol treatment recovered by ethanol precipitation in the presence of 0.3 M sodium acetate.
Binding of aptamer to TBP
Using sol–gel chip assay, binding of RNA pools or aptamers to TBP proteins were tested (Supplementary Fig. S3 and Fig. 3). Six duplicate TBP protein spots were printed along with negative controls (no protein) and reference controls (with Cy-3-labeled proteins) within 8–mm-diameter wells of 96-well plates (SPL). Spotting of sol–gel protein chip was done as described previously (Kim et al., 2006). After cleaning and blocking of wells, Cy-3-labeled RNA aptamers were incubated for 1 hour in each well and then washed 3 times for 15 minutes. The resulting plate chip well was scanned and analyzed using a 96-well fluorescence scanner and the appropriate software program (FLA-5100 and multigauge; Fuji). The background intensity was subtracted from the signal intensity of each spot (LAU/mm2).
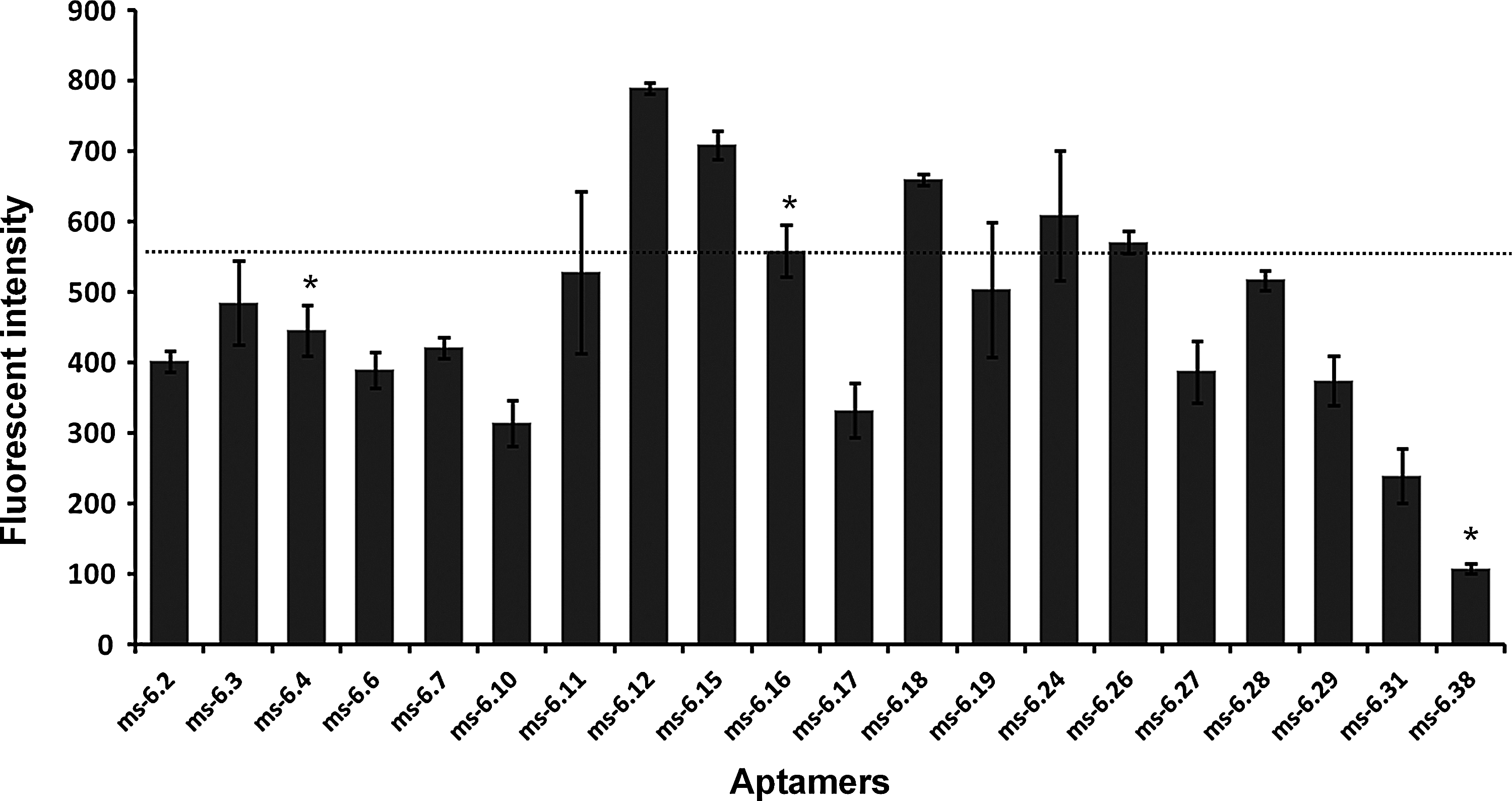
Aptamer binding results using the sol–gel-based array chip. Individual binding activity of selected aptamers was calculated and plotted as the fluorescent intensity of sol–gel spot as described in the Materials and Methods section. Each aptamer (200 pmol) was labeled with Cy-3 and separately incubated with sol–gel spots containing TBP for binding activity analysis. The images of binding assay are described in Supplementary Fig. S3. Aptamers ms 6.4, ms 6.16, and ms 6.38 matched previously selected aptamers (Fan et al., 2004) (see Table 1, Group I) and are marked with asterisk (*). All measured intensity was normalized by the value of RNA library as a control.
Results
Efficient selection strategy using microfluidic SELEX-on-a-chip
Previously, we demonstrated that specific aptamers bind their respective protein targets immobilized within a nanoporous sol–gel droplet and can be selectively eluted by micro-heating (Park et al., 2009). This sol–gel-based microfluidics system (SELEX-on-a-chip) was expected to provide rapid, and efficient selection of aptamers in a multiplex format for several reasons. First, because the microfluidic device allows the scale-down to very small quantities of reagents, the microscale creates favorable flow characteristics such as diffusion dominant chemical transport and laminar flow when fluid passes through the micro- to nano-sized channel. Thus, in comparison with conventional SELEX (Supplementary Table S1), resulting small volumes and reduced matrix surface of microfluidic SELEX offer high-speed partitioning of aptamer binding to targets by decreasing both diffusion time and nonspecific binding to the partitioning matrix. This can be explained by the fact that travel time is proportional to the length of the device squared as described by the equation:
where l is the length of device, tD is the time traveled, and D is the RNA diffusion coefficient (Weibel and WHITESIDES, 2006). Second, because the sol–gel system provides ultra-low volume handling, sol–gel as a 3D immobilizing material is ideal for a microfluidic system (Supplementary Fig. S1A). Additionally, immobilizing an array of different proteins in a single channel eliminates the need for a negative SELEX step which normally requires 3–5 additional selection cycles.
Overview of TBP aptamer selection using microfluidic SELEX on-a-chip
To show that the sol–gel-based microfluidic chip SELEX is more efficient than conventional SELEX in aptamer generation, we used yTBP as a first target, which was previously selected for aptamers using conventional filter binding SELEX (Fan et al., 2004). To evaluate the SELEX on-a-chip system, we performed yTBP aptamer selection using microfluidic SELEX and compared the results with conventional filter binding SELEX performed on the same protein (Fan et al., 2004) (Fig. 1). Only a total of 120 fmol of the protein could be immobilized in single microdevice, and this may not be sufficient for the first cycles of a SELEX experiment, where the initial complexity of random pool is extremely large (∼1015). The SELEX process in general uses protein to RNA ratio of at least 1:100. Therefore, we performed 2 initial rounds of conventional SELEX to reduce the size of the aptamer pool before starting 4 consecutive rounds of microfluidic SELEX.
In the case of conventional SELEX of yTBP, we obtained yTBP aptamers after 11 cycles of SELEX and several negative selection cycles (Fan et al., 2004). Here, the yTBP target protein was immobilized at the first position followed by other sol–gel droplets containing yTFIIA (position 2), yTFIIB (position 3), and hHSF1 (position 4) that served as competitors, because they are all factors that operate together on promoters and will be targets for future SELEX-on-a-chip experiments. This arrangement provides a negative selection with each cycle, and additional negative SELEX steps were not required to obtain the yTBP aptamers (Supplementary Fig. S1B).
Using the final selected pool from sixth round (ms 6), 38 individuals aptamers were obtained and sequenced. These individuals belong to 20 different sequences, which are listed in Table 1. Comparison of the sequences of aptamers isolated from this microfluidic SELEX with those previously selected by the conventional filter binding (Fan et al., 2004) shows that some of the same aptamers were selected by both protocols. The ms 6.16 and ms 6.38 matched perfectly previously isolated TBP Apt No. 17 and TBP Apt No. 1 respectively, and ms 6.4 had 98% homology with TBP Apt No. 13 in group I (Table 1). The sequence of aptamer ms 6.4, the most abundant sequence (8 of 38) from this microfluidic SELEX, only differs at one base from the sequence of aptamer TBP Apt No. 13, which was isolated in the previous study (Fan et al., 2004). In addition, newly isolated aptamer sequences were found as listed in group II. In conclusion, these results indicate that successful and efficient isolation of aptamers can be achieved using the microfluidic SELEX method.

Aptamers TBP Apt No. 1, 13, and 17 are the same one previously isolated from conventional filter binding method.
For our microfluidic SELEX experiments, “ms No.” represents an initial letter of Microfluidic SELEX and SELEX cycle.
We compared the aptamer sequences from microfluidic selection with previous conventional SELEX results (shaded sequences), and matched aptamers (marked as asterisks (*) characters) were listed in Group I and new (unmatched) aptamers were placed in Group II.
TBP, TATA-binding protein.
Microfluidic device can efficiently enrich for high affinity aptamers
To further investigate the enriching process of TBP aptamers in microfluidic SELEX on-a-chip device, we cloned and then sequenced members of each pool (ms 3, ms 4, ms 5, and ms 6) (Supplementary Table S2). Surprisingly, the aptamer, ms 3.3 (TBP Apt No. 1), was found among the 19 sequenced clones from the third cycle of selection (one round of microfluidic SELEX). This ms 3.3 aptamer is identical to TBP Apt No. 1 selected from conventional SELEX after 11 rounds of filter binding selection.
To obtain a more quantitative measure of the amounts of the ms 3.3 aptamer across the successive rounds of microfluidic SELEX, a specific primer for ms 3.3 and reverse primer for constant region of aptamer pool were designed for a quantitative real time PCR (RT-qPCR) experiment to monitor the enrichment progress of ms 3.3 aptamer (Fig. 2A). As shown in Fig. 2B, we can see the enrichment of ms 3.3 aptamer even after just one round of microfluidic SELEX. The maximum enrichment of this aptamer occurred in third round of microfluidic SELEX (ms 5), and this result is consistent with pool binding assay shown in Supplementary Fig. S3A. Based on these binding measurements of aptamer pools to TBP, the maximum enrichment of TBP-binding was also observed in the third round of microfluidic SELEX (ms 5).
To investigate further the binding activity of the isolated aptamers and to test whether sol–gel arrays can be used to conveniently assay relative binding affinities of specific aptamers, Cy-3-labeled aptamers were incubated with TBP spotted on sol–gel microarrays in wells of microtiter dishes. Individual binding activity was calculated by fluorescent intensity of sol–gel microdroplets (Supplementary Fig. S3B). As summarized in Fig. 3, some of the new ms 6 aptamers specifically bind TBP proteins better than those previously selected by the conventional filter binding. Then, for the binding affinity assay, 5 different concentrations of TBP (from 0 to 800 nM) within sol–gel mixture were prepared and arrayed on the surface of microwell plate. This sol–gel microarray assay showed that the ms 6.16 (TBP Apt No. 17) appeared to have affinity in the nanomolar range, 8.38 nM, agreeing with previous measurements by electrophoretic mobility shift assay (EMSA) and filter binding assays (Fan et al., 2004). We should point out that ms 6.16 (TBP Apt No. 17) only bound to TBP, not to other proteins immobilized (Supplementary Fig. S3C). This ms 6.16 aptamer was previously shown to strongly inhibit in vitro transcription at a concentration of only 40 nM, which is only twice that of TBP in these reactions (Fan et al., 2004).
Selected aptamers from microfluidics SELEX
From the analytical binding assay described in Fig. 3, we selected the 6 ms aptamers (ms 6.12, 6.15, 6.16, 6.18, 6.24, and 6.26) that showed the highest binding activity. Secondary structure models of aptamers were predicted with the Mfold program (http://mfold.bioinfo.rpi.edu/cgi-bin/rna-form1-2.3.cgi) (ZUKER, 2003). The most stable predicted folds are shown in Fig. 4. No apparent common sequence or secondary structure was observed among the 6 aptamers. The dissociation constants (Kd) of the all aptamers were found to be in the nanomolar range [Kd for ms 6.12, 2.7 nM; ms 6.15, 13.2 nM; ms 6.16, 8.3 nM; ms 6.18, 4.5 nM; ms 6.24, 92.53 nM; ms 6.26, 10.56 nM]. Aptamer ms 6.12, newly found from microfluidic SELEX, showed highest affinity (Kd ∼ 2.7 nM). In the case of aptamer No. 17 (ms 6.16), the binding affinity previously measured by EMSA was in the range from 3 to 10 nM (Fan et al., 2004) and ms 6.16 showed an ∼ 8 nM Kd from our sol–gel chip assay.
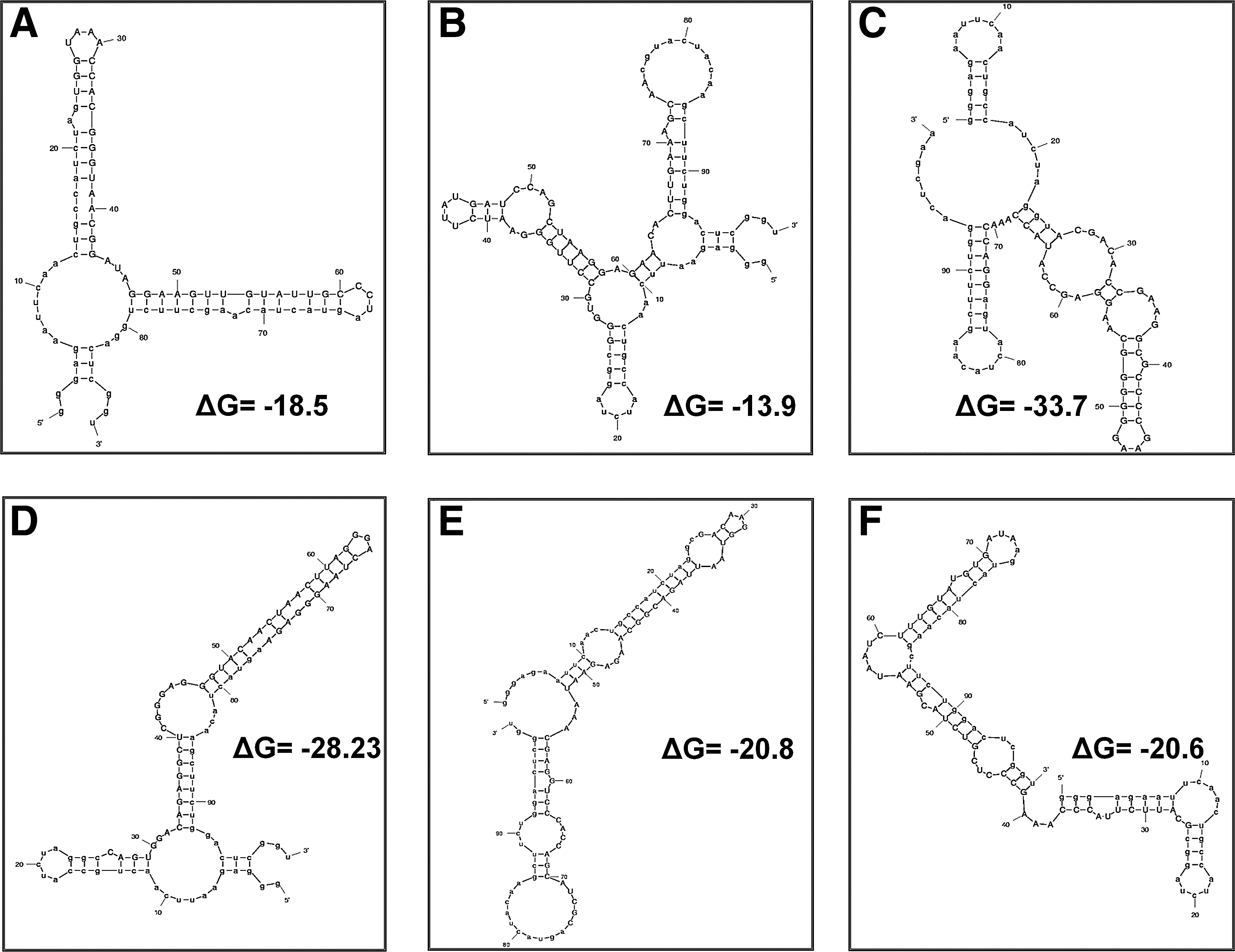
Mfold-generated secondary structure predictions.
Discussion
The SELEX process has been automated previously with the development of macro-robotic systems consisting of a PCR machine and a robotic manipulator to move reagents to multiple workstations. In addition to platform development, SELEX devices are being miniaturized. Hybarger et al. (2006) have reported the automated microline/valves based “start to finish” SELEX device. Even after these efforts, SELEX procedures are still usually focused on one or a few targets (Sefah et al., 2009).
A primary goal of most SELEX experiments is to obtain specific aptamers that bind to particular protein domains or to an active site of an enzyme. However, because target biomolecules are labile to denaturation by heat or solvents, target stability is an important issue in the SELEX experiment. Sol–gel technique allows the immobilization of target molecules in a biologically active form, and it presents these molecules to the aptamers at high density. It also produces a low background of nonspecific interaction (Kim et al., 2006).
Here, we described a strategy for selecting aptamers and demonstrate for the first time a complete selection protocol that uses a sol–gel derived microfluidic SELEX-on-a-chip with a device that was previously developed (Park et al., 2009). We focused on the aptamer selection of TBP, which is component of the RNA polymerase II general transcription factor machinery. yTBP was immobilized in a sol–gel droplet in a channel array along with sol–gel droplets containing yTFIIA, yTFIIB, or hHSF1 proteins as competitors for this micro-fluidic SELEX. The match between TBP aptamers selected by conventional and microfluidic SELEX demonstrated the effectiveness of using a microfluidic device to perform in vitro selection of aptamers against proteins. In this TBP aptamer selection, the microfluidic SELEX improved the efficiency of selection, reducing the number of cycles by 5 relative to a conventional binding assay that required 11 positive, as well as 3 additional negative cycles to obtain a high affinity TBP aptamer pool (Fan et al., 2004). Further, we can detect enrichment of an specific aptamer even after the third cycle of SELEX (the first cycle of microfluidic SELEX) even without negative selection cycles. Theoretically, a large array of proteins can be immobilized in sol–gel droplet arrays on a single glass slide. The different proteins in an array should serve each other as competitors in these selections of particular aptamers. Such competition may produce high-affinity aptamers with even fewer cycles of selection.
In principle, the binding reaction times can be reduced given that the diffusion time of aptamer to target molecules must be much shorter than milliseconds, in the chamber space of this device, because the transport of molecules by diffusion is proportional to the inverse square of the size of the chamber (Supplementary Table S1). Thus, the SELEX process in our microfluidic chip can be performed much faster than that of the existing conventional SELEX. Moreover, smaller-sized devices make it possible to build a highly integrated chip system for realizing the high-throughput SELEX to obtain aptamers against a large number of targets at once.
Footnotes
Acknowledgments
The authors acknowledge funding from National Research Laboratory from NRF (National Research Foundation) and Ministry of Knowledge Economy, and Industrial Technology Development (10032113-2010-13). S.K. would like to thank Korea Ministry of Environment as “The Eco-technopia 21 project” (2010-10002-0065-0/2010-09001-0076-0). J.-Y.A. would like to acknowledge the support of the National Research Foundation Grant (KRF-2008-532-D00003/2009-353-D00004). M.J. acknowledges the support from Korea Evaluation Institute of Industrial Technology (S1072576) and KIAT. D.-K.L. acknowledges the support from Global Research Laboratory grant by Korean Ministry of Education, Science and Technology. This work was supported by R&D Program of MKE/KEIT (10035501).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.