
Abstract
Hepatitis B virus (HBV) infection is a major health concern worldwide and only a minority of treated patients develop a sustained protective response following a short course of therapy, and most patients require prolonged treatment to suppress viral replication. However, several recent reports showed that inhibition of certain host cell proteins prevented viral infection, specifically the human abhydrolase domain containing 2 (ABHD2) has been confirmed by our previous study to be upregulated in HepG2.2.15 cells but downregulated by lamivudine. These observations suggested that ABHD2 was important for HBV propagation and could be a target of novel anti-HBV drugs. To assess the importance of ABHD2 to the HBV infection process, antisense oligonucleotides (ASODNs) were used to downregulate ABHD2 expression in HepG2.2.15 cells. From 5 ASODNS candidates tested, AB3 significantly downregulated ABHD2 mRNA and protein expression levels. Further, AB3 significantly reduced HBV DNA, hepatitis B surface antigen, and hepatitis B “e” antigen protein expression levels in cell medium without affecting cell viability. These results suggest that downregulation of ABHD2 using ASODNs blocked HBV replication and expression without affecting host cell physiology. Further, data demonstrated an essential role of ABHD2 in HBV propagation, suggesting it can serve as a novel target for anti-HBV drug development.
Introduction
Human abhydrolase domain containing 2 (ABHD2), an α/β hydrolase protein family member, possesses hydratase catalytic activity (HOLMQUIST, 2000). ABHD family genes were identified during a gene screen of human emphysematous tissues (Edgar and Polak, 2002) and ABHD2 was demonstrated to play a critical role in maintaining lung structural integrity by affecting lung homeostasis (Jin et al., 2009). It was later demonstrated that ABHD2 was involved in smooth muscle cell migration and in neointimal thickening of vascular smooth muscle cells (Miyata et al., 2005). However, no reports to date have described the relationship between HBV infection and ABHD2. Previous studies from our laboratory demonstrated that ABHD2 expression was upregulated in HepG2.2.15 cells but downregulated by lamivudine (Ding et al., 2008); however, as up- or downregulated genes may play different roles in facilitating viral replication, pathogenesis, or cellular responses associated with viral clearance, it was unclear what role upregulation of ABHD2 might play in the context of controlling viral replication. In this study, antisense oligonucleotides (ASODNs) were used to down-regulate ABHD2 expression as a means of assessing whether ABHD2 was essential to HBV propagation and whether ABHD2 can be used as a target for anti-HBV drugs.
Materials and Methods
Design and synthesis of ASODN
The entire ABHD2 mRNA sequence (GenBank accession number: NM 007011) was chosen as a potential target sequence. The secondary mRNA structure was predicted using the MFOLD Web server (ZUKER, 2003). Five regions with structural motifs deemed favorable for ASODN binding were chosen as targets (Patzel et al., 1999). Using a BLAST search, the ASODNs targeting these regions were determined to be specific and did not appear to inhibit the expression of other proteins (Table 1). All ASODNs were synthesized on an ABI8909 nucleic acid synthesis system and purified by Oligonucleotide Purification Cartridge (OPC) (Perkin-Elmer, Foster City, CA), and ASOD lengths were verified by gel electrophoresis. As ASODs containing phosphorothioate are nuclease resistant (Cohen, 1993; Uhlmann et al., 1999), all ASODNs generated were chemically modified with phosphorothioate by substituting the oxygen molecules of the phosphate backbone with sulfur.
ABHD2, abhydrolase domain containing 2.
Cell culture and transfection
HepG2.2.15 cells (clonal cells derived from HepG2 cells transfected with a plasmid containing more than full-length HBV DNA) that secrete hepatitis B virions (Sells et al., 1987, 1988) were kindly provided by the Beijing Medical University and were originally provided by the Mount Sinai Medical Center, New York. HepG2.2.15 cells were cultured in minimal essential medium (Invitrogen, Carlsbad, CA) supplemented with 10% fetal bovine serum (Gibco, Grand Island, NY), 380 μg/mL antibiotic G-418 sulfate (Promega, Madison, WI),
Detection of ABHD2 mRNA by semiquantitative reverse transcription–polymerase chain reaction
Seventy-two hours posttranscription, total cellular RNA was extracted from HepG2.2.15 cells using TRIzol Reagent (Invitrogen) according to the manufacturer's instructions. The quality of the purified RNA was assessed using formaldehyde agarose gels. Reverse transcription–polymerase chain reaction (RT-PCR) was performed as previously described (Yang et al., 2005). Briefly, cDNA templates were synthesized from 1 μg total RNA using SuperScript II reverse transcriptase (Invitrogen) and an oligo(dT) primer. The PCR was performed in a 20 μL reaction volume under the following conditions: 5 minutes at 94°C followed by 27 cycles of 94°C for 20 s, 60°C for 20 s, and 72°C for 20 s, with a final incubation for 5 minutes at 72°C. PCR products were then subjected to electrophoresis on a 2.0% agarose gel, bands visualized by ethidium bromide staining, and respective band intensities measured by scanning the gel with Gel Doc 1000 (Bio-Rad, Hercules, CA). PCR products were quantified by densitometry and normalized with respect to glyceraldehyde 3-phophate dehydrogenase (GAPDH) as an internal control. The forward (fp) and reverse (rp) primers used were ABHD2-fp (5′-GCCCCACCTGACCTCTACT-3′), ABHD2-rp (5′-AACGAAAGTGCGGATGTATT-3′), GAPDH-fp (5′-ACCACAGTCCATGCCATCAC-3′), and GAPDH-rp (5′-TCCACCACCCTGTTGCTGTA-3′).
Analysis of the ABHD2 by western blot
Rabbit anti-human ABHD2 antibodies were prepared by professor Yanning Xue and their specificity was confirmed. To detect cellular expression of ABHD2, HepG2.2.15 cells were trypsinized after transfection and lysed with lysis buffer (0.5% deoxycholic acid sodium salt, 0.1% sodium dodecyl sulfate, 1% Nonidet P-40, and a protease inhibitor cocktail [Roche Molecular Biochemicals, Mannheim, Germany]). Equal amounts (50 μg) of protein were run on a 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and then transferred onto Hybond–polyvinylidene difluoride membranes (Amersham, Arlington Heights, IL). Blots were blocked in Tris-buffered saline (TBS) containing 5.0% nonfat dry milk, washed with TBS + 0.2% Tween 20 (TBST), and then probed with rabbit anti-human ABHD2 antibodies or incubated with primary mouse anti-human actin (Pharmacia, Hong Kong, China) in TBS containing 2.5% nonfat dry milk at room temperature for 1 hour. After washing with TBST, blots were probed with a secondary peroxidase-conjugated antibody in TBS containing 2.5% nonfat dry milk at room temperature for 1 hour. Blots were then washed and immunoreactive proteins were visualized on a autoradiogram using ECL Western blot detection reagents (Santa Cruz Biotechnology, Santa Cruz, CA) and exposing blots to X-Omat BT Film (Kodak, Rochester, NY). Bands were quantified by densitometry using an ATTO Densitograph (ATTO Corporation, Tokyo, Japan) normalized against the β-actin internal control.
HBV DNA detection in cell culture medium by RT-PCR
To assess the effect of ASODNs on HBV DNA levels, HBV DNA concentrations in culture medium were determined by RT-PCR. Posttransfection, cell culture media were incubated at 94°C for 15 minutes. After centrifugation at 12,000 g for 10 minutes, supernatants were used as an RT-PCR template as previously described (He et al., 2001; Wang et al., 2002). The forward HBV DNA primer 5′-GGA GTA TGG ATT CGC ACT CCT C-3′ and the reverse primer 5′-TTG TTG TTG TAG GGG ACC TGC CT-3′, in addition to the fluorescent probe 5′-ACT TCC GGAAAC TAC TGT TAG ACG A-3′ and the quenching probe 5′-GTA GTT TCC GGA AGT-3′, were utilized. PCR amplification and analysis were performed using the iCycler real-time PCR detector (Bio-Rad). Assays were repeated in triplicate and the average threshold cycle values were used to determine HBV DNA concentrations. The inhibitory rate was calculated using the follow formula: IR (%) = (Ccontrol − Ctester)/Ccontrol × 100%, where Ccontrol represents HBV DNA copies in HepG2.2.15 cells and Ctester represents HBV DNA copies in cells transfected with ASODNs.
HBsAg and HBeAg detection assays
To assess the effect of ASODNs on HBV protein levels, HBsAg and HBeAg concentrations in culture medium were determined using diagnostic HBsAg and HBeAg enzyme-linked immunosorbent assay kits (Sino-American Biotechnology Co., Beijing, China) as described by the manufacturer. The inhibitory rates were calculated according to the following formula: inhibitory rate (%) = Acontrol − Atest/Acontrol × 100%. Assays were performed in triplicate and the average inhibitory rate expressed as the mean ± standard deviation.
Cell proliferation and apoptosis assays
MTS assays for cell proliferation were performed using the CellTiter 96 Aqueous One solution cell proliferation assay kit (Promega). The assay for apoptosis detection was performed using the Annexin V-FITC kit (Baosai, Beijing, China).
Statistical analysis
Data are expressed as mean ± standard deviation. Statistical analyses were performed using the Student's t-test (2-tailed) and one-way analysis of variance. All data represent at least 2 independent experiments.
Results
The effect of ASODNs on ABHD2 expression
To determine whether ABHD2 affects HBV replication, ASODNs were used to downregulate ABHD2 expression levels in HepG2.2.15 cells. To test the efficacy of respective ASODNs, their effect on ABHD2 mRNA and protein expression levels were first examined by semiquantitative RT-PCR and western blot analyses. HepG2.2.15 cells treated with AB1, AB3, AB4, and AB5 had significantly downregulated (76.71%, 85.97%, 77.85%, and 50.79%, respectively) ABHD2 mRNA expression levels (Fig. 1A–C) and the ABHD2 protein expression levels were significantly diminished by 93.83% and 92.15%, respectively, following treatment with AB3 and AB5 (Fig. 2A, B). Based on these results, AB3 was selected for use in subsequent experiments designed to examine ASODN effects on protein expression and viral infections.
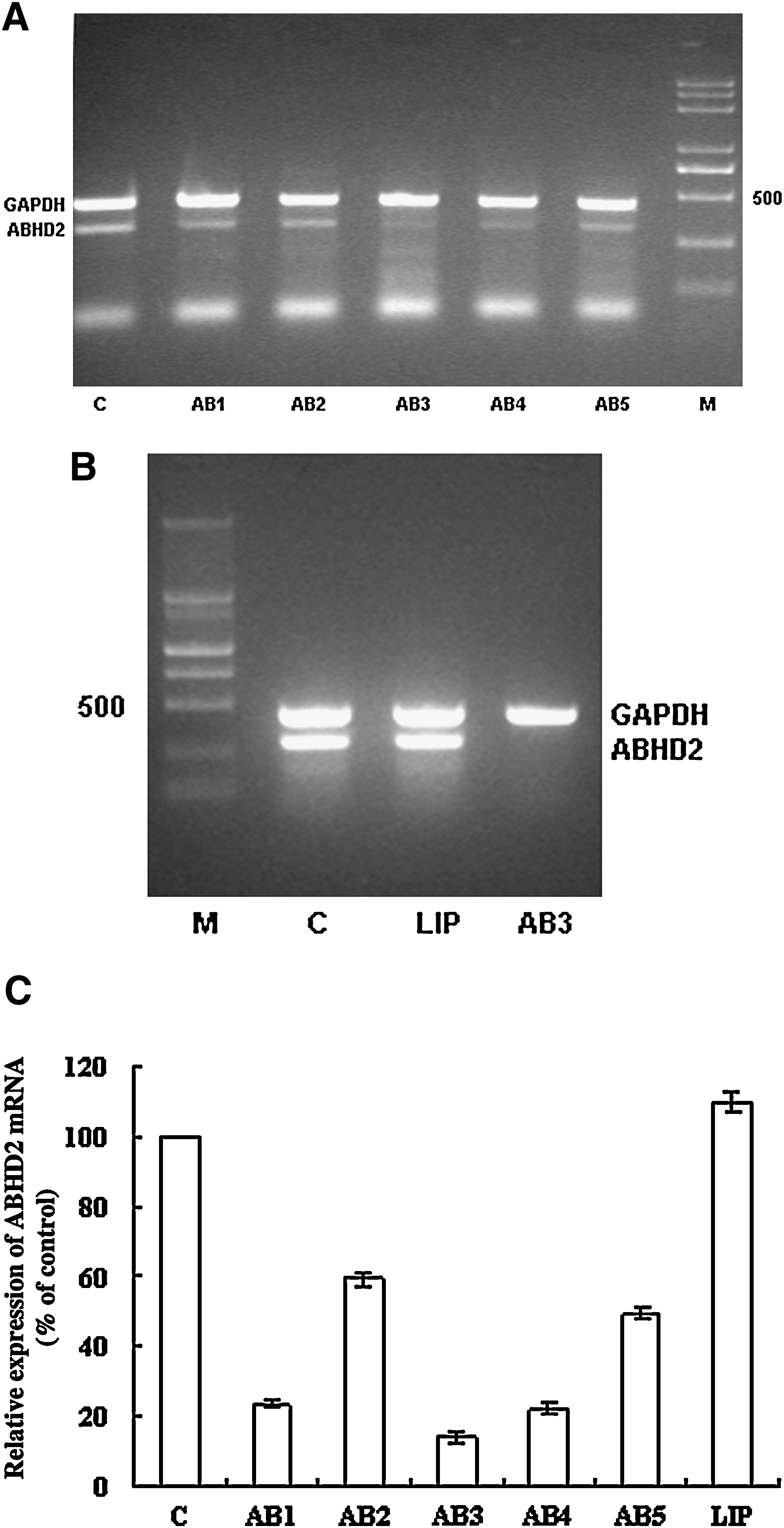
Expression of abhydrolase domain containing 2 (ABHD2) mRNA in antisense oligonucleotide (ASODN)–treated HepG2.2.15 cells.
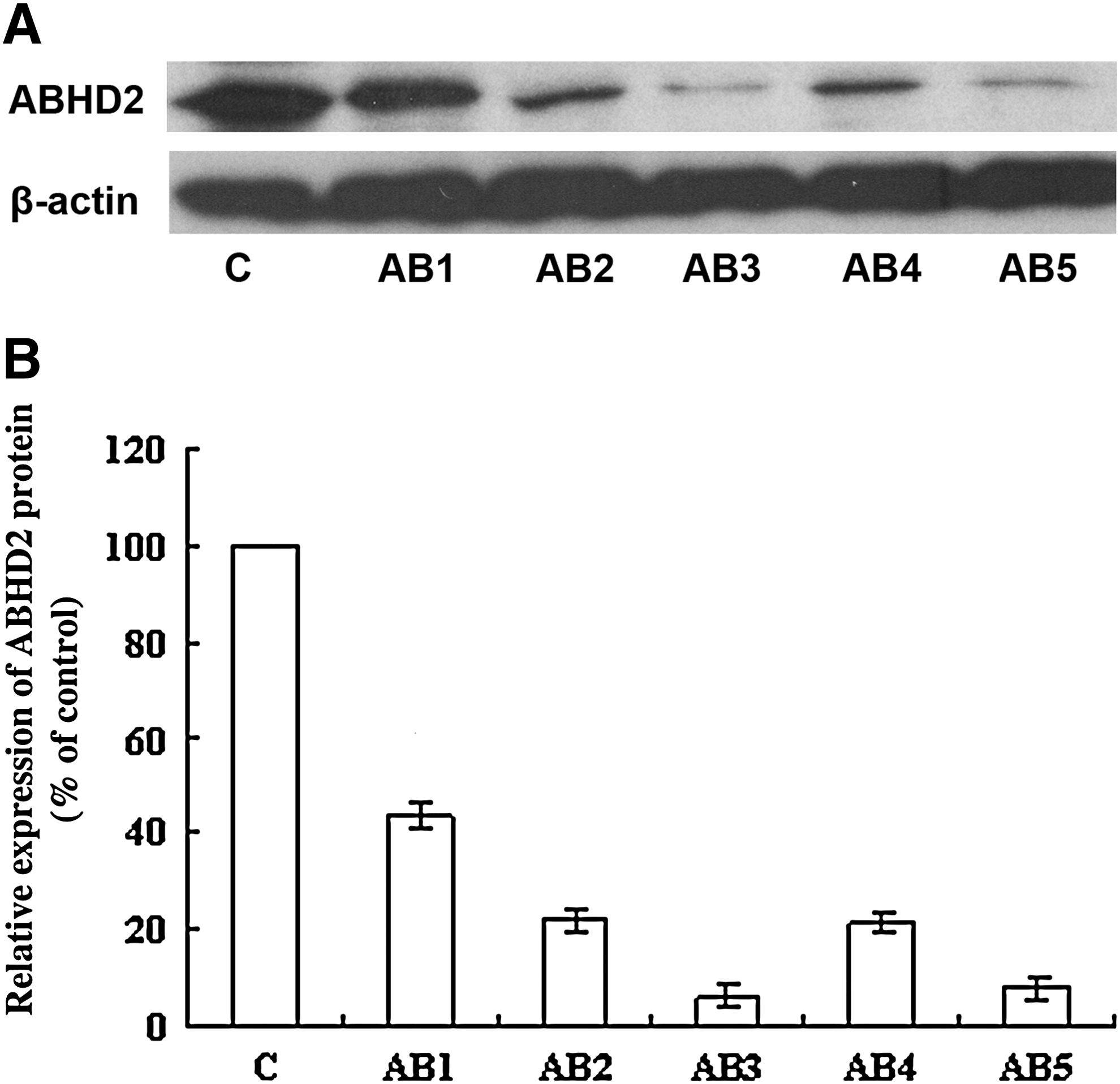
ABHD2 protein expression in ASODN-treated HepG2.2.15 cells.
AB3-mediated downregulation of ABHD2 expression is specific and dose dependent
To exclude nonspecific effects that may be conferred by AB3, 2 controls, SA3 (5′-GAAGGAGCCTGGGGATTTTA-3′) and RA3 (5′-TCCCTCCAGTACACGTCTAA-3′), were used. SA3 is directed against AB3 and used to determine the effect of phosphorothioate ODNs on cells. RA3 is a scrambled AB3 sequence used to test whether the AB3 sequence nonspecifically affects cell functions. These results showed that treatment with 0.8 μM SA3 or RA3 did not alter ABHD2 mRNA or protein expression levels compared with cells treated with serial dilutions of AB3, demonstrating a dose-dependent downregulation of ABHD2 mRNA and protein levels (Figs. 3–5).
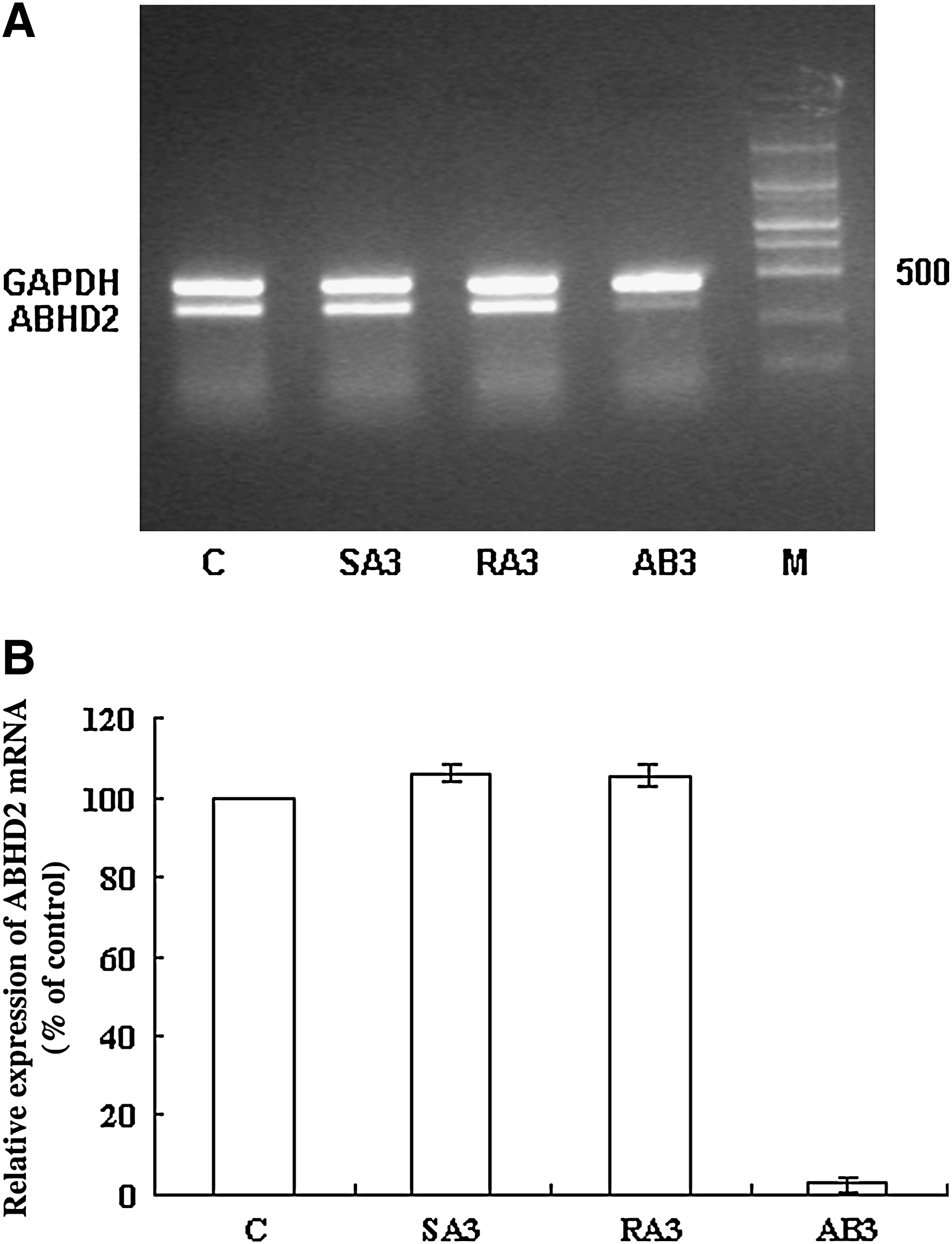
AB3 specifically downregulates ABHD2 mRNA.
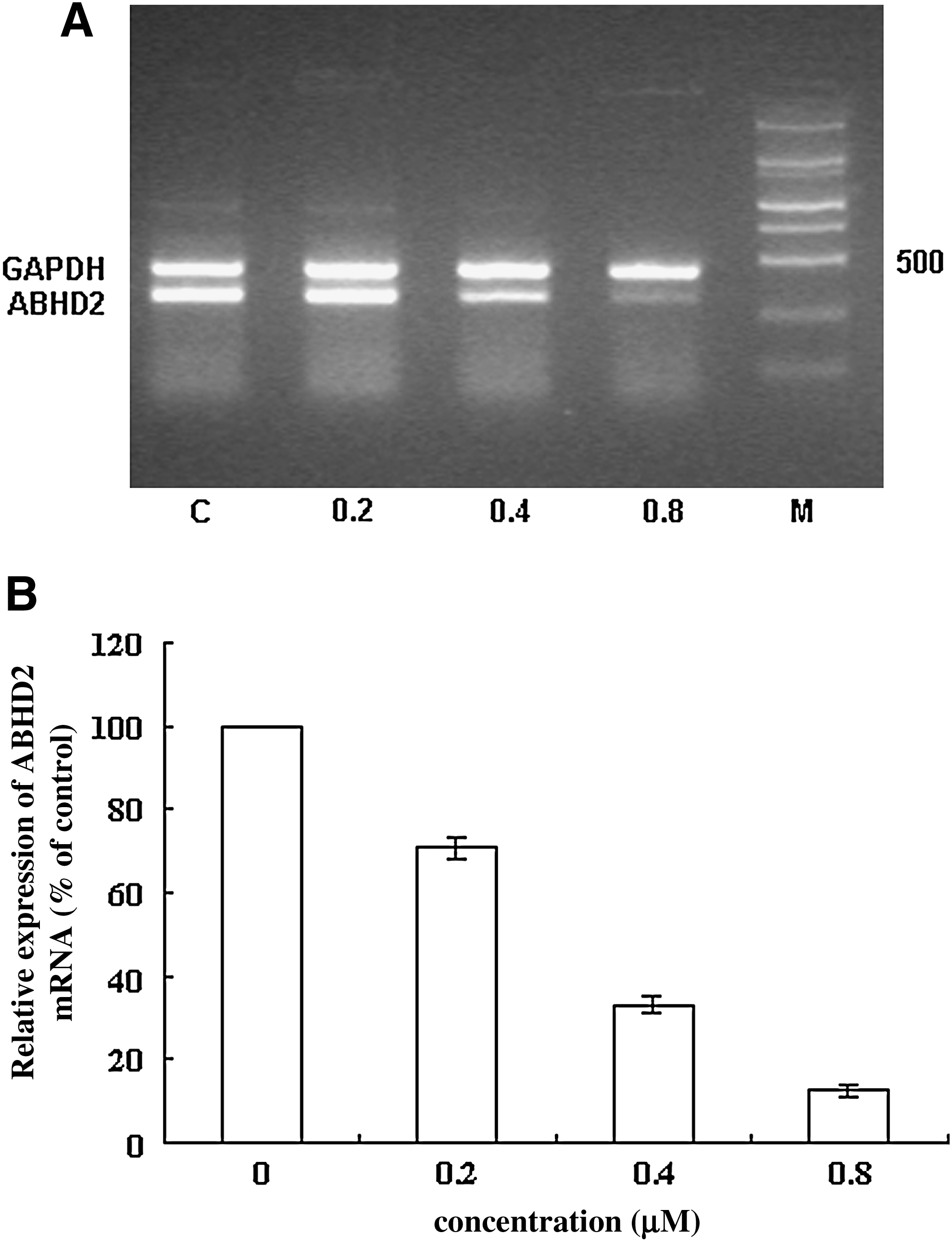
Downregulation of ABHD2 mRNA by AB3 is dose dependent.
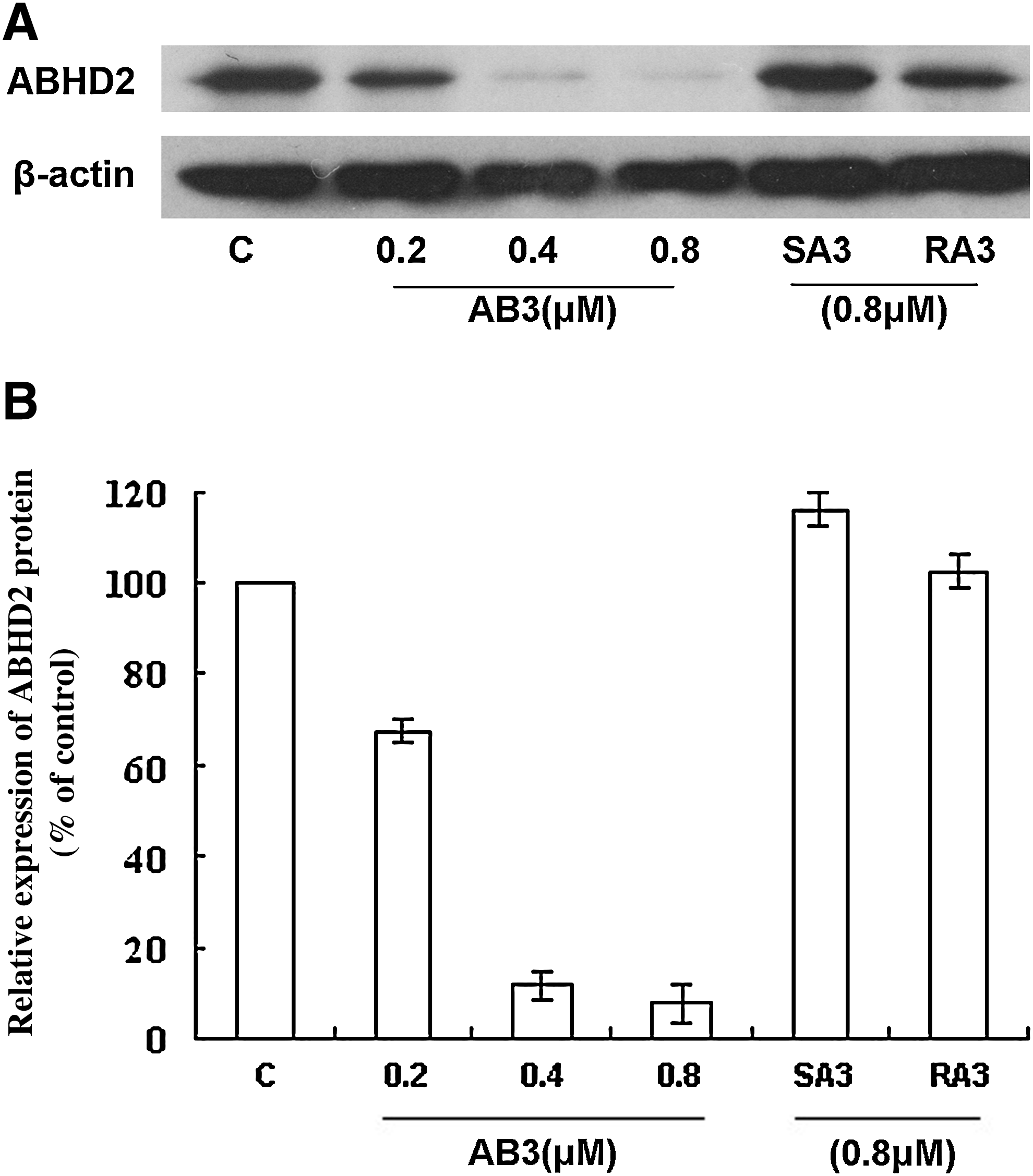
Downregulation of ABHD2 protein expression by AB3 is dose dependent and specific.
HBV replication is reduced following treatment with AB3
To detect whether blocking HBV propagation was specific, HepG2.2.15 cells were treated, respectively, with 0.8 μM AB3, SA3, or RA3 and the levels of HBV DNA, HBsAg, and HBeAg secreted into cell media were determined. These results demonstrated that SA3 and RA3 did not affect HBV replication in HepG2.2.15 cells (Fig. 6).
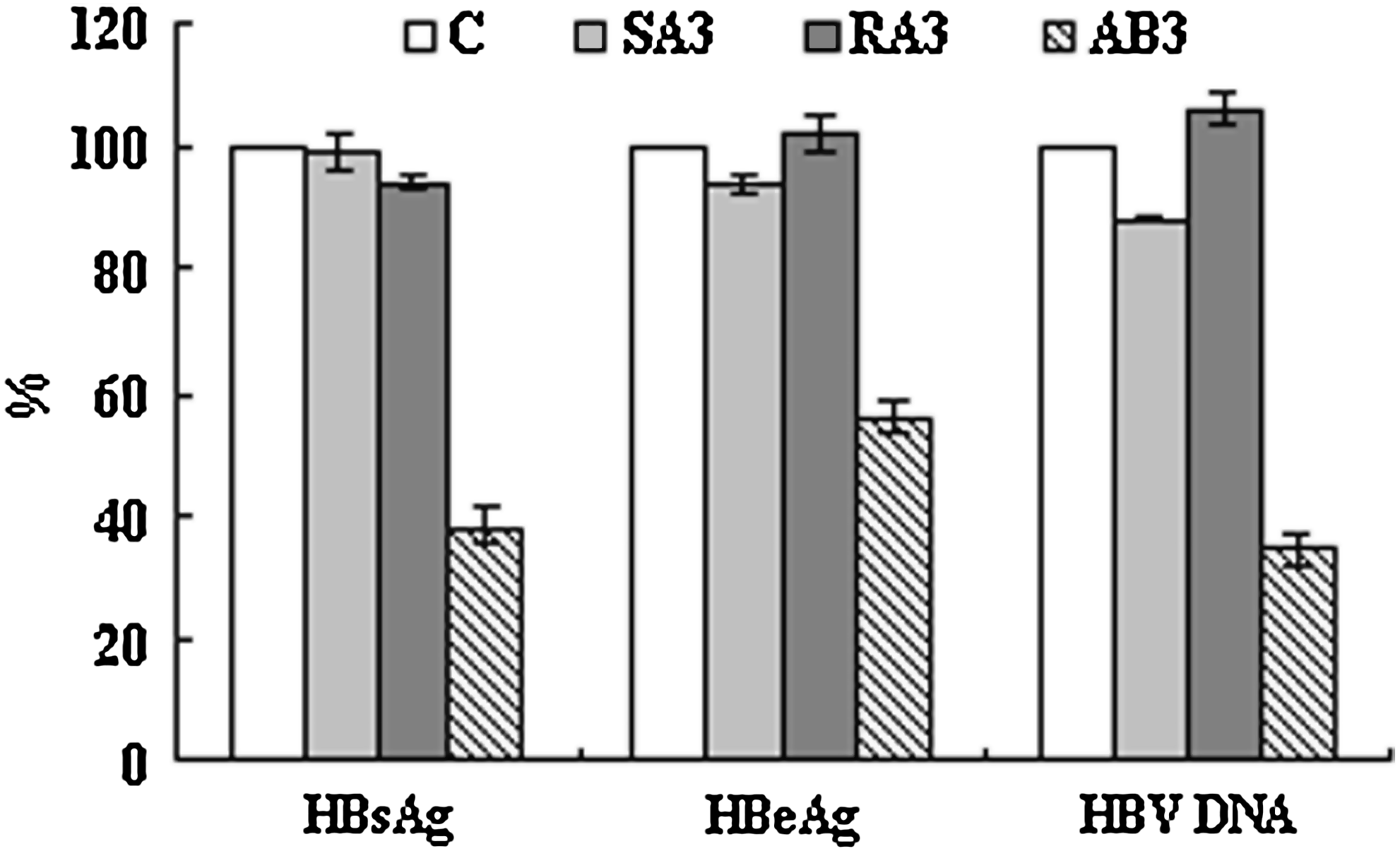
Hepatitis B virus (HBV) DNA, hepatitis B “e” antigen (HBeAg), and hepatitis B surface antigen (HBsAg) expression in HepG2.2.15 supernatants following treatment with either AB3, SA3, or RA3. Expression rates were calculated and expressed as the mean ± SD from 3 independent experiments, assuming that the expression levels of HBV DNA, HBsAg, and HBeAg in control cells were 100% (C, negative control).
AB3 reduced HBV production in HepG2.2.15 cells in a dose-dependent manner
To determine whether ABHD2 inhibition could block HBV propagation, HepG2.2.15 cells were treated with serial dilutions of AB3 and HBV DNA, HBsAg, and HBeAg were detected after 3 days (Fig. 7). These data demonstrated that AB3 (0.8 μM) inhibited HBV DNA, HBsAg, and HBeAg secretion into culture supernatants (65.50%, 67.47%, and 53.04% inhibition, respectively) (Fig. 7), suggesting that AB3-mediated inhibition of ABHD2 blocked HBV propagation in a dose-dependent manner.
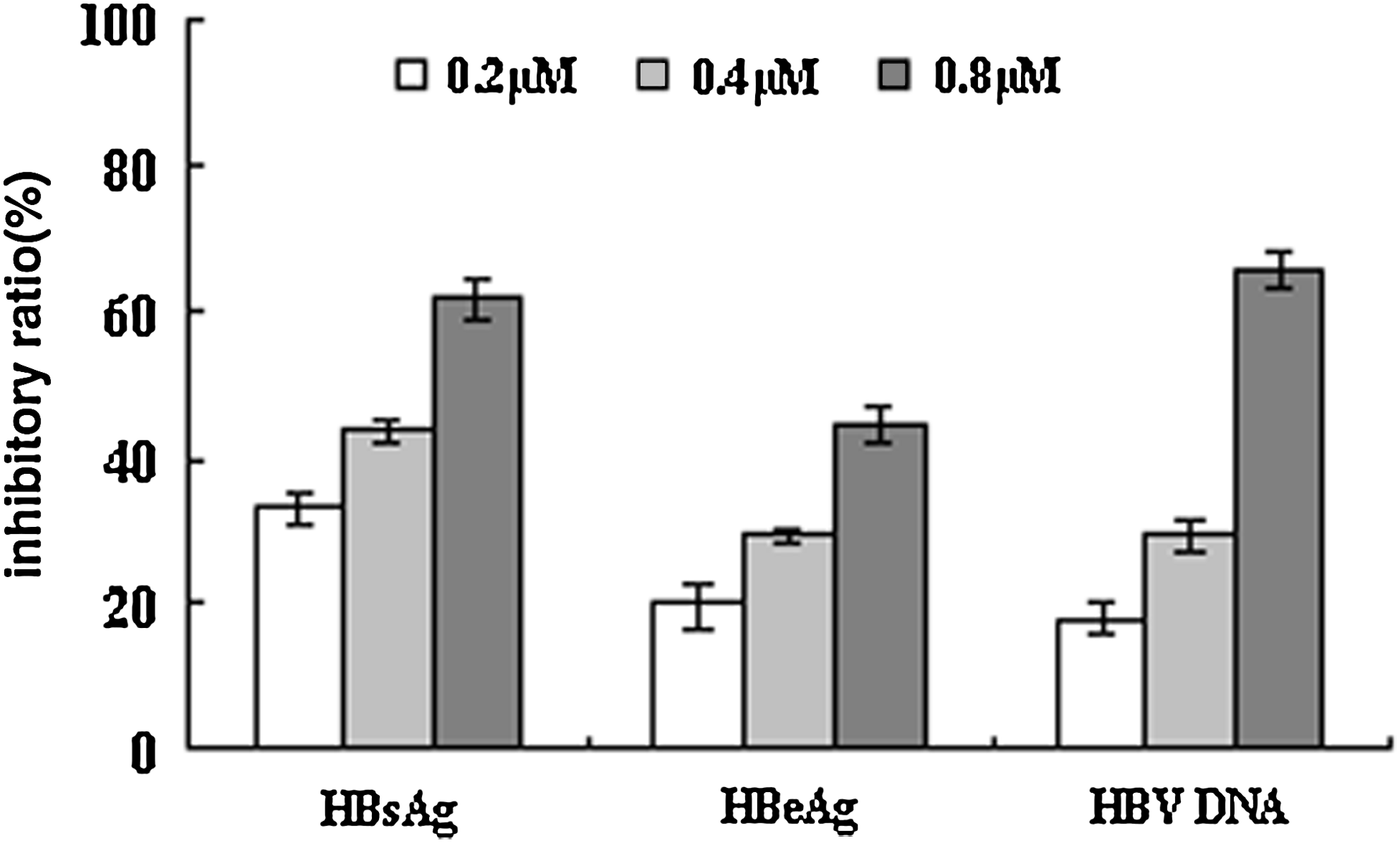
Reduction in HBV production by AB3 is dose dependent. HepG2.2.15 cells were treated with AB3 dilutions for 3 days. The inhibitory rates are expressed as the mean ± SD from 3 independent experiments.
Downregulation of ABHD2 does not affect cell viability
To examine the viability of HepG2.2.15 cells following AB3 treatment, cell proliferation was assessed using the MTS assay in addition to screening for apoptosis. Treatment of HepG2.2.15 cells with serial AB3 dilutions was not cytotoxic (determined by cellular proliferation rates) even at 6.25-fold higher concentrations of AB3 (5 μM, Fig. 8). Characterization of apoptosis showed that treatment with 5 μM AB3 cells did not induce apoptosis (data not shown), suggesting that HepG2.2.15 cell function was not affected by AB3 treatment over time at various concentrations.
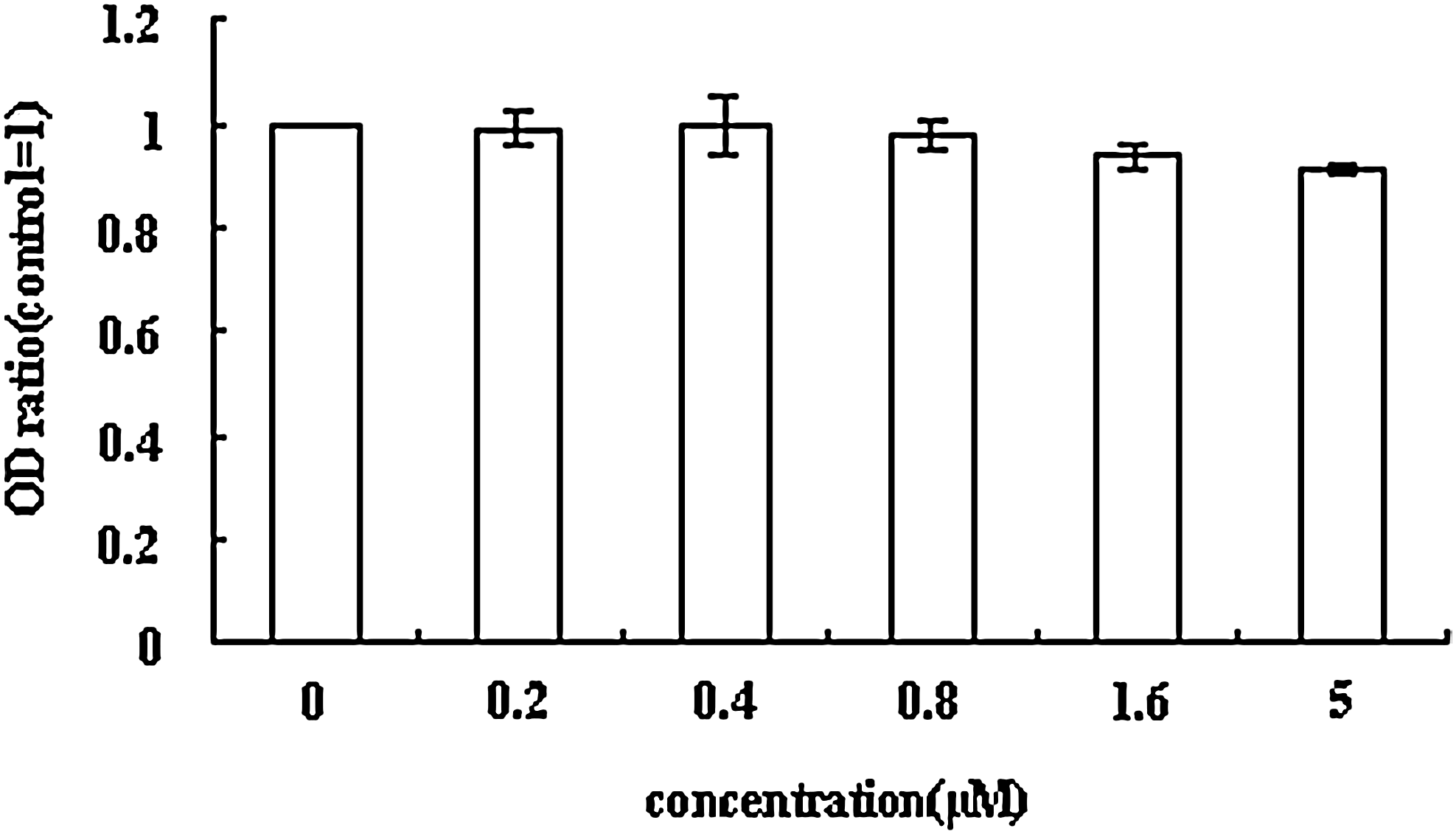
Downregulation of AHBD2 does not affect cell viability. We assumed that the optical density (OD) value of control cells was 1. The OD ratios are expressed as mean ± SD from 3 independent experiments.
Discussion
Although specific functions for ABHD2 have not been well defined to date, the characteristic α/β hydrolase fold of the protein family to which ABHD2 belongs sheds light on its potential function. The α/β hydrolase enzyme family is rapidly becoming one of the largest groups of structurally related enzymes with diverse catalytic functions. Each of these family members has a related protein fold referred to as the α/β hydrolase fold (HOLMQUIST, 2000). The protein core of each enzyme is also similar in structure and comprised of an α/β sheet consisting of 5–8 β-sheets connected by α-helices that form an α/β/α sandwich. All protein family members possess a nucleophile–histidine–acid catalytic triad borne on loops that are the most highly conserved structural features of the fold. However, only the histidine component of the catalytic triad is conserved among all family members, with the nucleophile and acidic loops containing more than one type of amino acid. The nucleophile could be a Ser, Cys, or Asp residue and the acid loops could accommodate an Asp or Glu residue. Members in this family generally act as enzymes and include acetylcholinesterases, dienelactone hydrolases, lipases, thioesterases, serine carboxypeptidases, proline iminopeptidases, praline oligopeptidases, haloalkane dehalogenase, haloperoxidases, epoxide hydrolases, hydroxynitrile lyases, and others.
The virus generally uses host-cell enzymes to carry out its replicative life cycle, including a number of enzymes with hydratase catalytic activities. Carboxypeptidase N, a plasma metalloprotease can bind to and activate the HBV core promoter (Zhang et al., 2003). Moreover, the cellular enzyme S-adenosyl-
To examine the effects of ABHD2 on HBV replication in vitro, we used ASODNs to downregulate ABHD2 expression levels in HepG2.2.15 cells. ASODNs are short, single-strand DNA sequences (13–25 nucleotides) thought to hybridize to specific mRNA sequences inducing target RNA degradation by RNase-H and/or by forming a stable DNA-RNA duplex and a special local structure, which might hinder RNase-H cleavage activity that results in blocking protein translation (Dias and Stein, 2002; Scherer and Rossij, 2003; Coma et al., 2004). The data presented in this report showed that downregulation of ABHD2 using AB3 significantly reduced HBV DNA, HBsAg, and HBeAg levels in HepG2.2.15 cell supernatants, suggesting that AB3-mediated ABHD2 suppression blocked HBV DNA replication and HBV mRNA transcription, stability, or translation and reduced the export of HBeAg and HBsAg. We therefore hypothesized that ABHD2 was essential to HBV replication and that ABHD2 might therefore be a potential target for anti-HBV drug development. In our previous work, the expression of ABHD2 was downregulated by lamivudine (Ding et al., 2008). However, several other reports showed that lamivudine treatment did not reduce HBV-specific RNAs that completely blocked HBV DNA replication (Doong et al., 1991; Lai and Yuen, 2000). Additional research will be required to identify the reasons for this discrepancy.
It is well known that phosphorothioate ASODNs are associated with nonspecific effects caused by the nonspecific cleavage of unintended targets (Woolf et al., 1992). For this reason, the specificity of AB3 was also analyzed by RT-PCR and western blot. These results showed that the levels of β-actin protein and GAPDH mRNA were unaffected by AB3 treatment; however, AB3 downregulated ABHD2 expression in a dose-dependent manner. These data suggested that AB3-mediated downregulation of ABHD2 did not occur as a consequence of the downregulation of either mRNAs or protein synthesis of housekeeping genes. As phosphorothioate ODNs (SA3) did not affect HBV propagation (nor did a scrambled sequence of AB3) it suggested that the reduction of HBV DNA levels were not the result of nonspecific effects of AB3 on cell function/physiology. Moreover, AB3 did not affect HepG2.2.15 cell proliferation or induce apoptosis even at high doses (5 μM). These results suggested that reductions in HBV DNA levels were not caused by abnormalities to cell function resulting from AB3 treatment.
The data presented in this report suggest that AB3 holds the potential of serving as a novel anti-HBV treatment. Current antiviral therapies for HBV include treatment with alpha-interferon– or lamivudine–based therapies, but long-term resolution of disease using these approaches has been disappointing because of low seroconversion rates and the development of drug-resistant viral mutants (Papatheodoridis et al., 2002; Lai et al., 2003; Rapti et al., 2007). Therefore, there is an urgent need to develop more-effective antiviral therapies that can reduce or eliminate viral infections completely, with fewer side effects than those already associated with present treatment modalities. The use of ASODNs as antiviral agents has emerged as a powerful new approach for treating HBV infections (Wagner and Flanagan, 1997; Veal et al., 1998) and many ASODN drugs are now being widely developed. For instance, Vitravene (ISIS Pharmaceuticals, Carlsbad, CA) is an antisense drug currently used for the treatment of cytomegalovirus infections, which has been already approved by the Food and Drug Administration (MARWICK, 1998; STIX, 1998; WACHECK, 2006). In this report, reduction of HBV replication by AB3 suggested that AB3 might be developed as a new drug for HBV therapy.
In conclusion, we identified ABHD2 as a new HBV infection-related gene, and targeting ABHD2 with ASODNs blocked HBV propagation, demonstrating that ABHD2 is essential to HBV replication and that it might therefore serve as a target for anti-HBV drug development.
Footnotes
Acknowledgments
This work was supported by a grant from the National Natural Science Foundation of China (30901825), a grant from The National Key Technologies R&D Program for New Drugs (2009ZX09301-002 and 2009ZX09503-021), a grant from the National High Technology Research and Development Program (863 Program) of China (2007AA02Z108), and grant from the Important National Science and Technology Specific Projects for Infectious Diseases (2008ZX10002-011).
Author Disclosure Statement
No competing financial interests exist.