
Abstract
Abstract
Physical exercise affects hematological equilibrium and metabolism. This study evaluated the biochemical and hematological responses of a male world-class athlete in sailing who is ranked among the top athletes on the official ISAF ranking list of windsurfing, class RS:X. The results describe the metabolic adaptations of this athlete in response to exercise in two training situations: the first when the athlete was using the usual training and dietary protocol, and the second following training and nutritional interventions based on a careful analysis of his diet and metabolic changes measured in a simulated competition. The intervention protocol for this study consisted of a 3-month facility-based program using neuromuscular training (NT), aerobic training (AT), and nutritional changes to promote anabolism and correct micronutrient malnutrition. Nutritional and training intervention produced an increase in the plasma availability of branched-chain amino acids (BCAAs), aromatic amino acids (AAAs), alanine, glutamate, and glutamine during exercise. Both training and nutritional interventions reduced ammonemia, uricemia, and uremia. In addition, we are able to correct a significant drop in potassium levels during races by correct supplementation. Due to the uniqueness of this experiment, these results may not apply to other windsurfers, but we nonetheless had the opportunity to characterize the metabolic adaptations of this athlete. We also proposed the importance of in-field metabolic analyses to the understanding, support, and training of world-class elite athletes.
Introduction
Some theories have suggested that the metabolism of amino acids plays an important role in producing central fatigue (for an elegant review, see Wilkinson et al., 2010). At the end of the 1980s, Blomstrand and colleagues (1988) proposed in a series of articles that central fatigue could be related to the plasma concentrations of aromatic (AAA) and branched-chain (BCAA) amino acids, their ratio and their relationship with tryptophan availability and serotonin production. However, some data have shown that tryptophan supplementation can extend exercise (Javierre et al., 2010; Segura and Ventura, 1988). In the 1990s, Banister and Cameron (1990) proposed that the increase in ammonia during exercise could lead to central fatigue. Because hyperammonemia has been linked to several central nervous system (CNS) conditions, it is probably linked to the glutamatergic system (Cauli et al., 2008). It is likely that adequate training and diet can improve the cellular machinery, thereby expanding chemical energy accumulation and transduction. These improvements can lead to better energy metabolism by decreasing the deleterious effects of metabolites and increasing both physical and mental performance (de Almeida et al., 2010; Prado et al., 2011).
Windsurfing is a water sport that mixes surfing and sailing (Sedlaczek, 2009). Its practice depends on three basic conditions: the athlete, environment, and equipment. Little is known about metabolism in windsurfing athletes because most articles in this area have focused on physiological parameters (Campillo et al., 2007; Castagna et al., 2007, 2008; Chamari et al., 2003; Melis et al., 2003; Pérez-Turpin et al., 2009; Vogiatzis et al., 2002) and injuries (Dyson et al., 2006; Hetsroni et al., 2006; McCormick and Davis, 1988; Nathanson and Reinert, 1999; Nathanson et al., 2008; Neville and Folland, 2009; Nickel et al., 2004; Orchard et al. 2002; Petersen et al., 2003).
The tactical and strategic decisions needed during a windsurfing competition require a mixture of force and resistance training. In this study, we evaluated a world champion windsurfer who is ranked among the top athletes by the official International Sailing Federation (ISAF) ranking. We investigated metabolic changes in response to a simulated competition in an in-field investigation and metabolic responses to a nutritional and training intervention.
Materials and Methods
Subject
This study evaluated a male world-class athlete in sailing from the windsurfing discipline, class RS:X. The athlete was well instructed regarding the procedures and signed a written informed consent. The study was approved by the Ethics Committee for Human Research of the Federal University of State of Rio de Janeiro and conformed to the requirements and guidance for carrying out research in human subjects (Health National Council, Brazil, 2010). During the trials, the athlete did not have health problems or use ergogenic substances or any other drugs. In addition, clinical examinations, anthropometric measurements, and laboratory tests were performed. Laboratory tests included hematological and biochemical analyses, which allowed any metabolic anomalies that could affect the results or impair their interpretation to be diagnosed.
Experimental design
We simulated a typical Olympic regatta. The athlete performed a race, rested for 30 min, and then competed in another race. Each race lasted 30 min. Blood samples were collected both before and after the races (T1–T4). An additional sample was collected after 120 min of rest (T5). Note that the experimental design provides information regarding the athlete's metabolic condition before and after the first exercise bout (T1–T2), before and after resting (T2–T3), before and after the second exercise bout (T3–T4), and before and after metabolic recuperation (T4–T5).
Figure 1 shows that blood was collected following in-field situations. The experimental time line and blood collection times (T1–T5) are shown.

Experimental design. Blood was collected following in field situations. The experimental timeline and blood collection times (T1–T5) are shown.
Two trials (A and B) were conducted under similar environmental conditions (°C; % relative humidity; wind velocity in km/h). The first trial was conducted using regular training and a regular diet. After analysis of the results, diet and training were modified, and the second trial was conducted.
Training interventions
The exercise program was constructed according to the American College of Sports Medicine Guidelines (ACSM Position Stand, 1998). The intervention protocol for this study consisted of a 3-month facility-based program using neuromuscular training (NT) and aerobic training (AT).
The training protocol was designed to strengthen the most important large muscle groups for windsurfing-specific activities and to improve cardiorespiratory capabilities. The athlete was asked to perform the NT twice weekly for 3 h each session for a total of 36 training sessions, and the AT twice weekly for 30 min each session for a total of 24 training sessions on different days from the NT. During each training session, the athlete was under direct supervision from a physical therapist and a trained exercise instructor.
The volume and intensity of the intervention protocol was controlled by oxygen uptake and by a cardiofrequency GPS (Garmin Forerunner® 405CX) that continuously recorded time, distance, pace and heart rate during the training.
Nutritional intervention
We suggested that the athlete should keep his regular dietary habits during the trials. During the intervention, we suggested nutritional changes to promote anabolism, achieving an intake of 16 MJ (20% from proteins, 60% from carbohydrates). The athlete was instructed to increase the number of meals to a minimum of six per day. We instructed the athlete to consume high biological-value proteins and carbohydrates in all meals, combining different absorption times from both nutrients (e.g., casein vs. whey protein; starch vs. maltodextrine). We also prescribed an intake of multivitamins and multiminerals to reload micronutrients, especially potassium, immediately before and after training.
Laboratory analysis
Samples for biochemical assays were collected from the antecubital vein and centrifuged (3,000 × g; 10 min). All of the blood sampling procedures, centrifugation, and blood storage were carried out immediately after collection to avoid the loss of volatile compounds. Serum and plasma were aliquoted, frozen in liquid nitrogen, and stored at −80°C for later analysis. The blood, serum, and plasma were analyzed in duplicate or triplicate, when necessary, and corrected against a standard curve with no fewer than five points.
Biochemical analyses were conducted using an automatic analyzer (ADVIA® 1200 Chemistry System, Siemens, USA). Alanine aminotransferase (ALT), alkaline phosphatase (ALP), aspartate aminotransferase (AST), creatine phosphokinase muscle-brain fraction (CKMB), lactate dehydrogenase (LDH), γ-glutamyltransferase (γGT), creatinine, glucose, urate, and urea were measured by enzymatic methods. Chloride, phosphate, serum iron, transferrin saturation, and total iron-binding capacity were quantified by colorimetric methods. Sodium and potassium were measured by the ion selective electrode method. Creatine phosphokinase (CK) was measured using a kinetic method, and α-1-acid glycoprotein (AGP) was measured using a turbidimetric method. Albumin and total protein were evaluated by an electrophoretic method. Amino acids were measured by an high-performance liquid chromatography (HPLC) method, and free testosterone was quantified by a radioimmunoassay method. Total testosterone and insulin were assessed by electrochemiluminescence. Erythropoietin, ferritin, and myoblobin were measured by chemiluminescence methods. Total cholesterol, phospholipids, high-density lipoprotein (HDL), low-density lipoprotein (LDL), very-low-density lipoprotein (VLDL), and total lipids and triacylglycerols (TG) were measured by the Charbroll & Charonnat method.
Samples for hematological assays were collected into a tube with K3-EDTA (Vacuette, Greiner Bio-One, Germany) and stored at 4°C for later automated analysis (COULTER® STKS™ Hematology Flow Cytometer, Beckman Coulter, Fullerton, CA). White cell counts (total and differential) were performed, and red blood cells and platelets were also measured. We also analyzed coagulation, bleeding time, capillary fragility, clot retraction, platelet count, prothrombin time, and activity by coagulometric methods.
Data presentation
To facilitate visualization and comparison, some data were normalized to the T1 measurement of each trial (absolute values are referenced in each figure). All other data are given as an absolute value.
Results
Branched chain amino acids
Plasma branched chain amino acids (BCAA) were assessed because they play important roles in protein synthesis and are used as a primary energy source by muscles. Our results showed that during trial A, isoleucine decreased by 23% after the first race and did not change thereafter. The plasma amino acid concentration reached 32% of the original level after the second race and remained at this value until after recuperation. The plasma concentration of valine decreased by 21% after the first race and did not change until the second race, during which it decreased an additional 41% (62% total). We were not able to measure a recovery until after recuperation. A similar curve was measured for leucine. Nutritional and training intervention produced a change in BCAA plasma availability. During trial B, isoleucine decreased by 42% during the first race and maintained the same concentration until the end of the second race. After recuperation, the plasma amino acid concentration returned to almost the T1 value. The same happened with both valine and leucine plasma concentrations (Fig. 2).

Intervention decreases BCAA consumption during the second regatta followed by faster restoration of plasma amino acid levels. Trial A (○) and trial B (●) plasma branched chain amino acid (isoleucine, valine, and leucine) levels normalized (%) to the T1 value. T1 values are shown in the inset.
Aromatic amino acids
Aromatic amino acids (AAA) are ketoglucogenic and can directly be used by hepatocytes for gluconeogenesis. During trial A, our results showed that AAA decreased by nearly 15% after the first race, did not change during the rest period, and then decreased again by approximately 40% after the second race. We did not measure an increase in the plasma amino acid pool after recuperation. After intervention, the plasma tyrosine concentration remained constant during the trial. Both plasma tryptophan and phenylalanine fluctuated almost 15–20% during the races, never dropping below 100%. Their concentration increased to levels ∼30% greater than the pretrial conditions (Fig. 3).

Intervention decreases the consumption of aromatic amino acids during exercise. Trial A (○) and trial B (●) plasma aromatic amino acid (tyrosine, tryptophan, and phenylalanine) levels normalized (%) to the T1 value. T1 values are shown in the inset.
Glutamine, glutamate, and alanine
The measurement of glutamine, glutamate, and alanine concentrations in plasma is important because of their roles in energy production in metabolism. The plasma concentrations of both glutamine and alanine decreased by more than 50% during trial A and failed to recover during the recuperation period. Plasma glutamate levels fluctuated less (up to 70% of rest). During trial B, no changes were observed in plasma glutamine or alanine levels. In contrast, glutamate showed opposite plasma concentration kinetics when compared to trial A (Fig. 4).
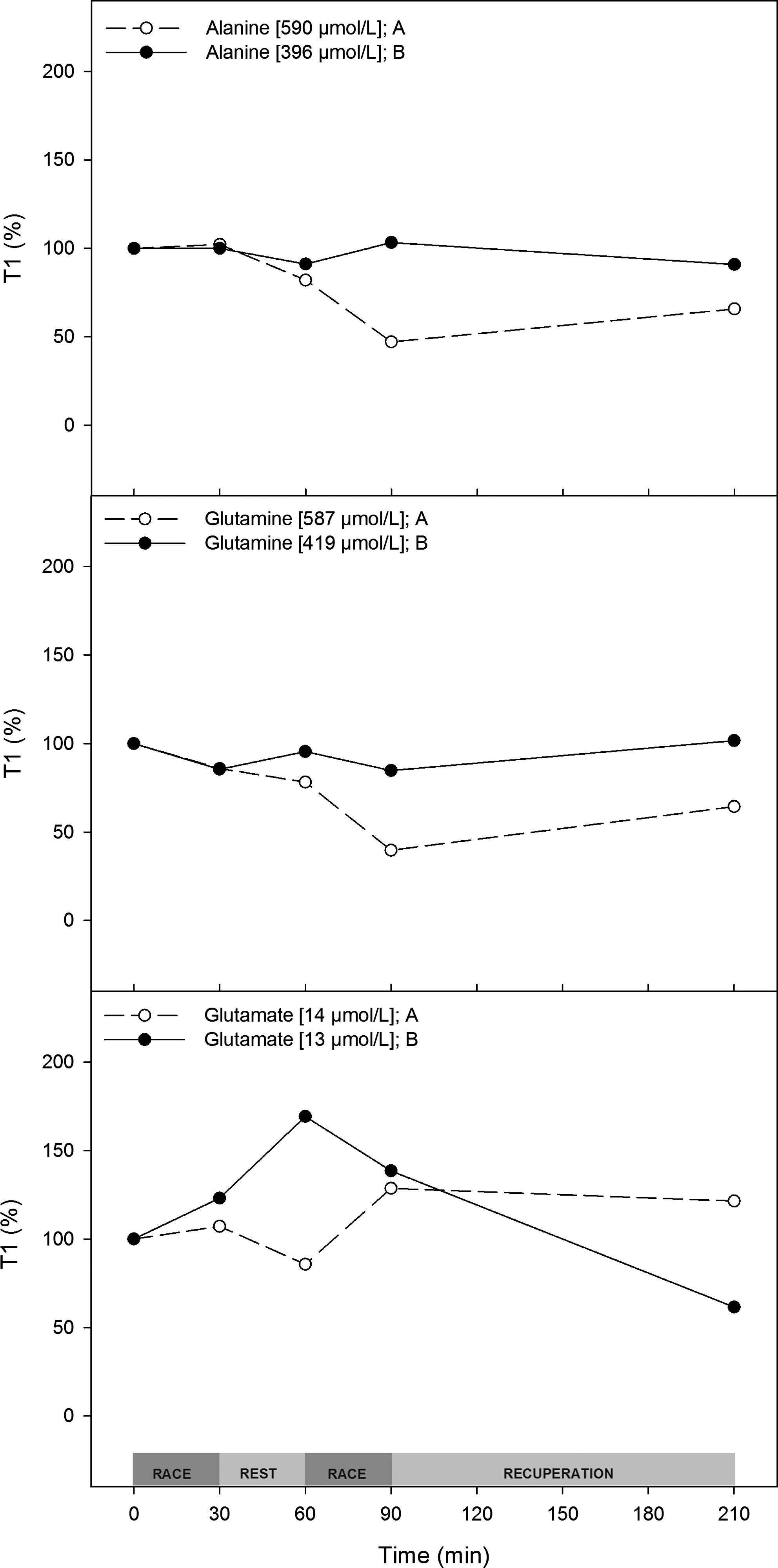
Both training and diet decrease the use of glutamine and alanine. Trial A (○) and trial B (●) plasma amino acid concentrations normalized (%) to the T1 value. T1 values are shown in the inset.
Metabolic pathways
Ammonia is a waste product from the metabolism of amino acids and other nitrogenous compounds. In trial A, ammonemia increased in response to exercise 20–30% more than it did in trial B. In both trials, exercise promoted approximately a sixfold rise in ammonemia after the races, and this returned to initial levels at rest (Fig. 5).

Intervention does not affect exercise-induced ammonemia. Trial A (○) and trial B (●) ammonia concentrations normalized (%) to the T1 value. T1 values are shown in the inset.
Urate is the final catabolite from purines and is an indicator of the rate of IMP production. To indirectly evaluate the contribution of myokinase to ATP production, we measured plasma urea concentrations. Urate rose nearly 20% more during trial A compared to trial B. During trial A, urate increased by up to 35%, reaching a 60% increase when compared with the prerace value. During trial A, urea consistently rose by approximately 20% with a slower increase during recuperation, reaching a final elevation of 25%. After intervention, the average increase in urea during the entire trial was 5% (Fig. 6).
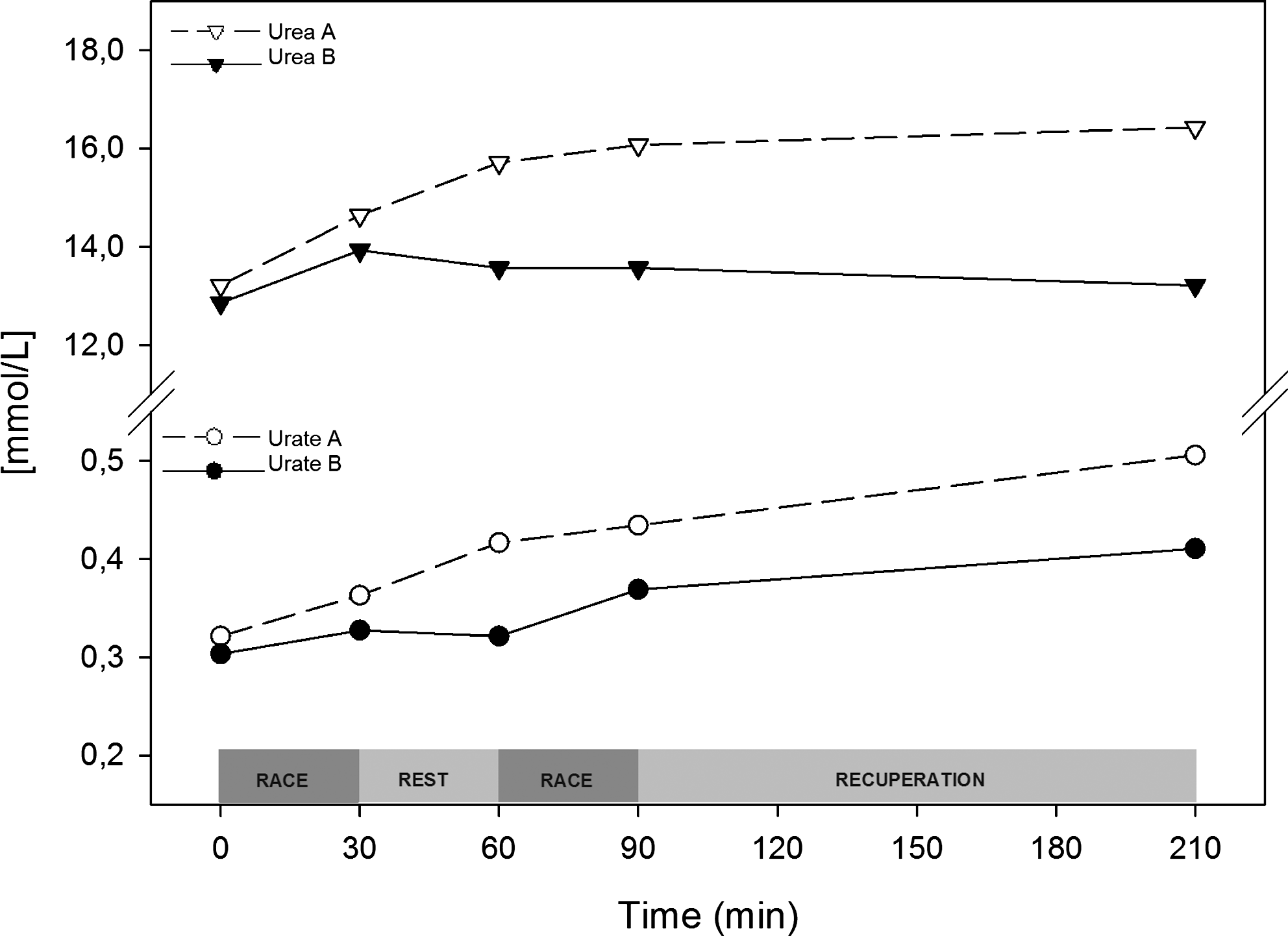
Interventions reduce both uremia and uricemia increases due to exercise. Trial A (○) and trial B (●) plasma urea and urate concentrations are shown as absolute values (mmol/L).
Gluconeogenesis is one of the destinations of amino acid metabolites. To understand the effects of windsurfing exercise on amino acid metabolism, we measured the changes in both serum glucose and insulin. During trial A, glycemia rose by ∼70% after the first race and decreased after the 30-min rest. In the second race, glycemia rose again, reaching a value 43% greater than the prerace value. During trial B, plasma glucose did not change in response to the first race and had a delayed increase compared to trial A. Glucose reached the pretrial values in both trials. Insulin decreased in both trials in response to the two races. In the first race of trial A, insulin decreased by 35% compared to preexercise levels. During the second race, insulin dropped by 55% compared with T1. Insulin returned to the initial concentration after recuperation. During trial B, a similar pattern was observed with a smaller variation when compared with trial A (Fig. 7).

Intervention decreases glycemic fluctuations. Trial A (○) and trial B (●) plasma insulin and glucose concentrations are shown as absolute values (mUI/mL and mmol/L).
Muscle stress biomarkers
Creatinine, an indicator of exercise muscle injury, increased in both trials. During trial A, the basal plasma creatinine concentration was 10% lower than in trial B, and it increased by ∼15–30% during the trial. AST increased by 15–20% during trial A and demonstrated a small fluctuation during trial B. However, the ALT plasma concentration fluctuated by 5–10% in both trials. Other hepatic injury biomarkers, such as ALP, γGT, and AGP, did not change during either trial (data not shown).
The classic muscle injury marker CK increased by approximately 150% during trial A. During trial B, basal CK was almost sevenfold higher than in trial A. However, the plasma concentration of this enzyme showed a 30% fluctuation during trial B. CKMB is a very specific muscle injury marker and acutely responded to exercise in both trials, with a much greater response in trial A.
LDH is a well-known delayed muscle injury marker, and it increased by up to 40%. In trial B, even with higher basal concentrations of CK and LDH, we observed a much smaller increase in these three markers than what was observed in trial A (Figs. 8 and 9).

Changes in stress markers between trials. Trial A (○) and trial B (●) plasma creatinine, aspartate aminotransferase (AST) and alanine aminotransferase (ALT) levels normalized (%) to the T1 value. T1 values are show in the inset.
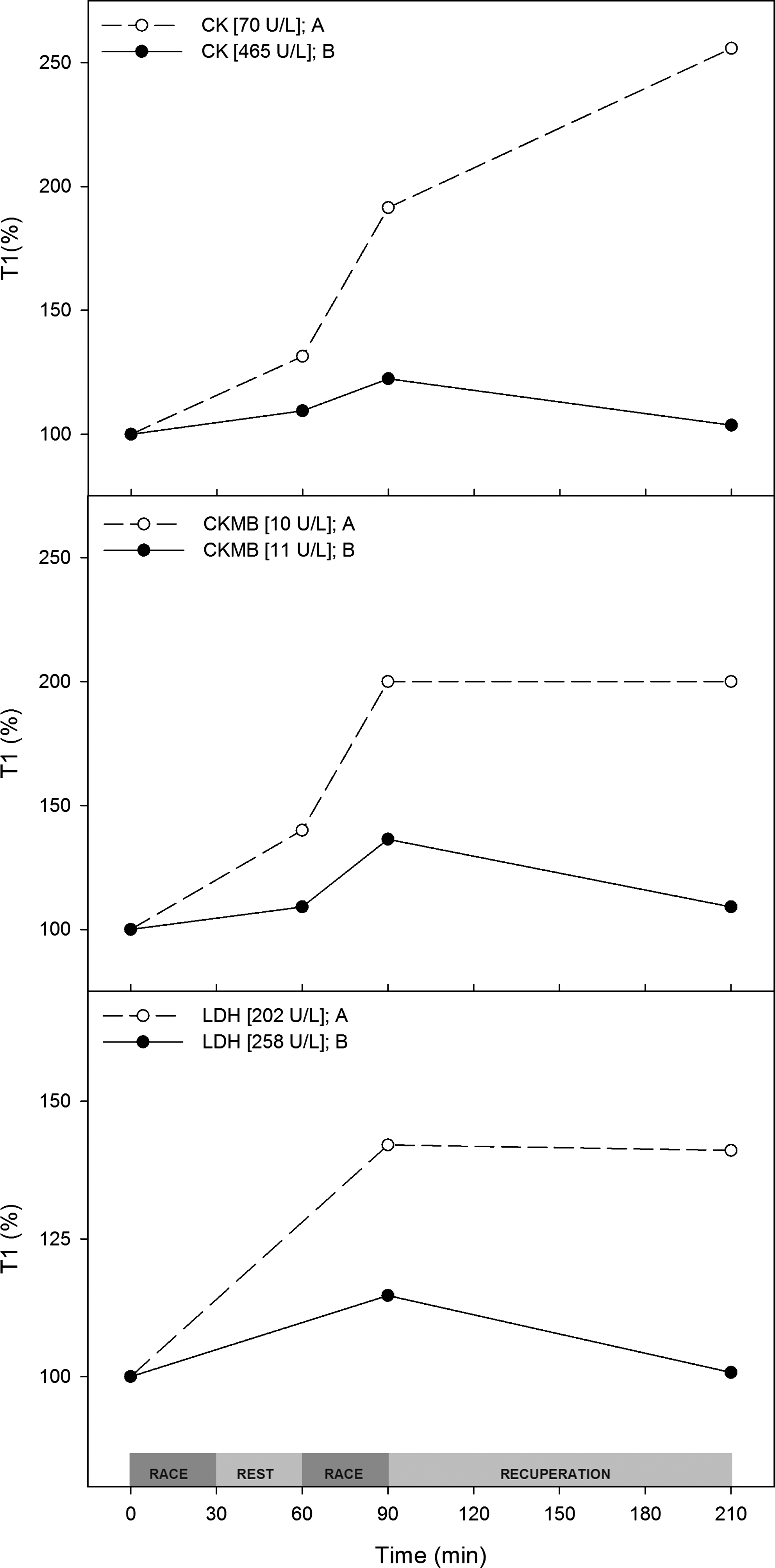
Physical training associated with diet reduces muscle microinjuries. Trial A (○) and trial B (●) plasma creatine phosphokinase (CK), creatine phosphokinase muscle-brain fraction (CKMB) and lactate dehydrogenase (LDH) levels normalized (%) to the T1 value. T1 values are shown in the inset.
During trial A, blood leukocytes increased by approximately 300% after the races and did not change after recuperation. The neutrophil count contributed the most to this increase. Lymphocytes in blood increased in response to exercise and decreased after resting (Fig. 10).
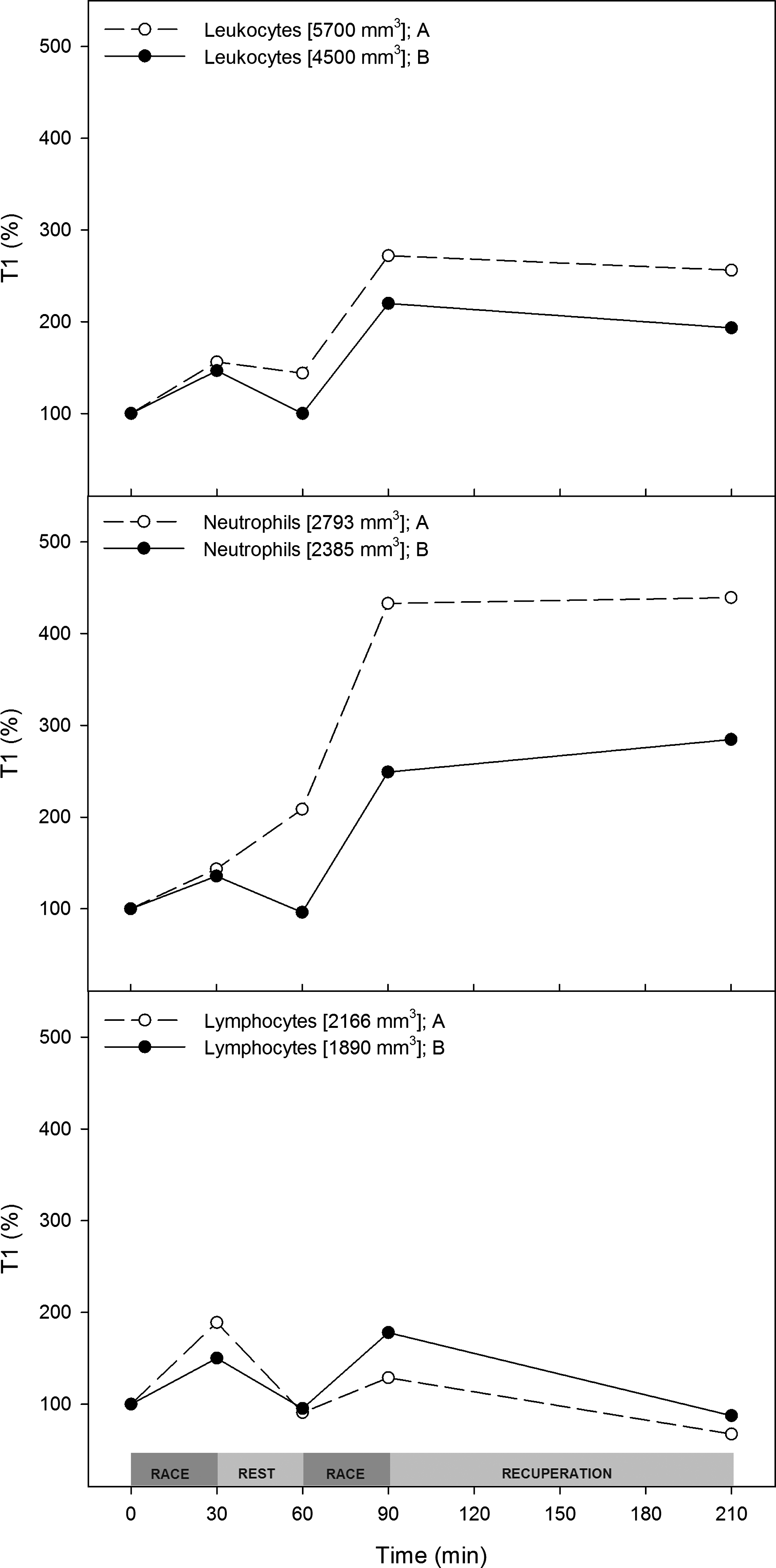
Intervention changes the neutrophil response. Trial A (○) and trial B (●) plasma white blood cell (leukocytes, neutrophils, and lymphocytes) numbers normalized (%) to the T1 value. T1 values are shown in the inset.
Hydroelectrolytic status
No changes in erythrocyte count or hematocrit were found at any time point during either trial (data not shown). Even without signals of dehydration, tropical windsurfing athletes can lose water and electrolytes due to sweating and low rehydration. To evaluate the athlete's blood mineral composition, we measured the main inorganic electrolytes. We did not detect any changes in sodium or chloride during either trial (data not shown). In trial A, phosphate increased by 50–70% in response to exercise; it decreased after rest but never reached the basal level. This effect was mitigated by the intervention. Potassium levels decreased during trial A to 3.3 mmol/L and slowly recovered during the experimental period without reaching basal levels. These changes were abolished by the intervention (Fig. 11).
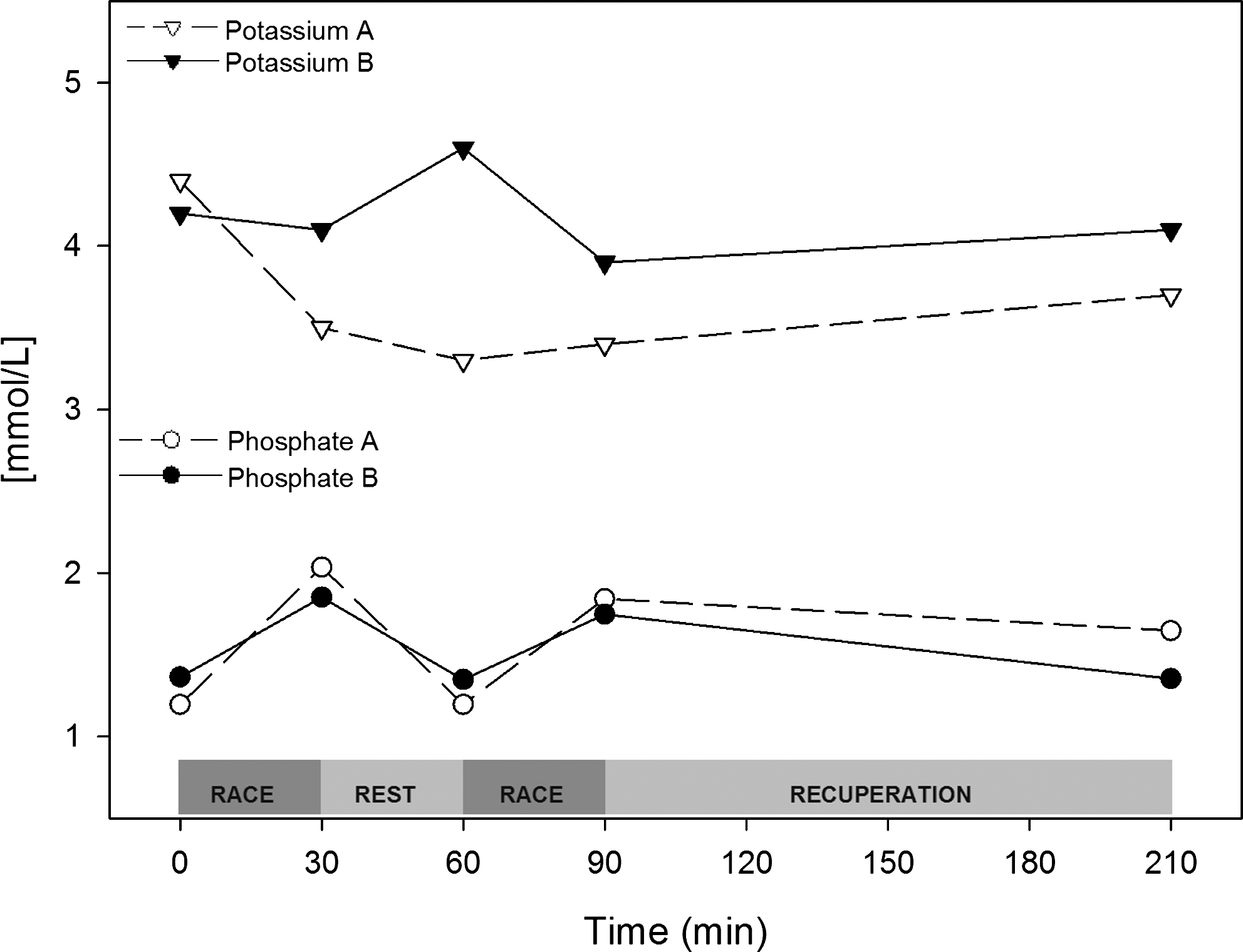
Interventions prevent kalemia from dropping during the regatta without changing the phosphate response. Trial A (○) and trial B (●) plasma potassium and phosphate concentrations are shown as absolute values (mmol/L).
Discussion
Windsurfing competitions require that physical preparation be combined with tactical and strategic decision making. Due to the need for rapid CNS responses, it is important to reduce metabolites that can be deleterious to short-term judgment and/or cognition. In this way, metabolic evaluations can lead to interventions that decrease exercise-induced stress and thereby improve physical and mental performance. Here, we describe a series of metabolic evaluations performed on a world-class windsurfing athlete.
Exercise intensity can be evaluated by increases in the plasma levels of CK, CKMB or LDH (Matsumoto et al., 2009; Rodrigues et al., 2010; Shave et al., 2002). Acute metabolic stress was clearly much greater in trial A than in trial B. In addition, the basal levels of both CK and LDH were higher in trial B. These results suggest that adaptation occurred due to exercise-induced muscle injury during training, resulting in an adjustment of the muscles to exercise-linked stress. In fact, during our studies, plasma AST levels were much higher than those of ALT. It has been show previously by our lab that the AST and ALT biomarkers are also related to hepatic injury. These biomarkers can be used to monitor muscle stress when compared with increases in γGT and alkaline phosphatase (Bassini-Cameron et al., 2007).
It is a well-documented fact that white blood cells are released in response to exercise, and neutrophil release is linked to exercise intensity and duration (Bessa et al., 2008). This effect seems to be regulated by differential cytokine release (Giraldo et al, 2009; Li and Gleeson, 2004). In our study, leukocytes increased by almost 300% in response to trial A, an effect mostly due to the increase in neutrophils (>400%). After intervention, leukocyte and neutrophils response increased by approximately 200 and 250%, respectively, compared with rest levels. Neither leukocytes nor neutrophils returned to their basal levels.
The response of white blood cells (WBCs) to exercise seems to depend on both cytokine and myokine regulation (Haugen et al., 2010; Mathur and Pedersen, 2008; Pedersen et al., 2007). In this study, we measured an acute increase of lymphocytes in response to each exercise bout, which returned to basal levels after exercise stress. Metabolic stress is correlated with the increase in WBCs in response to exercise. In fact, increases in carbohydrate availability and cortisol response, a gluconeogenic hormone, have been used as a link in integrated WBC and metabolic pathways (Peake et al., 2005). Some of our results suggest an increase in metabolic reserves after the intervention, measured as a decrease in gluconeogenesis.
Alanine, glutamine, and BCAA levels in the blood were severely depleted during the first trial. In fact, even after recuperation, their levels remained low. BCAAs have different oxidation rates in liver and muscle (for a review, see Harper et al., 1984). Isoleucine seemed to be somewhat more affected than valine and leucine, which can be explained by the fact that isoleucine is a precursor of glutamine and alanine, which are the most important gluconeogenic amino acids. Similarly, AAAs, which are generally metabolized in hepatocytes, were consumed less during the first exercise bout of trial A and then decreased quickly after the second bout. Phenylalanine and BCAA were similarly consumed in both trials, but with a different pattern regarding recuperation. However, the plasma concentration of Tyr and Trp did not change after the intervention. These data fit the current understanding of the interorgan metabolism of the BCAAs glutamine and arginine proposed by Wu's group (Li et al., 2007; Wu, 2009). In fact, it is reasonable to suppose that because the concentration of an amino acid in blood affects its ratio in the brain (Crowell et al., 1990), it is very important to establish training goals that lead to the maintenance of plasma amino acid levels.
In the first race of trial A, even with an ∼70% increase in glycemia, insulin decreased to 35% of its preexercise levels. This is a well-described metabolic effect that occurs during exercise in the presence of catecholamines (Bangsbo et al., 2007; Kraemer and Ratamess, 2005). In fact, in trial A, glucose increased during exercise in a typical gluconeogenic response. After the intervention, BCAAs, which are still being consumed during exercise, reacted better to the recuperation step. In addition, alanine, glutamine and AAA plasma concentrations were almost unaffected. These data suggest that in trial A, amino acids were being used by the muscles as an energy source and by the liver to produce glucose. A similar result showing that carbohydrate availability and insulin secretion are linked to training conditions was recently reported (Mikulski et al., 2010). These data reinforce the idea of a greater metabolic stress in trial A than in trial B.
Multiple studies have shown that various diet adequacies and supplementations can decrease ammonia production in response to exercise (Bassini-Cameron et al., 2008; Carvalho-Peixoto et al., 2008). This decrease in ammonemia is mainly due to decreased myokinase demand, which is in turn is due to an increase in ATP production from other metabolic sources (Almeida and Cabral, 2010). In both trials, ammonia increased after exercise bouts and returned to basal levels during rest and recuperation. To reinforce these analyses, we were able to show that urea increased in trial A, but we saw only a small increase in trial B. Urate levels also showed a smaller increase in trial B compared with trial A. Because urea is the final product of ammonia metabolism and urate is the final product of purine metabolism, it is reasonable to propose that the intervention could promote a metabolic adaptation that decreased the amount of ATP generated by myokinase by the use of amino acids.
Our data suggest that the intervention abolished the potassium response to exercise observed in trial A. Phosphate levels were also moderated by the intervention. These minerals participate in many metabolic processes, including muscle contraction, normal heart rhythm, nerve impulse conduction, oxygen transport, oxidative phosphorylation, enzyme activation, immune functions, antioxidant activity, bone health, and regulating blood pH. An adequate amount of minerals is necessary for optimal functioning of processes that are accelerated during exercise (Dressendorfer et al., 2002; Speich et al., 2001; Williams, 2005).
In this study, we evaluated metabolic adaptation in a world champion windsurfer in two different training situations: the first when the athlete was using the usual training and dietary protocols, and the second after a careful analysis of his diet and in-field metabolic changes as measured in a simulated competition. To summarize, we propose that, in response to the dietetic and training intervention, BCAA consumption was decreased during the second race and AAA, Ala, Glu, and Gln blood consumption was protected. It seems that the intervention also promoted a decrease in ammonemia, uricemia, and uremia in response to exercise. As a corollary to the diminished metabolic stress, inflammatory and muscle injury markers also decreased, which could show that the intervention had a positive effect on the athlete's response to exercise.
During the last decade in our laboratory, we have used exercise together with dietary modifications to cause metabolic stress in our studies of metabolism. Our studies are conducted in the field to mimic the real challenges faced during sports situations. Due to the uniqueness of this approach and the differences between it and standard experimental protocols, we call this approach “sportomics.” In this sense, sportomics is the use of “-omics” sciences together with classical clinical laboratory analysis (e.g., enzymatic determinations, ELISA, Western blotting, and other analytical procedures) to understand sport-induced modifications. Due to the uniqueness of this experiment, these results may not apply to other windsurfers, but we nonetheless had the opportunity to characterize the metabolic adaptations of this athlete. We also propose the importance of in-field metabolic analyses in the understanding, support, and training of elite athletes.
Footnotes
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.