
Abstract
Abstract
Surplus accumulation of regulatory T cells (Tregs) is known to be at the bottom of many morbid conditions, among them being neuropsychiatric diseases. In particular, Tregs may inhibit Th1 cells, including brain autoimmune lymphocytes, controlling the local microglial response and brain tissue homeostasis. The present study was undertaken in an attempt to suggest a novel approach for the treatment of maladaptation to mental stress associated with excessive Treg accumulation. Recently it was shown that alkylating drugs (ADs), such as melphalan and cyclophosphamide (Cy) in the dose 100-fold lower than cytostatic one are capable to disturb signal transduction by IL-2R. In this study we demonstrated that IL-2R is not a unique receptor, which may be blocked with ADs. Similar effect has been shown for two other surface receptors: TNFR and Fas. Molecular mechanisms of the receptor blockage were investigated on the model of TNF signaling. Study of NF-κB activity in nuclear extracts showed that alkylating agents act at the level of surface receptor or of the receptor platform. It was also shown that ADs administration in ultralow doses results in selective elimination of Tregs. In this study we used a new laboratory model of Treg accumulation in mice. Such Treg accumulation was associated with cognitive and behavioral abnormalities, which may be prevented by Cy administration.
Introduction
In this study we demonstrated that IL-2R is not a unique receptor, which may be blocked with ADs. Similar effect has been shown for at least two other surface receptors: TNFR and Fas. Recently it was shown that short-term simultaneous administration of dexamethasone (Dx) and IL-2 markedly expended functional suppressive FoxP3+CD4+CD25+ Tregs in murine peripheral lymphoid tissue including spleen, inguinal, and mesenteric lymph nodes (Chen et al., 2006). On the basis of these facts we created a laboratory model of Treg accumulation in mice. We believe that similar Treg accumulation takes place in the course of stress development. Using this animal model we also demonstrated that cognitive function impaired by superfluous Treg accumulation may be restored after cell surface receptor blockage.
Materials and Methods
Alkylating agents
Cyclophosphamide (Endoxan®, ASTA Medica, Frankfurt, Germany) and melphalan (Alkeran®, The Wellcome Foundation Ltd, London, UK) were used in the experiments. Cyclophosphamide (Cy) was applied in animal experiments and melphalan, which did not require metabolic activation by liver enzymatic systems, was used in the experiments with cell cultures.
Cell lines and reagents
L929 and HepG2 murine cell lines were obtained from Research Centre for Medical Genetics cell collection. The cells were seeded in 96-well plates (18×103 cells per well) in complete medium (RPMI/DMEM supplemented with 10% heat-inactivated donor horse serum, 2 mM L-glutamine, 20 μg/mL gentamycin) and incubated until monolayer formation. Then the cells were treated with actinomycin D (10 μg/mL) and melphalan (0.1–1 μg/mL) and exposed to tumor necrosis factor-alpha (TNF-α) or FasL for 18 h. Recombinant TNF-α and FasL (Bender Med Systems, San Diego, CA, USA) were used within the dose ranges 16–64 pg/mL and 6–12 ng/mL, respectively. Quadriplicate wells of each condition were assayed. For the purpose of cell viability assessment the cells were stained with 0.2% crystal violet and, then the plates were washed, dried, and read at 594 nm.
Analysis of NFκB activation
To determine whether melphalan treatment affects the activation of transcriptional factor NFκB, nuclear translocation of NFκB was examined with TransAM NFκB kit (Active Motif, Inc., Carlsbad, CA, USA). A pool of L929 cells was preincubated for 1 h with actinomycin D (10 μg/mL). Subsequently, cells were treated for another hour with melphalan (0.3 μg/mL) and then exposed to TNF-α (64 pg/mL) for 15 min. Nuclear extracts were prepared and tested for NFκB p65 according to the manufacturer's recommendation.
Animals
Male BALB/c mice at the age of 8–10 weeks were purchased from Nursery “Pushchino” (Inst. Bioorganic Chemistry, Russian Academy of Science). All the animals were kept in individual cages and had free access to food and water. The animals were housed at light- and temperature-controlled room. Group size is varied from 17 to 19 animals (see Table 1). All experimental procedures were approved by Ethics Committee of the Research Centre for Medical Genetics.
(1) Control animals. (2) Mice pretreated with Cy. (3) Mice injected with Dx/IL-2. (4) Mice injected with Dx/IL-2 after Cy pretreatment. The data are presented as: number of animals revealing the platform/total amount of animals in the group. p-Values were calculated using nonparametric Wilcoxon paired test.
On day 4 the task was complicated: the platform was placed at the opposite location.
Animal treatment
Mice were injected intraperitoneally with Cy at the dose of 15 mg/kg of body weight during 9 weeks (1 injection per week) until the start of the experiment. For Treg accumulation mice were injected with Dx (5 mg/kg) and recombinant IL-2 (100,000 IU per mouse).
Morris water maze (MWM) behavioral test
Spatial learning/memory was assessed by performance on a hippocampal-dependent visuospatial learning task in MWM. Mice were given four trials per day, for 4 consecutive days, to find a hidden platform located 1.5 cm below the water surface in a pool (60×50×40 cm). Within the testing room, only distal visuospatial cues were available to the mice for the location of the submerged platform. The escape latency, that is, the time required by the mouse to find and climb onto the platform, was recorded for up to 60 s. Then the mouse was moved from the maze to its home cage. If the mouse did not find the platform within 60 s, it was manually placed on the platform and returned to its home cage after 30 s. On day 4 the task was complicated; the platform was placed at the opposite location. Data was recorded by video filming.
Statistical analysis
Data were analyzed using SYSTAT SigmaStat statistical software. The effect of melphalan treatment on FasL- or TNF-α-induced cytotoxicity was evaluated using one-way analysis of variance (ANOVA). When significant differences were found, a Dunnett's post hoc test was used to identify group differences. For investigation differences between the animal groups in MWM Behavioral Test, statistical analysis was performed using nonparametric Wilcoxon paired-sample tests. Significance was accepted at a p-value <0.05.
Results
Noncytotoxic concentrations of alkylating agents disturb TNF-α and FasL signaling in the cells
As can be seen in Figure 1, relatively low concentrations of melphalan (0.1 and 0.03 μg/mL) protected murine fibroblastoid cells (L929) against TNF-α-induced cytotoxicity. In the case of further dose decrease (0.01 μg/mL) the protective effect of melphalan was weaker but revealed sufficiently clear. The similar results have been obtained for FasL. Thus, HepG2 cells treated with melphalan in different concentrations became relatively resistant to cytotoxic effect of FasL (Fig. 1B).
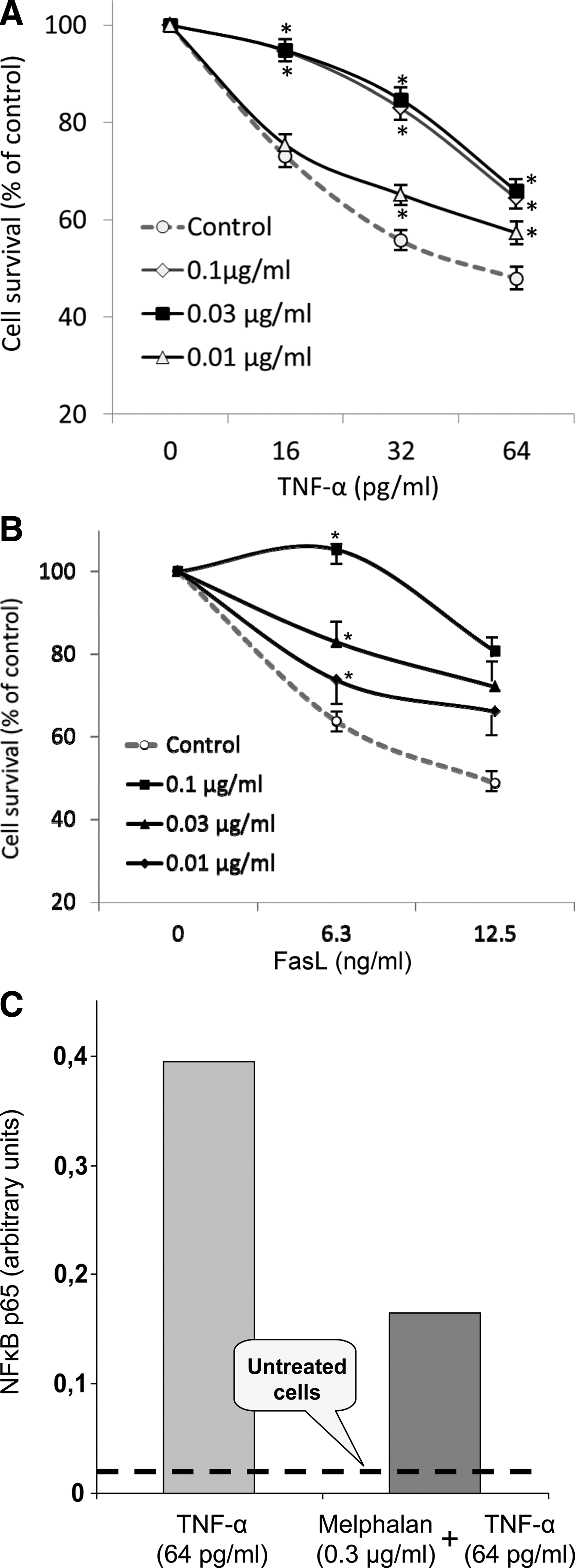
Influence of melphalan on TNF-α and FasL signaling. (
At the same time, 1-h treatment with melphalan markedly reduced the transcription factor activity in nuclear extracts of actinomycin D pretreated cells challenged with TNF-α (Fig. 1C). These data support the suggestion that specific alkylation of components in the cytoplasm or cell membrane by melphalan interferes with TNF-α signaling pathway.
Spatial learning/memory assessment
The scheme of experiment is presented at Figure 2. Cognitive abnormalities have been shown in mice treated with Dx/IL-2. The mice took longer to find the hidden platform compared with control animals. As can be seen in Figure 3, most dots of Dx/IL-2 curve are located above the control ones. According to Wilcoxon paired-sample test such differences have a tendency to be significant (p=0.06). However, the p-value was equal to 0.01 when we excluded all first trials in order that we might detect the learning capability in each of 4 days.

The scheme of murine experiments. BALB/c mice were divided into two groups: control animals and animals injected with Cy during 9 weeks (1 injection per week). Then half animals of each group were simultaneously injected with Dx and IL-2; the other half was injected with saline.

Cognitive abnormalities in mice treated with Dx/IL-2. Spatial learning/memory was assessed in the Morris Water Maze Behavioral Test. Mice were given four trials per day to find a hidden platform located 1.5 cm below the water surface. The time required by the mouse to find and climb onto platform (latency period) was evaluated. According to nonparametric Wilcoxon paired-sample test, p values are 0.06 and 0.01, after exclusion of all first trials (see text).
In addition we succeeded in demonstration that such cognitive disorders were significantly weaker in the mice pretreated with Cy. Although these mice looked for a platform contemporaneously as the Dx/IL-2-treated animals, percentage of the mice successfully revealing the platform was significantly higher (Table 1 and Fig. 4). As can be seen in Figures 3 and 4, untreated animals learned to find a platform within 1 day. On day 4 the task was complicated: the platform was placed at the opposite location. Such alteration of conditions resulted in decrease of percentage of mice finding the platform. Dx/IL-2 treated mice looked for a platform significantly longer and the percentage of animals successfully coped with the task was lower (p<0.001). However, the results of Cy pretreated mice did not significantly differ from those of intact control and were appreciably better in comparison with the mice treated with Dx/IL-2 only (p<0.05).
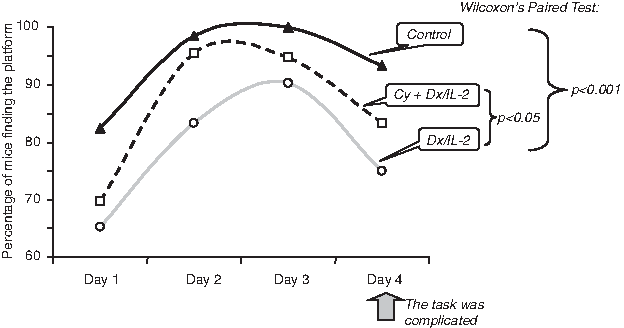
Dx/IL-2-induced cognitive disorders in mice pretreated with Cy. The data are presented as a percentage of animals revealing the platform. On day 4 the task was complicated: the platform was placed at opposite location. p Values were calculated using nonparametric Wilcoxon paired-sample test.
Discussion
Our data show that IL-2R is not a unique receptor, which may be affected by ADs. There are two other surface receptors, TNFR1 and Fas, that may be blocked by ultralow concentrations of the drugs. TNFR1 and Fas (CD95, APO-1) belong to the family of death receptors and transduce signals resulting in entirely different biological outcomes: they promote cell death via apoptosis but are also capable of inducing antiapoptotic signals through the activation of NF-κB or MAPK/ERK cascade resulting in cell protection and tissue regeneration (Schütze and Schneder-Brachert, 2009). In the case of TNFR1, ligand-dependent or -independent activation results in the recruitment of TRADD, which then becomes a platform for binding of additional cytoplasmic adaptor proteins. Therefore, two sorts of signalling cascades may be switched on. The first is implicated in increasing the transcriptional activity of antiapoptotic NF-κB. The second results in activation of caspase-8 and apoptotic machinery (Sheikh and Huang, 2003). In normal cell antiapoptotic pathways lead to antiapoptosis protein synthesis and prevail over the pro-apoptotic signals (Fig. 5A). However, inhibition of NF-κB-dependent signals by the transcription or protein synthesis inhibitors may tip a balance in favor of TNFR1-activated apoptotic signals. Thus, in our experiments combined treatment with actinomycin D and TNF-α induced both the death of L-929 cells in dose-dependent manner and pronounced NF-κB activation (see Figs. 1A and C). If the cells were preincubated with melphalan a marked decrease of NF-κB activity and a significant increase of L-929 cell survival have been observed. Such simultaneous inhibition of NF-κB activity and apoptosis development indicates that alkylating agent melphalan acts at the level of surface receptor or of the receptor platform. The signal cannot penetrate into the cell and both pro-apoptotic and antiapoptotic pathways have been switched off.

Tentative mechanism of protective effect of alkylating agents against TNF-α-induced cytotoxicity. (
In this work we succeeded in demonstration that prolonged treatment with Cy in the form of pulse-therapy attenuates the signs of cognitive disorders in Dx/IL-2 injected mice. As it was shown in mice, the short-term simultaneous administration of Dx and IL-2 results in a stress-like condition, which is attended by cognitive disorders. Mechanisms of such cognitive disorder development are most probably associated with Th1 cell inhibition by Tregs in the brain. The data by which severe conbined immunodeficient (SCID) mice and the mice with T-cell deficiency demonstrate similar cognitive disorders confirm this viewpoint. Moreover, normal T-cell transplantation restores their learning capability (Brynskikh et al., 2008; Cohen et al., 2006). It seems the favorable effect of Cy is associated with prevention of surplus Treg accumulation, although it is not a single mechanism of alkylating drug action. It was recently shown that Cy is not only capable of Treg elimination but also resets dendritic cell (DC) homeostasis (Radojcic et. al., 2010; Salem et. al., 2010a). In other words, alkylating metabolites of Cy find their cellular targets both in periphery and in bone marrow (Salem et al., 2010a, 2010b). Thus, in bone marrow Cy mobilizes DC progenitors, which are translocated into peripheral tissues and differentiated there in mature DCs, which demonstrate high pro-inflammatory activity including intensive production of IL-12 (Radojcic et. al., 2010). It is known that IL-12 plays an important role in promoting the differentiation of naïve T cells to Th1 cells. Glial cells supported by Th1 cells begin to clean the brain from waste products of cells. As a result the neuron function is restored.
Conclusion
The results reported in this study suggest that low noncytotoxic concentrations of alkylating agents have an effect upon cell at the level of surface receptors or recruited proteins. Although such influence is not a result of specific binding between the drug and a target molecule, the final biological effect has a selective mode. The selectivity depends on relatively low dose of the drug, which is insufficient for essential cell functions alteration, and on target molecule repertoire (repertoire of surface receptors). Now we can confidently consider that the capability of ADs to eliminate Tregs is associated with IL-2R blockage. The mechanisms of the interference of ADS in DC homeostasis and the role in this process of Fas and TNFR are not so clear.
Footnotes
Acknowledgments
This work was supported by Russian Foundation for Basic Research; Grant No 10-04-01342a.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.