
Abstract
Abstract
Soil salinity is one of the major abiotic stress limiting crop productivity and the geographical distribution of many important crops worldwide. To gain a better understanding of the salinity stress responses at physiological and molecular level in cultivated tomato (Solanum lycopersicum. cv. Supermarmande), we carried out a comparative physiological and proteomic analysis. The tomato seedlings were cultivated using a hydroponic system in the controlled environment growth chamber. The salt stress (NaCl) was applied (0, 50, 100, 150 and 200 mM), and maintained for 14 days. Salt treatment induced a plant growth reduction estimated as fresh-dry weight. Photosynthetic pigments (chlorophyll a, b) content of NaCl-treated tomato plants was significantly decreased as the salinity level increased. Proline accumulation levels in leaf and root tissues increased significantly with increasing NaCl concentration. Relative electrolyte leakage known as an indicator of membrane damage caused by salt stress was increased proportionally according to the NaCl concentrations. Roots of control and salt-stressed plants were also sampled for phenol protein extraction. Proteins were separated by two-dimensional gel electrophoresis (2-DGE). Several proteins showed up- and downregulation during salt stress. MALDI-TOF/MS analysis and database searching of some of the identified proteins indicated that the proteins are known to be in a wide range of physiological processes, that is, energy metabolism, ROS (reactive oxygen species) scavenging and detoxification, protein translation, processing and degradation, signal transduction, hormone and amino acid metabolism, and cell wall modifications. All proteins might work cooperatively to reestablish cellular homeostasis under salt stress, water deficiency, and ionic toxicity.
Introduction
The plant response to salt stress may be studied by comparative proteomic research, because the proteome, in contrast to the genome, may markedly change as a result of environmental factors (Williams and Hochstrasser, 1997). In salt resistance, specific changes in the profile of proteins, whose biological functions are related to such environmental stress tolerance, have been observed in many plants (Abbasi and Komatsu, 2004; Kav et al., 2004), and several proteins have been characterized to play prominent roles in response to salt stress. However, changes in protein profiles also differ depending on plant organs and species (Jiang et al., 2007, Razavizadeh et al., 2009). The proteomic approach, based on 2-DE and MS analysis, may offer the possibility to identify proteins associated with a particular environmental and/or developmental signal (Chen and Harmon, 2006; Jiang et al., 2007; Ndimba et al., 2005). The aim of the present research was to determine physiological responses of tomato plants (Solanum lycopersicum. cv. Supermarmande), exposed to salt stress and to identify the changes in the levels of root protein expression, by comparative proteomics between control and salt-treated plants, in order to increase the data set of proteins involved in plant tolerance to salinity.
Materials and Methods
Plant materials and stress treatment
Tomato seeds (Solanum lycopersicum var. Supermarmande) were surface-sterilized by soaking in the 5% (v/v) sodium hypochlorite solution for 15 min followed by three washes with sterile distilled water. Then seeds were germinated in Petri dishes with moistened filter paper and were incubated in the culture room under fluorescent light (90 mmolm−2 s−1 with a 16-h photoperiod) at 25°C. The obtained seedlings were transferred to continuously aerated hydroponic nutrient solution containing: KNO3 3 mM, Ca(NO3)2 1 mM, KH2PO4 2 mM, MgSO4 0.5 mM, Fe-K-EDTA 32.9 μM, and micronutrients: H3BO4 30 μM, MnSO4 5 μM, CuSO4 1 μM, ZnSO4 1 μM, (NH4)6Mo7O24 1 μM. Culture was conducted in a growth chamber: 25°C/70% relative humidity during the day and 20°C/90% relative humidity during the night; photoperiod: 16 h daily with a light irradiance of 150 mmolm−2 s−1. Plants were grown for 14 days in control medium, then given NaCl stress (50, 100, 150, and 200 mM). Salt stress was maintained for 2 weeks. Roots harvested from control or NaCl-treated plants under 100 mM NaCl, washed with distilled wafter at least five times, and immediately frozen in liquid nitrogen, and stored at −80°C for protein extraction.
Physiological analyses
Control and NaCl-treated plants, growing under 200 mM NaCl, were harvested and separated into leaves, stems, and roots. Fresh weight was measured and dry weight was determined after desiccation at 80°C for 48 h. Estimation of chlorophyll pigment contents was achieved by application of the method of Torrecillas et al. (1984): 80% acetone extract was colorimetric all assayed at 649 and 665 nm. For membrane permeability (solute leakage), electrolyte leakage was determined as described by Dionisio-Sese and Tobita (1998). Leaf discs of fresh seedlings were cut into 2–3-mm pieces and placed in test tubes containing 10 mL distilled water. The tubes were incubated in a water bath at 32°C for 2 h and the initial electrical conductivity of the medium (EC1) was measured using a conductivity meter (CDM92, Conductivity Meter). The samples were autoclaved at 121°C for 20 min to release all electrolytes, cooled to 25°C, and the final electrical conductivity (EC2) was measured. The method is as followed: EL=EC1/EC2×100.
The proline content was determined using the method of Bates et al. (1973). Proline was extracted from organs samples of 200 mg fresh weight (FW) with 10 mL of 3% sulphosalycylic acid at 70°C for 30 min. After addition of acid ninhydrin and glacial acetic acid to the extracts, the mixture was heated at 90°C for 1 h in water bath. Reaction was then stopped by using an ice bath. The mixture was extracted with toluene and the absorbance of fraction with toluene aspired from the liquid phase was spectrophotometrically determined at 520 nm. Proline concentration was determined using calibration curve as μmol proline g−1 FW.
Preparation of total protein extracts
Control and NaCl-treated root were ground to a fine powder in liquid nitrogen and stored at −80°C. For each sample, four independent protein extractions were performed as previously described (Rocco et al., 2006). Root powder was suspended in 3×vol of an extraction buffer containing [700 mM sucrose, 500 mM Tris (pH 8), 100 mM KCl, 2% (v/v) β-mercaptoethanol, 2 mM PMSF, pH 8.5] and incubated for 10 min on ice. Afterward, an equal volume of Tris-saturated phenol was added. The samples were shaken for 10 min at room temperature and then centrifuged (10 min, 5,525×g, 4°C) to separate the phenolic and the aqueous phase. The phenolic phase was recovered and reextracted with the same volume of extraction buffer. Subsequently, centrifugation was repeated and 5 vol of precipitation solution (0.1 M ammonium acetate in methanol) was added to the recovered phenol phase. Proteins were allowed to precipitate at −20°C overnight. After centrifugation (10 min, 5,525×g, 4°C), the protein pellet was washed three times with the precipitation solution and once with acetone. Each washing step was followed by 5 min of centrifugation as described above. After drying under vacuum, the pellet was resuspended in lysis buffer [9 M Urea, 4% (w/v) CHAPS, 0.5 % (v/v) Triton X-100, 20 mM DTT, 1.2% (v/v) pharmalytes pH 3–10], and protein concentration was measured according to a modified Bradford assay (Ramagli and Rodriguez, 1985).
Two-dimensional electrophoresis
Proteins were first separated according to their charge after passive rehydration of 24 cm-long Immobiline dry strips, pH 4–7 (Amersham Bioscience, Uppsala, Sweden) with 500 μg of resuspended proteins, 9 μL IPG buffer pH 4–7 and rehydration buffer [8 M Urea, 2% (w/v) CHAPS, 0.3 % (w/v) DTT, 2% (v/v) pharmalytes pH 3–10], to a final volume of 450 μL. Isoelectric focusing was performed with the Multiphor II (Amersham Bioscience, Uppsala, Sweden) according to the following program: 2 h at 150 V, 2 h at 400 V, 2 h to increase the voltage from 400 V to 3,500 V, 25 h at 3,500 V. After migration, IEF strips were stored at −80°C or immediately incubated in equilibration buffer [6 M Urea, 50 mM Tris-HCl, pH 8.8, 30% glycerol, 2% (w/v) SDS, with addition of 2% (w/v) DTT in the first equilibration step, and 2.5% (w/v) iodoacetamide in the second equilibration step, respectively] for 20 min.
SDS-PAGE was carried out with 11% acrylamide gels in the Biorad Protean Plus Dodeca cell electrophoresis chamber (2 h at 70 V, 15 h at 100 V).
Four 2-DE gels per sample were fixed in 2% phosphoric acid–50% ethanol, washed in 2% phosphoric acid, and stained 3 days in 2% phosphoric acid–15% ammonium sulfate–17% ethanol and 0.1% Coomassie Brilliant Blue G-250. This staining allowed a reproducible quantification because the relation between protein abundance and optical density of the spots is linear. Protein spot detection and quantification were obtained using the image-analysis software Progenesis SameSpots v3.0 (Nonlinear Dynamics Ltd, Japan).
Protein identification by mass spectrometry
In-gel digestion was performed with the Progest system (Genomic Solution) according to a standard trypsin protocol. Gel pieces were washed twice in baths of 10% acetic acid, 40% ethanol, and acetonitrile (ACN). They were then washed twice in baths of 25 mM NH4CO3 and ACN. Digestion was subsequently performed for 6 h at 37°C with 125 ng of modified trypsin (Promega, Madison, WI) dissolved in 20% methanol and 20 mM NH4CO3. The peptides were extracted successively with 2% trifluoroacetic acid (TFA) and 50% ACN and then with ACN. Peptide extracts were dried in a vacuum centrifuge and suspended in 20 μL of 0.05% TFA, 0.05% HCOOH, and 2% ACN. High-performance liquid chromatography (HPLC) was performed on an Ultimate LC system combined with a Famos autosampler and a Switchos II microcolumn switch system (Dionex, Sunnyvale, CA). A 4-μL sample was loaded at 5 μL/min−1 on a precolumn cartridge (stationary phase: C18 PepMap 100, 5 μm; column: 300 μm i.d., 5 mm; Dionex) and desalted with 0.05% TFA, 0.05% HCOOH, and 2% ACN. After 2.5 min, the precolumn cartridge was connected to the separating PepMap C18 column (stationary phase: C18 PepMap 100, 3 μm; column: 75 μm i.d., 150 mm; Dionex). Buffers were 0.1% HCOOH, 3% ACN (A) and 0.1% HCOOH and 95% ACN (B). The peptide separation was achieved with a linear gradient from 5 to 30% B for 25 min at 200 nL/min−1. Including the regeneration step at 100% B and the equilibration step at 100% A, one run took 45 min.
Peptide ions were analyzed using Xcalibur 1.4 with the following data-dependent acquisition steps: (1) full MS scan [mass-to-charge ratio (m/z) 400 to 1900, centroid mode], (2) ZoomScan on a selected precursor (scan at high resolution in profile mode on an m/z window of 4), and (3) MS/MS (qz=0.22, activation time=50 ms, and collision energy=40%; centroid mode). Steps 2 and 3 were repeated for the two major ions detected in step 1. Dynamic exclusion was set to 30 s.
Statistical analysis
The statistical analyses were performed with the Statistica software (version 6.0). All physiological parameters mean values and standard error (SE) were obtained from of at least 10 to 20 replicates and analyzed using Duncan's multiple range test. A p-value of <0.05 was considered to be statistically significant. For gel image analysis, Samespots software was used to detect varying spots using one-way analysis of variance (ANOVA) on normalized spot volume from the four gel repeats with p<0.01.
Results and Discussion
Physiological responses to NaCl-stress in tomato
The present results showed that salt stress treatment (200 mM NaCl) significantly decreased the whole plant growth (dry and fresh weight production) of the tomato plant. The fresh and dry mass was decreased with salt treatment. Reduction value of FW and dry weight (DW), expressed as the ratio of the treatment to the control, ranged respectively by 67% and 64% (Fig. 1). The same results were observed in tomato cultivars (Juan et al., 2005; Maggio et al., 2007). As known, the excess foliar accumulation of toxic (Na+ and Cl−) inhibits plant growth and development (Munns et al., 2002). One of the most widely used agricultural indices to define stress tolerance is data for plant biomass and yield (Sairam et al., 2002). Decreased pigment contents due to salinity have previously been reported, with chlorophyll and carotenoid contents being suggested as one of the parameters of salt tolerance in crop plants (Juan et al., 2005; Sairam et al., 2002), prompting their wide use in screening programs. Here we found a decrease in pigment content in different parts of the tomato plant. Both chlorophyll a and b was gradually decreased with increasing external salinity in the tomato (Fig. 2).
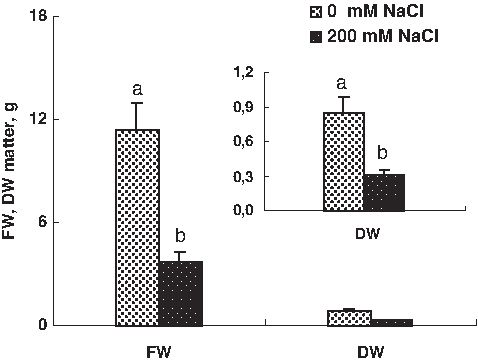
Effect of salinity on seedling growth. Dry and fresh weight comparison of control plants and plants submitted to salinity (200 mM of NaCl) in whole plants. Data are means of 32 replicates±SE at p<0.05. Letters correspond to Duncan's multiple range test at 95%.
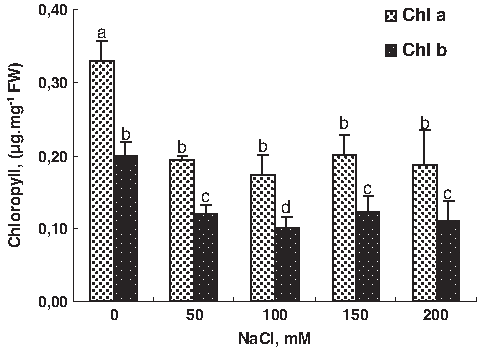
Effect of NaCl treatment (0, 50, 100, 150, and 200 mM) on pigment chlorophyll content in tomato leaves. Data are means of 10 replicates±SE at p<0.05. Letters correspond to Duncan's multiple range test at 95%.
Among the compounds considered compatible, perhaps the most thoroughly studied in relation to the response to saline stress in plants is proline. Stewart and Lee (1974) suggested that proline might act as acyteoplasmic osmoticums counteracting the effect of salt accumulated in the vacuole. However, the role of the proline in osmoregulation and salt tolerance has generally been questioned. For example, higher proline accumulation was found in salt tolerant Brassica juncea plants with better growth than the control (Kirti et al., 1991).
Also, Juan et al. (2005) found that the most salt-tolerant cultivars, Brillante and Jaguar, registered the highest foliar concentrations in proline than the salt-sensitive cultivars.
In contrast, Tal et al. (1979) reported more proline accumulation in salt-sensitive species of tomato than in tolerant wild relatives. Aziz et al. (1998) reported a negative relationship between proline accumulation and salt tolerance in tomato plants. In the present study, under salt condition we found an accumulation of proline. This accumulation was gradually increased with increasing external salinity in tomato (Fig. 3).

The effect of NaCl treatments (0, 100, and 200 mM) on proline content in different parts of tomato plant (leaves, shoots, and roots). Data are means of 10 replicates±SE at p<0.05. Letters correspond to Duncan's multiple range test at 95%.
Relative electrolyte leakage (REL) is an indicator of membrane damage caused by NaCl stress (Cao et al., 2007). Previous studies found an accumulation of solute leakage and an increased REL in leaves of five cultivars of sugar beet (Ghoulam et al., 2002) and in leaves of a salt-sensitive poplar species (Populus x canescens) (Bolu and Polle, 2004). As shown in Figure 4, REL in tomato leaves was gradually increased with increasing external salinity.
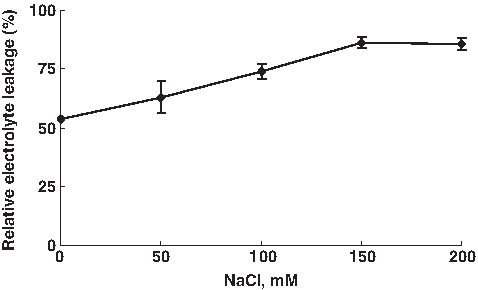
The effect of NaCl treatments (0, 100, and 200 mM) on electrolyte leakage from disc of tomato leaves. Data are means of 10 replicates±SE at p<0.05.
Proteomic analysis of root proteins in the salt-stressed tomato
In order to investigate the changes of root proteome in response to salt stress, 2-DGE analysis of the total proteins in tomato roots from three or four biologically independent replicate experiments was carried out. Figure 5A shows the reference Coomassie brilliant blue-stained gels obtained from control and Figure 5B from salt stress plants (100 mM NaCl). The protein spots showed a broad distribution in the pI range from 4.0 to 7.0 and the mass range from 10 to 120 kDa. Software image analysis revealed a total of 60 protein spots that changed their abundance (vol %) significantly (p<0.05) by <1.5-fold in average normalized volume according to some previous proteomics studies (Jellouli et al., 2008; Jiang et al., 2007). At least 33 spots showed significant changes under salt stress; 27 proteins were upregulated and 19 proteins were downregulated in salt-stressed tomato roots. In this article we represent only 23 differentially expressed proteins.

Root proteome patterns under control condition (
LC-MS/MS identification and classification of differentially expressed proteins
We selected 60 statistically confirmed proteins spots; all of them were excised from gels for tryptic digestion and analysis by LC-MS/MS to determine protein identity. The database was elaborated from the Solanaceae Genomics Network available at the Web address http://www.sgn.cornell.edu and identified proteins were classified according to the Funcat automatic classification (http://mips.gsf.de/proj/funcatDB). In total, it was possible to obtain good-quality protein identification results with a high probability score for 51 out of the 60 protein spots.
Salt stress-responsive proteins identified
Table 1 shows that of the 23 differentially expressed and identified proteins, 6 of them were downregulated and 14 proteins were upregulated in tomato roots under salt treatment. Exposure of plants to salt stress increase the production of reactive oxygen species (ROS) such as singlet oxygen (O21), superoxide radical (O2−), hydrogen peroxide (H2O2) and hydroxyl radical (OH−), and these ROS cause oxidative damage to different cellular components including membrane lipids, protein, and nucleic acids (Apel and Hirt, 2004).
Number of polypeptide matched.
Sequence coverage percentage.
Salt effect; (–) indicates the downregulation of protein, (+) indicates the upregulation of protein.
Recent published articles revealed that the balance of ROS formation and removal is a determinant factor for the severity of oxidative stress and cell damage (Apel and Hirt, 2004; Sahi et al., 2006). One of the antioxidant enzymes is peroxidase, which can detoxify H2O2 by oxidizing specific substrates such as ascorbate. Peroxidases have been found to be upregulated by salt stress in many plants (Hernandez et al., 2000). Glutathione, as well as ascorbate, can remove many free radicals through a series of oxidation/reduction, and the glutathione-based detoxification is considered as a very important salt adaptation mechanism in plant (Apel and Hirt, 2004; Askari et al., 2006). Moreover, overexpression of a cDNA encoding an enzyme with the dual activities of glutathione S-transferase (GST) and glutathione peroxidase (GPX) in tobacco increased the GST and GPX activities and resulted in higher salt-stress tolerance (Roxas et al., 2000). In our study, var. supermarmande represents significantly upregulation of ascorbate peroxidase (spot 17), GST (spot 16), and GPX (spot 20) under salt treatment. Similar results were obtained by Mittova et al. (2004) in the case of the wild salt-tolerant tomato species Solanum pennellii. However, two spots (spot 22 and 9), corresponding to the same cell wall peroxidase protein, were downregulated after salt stress. These data are in opposition to previous studies that demonstrated the upregulation of peroxidase by salt stress in rice root (Yan et al., 2005) and in Arabidopsis root (Jiang et al., 2007) roots. Such a pattern of variations suggested that, in sensitive genotype, a prolonged oxidative stress occurred, whereas in tolerant genotype, it was more rapidly overcome. Furthermore, we found induction of Thioredoxin H-type 1 (spot 19) under salt treatment. In some studies we demonstrate that TRX-H-1 proteins were associated with oxidative stress. It serves as a major defense system against oxidative salt stress in Schizosaccharomyces pombe (Song and Roe, 2008), and could contribute by reducing disulfide bonds of oxidized proteins, and by supporting oxidant-scavenging proteins such as thioredoxin-dependent peroxidase Tpx1.
On the other hand, plants have evolved a complex sensory mechanism to monitor their surroundings and to adapt their growth and development to the prevailing environmental conditions. Salt treatment can interrupt several physiological and biochemical processes in cells. Therefore, a wide range of stress-related proteins have been identified. In our study, four heat-shock proteins were identified; Class II small heat-shock protein Hsp17.6 (spot 3), heat-shock Hsp70 protein (spot 8), Hsp90 protein (spot 7). Their upregulation that they play a crucial role in preventing aggregation of the denatured proteins and in facilitating the refolding under salt stress similarly to that was found in previous studies (Chang et al., 2000; Frydman et al., 1994, Lindquist and Craig, 1988; Wang et al., 2009). In addition, sHsps are involved in many developmental processes, such as embryo development, seed germination, somatic embryogenesis, pollen development, and fruit maturation (Waters et al., 1996). In this investigation, small heat-shock proteins (sHsp 26.5) were downregulated dramatically during salt treatment. In contrast, the other Hsps (Hsp70, Hsp90, and Hsp17.6) were increased in the salt-stressed root. Previous studies showed that Hsps were upregulated in stressed plants under salt stress. Hamilton and Heckathorn (2001) demonstrated that in plants, mitochondrial sHsps may protect cellular proteins through a mechanism involving ROS scavenging. It is not clear whether the expression of sHsp genes during oxidative stress is induced by H2O2, which may function as a signaling molecule, or by the damaged proteins. Probably, both mechanisms are involved (Volkov et al., 2006). Downregulation of sHSPs such as sHsp26.5 would probably cause impairments in protein structure and function (Timperio et al., 2008).
In higher plants, glycolysis is a central pathway for energy production. At the same time, the activity of enzymes for the citric acid cycle is controlled by the energy status of the cell. Several glycolysis and carbohydrate metabolism-associated proteins were changed following salt and osmotic stress treatments in several plant species (Jiang and Deyholos, 2006; Ndimba et al., 2005). It is interesting that one of these energy metabolism related proteins Vacuolar ATPase subunit B (spot2) was significantly downregulated in the tomato root under salt conditions. In contrast, ATP synthase F1 subunit 1 (spot 15) increased in NaCl-treated roots. Little is known on the response of expression of ATPase subunit genes against environmental stresses. Sweetlove et al. (2002) exposed Arabidopsis cells to oxidative stresses, and found that the ATP synthase a-subunit and b-subunit were significantly decreased by the stress treatments. The authors speculated that the ATP synthase subunits are the greatest casualties of oxidative stress.
Enolase (spot 11), known as 2-phospho-D-glycerate hydrolases, was classically known to catalyze the dehydration of 2-phosphoglycerate to phosphoenolpyruvate. Enolase is one of the most important enzymes in glycolysis. It is responsive to many environmental stresses, including salt stress, drought, cold, and anaerobic stress in different plant species (Riccardi et al., 1998, Yan et al., 2006). In our experiment, enolase proteins was induced in by salt stress in the tomato root.
The abundance of others proteins involved in glycolysis, citrate circle, and metabolism like; UDP-glucose pyrophosphorylase (spot 12) Malate dehydrogenase cytosolic (cMDH,spot 13), Pyruvate dehydrogenase (spot 18), were strongly increased by salt stress on the root. Malate dehydrogenase (MDH) catalyzes the reversible reaction of oxaloacetate (OAA) to malate. Involved in malic acid concentration regulation (Miller et al., 1998), MDH was reported to be responsive to salinity stress in the root and suspension cell of Arabidopsis (Jiang et al., 2007; Ndimba et al., 2005), and pea root as well (Kav et al., 2004). In addition, cytosolic malate dehydrogenase was upregulated by long-term salinity stress in Thelluniella halophila leaves.
In the present work cMDH was significantly upregulated following salt treatment in the tomato root. All data suggested that MDH has a crucial biological function in plant metabolism in various tissues and species. Hyperosmotic stresses have been shown to cause changes in the pool of various free amino acids in plant cells (Martino et al., 2003). It was observed that the abundance of amino acid biosynthesis-related enzymes was influenced by NaCl. Glutamine synthetase GS58 (SGN-U314517, spot 10) decreased in abundance following NaCl treatment on the root of the tomato. Glutamine synthetase (GS) is reported to play a pivotal role in nitrogen assimilation through reassimilation of NH4 from photorespiration and proteolysis processes in plants subjected to salt stress and water deficiency (Tsai et al., 2002). Also, it was essential to synthesize the precursors of proline, which can serve as an osmolyte (Silveira et al., 2003). Yan et al. (2005) demonstrated that downregulation of GS protein and consequently reduced GS activity in rice may result in less proline production, which might be part of the reason for its salt sensitivity. These data, coupled with our studies, suggest that GS might be a determinant component for salt stress tolerance in different plant species.
Finally, we identified a transcription factor BTF3 (spot 14), which was upregulated in stressed root. This protein plays a central role in the elongation step of protein biosynthesis or cell-cycle regulation (Freire et al., 2005). The induction of this proteins under stress was studied previously, and we found that BTF3-like transcription factor activity was downregulated in the tomato root in response to waterlogging stress (Ahsan et al., 2007) but increased in the Arabidopsis root following NaCl treatment (Jiang and Deyholos, 2006; Jiang et al., 2007). Downregulation or decreased activity stress suggests that normal protein biosynthesis, particularly transcription or translation, and cell cycle regulation in roots are highly disrupted or inhibited under stress conditions, which may reduce or halt protein synthesis, and ultimately reduce plant growth and development (Chang et al., 2000).
Conclusion
In this study, physiological and proteomic analysis was conducted with tomato plants subjected to NaCl treatment. Symptoms of stress, such as growth inhibition, proline accumulation, reduction in pigment content, and electrolyte leakage was detected. Using two-dimensional electrophoresis (2-DE) we identified 24 proteins that significantly upregulated or downregulated due to NaCl stress (Table 1). This study may reflect the fact that these identified proteins seems to be involved in diverse biochemical reactions imparting tolerance to NaCl stress in the tomato. Our findings reveal a series of changes at the protein level that may provide insight into the stress-tolerance nature of the tomato.
Footnotes
Acknowledgments
The authors acknowledge the Ministry of Higher Education and Scientific Research of Tunisia for financial support. Many thanks to Karine Leyre and Caroline Callot for technical assistance and Emmanuel Botton for taking care of the plants.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.