
Abstract
Abstract
In Tunisia, like in the other countries of the Mediterranean, tomato is ranked among the important vegetables in the economic sphere. Tunisia ranks as the first consumer of this vegetable in the world. However, tomatoes are exposed to multiple environmental stresses. In particular, salinity is the most stressful limiting factor to productivity. Salt tolerance of the tomato is susceptible to be ameliorated by genetic and physiologic ways. Salicylic acid (SA), a plant phenolic, is now considered as a hormone-like endogenous regulator, and its role in the defense mechanisms against biotic and abiotic stressors has been well documented. So, the aim of this study was to investigate the impact of exogenous application of SA (0.01 mM) on growth, nutritional behavior, and some metabolic parameters (total chlorophyll, soluble sugars, proline, and proteins) of tomato plants cv. Moneymaker exposed to NaCl (100 mM). Our results showed that the application of 0.01 mM SA to tomato plants via root drenching attenuate the depressive effect of salinity on plants. This amelioration results in stimulation of growth and development of plant. Under stress conditions, SA-treated plants exhibited more accumulation of photosynthetic pigments and K+ contents. Thus, SA induces an increase in soluble sugars in roots and leaves; also, we noted the increase of proteins only in roots. Overall, the adverse effects of salt stress tomato plants were alleviated by the exogenous application of SA at vegetative stage, which upregulated nutrition and the accumulation of some organic solutes and osmoprotectors such sugars, proline, and proteins. So salicylic acid can be greatly used to enhance salt tolerance of tomato plants.
Introduction
Materials and Methods
Plant material and growth conditions
Seeds of tomato (Solanum lycopersicum cv Moneymaker) were sterilized with 20% sodium hypochlorite solution for 10 min and rinsed with distilled water. Seeds were then germinated in Petri plates double lined with filter paper moistened with distilled water in the dark at 25°C. In 8 days the germinated seedlings were transferred to nutrient solution. The composition of this one was made of macroelements (mM): K+ 3.0; Ca2+ 3.5; Mg2+ 1.0; NO3− 8.0; SO42− 1.0; PO4H2− 1.5, and microelements (ppm): Fe 1.4; Mn 0.25; B 0.16; Cu 0.03; Zn 0.03, and Mo 0.01. The experiment was led in air-conditioned room, under artificial light (150 μmol·m−2·s−1; 16h photoperiod), at 25°C day/20°C night and air humidity 60–80%. Ten days after the transfer, the plants were at the third leaf stage. Treatments applied were designed as follows: (1) control (only nutrient solution), (2) NaCl 100 mM, (3) SA 0.01 mM, (4) NaCl 100 mM plus 0.01 mM SA. pH of the nutrient solution was adjusted at 6 with KOH (1.0 N). After 17 days of treatments, plants were harvested and divided into roots, stems, and leaves for physiological and biochemical parameters determination.
Measured parameters
Plant growth measurement
Dry weight was determined after desiccation at 80°C for 48 h.
Ion content
Determination of K+, Na+, and Cl−: extraction of ions was achieved in HNO3 (0.1 N). K+and Na+ were determined by flame spectrophotometry and Cl− by coulometry.
Chlorophyll content
Total chlorophyll (Chlt) concentrations were determined following the method of Torrecillas et al. (1984). Total chlorophyll was calculated according to the following expression:
Chlorophyll fluorescence
Chlorophyll fluorescence emission from the adaxial surface of the third and fifth leaf from the shoot tip was measured by a fluorimeter type P.E.A (Plant Efficiency Analyzer from Hansatech, Norfolk, UK).
The unit consists of a fluorescence sensor that allows the illumination and detection of fluorescence of a leaf area of 50 mm2. The sensor is connected to a control unit for processing and storing signals from the sensor. To visualize the induction curves of fluorescence measuring, the unit is connected to a PC microcomputer. Data are processed using software provided by Hansatech. At each measurement, the leaf is fixed by the intact middle portion of a clamp ensuring total darkness on an area of 50 mm2. After an adaptation time of 30 min in the dark, the area bounded by the clamp is exposed to red light (650 nm), with an intensity of 3,000 μmol·m−2·s−2, prior to fluorescence measurements.
Proline
The proline content was determined using the method of Bates et al. (1973). Proline was extracted from organs samples of 200 mg FW with 10 mL of 3% sulphosalycylic acid at 70°C for 30 min. Extracts were filtered through filter paper. After addition of acid ninhydrin and glacial acetic acid to the extracts, the mixture was heated at 90°C for 1 h in water bath. Reaction was then stopped by using ice bath. The mixture was extracted with toluene, and the absorbance of fraction with toluene evaporated from liquid phase was spectrophotometrically determined at 520 nm. Proline concentration was determined using calibration curve as μmol proline·g−1 FW.
Soluble sugars
Total soluble sugars were estimated by the anthrone reagent method using glucose as the standard (Yemm and Willis, 1954).
Protein contents
Leaf and root were ground in mortar with liquid nitrogen and the soluble proteins were extracted by adding 50 mM pH 7.5 potassium phosphate buffer, 1 mM EDTA, 1 mM DTT, 5% glycerol, and 5% polyvinylpyrrolidone. The mixture was centrifuged at 13,000×g for 10 min at 4°C. Protein concentration was determined according to the method of Bradford (1976), using bovine serum albumin as a standard.
Statistical analysis
The data was analyzed using analysis of variance (ANOVA) and the significance of the differences between means were determined at p<0.05 using Duncun's multiple range test (Statistica TM software).
Results
Growth parameters
The dry weight of tomato plants was reduced significantly with salt stress. Shoot dry weight was reduced by approximately 49% compared to control; in details, this reduction was about 59 and 43%, respectively, in stems and in leaves. Roots were less sensitive to NaCl; the reduction was only 25%. Application of 0.01 mM SA through the rooting medium improved plant growth of both stressed and nonstressed tomato plants. SA (0.01 mM) added to control plants enhanced shoot growth by 17% compared with control plants, whereas in saline conditions SA ameliorate shoot growth with 39% compared to plants growing only in saline medium (Fig. 1). Salinity (NaCl) caused a marked reduction in leaf area. The reduction was about 63% at stage 3 and 65% at stage 5 compared with control plants. The addition of SA in culture medium affected positively leaf area compared to control under salt stress (Fig. 2).

Dry weight of leaves, stems, and roots of tomato plants grown for 17 days in nutrient solution with 100 mM NaCl±0.01 mM salicyclic acid. Means of 12 replicates. Bars indicate±standard error (p=0.05).
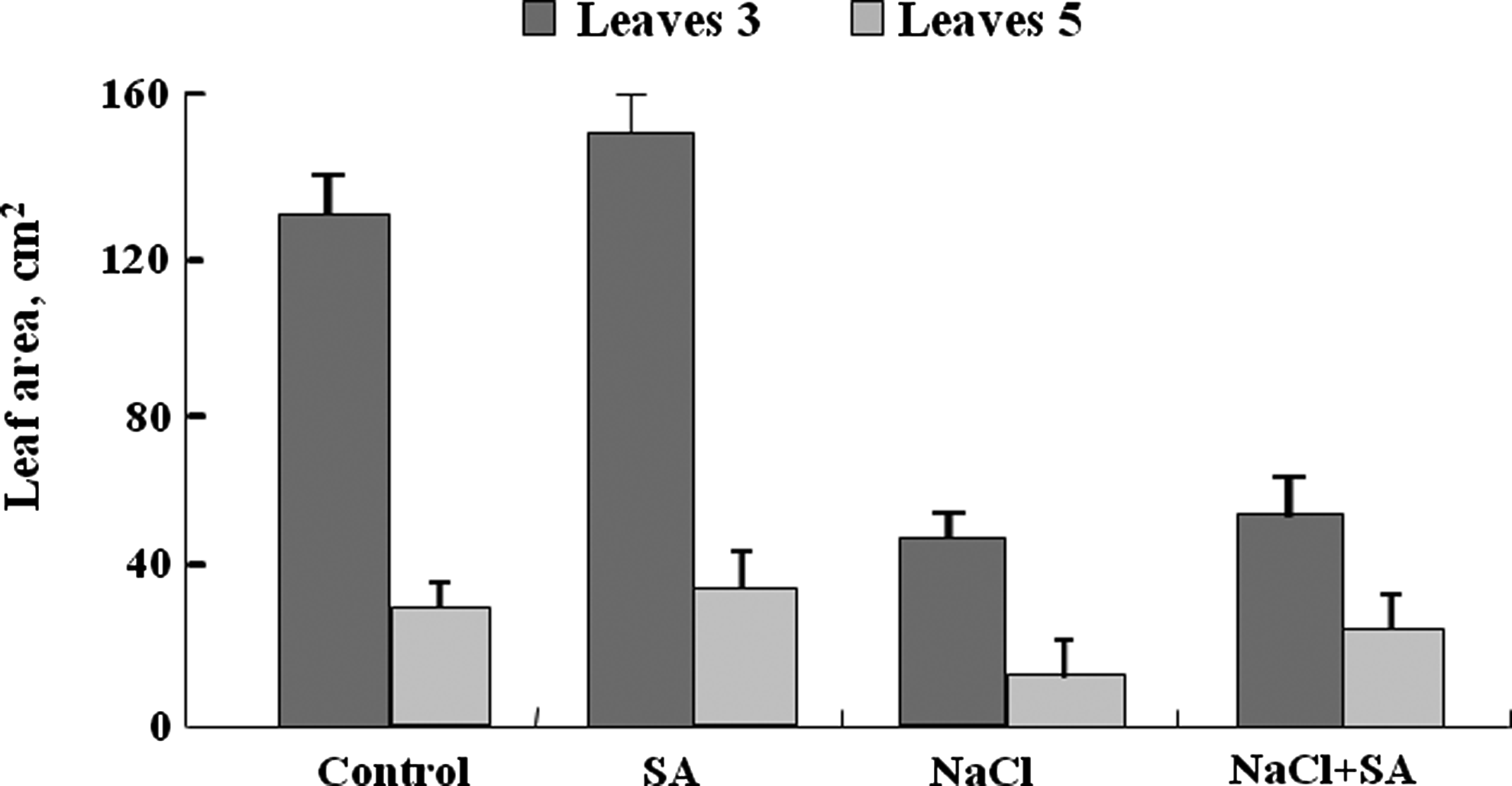
Leaf area of stages 3 and 5 of tomato plants grown for 17 days in nutrient solution in 100 mM NaCl±0.01 mM salicylic acid. Means of six replicates. Bar indicates±standard error (p=005).
Ion content
External NaCl salinity decreased the mineral content in shoot and root of tomato plants. The amounts of K+ absorbed by roots and transported to the shoots were restricted (Fig. 3). Content of this ion decreased in saline conditions, which suggests a deficit in providing the plants with essential ions to the growth. Salinity reduced K+ concentration in leaves and stems respectively, with 45 and 33% in tomato plants. The addition of salicylic acid in the culture medium improves accumulation of K+ on the stems compared to plant treated by NaCl only. Moreover, in the presence of NaCl, sodium and chloride were accumulated in leaves, stems, and roots of all plants grown under saline conditions. Stems are the organs most charged with Na+ and Cl−(Fig. 4). However, application of SA induced a significantly decreases in Na+ and Cl− content compared to the plants treated only by NaCl 100 mM. Thus, addition of 0.01 mM SA to salted plants reduced Na+ content in shoots and roots by 29% after 17 days. Similarly, SA reduced Cl− content by 22% in the two organs.

K+ contents in tomato plants grown for 17 days in nutrient solution with 100 mM NaCl±0.01 mM salicylic acid. Means of 12 replicates. Bars indicate±standard error (p=0.05).

Na+ and Cl− contents in tomato plants grown for 17 days in nutrient solution with 100 mM NaCl±0.01 mM salicylic acid. Means of 12 replicates. Bars indicate±standard error (p=0.05).
Chlorophyll contents
Chlorophyll contents of control plants were higher in leaves stage 3 than those in leaves stage 5. NaCl 100 mM, decreased chlorophyll content in leaves almost 50% compared to control plants. Adding 0.01 mM salicylic acid to NaCl-treated plants increased chlorophyll accumulation in the two types of leaves, while adding salicylic to control plants decreased these contents (Fig. 5).
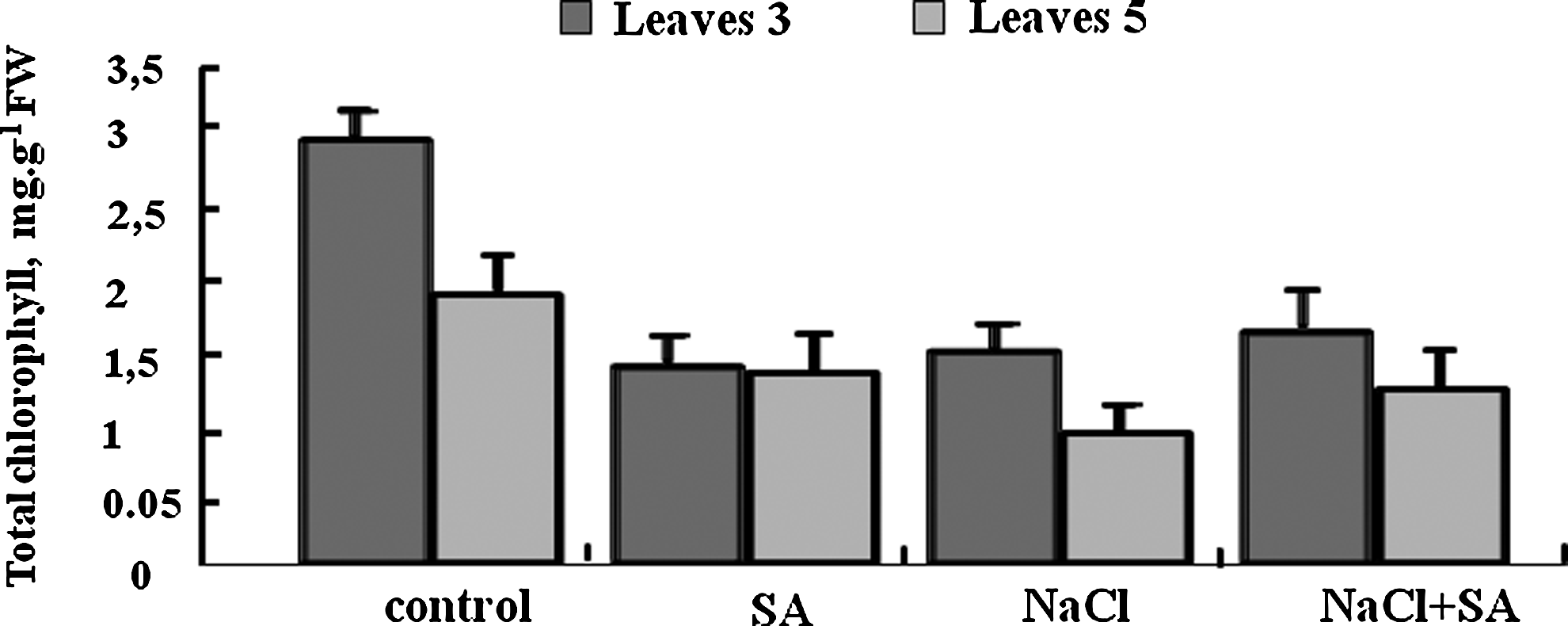
Total chlorophyll contents in leaves at stages 3 and 5 of tomate plants grown for 17 days in nutrient solution with 100 mM NaCl±0.01 mM salicylic acid. Means of six replicates. Bars indicates±standard error (p=0.05).
Chlorophyll fluorescence
Chlorophyll fluorescence was measured on the leaf at stages 3 and 5. Figure 6 shows that the maximum quantum yield of PS2, estimated by the ratio (Fv/Fm), in tomato seedlings subjected to NaCl 100 mM or treated with SA only decreases compared with control plants. In leaves at stage 3, an Fv/Fm change of 0.82 in control medium (−NaCl) to 0.66 in stressful conditions (+NaCl). Adding SA, in the presence or absence of NaCl, did not increase this value in leaf 3. In contrast, in the leaf at stage 5, adding SA to the medium enriched with NaCl 100 mM made this value closer to the values of control plants.

Effect of NaCl (100 mM) and salicylic acid (0.01) on the quantum yield of PS2 estimated by the ratio (Fv/Fm) of the leaves at stages 3 and 5 after 17 days of treatment.
Leaf proline
Figure 7 shows that the content of proline was much higher in leaves of plants subjected to 100 mM NaCl compared with control plants. Leaf 5 accumulated more of amino acid than in at stage 3. But the level of proline decreased with the use of salicylic acid in stressed plants at 100 mM NaCl.
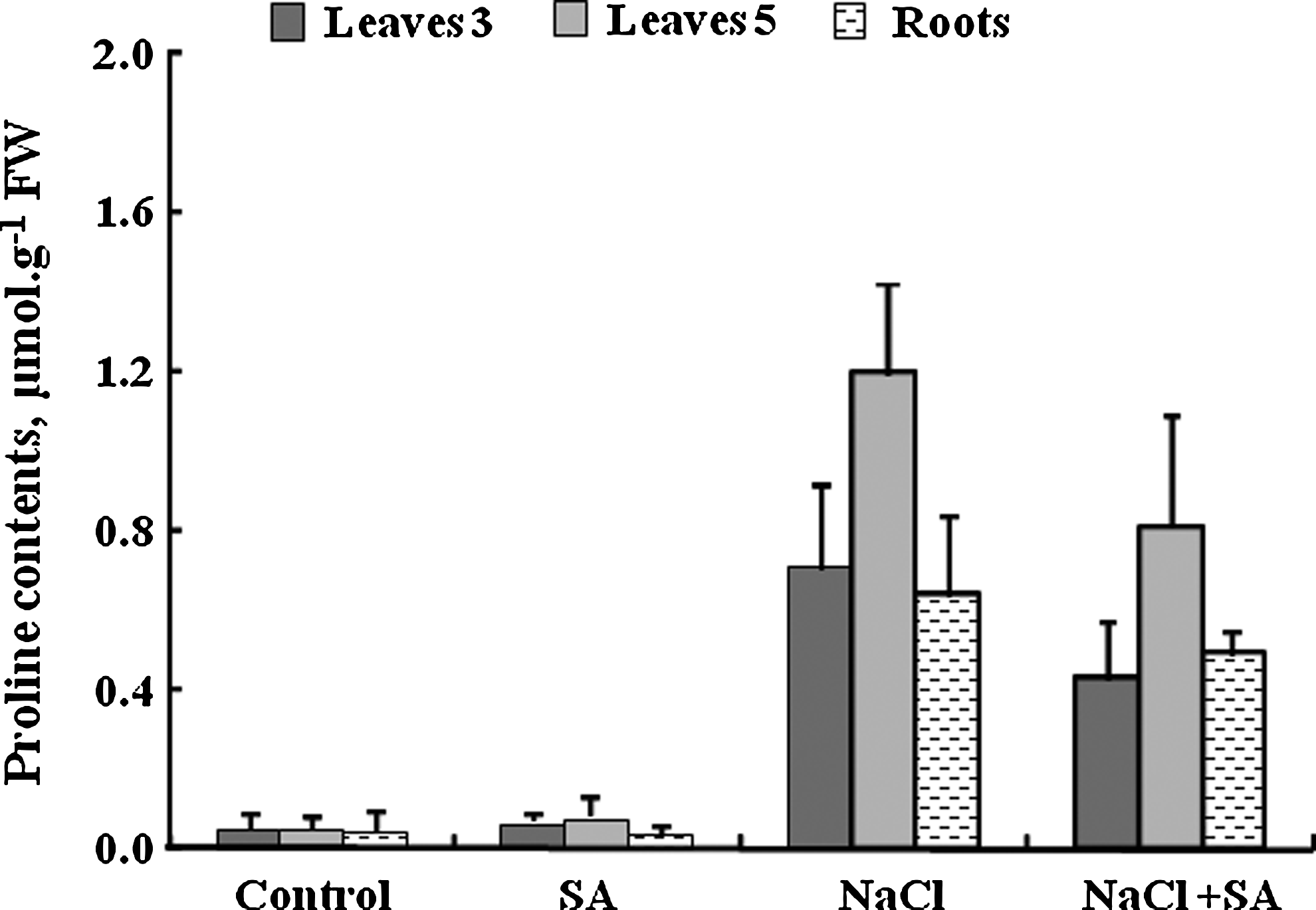
Proline contents in leaves at stages 3 and 5, and roots of tomato plants grown for 17 days in nutrient solution with 100 mM NaCl±0.01 mM salicylic acid. Means of six replicates. Bars indicate±standard error (p=0.05).
Root proline
Figure 7 shows that salinity increased the root proline. But with 0.01 mM SA concentration, the proline decreased.
Leaf sugar
At 100 mM NaCl concentration, the level of sugar in leaf stage 5 increased with 37%, but decreased slightly in leaf stage 3. Salicylic acid increased the sugar level; this increase is about 4, 13% respectively in leaves stages 3 and stage 5 (Fig. 8).

Soluble sugars contents in leaves at stages 3 and 5 and roots of tomato plants grown for 17 days in nutrient solution with 100 mM naCl±0.01 mM salicylic acid. Means of six replicates. Bars indicate±standard error (p=0.05).
Root sugar
According to Figure 8, there was no significant change at 100 mM NaCl; however, adding salicylic acid increased the root sugar level.
Leaf protein
According to Figure 9, salinity decreased the leaf protein level. Salicylic acid decreased more protein concentration at the 100 mM salinity level; also, a large decrease in leaf 5 was observed.
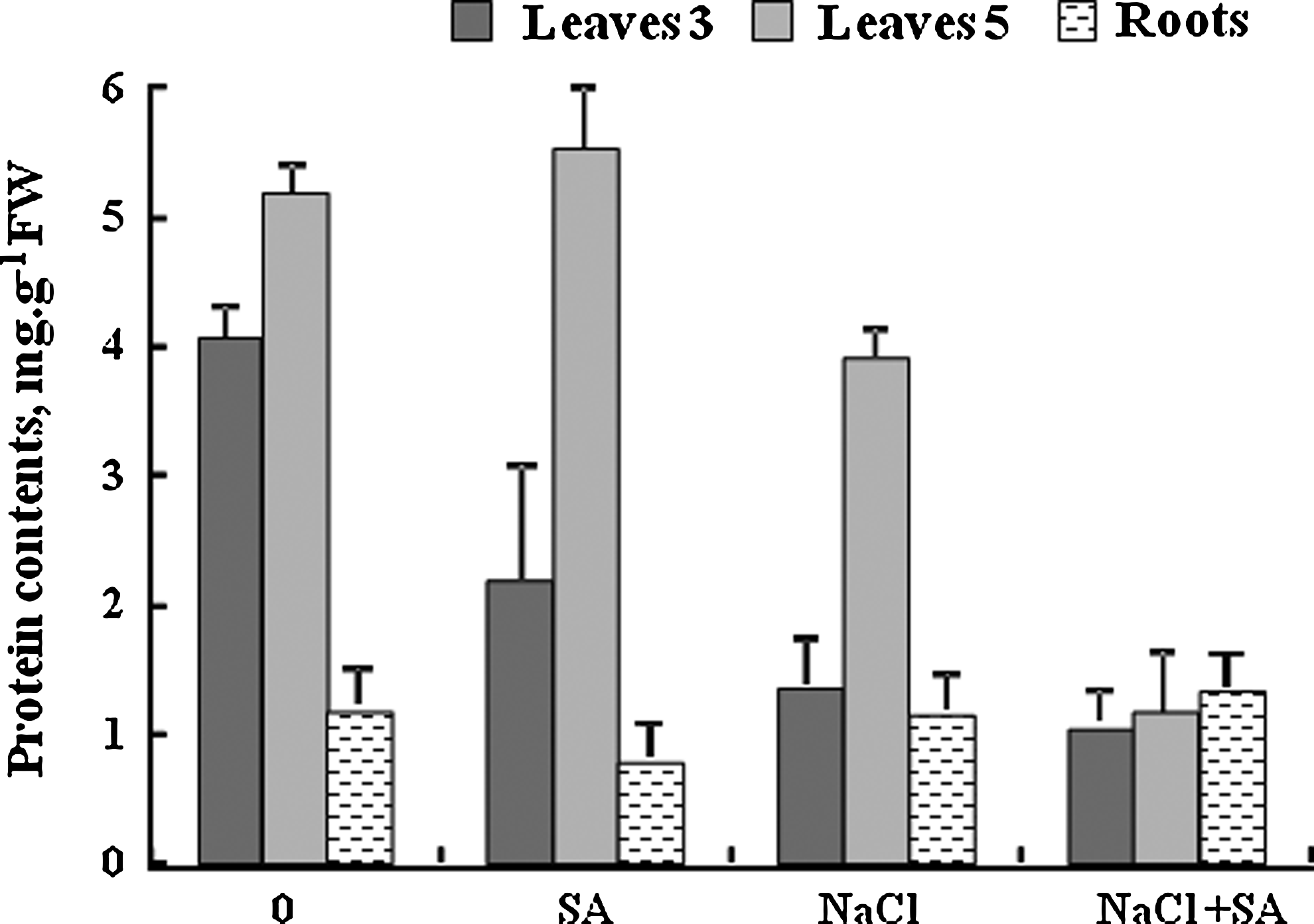
Protein contents in leaves at stages 3 and 5 and roots of tomato plants grown for 17 days in nutrient solution with 100 mM NaCl±0.01 mM salicylic acid. Means of six replicates. Bars indicate±standard error (p=0.05).
Root protein
Figure 9 shows no change in the root protein in the presence of NaCl. Adding salicylic acid in the saline condition increased protein level.
Discussion
Salt (NaCl) stress is among the factors most limiting to plant productivity (Shi et al., 2002). Plants exposed to salt stress adapt their metabolism in order to cope with the changed environment. Survival under these stressful conditions depends on the plant's ability to perceive the stimulus, generate and transmit signals, and instigate biochemical changes that adjust the metabolism accordingly (Hussain et al., 2008).
In our study we found that application of NaCl 100 mM to tomato plants adversely influenced their growth pattern (dry weights of shoots and roots and leaf area), compared with control plants (Fig. 1). These results are in agreement with those of Ghoulam et al. (2001), who showed that NaCl salinity caused a marked reduction in growth of sugar beet plants. The reduction in growth observed for many plant species subjected to excess salinity is often associated with a reduction in photosynthetic capacity. Salicylic acid-treated tomato plants exhibited an increase in tolerance to salt treatment. This increase was reflected in the measured growth criteria: dry weight of shoots and roots as well as leaf area was increased compared with plants receiving NaCl only (Fig. 1). Khodary (2004) also reported a similar increase in the growth of shoots and roots of Maize plants in response to salicylic acid treatment. Also, Kaydan et al. (2007) noted that SA promotes the growth of young wheat plants grown on saline medium. Increase in growth of wheat under nonsaline or saline conditions from which SA treatment resulted, can be attributed to an increase in photosynthesizing tissue, that is, the leaves (Dhaliwal et al., 1997).
According to the data provided in the litterature, macro element (K, Ca) uptake and transportation are usually reduced in plants under NaCl stress
Figure 5 shows that total chlorophyll content of NaCl-treated tomato plants was decreased below that of the controls in both leaf 3 and 5. Similarly, Turan et al. (2007) found an inhibition in chlorophyll biosynthesis lentil plants because of salt stress. The exogenous SA application enhanced pigment contents in saline conditions (Fig. 5). Also, Khodary (2004) observed a significant increase in pigment contents in maize sprayed with SA. In contrast, SA in control plants, decrease total chlorophyll contents; these results were in accordance with Pancheva et al. (1996). By chlorophyll fluorescence measurements, it can be concluded that the maximum quantum yield of PS2 as measured by Fv/Fm was decreased from that of the control in the treated leaves stages 3 and 5 with NaCl 100 mM and those treated only with SA. After 17 days of treatment by NaCl, Fv/Fm of PS2 in the leaves stage 5 treated with SA were enhanced from that of the stressed one treated with only NaCl 100 mM. In contrast, in leaves stage 3, SA does not increase Fv/Fm. Similar results were observed by Szepesi (2006) and Tari et al. (2002).
Sugars are considered to play a major role in osmoregulation under abiotic stress conditions (Fallon and Phillips, 1988). In fact, the first effect of salinity on plants is a drought effect or water deficit. Plants try to reduce their osmotic potential via increasing mineral ions content and compatible solutes synthesis to better water uptake under salinity. Total soluble carbohydrates are important solutes that are synthesized and accumulated in cytosol under salt stress.Thus, they are necessary for the survival of the plant, because they are a source of carbon and energy (Rejskova et al., 2007). In our study, tomato plants submitted to NaCl salinity treatment showed an increase in their soluble sugar content in leaf 5 and a decrease in leaf 3 but no change in root sugar level (Fig. 6). Kerepsi and Galiba (2000) reported that carbohydrates changes are important because of their relationship with such physiological processes as photosynthesis, translocation, and respiration. Some studies reported that plants sugars content rose (Munns and Weir, 1981) or remained constant (Morgan, 1992) under salinity. The increase in soluble sugars due to salt stress has also be reported in wheat by Kumar et al. (2002). The soluble sugar in oat organ plant (root and bud) increased with NaCl increasing (El-Tayeb, 2005). In contrast, Patakas et al. (2002) observed a nonsignificant difference in sugars contents, for example, grapevine plants under stress conditions. The salt stress in soybean varieties decreased sugar level
In contrast, Khodary (2004) reported a decrease in soluble sugar content associated with an increase in polysaccharides in NaCl-stressed maize plants with SA application. It is indicated that SA application may activate the metabolic consumption of soluble sugars to form new cell constituents as a mechanism to stimulate the growth of barely plants. It is also assumed that SA treatments may inhibit polysaccharide-hydrolyzing enzymes and/or accelerate the incorporation of soluble sugars into polysaccharides. This assumption could be supported by the observation that SA increased insoluble sugars level at the expense of soluble sugars (El-Tayeb, 2005).
Amino acid proline, which is known to occur widely in higher plants, normally accumulates in large amounts in response to various abiotic stresses (Ali et al., 1999; Ashraf, 1994). In addition to its role as an osmolyte for osmotic adjustment, proline contributes to stabilizing subcellular structures, for example, membrane and proteins, scavenging free radicals and buffering cellular redox potential under stress conditions (Hsu et al., 2003; Ozturk and Demir, 2002). It has long been suggested that accumulation of proline in plant tissue under salt stress is an adaptative response even though investigators have obtained contrasting results regarding the role of proline in stress tolerance of plants (Ashraf, 1994; Rhodes et al., 1999). In the tomato, several studies have demonstrated the accumulation of proline in roots and leaves under salt stress (Amini and Ehsanpour, 2005; Szepesi, 2006).
The data showed that the content of proline increased in leaves (3, 5) and roots of the tomato plants at 100 mM NaCl (Fig. 7). Also, Sakhabutdinova et al. (2003) reported that salinity and water deficit induces accumulation of proline in wheat plants. In control plants, SA treatment induced an increase in the contents of this compound in leaves. In this regard, Sakhabutdinova et al. (2003) found that SA increased proline accumulation in normal wheat seedlings. But under salt stress, SA decreases proline level. These results are in accordance with Shahba et al. (2010), who found that SA decreased proline level in tomato under salinity stress.
Soluble protein is generally decreased in response to salinity (Abed-Latef, 2005; Parida et al., 2002). It has also been reported that high salt concentration causes an increase in high protein content in some glycophytic plants (Abed El-Baki, 1996; Jones and MacMillan, 1987) or increase in soluble proteins (Shaddad et al., 2005). Amini and Ehsanpour (2005) reported that the accumulation of proteins in plants grown under saline condition may provide a storage form of nitrogen that is reutilized when stress is over and may play a role in osmotic adjustment. Increasing of soluble proteins may be due to synthesis of osmotin like protein or structural protein, in particular, synthesis of those proteins that are involved in modification of a cell wall.
According to this research, the leaf protein level decreased by salt stress. Shahba et al. (2010) reported that the leaf protein level decreased in tomato because of a salinity effect. The cause of protein reduction at the salinity condition is the inhibition of nitrate reductase activity (Undovenko, 1971). Salt stress induced some changes on the protein of rice leaf shoots and roots, but was not effective on the leaf blade. The level of some protein decreases because of protein synthesis reduction (Kong-Ngern et al., 2005). In control plants, SA treatment induced an increase in the content of proteins only in leaf 5. In NaCl-treated plants, proteins content decreased in both leaves 3 and 5 when SA was applied than those under the corresponding stress; the reduction was very pronounced in leaf 5. Singh and Usha (2003) have observed a similar reduction in soluble proteins content in water-stressed plant with (1 or 2 mM) SA application, unlike roots, which marked a slight increase after adding salicylic acid.
Conclusions
Taken overall, it might be assumed that SA, given through root medium in a lower dose (0.01), positively affected the growth parameters when compared to the control plants. The enhancement of salt tolerance in tomato cv Moneymaker was positively associated with the decreasing transport and the preventing of the accumulation of toxic ions as sodium and chloride at toxic levels. Salt stress resistance was also probably in relation with modifications in some metabolites, for example, soluble sugar, which played a role of compatibles osmolytes to compensate the decrease of water potential in the presence of Na+ and Cl−.
Footnotes
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.